
Dysregulation of the mRNA Expression of Human Renal Drug Transporters by Proinflammatory Cytokines in Primary Human Proximal Tubular Epithelial Cells

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Isolation and Culture
2.3. Proinflammatory Cytokine Treatments
2.4. RNA Isolation, cDNA Synthesis and Quantification via Real-Time Quantitative PCR (qPCR)
2.5. Data and Statistical Analysis
3. Results
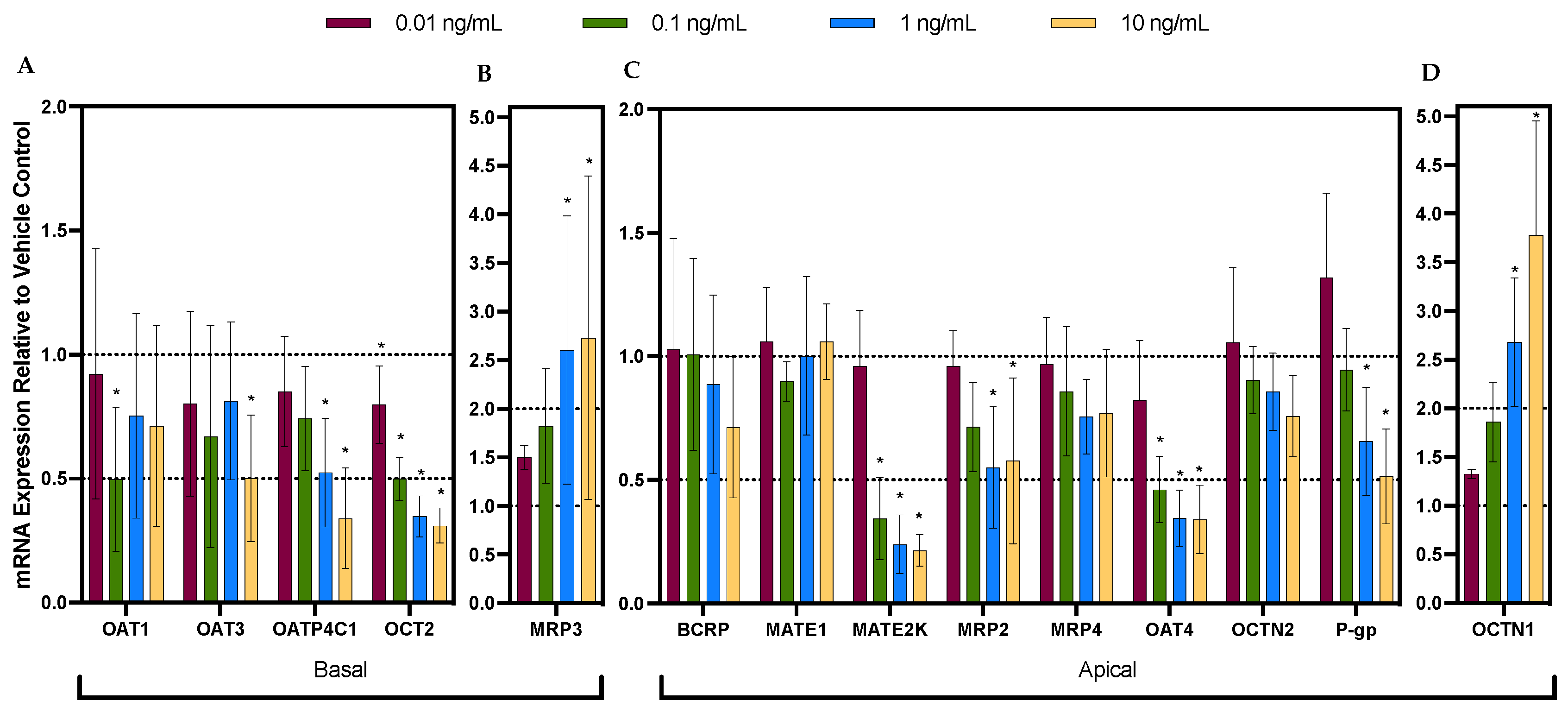
3.1. Cytokine Cocktail Significantly and Differentially Dysregulated the mRNA Expression of Renal Drug Transporters
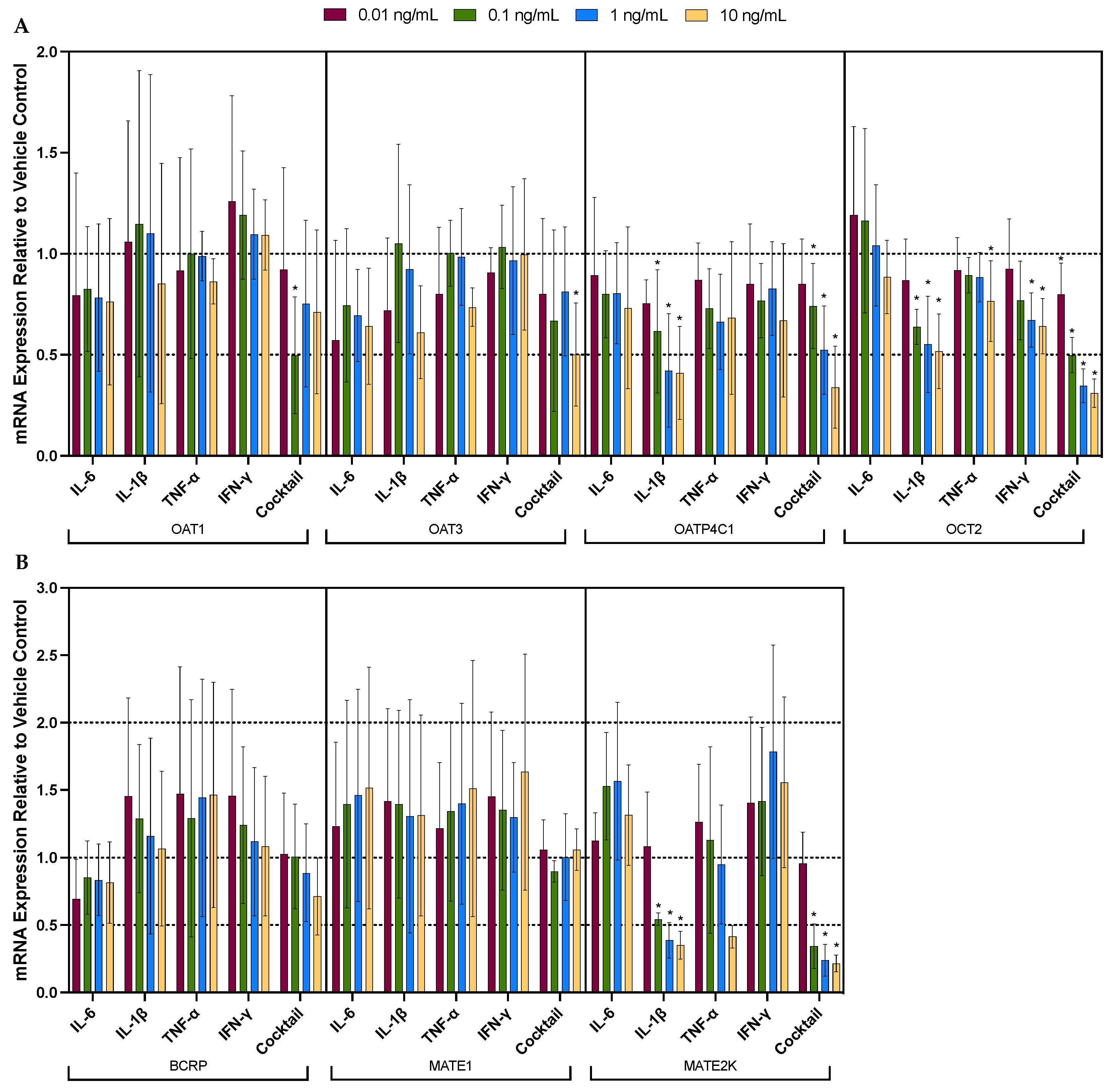
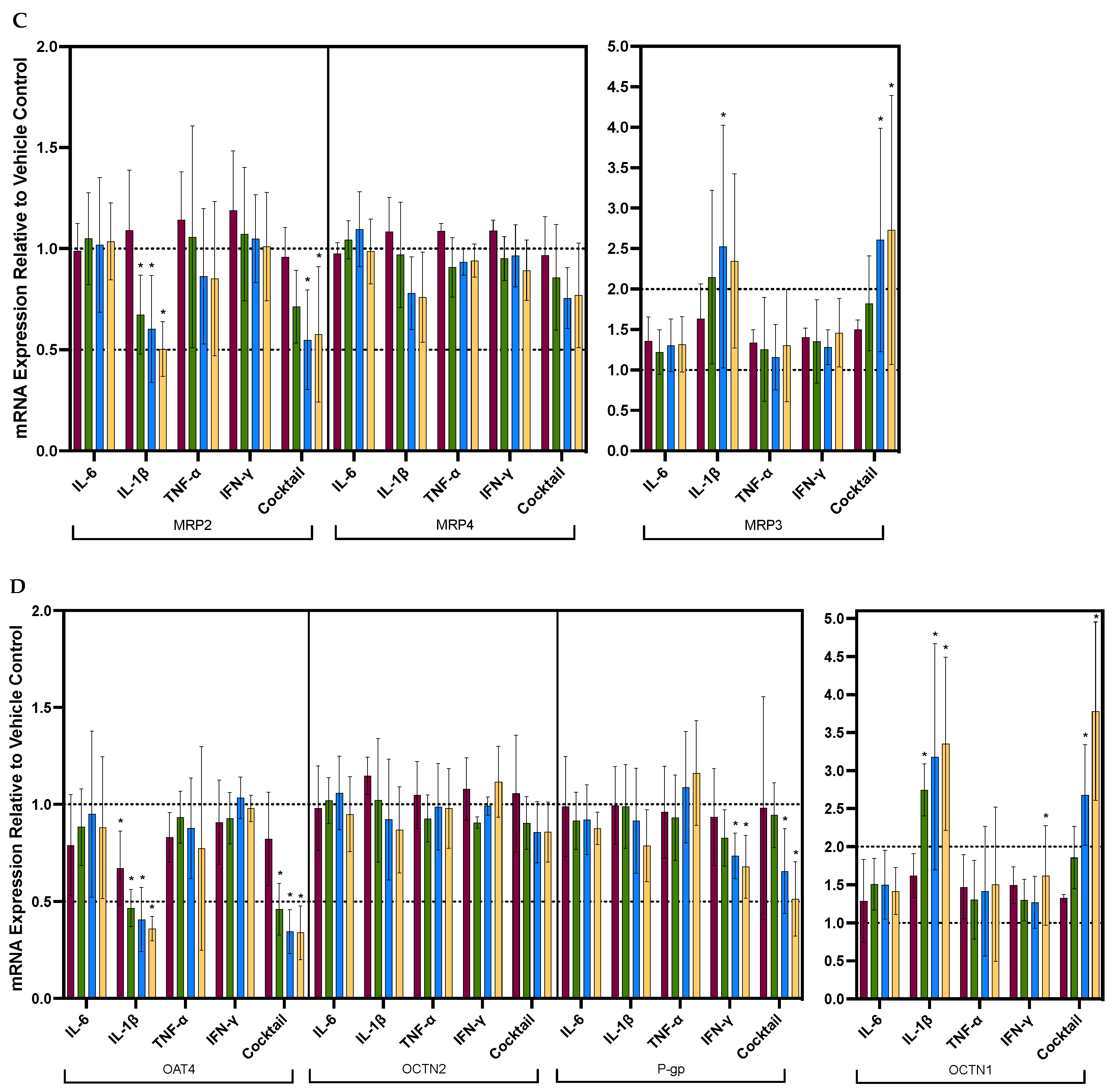
3.2. Individual Cytokines Significantly and Differentially Dysregulated the mRNA Expression of Renal Drug Transporters
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stanke-Labesque, F.; Gautier-Veyret, E.; Chhun, S.; Guilhaumou, R. Inflammation Is a Major Regulator of Drug Metabolizing Enzymes and Transporters: Consequences for the Personalization of Drug Treatment. Pharmacol. Ther. 2020, 215, 107627. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, V.; Teng, S.; Piquette-Miller, M. Regulation of Drug Transporters: During Infection and Inflammation. Mol. Interv. 2007, 7, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, M.J.; Furukawa, C.T.; Koup, J.R.; Shapiro, G.G.; Pierson, W.E.; Bierman, C.W. Altered Theophylline Clearance during an Influenza B Outbreak. Pediatrics 1982, 69, 476–480. [Google Scholar] [CrossRef] [PubMed]
- Morgan, E. Impact of Infectious and Inflammatory Disease on Cytochrome P450–Mediated Drug Metabolism and Pharmacokinetics. Clin. Pharmacol. Ther. 2009, 85, 434–438. [Google Scholar] [CrossRef] [PubMed]
- Sanaee, F.; Clements, J.D.; Waugh, A.W.G.; Fedorak, R.N.; Lewanczuk, R.; Jamali, F. Drug−disease Interaction: Crohn’s Disease Elevates Verapamil Plasma Concentrations but Reduces Response to the Drug Proportional to Disease Activity. Br. J. Clin. Pharmacol. 2011, 72, 787–797. [Google Scholar] [CrossRef] [PubMed]
