
The Effect of Cholesterol Content on the Adjuvant Activity of Nucleic-Acid-Free Lipid Nanoparticles
 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Materials
2.3. Methods
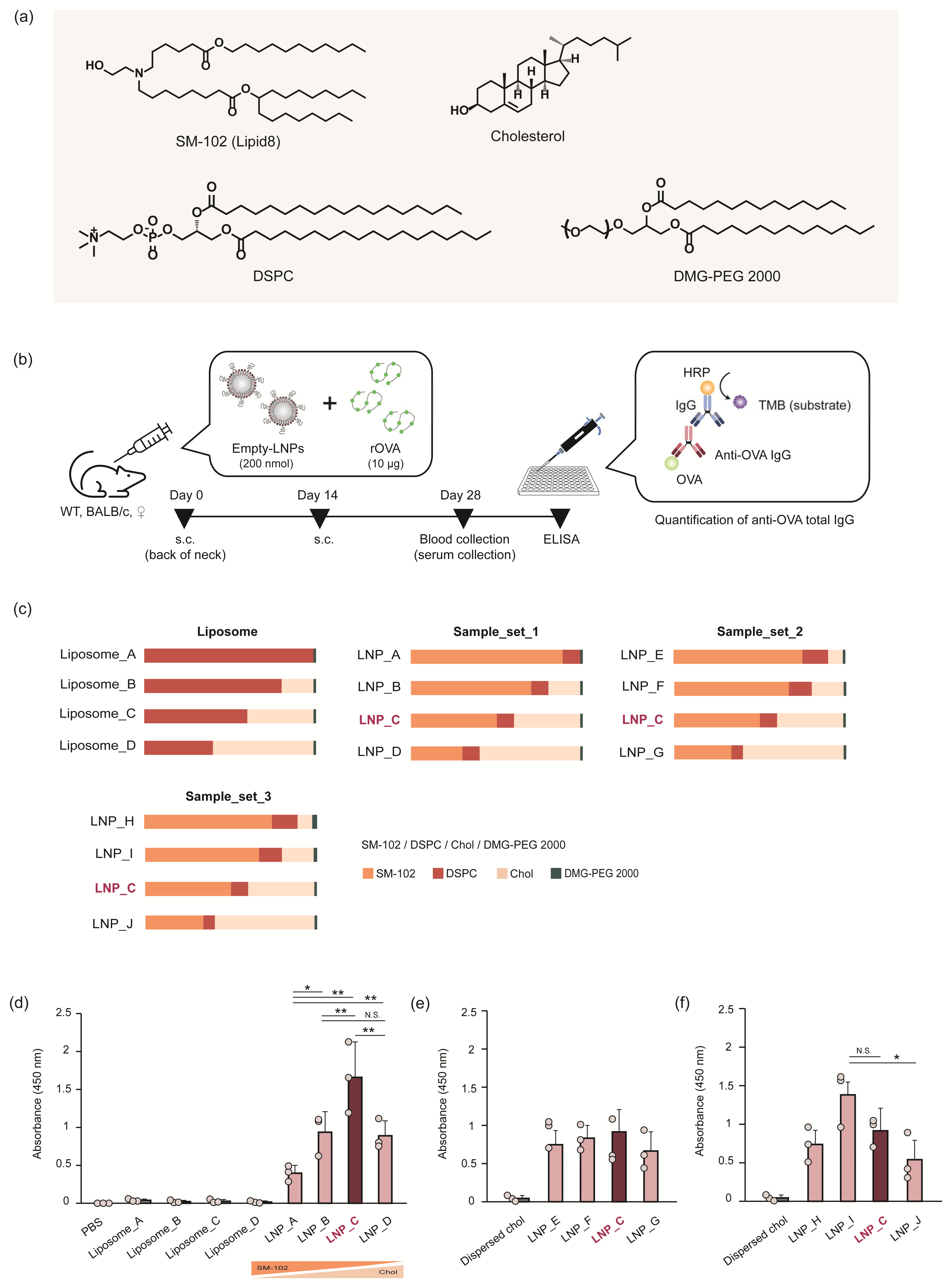
2.3.1. Preparation of LNPs
2.3.2. Characterization of the LNPs
2.3.3. OVA Antigen-Specific Total IgG Quantification
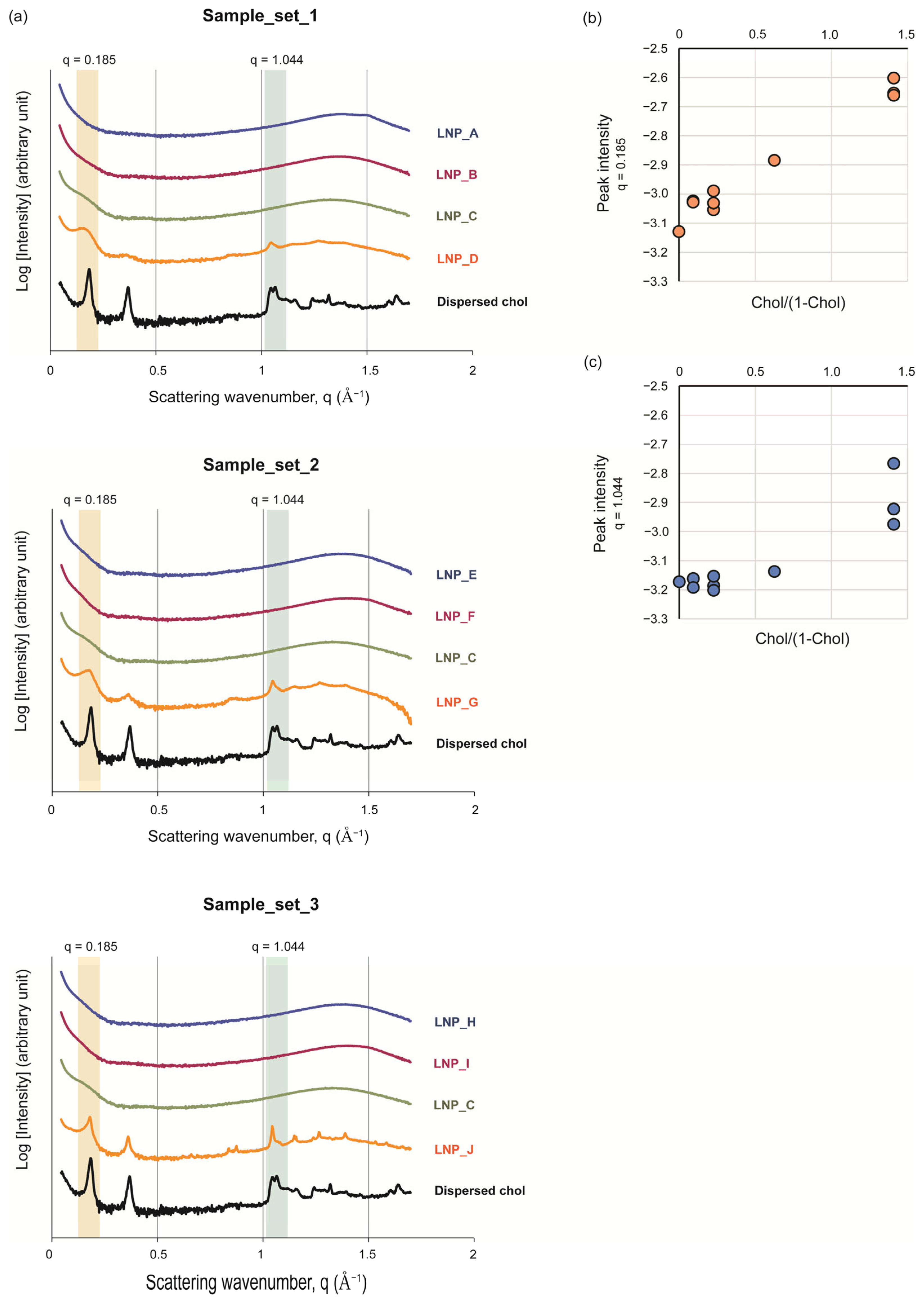
2.3.4. SAXS
2.3.5. Cryo-TEM
3. Results
3.1. Adjuvant Activity of the LNPs Did Not Depend on the Amount of Ionizable Lipids
3.2. The Amounts of Ionizable Lipid and Cholesterol Affect the Particle Morphology
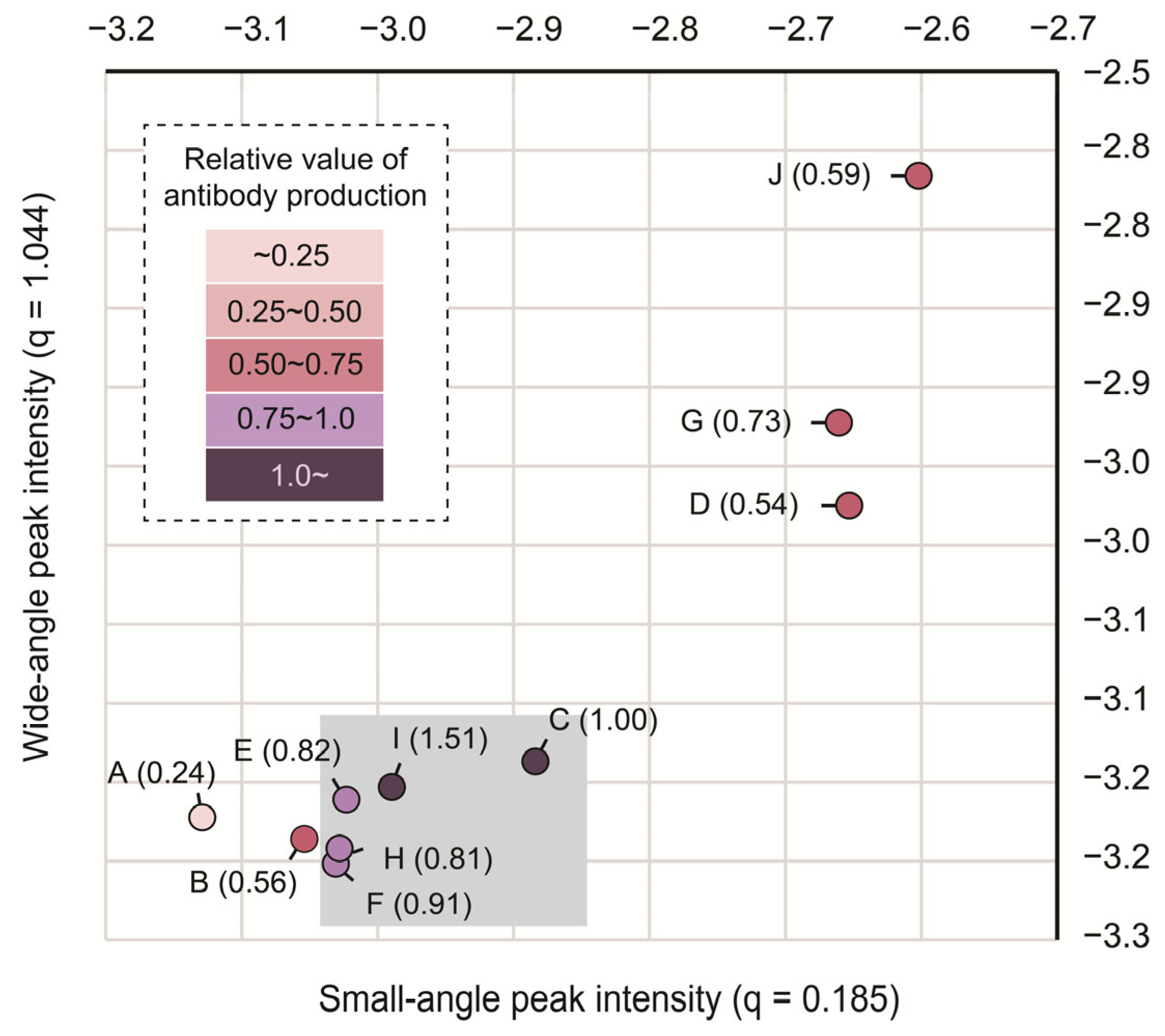
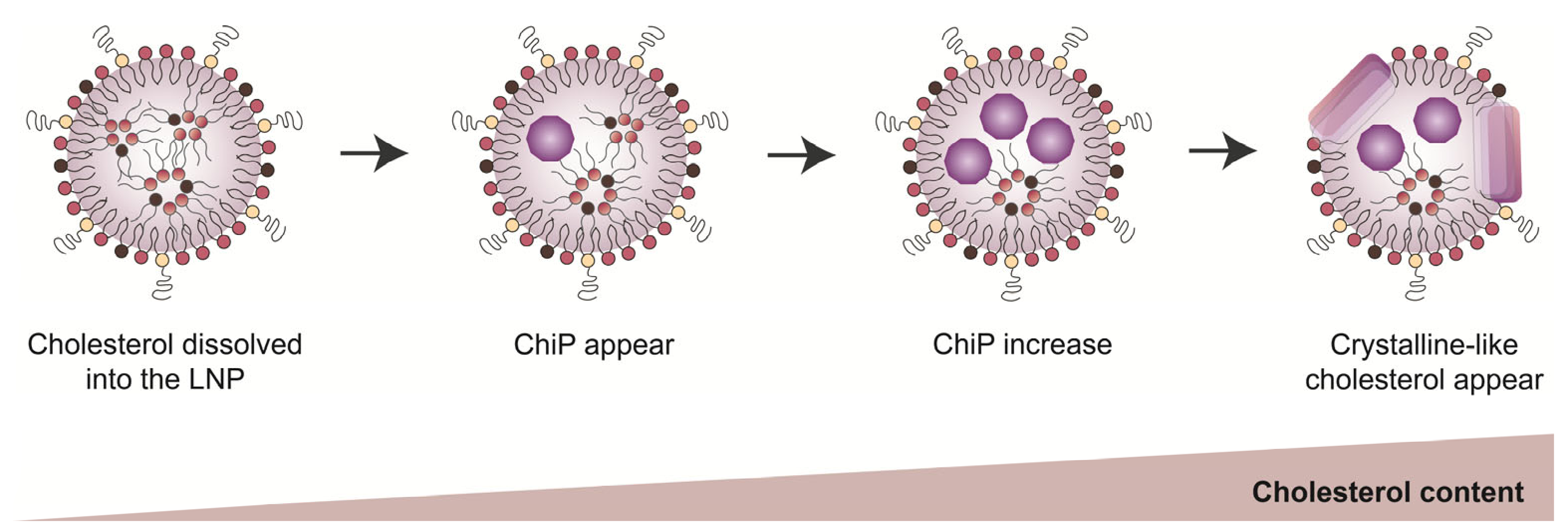
3.3. The Intra-Particle State of Cholesterol Varied Depending on Its Amount in the LNPs
3.4. The Intermediate State of the Cholesterol in LNPs Could Be Important for Adjuvant Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. MRNA Vaccines—A New Era in Vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef]
- Chaudhary, N.; Weissman, D.; Whitehead, K.A. MRNA Vaccines for Infectious Diseases: Principles, Delivery and Clinical Translation. Nat. Rev. Drug Discov. 2021, 20, 817–838. [Google Scholar] [CrossRef] [PubMed]
- Gote, V.; Bolla, P.K.; Kommineni, N.; Butreddy, A.; Nukala, P.K.; Palakurthi, S.S.; Khan, W. A Comprehensive Review of MRNA Vaccines. Int. J. Mol. Sci. 2023, 24, 2700. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.; Kwon, M.; Im, S.; Lee, K.; Lee, H. MRNA Vaccines: The Most Recent Clinical Applications of Synthetic MRNA. Arch. Pharm. Res. 2022, 45, 245–262. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.; Hur, S. Immunogenicity of in Vitro-Transcribed RNA. Acc. Chem. Res. 2021, 54, 4012–4023. [Google Scholar] [CrossRef] [PubMed]
