

Escherichia coli Isolated from Vegans, Vegetarians and Omnivores: Antibiotic Resistance, Virulence Factors, Pathogenicity Islands and Phylogenetic Classification
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Antimicrobial Susceptibility
2.3. Detection of β-Lactamases
2.4. Detection of Virulence-Associated Genes
2.5. Detection of Pathogenicity Island (PAI) Markers
2.6. Phylogenetic Classification
2.7. Statistical Analysis
3. Results
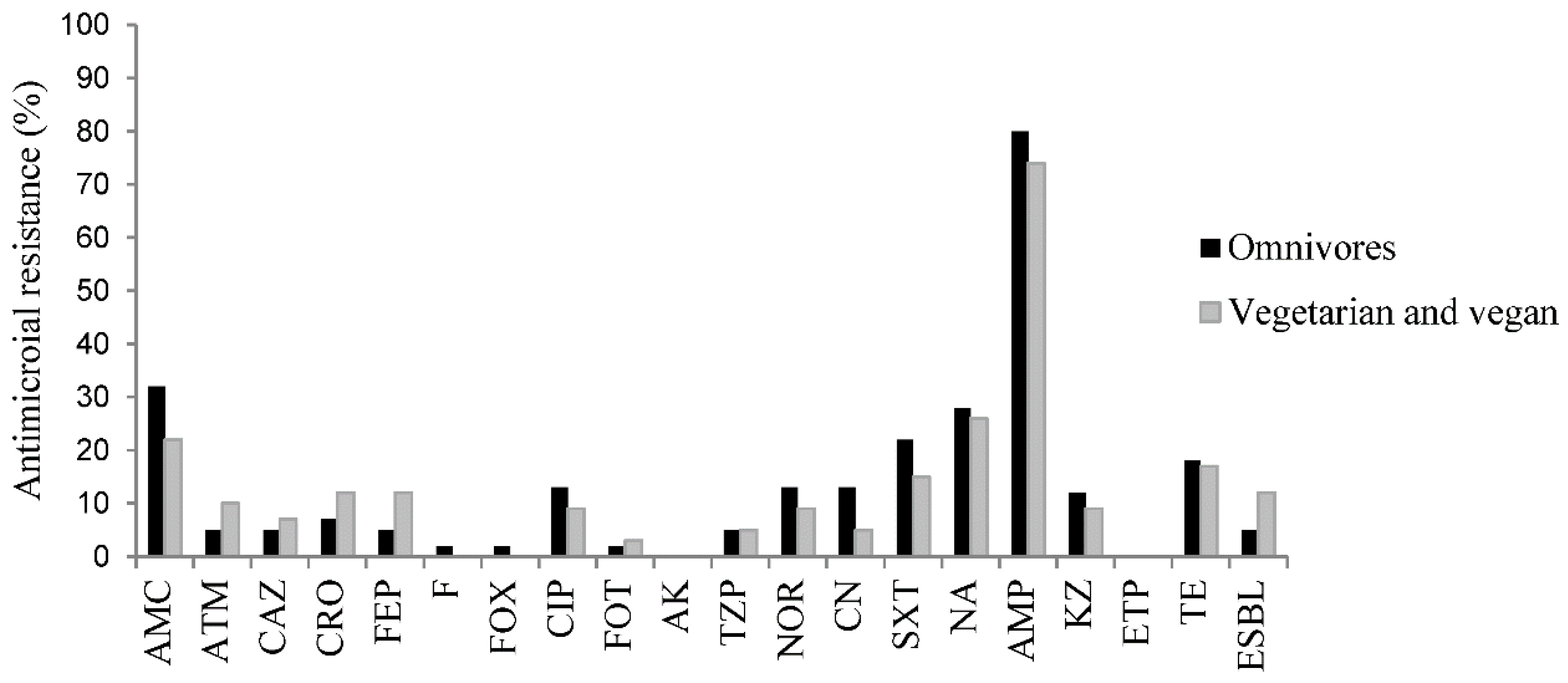
3.1. Antibiotic Resistance
3.2. Virulence-Associated Genes
3.3. PAIs
3.4. Phylogeny
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Micenková, L.; Beňová, A.; Frankovičová, L.; Bosák, J.; Vrba, M.; Ševčíková, A.; Kmeťová, M.; Šmajs, D. Human Escherichia coli isolates from hemocultures: Septicemia linked to urogenital tract infections is caused by isolates harboring more virulence genes than bacteremia linked to other conditions. Int. J. Med. Microbiol. 2017, 307, 182–189. [Google Scholar] [CrossRef]
- Pitout, J.D.D. Extraintestinal pathogenic Escherichia coli: A combination of virulence with antibiotic resistance. Front. Microbiol. 2012, 3, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köhler, C.-D.; Dobrindt, U. What defines extraintestinal pathogenic Escherichia coli. Int. J. Med. Microbiol. 2011, 301, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Raeispour, M.; Ranjbar, R. Antibiotic resistance, virulence factors and genotyping of Uropathogenic Escherichia coli strains. Antimicrob. Resist. Infect. Control 2018, 7, 118. [Google Scholar] [CrossRef] [PubMed]
- Daga, A.P.; Koga, V.L.; Soncini, J.G.M.; De Matos, C.M.; Perugini, M.R.E.; Pelisson, M.; Kobayashi, R.; Vespero, E.C. Escherichia coli Bloodstream Infections in Patients at a University Hospital: Virulence Factors and Clinical Characteristics. Front. Cell Infect. Microbiol. 2019, 9, 191. [Google Scholar] [CrossRef]
- Starčič Erjavec, M.; Žgur-Bertok, D. Virulence potential for extraintestinal infections among commensal Escherichia coli isolated from healthy humans—The Trojan horse within our gut. FEMS Microbiol. Lett. 2015, 362, fnu061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fegan, N.; Gobius, K.S. Pathogenic Escherichia coli and One Health Implications. Curr. Top. Microbiol. Immunol. 2013, 366, 49–62. [Google Scholar] [PubMed]
- Schjørring, S.; Krogfelt, K.A. Assessment of bacterial antibiotic resistance transfer in the gut. Int. J. Microbiol. 2011, 2011, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Soncini, J.G.M.; Cerdeira, L.; Sano, E.; Koga, V.L.; Tizura, A.T.; Tano, Z.N.; Nakazato, G.; Kobayashi, R.K.T.; Aires, C.A.M.; Lincopan, N.; et al. Genomic insights of high-risk clones of ESBL-producing Escherichia coli isolated from community infections and commercial meat in southern Brazil. Sci. Rep. 2022, 12, 9354. [Google Scholar] [CrossRef]