- Kruger, P.S.; Freir, N.M.; Venkatesh, B.; Robertson, T.A.; Roberts, M.S.; Jones, M. A Preliminary Study of Atorvastatin Plasma Concentrations in Critically Ill Patients with Sepsis. Intensive Care Med. 2009, 35, 717–721. [Google Scholar] [CrossRef] [PubMed]
- Patilea-Vrana, G.; Unadkat, J. Transport vs. Metabolism: What Determines the Pharmacokinetics and Pharmacodynamics of Drugs? Insights from the Extended Clearance Model. Clin. Pharmacol. Ther. 2016, 100, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Le Vee, M.; Gripon, P.; Stieger, B.; Fardel, O. Down-Regulation of Organic Anion Transporter Expression in Human Hepatocytes Exposed to the Proinflammatory Cytokine Interleukin 1β. Drug Metab. Dispos. 2008, 36, 217–222. [Google Scholar] [CrossRef]
- Le Vee, M.; Lecureur, V.; Stieger, B.; Fardel, O. Regulation of Drug Transporter Expression in Human Hepatocytes Exposed to the Proinflammatory Cytokines Tumor Necrosis Factor-α or Interleukin-6. Drug Metab. Dispos. 2009, 37, 685–693. [Google Scholar] [CrossRef]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.-H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An Inflammatory Cytokine Signature Predicts COVID-19 Severity and Survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef]
- Ghazavi, A.; Ganji, A.; Keshavarzian, N.; Rabiemajd, S.; Mosayebi, G. Cytokine Profile and Disease Severity in Patients with COVID-19. Cytokine 2021, 137, 155323. [Google Scholar] [CrossRef]
- Kahle, E.M.; Bolton, M.; Hughes, J.P.; Donnell, D.; Celum, C.; Lingappa, J.R.; Ronald, A.; Cohen, C.R.; de Bruyn, G.; Fong, Y.; et al. Plasma Cytokine Levels and Risk of HIV Type 1 (HIV-1) Transmission and Acquisition: A Nested Case-Control Study Among HIV-1–Serodiscordant Couples. J. Infect Dis. 2015, 211, 1451–1460. [Google Scholar] [CrossRef] [PubMed]