- Karikó, K.; Muramatsu, H.; Welsh, F.A.; Ludwig, J.; Kato, H.; Akira, S.; Weissman, D. Incorporation of Pseudouridine Into MRNA Yields Superior Nonimmunogenic Vector with Increased Translational Capacity and Biological Stability. Mol. Ther. 2008, 16, 1833–1840. [Google Scholar] [CrossRef]
- Andries, O.; Mc Cafferty, S.; De Smedt, S.C.; Weiss, R.; Sanders, N.N.; Kitada, T. N1-Methylpseudouridine-Incorporated MRNA Outperforms Pseudouridine-Incorporated MRNA by Providing Enhanced Protein Expression and Reduced Immunogenicity in Mammalian Cell Lines and Mice. J. Control. Release 2015, 217, 337–344. [Google Scholar] [CrossRef]
- Decroly, E.; Ferron, F.; Lescar, J.; Canard, B. Conventional and Unconventional Mechanisms for Capping Viral MRNA. Nat. Rev. Microbiol. 2011, 10, 51–65. [Google Scholar] [CrossRef]
- Ramanathan, A.; Robb, G.B.; Chan, S.-H. MRNA Capping: Biological Functions and Applications. Nucleic Acids Res. 2016, 44, 7511–7526. [Google Scholar] [CrossRef]
- Baiersdörfer, M.; Boros, G.; Muramatsu, H.; Mahiny, A.; Vlatkovic, I.; Sahin, U.; Karikó, K. A Facile Method for the Removal of DsRNA Contaminant from In Vitro-Transcribed MRNA. Mol. Ther. Nucleic Acids 2019, 15, 26–35. [Google Scholar] [CrossRef]
- Piao, X.; Yadav, V.; Wang, E.; Chang, W.; Tau, L.; Lindenmuth, B.E.; Wang, S.X. Double-Stranded RNA Reduction by Chaotropic Agents during in Vitro Transcription of Messenger RNA. Mol. Ther. Nucleic Acids 2022, 29, 618–624. [Google Scholar] [CrossRef]
- Huang, P.; Deng, H.; Zhou, Y.; Chen, X. The Roles of Polymers in MRNA Delivery. Matter 2022, 5, 1670–1699. [Google Scholar] [CrossRef]
- Yang, W.; Mixich, L.; Boonstra, E.; Cabral, H. Polymer-Based MRNA Delivery Strategies for Advanced Therapies. Adv. Healthc. Mater. 2023, 12, 2202688. [Google Scholar] [CrossRef]