- Lawson, B.; De Pinna, E.; Horton, R.A.; MacGregor, S.K.; John, S.K.; Chantrey, J.; Duff, J.P.; Kirkwood, J.K.; Simpson, V.R.; Robinson, R.A.; et al. Epidemiological Evidence That Garden Birds Are a Source of Human Salmonellosis in England and Wales. PLoS ONE 2014, 9, e88968. [Google Scholar] [CrossRef]
- Done, H.Y.; Venkatesan, A.K.; Halden, R.U. Does the Recent Growth of Aquaculture Create Antibiotic Resistance Threats Different from those Associated with Land Animal Production in Agriculture? AAPS J. 2015, 17, 513–524. [Google Scholar] [CrossRef] [Green Version]
- Gozi, K.S.; Froes, J.R.; Ajude, L.P.T.D.; Da Silva, C.R.; Baptista, R.S.; Peiró, J.R.; Marinho, M.; Mendes, L.C.N.; Nogueira, M.C.L.; Casella, T. Dissemination of Multidrug-Resistant Commensal Escherichia coli in Feedlot Lambs in Southeastern Brazil. Front. Microbiol. 2019, 10, 1394. [Google Scholar] [CrossRef]
- Mulder, M.; Jong, J.C.K.; Goessens, W.H.F.; de Visser, H.; Ikram, M.A.; Verbon, A.; Stricker, B.H. Diet as a risk factor for antimicrobial resistance in community-acquired urinary tract infections in a middle-aged and elderly population: A case–control study. Clin. Microbiol. Infect. 2019, 25, 613–619. [Google Scholar] [CrossRef]
- De Filippo, C.; Cavalieri, D.; di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [Green Version]
- Marano, R.B.; Fernandes, T.; Manaia, C.M.; Nunes, O.; Morrison, D.; Berendonk, T.U.; Kreuzinger, N.; Tenson, T.; Corno, G.; Fatta-Kassinos, D.; et al. A global multinational survey of cefotaxime-resistant coliforms in urban wastewater treatment plants. Environ. Int. 2020, 144, 106035. [Google Scholar] [CrossRef]
- CLSI. Suggested Grouping of US-FDA Approved Antimicrobial Agents That Should Be Considered for Routine Testing and Reporting on Nonfastidious Organisms by Clinical Laboratories, 29th ed.; CLSI Guideline M100-S29; Clinical and Laboratory Institute: Wayne, PA, USA, 2019. [Google Scholar]
- Dallenne, C.; Da Costa, A.; Decré, D.; Favier, C.; Arlet, G. Development of a set of multiplex PCR assays for the detection of genes encoding important beta-lactamases in Enterobacteriaceae. J. Antimicrob. Chemother. 2010, 65, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Leflon-Guibout, V.; Jurand, C.; Bonacorsi, S.; Espinasse, F.; Guelfi, M.C.; Duportail, F.; Heym, B.; Bingen, E.; Nicolas-Chanoine, M.H. Emergence and spread, of three clonally related virulent isolates of CTX-M-15-producing Escherichia coli with variable resistance to aminoglycosides and tetracycline in a French geriatric hospital. Antimicrob. Agents Chemother. 2004, 48, 3736–3742. [Google Scholar] [CrossRef] [Green Version]