- Mandala, W.L.; Msefula, C.L.; Gondwe, E.N.; Drayson, M.T.; Molyneux, M.E.; MacLennan, C.A. Cytokine Profiles in Malawian Children Presenting with Uncomplicated Malaria, Severe Malarial Anemia, and Cerebral Malaria. Clin. Vaccine Immunol. 2017, 24, e00533-16. [Google Scholar] [CrossRef]
- Kim, H.O.; Kim, H.-S.; Youn, J.-C.; Shin, E.-C.; Park, S. Serum Cytokine Profiles in Healthy Young and Elderly Population Assessed Using Multiplexed Bead-Based Immunoassays. J. Transl. Med. 2011, 9, 113. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, A.E.; Pizzoferrato, T.; Bedford, J.; Morris, A.; Hoffman, R.; Braden, G. Tenofovir-Associated Acute and Chronic Kidney Disease: A Case of Multiple Drug Interactions. Clin. Infect. Dis. 2006, 42, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Arancibia, A.; Drouguett, M.T.; Fuentes, G.; González, G.; González, C.; Thambo, S.; Palombo, G. Pharmacokinetics of Amoxicillin in Subjects with Normal and Impaired Renal Function. Int. J. Clin. Pharmacol. 1982, 20, 447–453. [Google Scholar]
- Benzi, J.R.d.L.; dos Melli, P.P.S.; Duarte, G.; Unadkat, J.D.; Lanchote, V.L. The Impact of Inflammation on the In Vivo Activity of the Renal Transporters OAT1/3 in Pregnant Women Diagnosed with Acute Pyelonephritis. Pharmaceutics 2023, 15, 2427. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Dunvald, A.D.; Järvinen, E.; Mortensen, C.; Stage, T.B. Clinical and Molecular Perspectives on Inflammation-Mediated Regulation of Drug Metabolism and Transport. Clin. Pharmacol. Ther. 2022, 112, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Chousterman, B.G.; Swirski, F.K.; Weber, G.F. Cytokine Storm and Sepsis Disease Pathogenesis. Semin. Immunopathol. 2017, 39, 517–528. [Google Scholar] [CrossRef]