- Hou, X.; Zaks, T.; Langer, R.; Dong, Y. Lipid Nanoparticles for MRNA Delivery. Nat. Rev. Mater. 2021, 6, 1078–1094. [Google Scholar] [CrossRef]
- Tenchov, R.; Bird, R.; Curtze, A.E.; Zhou, Q. Lipid Nanoparticles─From Liposomes to MRNA Vaccine Delivery, a Landscape of Research Diversity and Advancement. ACS Nano 2021, 15, 16982–17015. [Google Scholar] [CrossRef] [PubMed]
- Spadea, A.; Jackman, M.; Cui, L.; Pereira, S.; Lawrence, M.J.; Campbell, R.A.; Ashford, M. Nucleic Acid-Loaded Lipid Nanoparticle Interactions with Model Endosomal Membranes. ACS Appl. Mater. Interfaces 2022, 14, 30371–30384. [Google Scholar] [CrossRef] [PubMed]
- Aliakbarinodehi, N.; Gallud, A.; Mapar, M.; Wesén, E.; Heydari, S.; Jing, Y.; Emilsson, G.; Liu, K.; Sabirsh, A.; Zhdanov, V.P.; et al. Interaction Kinetics of Individual MRNA-Containing Lipid Nanoparticles with an Endosomal Membrane Mimic: Dependence on PH, Protein Corona Formation, and Lipoprotein Depletion. ACS Nano 2022, 16, 20163–20173. [Google Scholar] [CrossRef] [PubMed]
- Hald Albertsen, C.; Kulkarni, J.A.; Witzigmann, D.; Lind, M.; Petersson, K.; Simonsen, J.B. The Role of Lipid Components in Lipid Nanoparticles for Vaccines and Gene Therapy. Adv. Drug Deliv. Rev. 2022, 188, 114416. [Google Scholar] [CrossRef] [PubMed]
- Swetha, K.; Kotla, N.G.; Tunki, L.; Jayaraj, A.; Bhargava, S.K.; Hu, H.; Bonam, S.R.; Kurapati, R. Recent Advances in the Lipid Nanoparticle-Mediated Delivery of MRNA Vaccines. Vaccines 2023, 11, 658. [Google Scholar] [CrossRef] [PubMed]