- Koga, V.L.; Scandorieiro, S.; Vespero, E.C.; Oba, A.; de Brito, B.G.; de Brito, K.C.T.; Nakazato, G.; Kobayashi, R.K.T. Comparison of antibiotic resistance and virulence factors among Escherichia coli isolated from conventional and free-range poultry. BioMed. Res. Int. 2015, 2015, 618752. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Stell, A.L. Extended virulence genotypes of Escherichia coli strains from patients with urosepsis in relation to phylogeny and host compromise. J. Infect. Dis. 2000, 181, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Sabaté, M.; Moreno, E.; Perez-Berezo, T.; Andreu, A.; Prats, G. Pathogenicity island markers in commensal and Uropathogenic Escherichia coli isolates. Clin. Microbiol. Infect. 2006, 12, 880–886. [Google Scholar] [CrossRef]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef]
- Von Baum, H.; Marre, R. Antimicrobial resistance of Escherichia coli and therapeutic implications. Int. J. Med. Microbiol. 2005, 295, 503–511. [Google Scholar] [CrossRef]
- Samei, A.; Haghi, F.; Zeighami, H. Distribution of pathogenicity island markers in commensal and uropathogenic Escherichia coli isolates. Folia. Microbiol. 2016, 61, 261–268. [Google Scholar] [CrossRef]
- Shurina, B.A.; Page, R.C. Structural Comparisons of Cefotaximase (CTX-M-ase) Sub Family 1. Front. Microbiol. 2021, 12, 688509. [Google Scholar] [CrossRef]
- Poirel, L.; Gniadkowski, M.; Nordmann, P. Biochemical analysis of the ceftazidime-hydrolysing extended-spectrum β-lactamase CTX-M-15 and of its structurally related β-lactamase CTX-M-3. J. Antimicrob. Chemother. 2002, 50, 1031–1034. [Google Scholar] [CrossRef] [Green Version]
- Naas, T.; Oueslati, S.; Bonnin, R.A.; Dabos, M.L.; Zavala, A.; Dortet, L.; Retailleau, P.; Iorga, B.I. Beta-lactamase database (BLDB) – structure and function. J. Enzym. Inhib. Med. Chem. 2017, 32, 917–919. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Zhang, H.; Guo, Y.; Tian, T. Influence of chicken manure fertilization on antibiotic-resistant bacteria in soil and the endophytic bacteria of pakchoi. Int. J. Environ. Res. Public Health 2016, 13, 662. [Google Scholar] [CrossRef]
- Yang, Y.; Ashworth, A.; Willett, C.; Cook, K.; Upadhyay, A.; Owens, P.R.; Ricke, S.C.; DeBruyn, J.; Moore, P.A., Jr. Review of Antibiotic Resistance, Ecology, Dissemination, and Mitigation in U. S. Broiler Poult. Syst. 2019, 10, 2639. [Google Scholar]
- Sneeringer, S.; MacDonald, J.M.; Key, N.; McBride, W.D.; Mathews, K. Economics of Antibiotic Use in U.S. livestock production. USDA Econ. Res. Rep. 2015. Available online: https://www.ers.usda.gov/webdocs/publications/err200/55529_err200.pdf (accessed on 10 October 2022).