- Tisoncik, J.R.; Korth, M.J.; Simmons, C.P.; Farrar, J.; Martin, T.R.; Katze, M.G. Into the Eye of the Cytokine Storm. Microbiol. Mol. Biol. Rev. 2012, 76, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Yin, Z.; Hu, Y.; Mei, H. Controlling Cytokine Storm Is Vital in COVID-19. Front. Immunol. 2020, 11, 570993. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, K.; Terada, T.; Motohashi, H.; Asaka, J.; Aoki, M.; Katsura, T.; Kamba, T.; Ogawa, O.; Inui, K. Analysis of Regulatory Polymorphisms in Organic Ion Transporter Genes (SLC22A) in the Kidney. J. Hum. Genet 2008, 53, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Nigam, S.K.; Bush, K.T.; Martovetsky, G.; Ahn, S.-Y.; Liu, H.C.; Richard, E.; Bhatnagar, V.; Wu, W. The Organic Anion Transporter (OAT) Family: A Systems Biology Perspective. Physiol. Rev. 2015, 95, 83–123. [Google Scholar] [CrossRef] [PubMed]
- Karimian Pour, N.; McColl, E.R.; Piquette-Miller, M. Impact of Viral Inflammation on the Expression of Renal Drug Transporters in Pregnant Rats. Pharmaceutics 2019, 11, 624. [Google Scholar] [CrossRef]
- Ando, H.; Nishio, Y.; Ito, K.; Nakao, A.; Wang, L.; Zhao, Y.L.; Kitaichi, K.; Takagi, K.; Hasegawa, T. Effect of Endotoxin on P-Glycoprotein-Mediated Biliary and Renal Excretion of Rhodamine-123 in Rats. Antimicrob. Agents Chemother. 2001, 45, 3462–3467. [Google Scholar] [CrossRef]
- Maeda, T.; Hirayama, M.; Kobayashi, D.; Miyazawa, K.; Tamai, I. Mechanism of the Regulation of Organic Cation/Carnitine Transporter 1 (SLC22A4) by Rheumatoid Arthritis-Associated Transcriptional Factor RUNX1 and Inflammatory Cytokines. Drug Metab. Dispos. 2007, 35, 394–401. [Google Scholar] [CrossRef]
- Yin, J.; Wang, J. Renal Drug Transporters and Their Significance in Drug–Drug Interactions. Acta Pharm. Sin. B 2016, 6, 363–373. [Google Scholar] [CrossRef]
- Postler, T.S.; Peng, V.; Bhatt, D.M.; Ghosh, S. Metformin Selectively Dampens the Acute Inflammatory Response through an AMPK-Dependent Mechanism. Sci. Rep. 2021, 11, 18721. [Google Scholar] [CrossRef] [PubMed]
- Bai, B.; Chen, H. Metformin: A Novel Weapon Against Inflammation. Front. Pharmacol. 2021, 12, 622262. [Google Scholar] [CrossRef] [PubMed]