- Jackson, L.A.; Anderson, E.J.; Rouphael, N.G.; Roberts, P.C.; Makhene, M.; Coler, R.N.; McCullough, M.P.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; et al. An MRNA Vaccine against SARS-CoV-2—Preliminary Report. N. Engl. J. Med. 2020, 383, 1920–1931. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the MRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Walsh, E.E.; Frenck, R.W.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and Immunogenicity of Two RNA-Based Covid-19 Vaccine Candidates. N. Engl. J. Med. 2020, 383, 2439–2450. [Google Scholar] [CrossRef] [PubMed]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 MRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Linares-Fernández, S.; Lacroix, C.; Exposito, J.-Y.; Verrier, B. Tailoring MRNA Vaccine to Balance Innate/Adaptive Immune Response. Trends Mol. Med. 2020, 26, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Ishii, K.J.; Akira, S. Innate Immune Recognition of Nucleic Acids: Beyond Toll-like Receptors. Int. J. Cancer 2005, 117, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Desmet, C.J.; Ishii, K.J. Nucleic Acid Sensing at the Interface between Innate and Adaptive Immunity in Vaccination. Nat. Rev. Immunol. 2012, 12, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Kobiyama, K.; Ishii, K.J. Making Innate Sense of MRNA Vaccine Adjuvanticity. Nat. Immunol. 2022, 23, 474–476. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Lee, A.; Grigoryan, L.; Arunachalam, P.S.; Scott, M.K.D.; Trisal, M.; Wimmers, F.; Sanyal, M.; Weidenbacher, P.A.; Feng, Y.; et al. Mechanisms of Innate and Adaptive Immunity to the Pfizer-BioNTech BNT162b2 Vaccine. Nat. Immunol. 2022, 23, 543–555. [Google Scholar] [CrossRef]
- Swaminathan, G.; Thoryk, E.A.; Cox, K.S.; Meschino, S.; Dubey, S.A.; Vora, K.A.; Celano, R.; Gindy, M.; Casimiro, D.R.; Bett, A.J. A Novel Lipid Nanoparticle Adjuvant Significantly Enhances B Cell and T Cell Responses to Sub-Unit Vaccine Antigens. Vaccine 2016, 34, 110–119. [Google Scholar] [CrossRef]