- Aslam, N.; Khan, S.-U.; Usman, T.; Ali, T. Phylogenetic genotyping, virulence genes and antimicrobial susceptibility of Escherichia coli isolates from cases of bovine mastitis. J. Dairy Res. 2021, 88, 78–79. [Google Scholar] [CrossRef]
- Sarowska, J.; Futoma-Koloch, B.; Jama-Kmiecik, A.; Frej-Madrzak, M.; Ksiazczyk, M.; Bugla-Ploskonska, G.; Choroszy-Krol, I. Virulence factors, prevalence and potential transmission of extraintestinal pathogenic Escherichia coli isolated from different sources: Recent reports. Gut Pathog. 2019, 11, 10. [Google Scholar] [CrossRef] [Green Version]
- Nowrouzian, F.; Hesselmar, B.; Saalman, R.; Strannegård, I.; Åberg, N.; Wold, A.E.; Adlerberth, I. Escherichia coli in infants’ intestinal microflora: Colonization rate, strain turnover, and virulence gene carriage. Pediatr. Res. 2003, 54, 8–14. [Google Scholar] [CrossRef]
- Kline, K.A.; Fälker, S.; Dahlberg, S.; Normark, S.; Henriques-Normark, B. Bacterial Adhesins in Host-Microbe Interactions. Cell Host Microbe 2009, 5, 580–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mjailovic, H.; Smith, S.G. Bacterial self-defence: How Escherichia coli evades serum killing. FEMS Microbiol. Lett. 2014, 354, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, C. Structure and Assembly of Escherichia coli Capsules. EcoSal Plus 2009, 354, 1–9. [Google Scholar] [CrossRef]
- Knust, Z.; Schmidt, G. Cytotoxic Necrotizing Factors (CNFs)−A Growing Toxin Family. Toxins 2010, 2, 116–127. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-C.; Chang, C.-C.; Chiu, T.H.T.; Lin, M.-N.; Lin, C.-L. The risk of urinary tract infection in vegetarians and non-vegetarians: A prospective study. Sci. Rep. 2020, 10, 906. [Google Scholar] [CrossRef]


{kind=link}
| Enzymes | Genes | Primer Sequences (5′–3′) | Amplion Size (bp) | References |
|---|---|---|---|---|
| CTX-M-1 | blaCTX-M1 | TTAGGAARTGTGCCGCTGYA CGATATCGTTGGTGGTRCCAT | 688 | Dallene et al., 2010 [17] |
| CTX-M-2 | blaCTX-M2 | CGTTAACGGCACGATGAC CGATATCGTTGGTGGTRCCAT | 404 | Dallene et al., 2010 [17] |
| CTX-M-8/25 | blaCTX-M8 | AACRCRCAGACGCTCTAC TCGAGCCGGAASGTGTYAT | 326 | Dallene et al., 2010 [17] |
| CTX-M-9 | blaCTX-M9 | TCAAGCCTGCCGATCTGGT TGATTCTCGCCGCTGAAG | 561 | Dallene et al., 2010 [17] |
| CTX-M-15 | blaCTX-M15 | ATA AAA CCG GCA GCG GTGGAA TTT TGA CGA TCG GGG | 483 | Leflon-Guibout et al., 2004 [18] |
| Genes | Primer Sequences (5′–3′) | VAGs | Amplion Size (bp) | References |
|---|---|---|---|---|
| kpsMTII | GCG CAT TTG CTG ATA CTG TTG | Group 2 of capsular antigens | 272 | Johnson and Stell, 2000 [20] |
| CAT CCA GAC GAT AAG CAT GAC CA | ||||
| KpsMTIII | TCC TCTT GCT ACT ATT CCC CCT | Group 3 of capsular antigens | 392 | Johnson and Stell, 2000 [20] |