- Widemann, B.C.; Balis, F.M.; Kempf-Bielack, B.; Bielack, S.; Pratt, C.B.; Ferrari, S.; Bacci, G.; Craft, A.W.; Adamson, P.C. High-Dose Methotrexate-Induced Nephrotoxicity in Patients with Osteosarcoma. Cancer 2004, 100, 2222–2232. [Google Scholar] [CrossRef]
- Haustein, K.-O. Therapeutic Range of Cardiac Glycosides. In Cardiac Glycoside Receptors and Positive Inotropy; Steinkopff: Heidelberg, Germany, 1984; pp. 147–153. [Google Scholar]
- Yin, J.; Duan, H.; Wang, J. Impact of Substrate-Dependent Inhibition on Renal Organic Cation Transporters HOCT2 and HMATE1/2-K-Mediated Drug Transport and Intracellular Accumulation. J. Pharmacol. Exp. Ther. 2016, 359, 401–410. [Google Scholar] [CrossRef]
- Brouwer, K.L.R.; Evers, R.; Hayden, E.; Hu, S.; Li, C.Y.; Meyer zu Schwabedissen, H.E.; Neuhoff, S.; Oswald, S.; Piquette-Miller, M.; Saran, C.; et al. Regulation of Drug Transport Proteins—From Mechanisms to Clinical Impact: A White Paper on Behalf of the International Transporter Consortium. Clin. Pharmacol. Ther. 2022, 112, 461–484. [Google Scholar] [CrossRef]
- Morgan, E.T.; Goralski, K.B.; Piquette-Miller, M.; Renton, K.W.; Robertson, G.R.; Chaluvadi, M.R.; Charles, K.A.; Clarke, S.J.; Kacevska, M.; Liddle, C.; et al. Regulation of Drug-Metabolizing Enzymes and Transporters in Infection, Inflammation, and Cancer. Drug Metab. Dispos. 2008, 36, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Abualsunun, W.A.; Piquette-Miller, M. STAT3 Is Involved in IL-6-Mediated Downregulation of Hepatic Transporters in Mice. J. Pharm. Pharm. Sci. 2018, 21, 325s–334s. [Google Scholar] [CrossRef] [PubMed]
- Keller, R.; Klein, M.; Thomas, M.; Dräger, A.; Metzger, U.; Templin, M.F.; Joos, T.O.; Thasler, W.E.; Zell, A.; Zanger, U.M. Coordinating Role of RXRα in Downregulating Hepatic Detoxification during Inflammation Revealed by Fuzzy-Logic Modeling. PLoS Comput. Biol. 2016, 12, e1004431. [Google Scholar] [CrossRef] [PubMed]