- Thoryk, E.A.; Swaminathan, G.; Meschino, S.; Cox, K.S.; Gindy, M.; Casimiro, D.R.; Bett, A.J. Co-Administration of Lipid Nanoparticles and Sub-Unit Vaccine Antigens Is Required for Increase in Antigen-Specific Immune Responses in Mice. Vaccines 2016, 4, 47. [Google Scholar] [CrossRef]
- Alameh, M.-G.; Tombácz, I.; Bettini, E.; Lederer, K.; Sittplangkoon, C.; Wilmore, J.R.; Gaudette, B.T.; Soliman, O.Y.; Pine, M.; Hicks, P.; et al. Lipid Nanoparticles Enhance the Efficacy of MRNA and Protein Subunit Vaccines by Inducing Robust T Follicular Helper Cell and Humoral Responses. Immunity 2021, 54, 2877–2892.E7. [Google Scholar] [CrossRef]
- Ndeupen, S.; Qin, Z.; Jacobsen, S.; Bouteau, A.; Estanbouli, H.; Igyártó, B.Z. The MRNA-LNP Platform’s Lipid Nanoparticle Component Used in Preclinical Vaccine Studies Is Highly Inflammatory. iScience 2021, 24, 103479. [Google Scholar] [CrossRef]
- Tanaka, H.; Oasa, S.; Kinjo, M.; Tange, K.; Nakai, Y.; Harashima, H.; Akita, H. Temperature and PH Sensitivity of a Stabilized Self-Nanoemulsion Formed Using an Ionizable Lipid-like Material via an Oil-to-Surfactant Transition. Colloids Surf. B Biointerfaces 2017, 151, 95–101. [Google Scholar] [CrossRef]
- Frey, M.; Bobbala, S.; Karabin, N.; Scott, E. Influences of Nanocarrier Morphology on Therapeutic Immunomodulation. Nanomedicine 2018, 13, 1795–1811. [Google Scholar] [CrossRef] [PubMed]
- Niikura, K.; Matsunaga, T.; Suzuki, T.; Kobayashi, S.; Yamaguchi, H.; Orba, Y.; Kawaguchi, A.; Hasegawa, H.; Kajino, K.; Ninomiya, T.; et al. Gold Nanoparticles as a Vaccine Platform: Influence of Size and Shape on Immunological Responses in Vitro and in Vivo. ACS Nano 2013, 7, 3926–3938. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Sun, L.; Zhang, Y.; Dove, A.P.; O’Reilly, R.K.; Chen, G. Shape Effect of Glyco-Nanoparticles on Macrophage Cellular Uptake and Immune Response. ACS Macro Lett. 2016, 5, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Ridolfo, R.; Tavakoli, S.; Junnuthula, V.; Williams, D.S.; Urtti, A.; van Hest, J.C.M. Exploring the Impact of Morphology on the Properties of Biodegradable Nanoparticles and Their Diffusion in Complex Biological Medium. Biomacromolecules 2021, 22, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Hassett, K.J.; Benenato, K.E.; Jacquinet, E.; Lee, A.; Woods, A.; Yuzhakov, O.; Himansu, S.; Deterling, J.; Geilich, B.M.; Ketova, T.; et al. Optimization of Lipid Nanoparticles for Intramuscular Administration of MRNA Vaccines. Mol. Ther. Nucleic Acids 2019, 15, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Corbett, K.S.; Edwards, D.K.; Leist, S.R.; Abiona, O.M.; Boyoglu-Barnum, S.; Gillespie, R.A.; Himansu, S.; Schäfer, A.; Ziwawo, C.T.; DiPiazza, A.T.; et al. SARS-CoV-2 MRNA Vaccine Design Enabled by Prototype Pathogen Preparedness. Nature 2020, 586, 567–571. [Google Scholar] [CrossRef]
- Sabnis, S.; Kumarasinghe, E.S.; Salerno, T.; Mihai, C.; Ketova, T.; Senn, J.J.; Lynn, A.; Bulychev, A.; McFadyen, I.; Chan, J.; et al. A Novel Amino Lipid Series for MRNA Delivery: Improved Endosomal Escape and Sustained Pharmacology and Safety in Non-Human Primates. Mol. Ther. 2018, 26, 1509–1519. [Google Scholar] [CrossRef]
- Shimizu, N.; Yatabe, K.; Nagatani, Y.; Saijyo, S.; Kosuge, T.; Igarashi, N. Software Development for Analysis of Small-Angle X-ray Scattering Data. AIP Conf. Proc. 2016, 1741, 050017. [Google Scholar]
- Tanaka, H.; Hagiwara, S.; Shirane, D.; Yamakawa, T.; Sato, Y.; Matsumoto, C.; Ishizaki, K.; Hishinuma, M.; Chida, K.; Sasaki, K.; et al. Ready-to-Use-Type Lyophilized Lipid Nanoparticle Formulation for the Postencapsulation of Messenger RNA. ACS Nano 2023, 17, 2588–2601. [Google Scholar] [CrossRef]