| AGG CGT ATC CAT CCC TCC TAA C | ||||
| KpsMTk1 | TAG CAA ACG TTC TAT TGG TGC | K1 capsule | 153 | Johnson and Stell, 2000 [20] |
| CAT CCA GAC GAT AAG CAT GAC CA | ||||
| kpsMTk5 | CAG TAT CAG CAA TCG TTC TGT A | K5 capsule | 159 | Johnson and Stell, 2000 [20] |
| CAT CCA GAC GAT AAG CAT GAC CA | ||||
| cvaC | CAC ACA CAA ACG GGA GCT GTT | Colicin V | 680 | Johnson and Stell, 2000 [20] |
| CTT CCC GCA GCA TAG TTC CAT | ||||
| iutA | GGC TGG ACA TCA TGG GAA CTG G | Aerobactin siderophore receptor | 300 | Johnson and Stell, 2000 [20] |
| CGT CGG GAA CGG GTA GAA TCG | ||||
| fimH | TGC AGA ACG GAT AAG CCG TGG | Fimbriae type 1 | 508 | Johnson and Stell, 2000 [20] |
| GCA GTC ACC TGC CC TCC GGT A | ||||
| fyuA | TGA TTA ACC CCG CGA CGG AA | Yersiniobactinsiderophore receptor | 880 | Johnson and Stell, 2000 [20] |
| CGC AGT AGG CAC GAT CTT GTA | ||||
| papC | GAC GGC TGT ACT GCA GGG TGT GGC G | P Fimbriae | 328 | Johnson and Stell, 2000 [20] |
| ATA TCC TTT CTG CAG GCA GGG TGT GGC | ||||
| papG | CTG TAA TTA CGG AAG TGA TTT CTG | P Fimbriae | 1070 | Johnson and Stell, 2000 [20] |
| CTG TAA TTA CGG AAG TGA TTT CTG | ||||
| sfaA | CTC CGG AGA ACT GGG TGC ATC TTA C | Sfa fimbriae | 410 | Johnson and Stell, 2000 [20] |
| CGG AGG AGT AAT TAC AAA CCT GGC A | ||||
| sfaS | GTG GAT ACG ACG ATT ACT GTG | Sfa fimbriae | 240 | Johnson and Stell, 2000 [20] |
| CCG CCA GCA TTC CCT GTA TTC | ||||
| afa | GGC AGA GGG CCG GCA ACA GGC | Afa fimbriae | 750 | Johnson and Stell, 2000 [20] |
| CCC GTA ACG CGA CAG CAT CTC | ||||
| ibeA | AGG CAG GTG TGC GCC GCG TAC | Invasion of brain endothelium | 170 | Johnson and Stell, 2000 [20] |
| TGG TGC TCC GGC AAA CCA TGC | ||||
| Hly | AAC AAG GAT AAG CAC TGT TCT GGC | Hemolysin | 1177 | Johnson and Stell, 2000 [20] |
| ACC ATA TAA GCG GTC ATT CCC GTC | ||||
| cnf1 | AGG AAG TTA TAT TTC CGT AGG | Cytotoxic necrotizing factor 1 | 498 | Johnson and Stell, 2000 [20] |
| GTA TTT GCC TGA ACC GTA A | ||||
| cnf2 | AAT CTA ATT AAA GAG AAC | Cytotoxic necrotizing factor 2 | 543 | Johnson and Stell, 2000 [20] |
| CAT GCT TTG TAT ATC TA | ||||
| traT | GGT GTG GTG CGA TGA GCA CAG | Serum resistance | 290 | Johnson and Stell, 2000 [20] |
| GGT GTG GTG CGA TGA GAC CAG | ||||
| iroN | AAT CCG GCA AAG AGA CGA ACC GCC T | Salmochelinsiderophore receptor | 553 | Johnson and Stell, 2000 [20] |
| GTT CGG GCA ACC CCT GCT TTG ACT TT | ||||
| ompT | TCA TCC CGG AAG CCT CCC TCA CTA CTA T | Episomal outer membrane protease | 496 | Johnson and Stell, 2000 [20] |
| TAG CGT TTG CTG CAC TGG CTT CTG ATA C | ||||
| hlyF | GGC CAC AGT CGT TTA GGG TGC TTA CC | Putative avian hemolysin | 450 | Johnson and Stell, 2000 [20] |
| GGC GGT TTA GGC ATT CCG ATA CTC AG | ||||