- Beigneux, A.P.; Moser, A.H.; Shigenaga, J.K.; Grunfeld, C.; Feingold, K.R. The Acute Phase Response Is Associated with Retinoid X Receptor Repression in Rodent Liver. J. Biol. Chem. 2000, 275, 16390–16399. [Google Scholar] [CrossRef] [PubMed]
- Pascussi, J.-M.; Gerbal-Chaloin, S.; Pichard-Garcia, L.; Daujat, M.; Fabre, J.-M.; Maurel, P.; Vilarem, M.-J. Interleukin-6 Negatively Regulates the Expression of Pregnane X Receptor and Constitutively Activated Receptor in Primary Human Hepatocytes. Biochem. Biophys. Res. Commun. 2000, 274, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Beigneux, A.P.; Moser, A.H.; Shigenaga, J.K.; Grunfeld, C.; Feingold, K.R. Reduction in Cytochrome P-450 Enzyme Expression Is Associated with Repression of CAR (Constitutive Androstane Receptor) and PXR (Pregnane X Receptor) in Mouse Liver during the Acute Phase Response. Biochem. Biophys. Res. Commun. 2002, 293, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Teng, S.; Piquette-Miller, M. The Involvement of the Pregnane X Receptor in Hepatic Gene Regulation during Inflammation in Mice. J. Pharmacol. Exp. Ther. 2005, 312, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, M.; Naito, S. Tissue-Specific MRNA Expression Profiles of Human ATP-Binding Cassette and Solute Carrier Transporter Superfamilies. Drug Metab. Pharmacokinet. 2005, 20, 452–477. [Google Scholar] [CrossRef] [PubMed]
- Human Protein Atlas. Available online: https://www.proteinatlas.org/ (accessed on 22 January 2024).
- Greenbaum, D.; Colangelo, C.; Williams, K.; Gerstein, M. Comparing Protein Abundance and MRNA Expression Levels on a Genomic Scale. Genome Biol. 2003, 4, 117. [Google Scholar] [CrossRef] [PubMed]
- Pou Casellas, C.; Jansen, K.; Rookmaaker, M.B.; Clevers, H.; Verhaar, M.C.; Masereeuw, R. Regulation of Solute Carriers Oct2 and OAT1/3 in the Kidney: A Phylogenetic, Ontogenetic, and Cell Dynamic Perspective. Physiol. Rev. 2022, 102, 993–1024. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, P.; Chowdhury, S.K.; Yucha, R.; Kelly, E.J.; Xiao, G. Emerging Kidney Models to Investigate Metabolism, Transport, and Toxicity of Drugs and Xenobiotics. Drug Metab. Dispos. 2018, 46, 1692–1702. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, P.; Chung, G.; Pye, K.; Yukawa, T.; Imanishi, A.; Takai, Y.; Brown, C.; Wagoner, M.P. Freshly Isolated Primary Human Proximal Tubule Cells as an in Vitro Model for the Detection of Renal Tubular Toxicity. Toxicology 2020, 442, 152535. [Google Scholar] [CrossRef]
- Capaldo, C.T.; Farkas, A.E.; Hilgarth, R.S.; Krug, S.M.; Wolf, M.F.; Benedik, J.K.; Fromm, M.; Koval, M.; Parkos, C.; Nusrat, A. Proinflammatory Cytokine-Induced Tight Junction Remodeling through Dynamic Self-Assembly of Claudins. Mol. Biol. Cell 2014, 25, 2710–2719. [Google Scholar] [CrossRef]
- Li, N.; Gu, L.; Qu, L.; Gong, J.; Li, Q.; Zhu, W.; Li, J. Berberine Attenuates Pro-Inflammatory Cytokine-Induced Tight Junction Disruption in an in vitro Model of Intestinal Epithelial Cells. Eur. J. Pharm. Sci. 2010, 40, 1–8. [Google Scholar] [CrossRef]
- Chen, M.L.; Ge, Z.; Fox, J.G.; Schauer, D.B. Disruption of Tight Junctions and Induction of Proinflammatory Cytokine Responses in Colonic Epithelial Cells by Campylobacter Jejuni. Infect Immun. 2006, 74, 6581–6589. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Condition | IL-6 | IL-1β | TNF-α | IFN-γ | Group Size (N) | References |
|---|---|---|---|---|---|---|
| COVID-19 | 1.6–4823.0 [10] | 1.6–8.3 [10] | 0.8–112.4 [10] | 2.7–434.7 [11] | 1959 [10], 63 [11] | [10,11] |
| HIV-1 infection | 1.5–933.2 | 1.5–2751.2 | 1.5–389.1 | 1.5–2511.9 | 120 | [12] |
| Malaria | 1.2–812.8 | 10.2–812.83 | 1.1–44.7 | 1.5–309.0 | 113 | [13] |
| Healthy adults under the age of 45 | 0.16–37.7 | 0.17–24.0 | 0.93–26.8 | 0.14–126.8 | 55 | [14] |
| Donor ID | Age | Body Mass Index (BMI) | Gender | Race | Cause of Death | Alcohol Use | Tobacco Use | Substance Use |