| Iss | CAG CAA CCC GAA CCA CTT GAT G | Episomal increased serum survival | 323 | Johnson and Stell, 2000 [20] |
| AGC ATT GCC AGA GCG GCA GAA |
| PAI | Primers | Sequences (5′–3′) | Amplicon Size (bp) | References |
|---|---|---|---|---|
| PAI I536 | I.9 | TAA TGC CGG AGA TTC ATT GTC | 1800 | Koga et al., 2014 [19] |
| I.10 | AGG ATT TGT CTC AGG GCT TT | |||
| PAI II536 | orf1up | CAT GTC CAA AGC TCG AGC C | 1000 | Sabaté et al., 2006 [21] |
| orf1down | CTA CGT CAG GCT GGC TTT G | |||
| PAI III536 | sfaAI.1 | CGG GCA TGC ATC AAT TAT CTT TG | 161 | Sabaté et al., 2006 [21] |
| sfaAI.2 | TGT GTA GAT GCA GTC ACT CCG | |||
| PAI IV536 | IRP2 FP | AAG GAT TCG CTG TTA CCG GAC | 300 | Sabaté et al., 2006 [21] |
| IRP2 RP | TCG GGC AGC GTT TCT TCT | |||
| PAI ICFT073 | RPAi | GGA CAT CCT GTT ACA GCG CGC A | 930 | Sabaté et al., 2006 [21] |
| RPAf | TCG CCA ATC ACA GC GAA C | |||
| PAI IICFT073 | cft073.2Ent1 | ATG GAT GTT GTA TCG CGC | 400 | Sabaté et al., 2006 [21] |
| cft073.2Ent2 | ACG AGC ATG TGG ATC TGC | |||
| PAI IJ96 | papGIf | TCG TGC TCA GGT CCG GAA TTT | 400 | Sabaté et al., 2006 [21] |
| papGIr | TGG CAT CCC ACA TTA TCG | |||
| PAI IIJ96 | hlyd | GGA TCC ATG AAA ACA TGG TTA ATG GG | 2300 | Sabaté et al., 2006 [21] |
| cnf | GAT ATT TTT GTT GCC ATT GGT TAC C |
| PCR | Genes | Primer Sequences (5′–3′) | Amplicon Size (bp) | References |
|---|---|---|---|---|
| Multiplex | chuA | GAC GAA CCA ACG GTC AGG AT | 279 | Clermont et al., 2013 [22] |
| TGC CGC CAG TAC CAA AGA CA | ||||
| yjaA | TGA AGT GTC AGG AGA CGC TG | 211 | ||
| ATG GAG AAT GCG TTC CTC AAC | ||||
| TSPE4.C2 | GAG TAA TGT CGG GGC ATT CA | 152 | ||
| CGC GCC AAC AAA GTA TTA CG | ||||
| E group | arpA | GAT TCC ATC TTG TCA AAA TAT GCC | 219 | Clermont et al., 2013 [22] |
| GAA AAG AAA AAG AAT TCC CAA GAG | ||||
| C group | trpA | AGT TTT ATG CCC AGT GCG AG | 489 | Clermont et al., 2013 [22] |
| TCT GCG CCG GTC ACG CCC |
| Genes | Omnivores n (%) | Vegetarians/Vegans n (%) |
|---|---|---|
| CTX-M-1 | - | 4 (57) |
| CTX-M-2 | 1 (33) | 3 (43) |
| CTX-M-8/25 | - | - |
| CTX-M-9 | 2 (67) | 5 (71) |
| CTX-M-15 | - | 2 (29) |
| Total of isolates | 3 | 10 |
| Virulence-Associated Genes | Coding For | Omnivores n = 60 (%) | Vegetarians Vegans n = 58 (%) | p-Value |
|---|---|---|---|---|
| kpsMT II | Group 2 of capsular antigens | 15 (25) | 10 (17) | 0.349 |
| kpsMT III | Group 3 of capsular antigens | 0 | 9 (15) | 0.001 * |
| kpsMT k1 | K1 capsule | 9 (15) | 16 (28) | 0.078 |
| kpsMT k5 | K5 capsule | 14 (23) | 8 (14) | 0.214 |
| cvaC | Colicin V | 3 (5) | 7 (12) | 0.152 |
| iutA | Aerobactinsiderophore receptor | 19 (32) | 13 (22) | 0.308 |
| fimH | Fimbriae type 1 | 42 (70) | 7 (12) | <0.001 * |
| fyuA | Yersiniobactinsiderophore receptor | 19 (32) | 11 (19) | 0.206 |