|---|---|---|---|---|---|---|---|---|
| AJJM288 (D3) | 47 | 26.45 | Female | African American | Stroke | No | No | No |
| AJLA067 (D4) | 34 | 34.9 | Male | African American | Anoxia | No | No | No |
| AJLL353 (D6) | 48 | 28.36 | Female | Caucasian | Stroke | No | Yes | No |
| AKAT206 (D7) | 31 | 24.56 | Female | Caucasian | Anoxia | No | No | No |
| AKAG138 (D9) | 22 | 22.5 | Female | Caucasian | Anoxia | No | No | No |
| Basolateral Transporters | IL-6 | IL-1β | TNF-α | IFN-γ | Cocktail |
|---|---|---|---|---|---|
| OAT1 | ↔ | ↓ 50% a | |||
| OAT3 | ↔ | ↓ 50% | |||
| OATP4C1 | ↔ | ↓ 59% | ↔ | ↔ | ↓ 66% |
| OCT2 | ↔ | ↓ 48% | ↔ | ↔ | ↓ 69% |
| Apical Transporters | |||||
| BCRP | ↔ | ||||
| MATE1 | ↔ | ||||
| MATE2-K | ↔ | ↓ 65% | ↓ 59% | ↔ | ↓ 78% |
| MRP2 | ↔ | ↓ 50% | ↔ | ↔ | ↓ 45% b |
| MRP3 | ↔ | ↑ 152% b | ↔ | ↔ | ↑ 173% |
| MRP4 | ↔ | ||||
| OAT4 | ↔ | ↓ 64% | ↔ | ↔ | ↓ 66% |
| OCTN1 | ↔ | ↑ 235% | ↔ | ↔ | ↑ 278% |
| OCTN2 | ↔ | ||||
| P-gp | ↔ | ↔ | ↔ | ↓ 32% | ↓ 49% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsang, Y.P.; Hao, T.; Mao, Q.; Kelly, E.J.; Unadkat, J.D. Dysregulation of the mRNA Expression of Human Renal Drug Transporters by Proinflammatory Cytokines in Primary Human Proximal Tubular Epithelial Cells. Pharmaceutics 2024, 16, 285. https://doi.org/10.3390/pharmaceutics16020285
Tsang YP, Hao T, Mao Q, Kelly EJ, Unadkat JD. Dysregulation of the mRNA Expression of Human Renal Drug Transporters by Proinflammatory Cytokines in Primary Human Proximal Tubular Epithelial Cells. Pharmaceutics. 2024; 16(2):285. https://doi.org/10.3390/pharmaceutics16020285
Chicago/Turabian StyleTsang, Yik Pui, Tianran Hao, Qingcheng Mao, Edward J. Kelly, and Jashvant D. Unadkat. 2024. "Dysregulation of the mRNA Expression of Human Renal Drug Transporters by Proinflammatory Cytokines in Primary Human Proximal Tubular Epithelial Cells" Pharmaceutics 16, no. 2: 285. https://doi.org/10.3390/pharmaceutics16020285