| papC | P Fimbriae | 7 (12) | 7 (12) | 0.894 |
| papG | P Fimbriae | 2 (3) | 2 (3) | 0.946 |
| sfaA | Sfa fimbriae | 6 (10) | 4 (7) | 0.582 |
| sfaS | Sfa fimbriae | 4 (7) | 2 (3) | 0.451 |
| afa | Afa fimbriae | 2 (3) | 2 (3) | 0.946 |
| ibeA | Invasion of brain endothelium | 5 (8) | 3 (5) | 0.526 |
| hly | Hemolysin | 4 (7) | 8 (14) | 0.180 |
| cnf1 | Cytotoxic necrotizing factor 1 | 0 | 4 (7) | 0.035 * |
| cnf2 | Cytotoxic necrotizing factor 2 | 3 (5) | 4 (7) | 0.631 |
| traT | Serum resistance | 19 (32) | 36 (62) | 0.001 * |
| iroN | Salmochelinsiderophore receptor | 8 (13) | 9 (15) | 0.682 |
| ompT | Episomal outer membrane protease | 4 (7) | 9 (15) | 0.110 |
| hlyF | Putative avian hemolysin | 2 (3) | 6 (10) | 0.118 |
| Iss | Episomal increased serum survival | 12 (20 | 3 (5) | 0.019 * |
| Genes | Omnivores n = 60 (%) | Vegetarians/Vegans n = 58 (%) | p-Value |
|---|---|---|---|
| PAI | |||
| PAI I536 | 33 (55) | 7 (12) | <0.001 * |
| PAI II536 | 0 | 0 | |
| PAI III536 | 7 (12) | 0 | 0.008 * |
| PAI IV536 | 23 (38) | 34 (59) | 0.018 * |
| PAI ICFT073 | 6 (10) | 8 (14) | 0.483 |
| PAI IICFT073 | 11 (18) | 9 (15) | 0.744 |
| PAI IJ96 | 0 | 3 (5) | 0.070 |
| PAI IIJ96 | 3 (5) | 0 | 0.090 |
| Genes | Omnivores n = 60 (%) | Vegetarians/Vegans n = 58 (%) | p-Value |
|---|---|---|---|
| Phylogenetic Classification | |||
| A | 15 (25) | 13 (22) | 0.818 |
| B1 | 8 (13) | 10 (17) | 0.506 |
| B2 | 4 (7) | 11 (19) | 0.079 |
| C | 2 (3) | 2 (3) | 0.703 |
| D | 12 (20) | 8 (14) | 0.414 |
| E | 9 (15) | 10 (17) | 0.506 |
| F | 10 (17) | 4 (7) | 0.073 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tizura, A.T.; Material Soncini, J.G.; Koga, V.L.; Takayama Kobayashi, R.K.; Tano, Z.N.; Vespero, E.C. Escherichia coli Isolated from Vegans, Vegetarians and Omnivores: Antibiotic Resistance, Virulence Factors, Pathogenicity Islands and Phylogenetic Classification. Microbiol. Res. 2022, 13, 825-835. https://doi.org/10.3390/microbiolres13040058
Tizura AT, Material Soncini JG, Koga VL, Takayama Kobayashi RK, Tano ZN, Vespero EC. Escherichia coli Isolated from Vegans, Vegetarians and Omnivores: Antibiotic Resistance, Virulence Factors, Pathogenicity Islands and Phylogenetic Classification. Microbiology Research. 2022; 13(4):825-835. https://doi.org/10.3390/microbiolres13040058
Chicago/Turabian StyleTizura, Ariane Tiemy, João Gabriel Material Soncini, Vanessa Lumi Koga, Renata Katsuko Takayama Kobayashi, Zuleica Naomi Tano, and Eliana Carolina Vespero. 2022. "Escherichia coli Isolated from Vegans, Vegetarians and Omnivores: Antibiotic Resistance, Virulence Factors, Pathogenicity Islands and Phylogenetic Classification" Microbiology Research 13, no. 4: 825-835. https://doi.org/10.3390/microbiolres13040058