- Chander, N.; Basha, G.; Yan Cheng, M.H.; Witzigmann, D.; Cullis, P.R. Lipid Nanoparticle MRNA Systems Containing High Levels of Sphingomyelin Engender Higher Protein Expression in Hepatic and Extra-Hepatic Tissues. Mol. Ther. Methods Clin. Dev. 2023, 30, 235–245. [Google Scholar] [CrossRef]
- Schoenmaker, L.; Witzigmann, D.; Kulkarni, J.A.; Verbeke, R.; Kersten, G.; Jiskoot, W.; Crommelin, D.J.A. MRNA-Lipid Nanoparticle COVID-19 Vaccines: Structure and Stability. Int. J. Pharm. 2021, 601, 120586. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.N.; Brown, D.A.; London, E. On the Origin of Sphingolipid/Cholesterol-Rich Detergent-Insoluble Cell Membranes: Physiological Concentrations of Cholesterol and Sphingolipid Induce Formation of a Detergent-Insoluble, Liquid-Ordered Lipid Phase in Model Membranes. Biochemistry 1997, 36, 10944–10953. [Google Scholar] [CrossRef] [PubMed]
- Soto-Arriaza, M.A.; Olivares-Ortega, C.; Quina, F.H.; Aguilar, L.F.; Sotomayor, C.P. Effect of Cholesterol Content on the Structural and Dynamic Membrane Properties of DMPC/DSPC Large Unilamellar Bilayers. Biochim. Biophys. Acta (BBA)-Biomembr. 2013, 1828, 2763–2769. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.C.; Wrenn, S.P. Nanoscale Phase Separation in DSPC–Cholesterol Systems. Langmuir 2013, 29, 9832–9840. [Google Scholar] [CrossRef]
- Stevens, M.M.; Honerkamp-Smith, A.R.; Keller, S.L. Solubility Limits of Cholesterol, Lanosterol, Ergosterol, Stigmasterol, and β-Sitosterol in Electroformed Lipid Vesicles. Soft Matter 2010, 6, 5882. [Google Scholar] [CrossRef] [PubMed]
- Connors, J.; Joyner, D.; Mege, N.J.; Cusimano, G.M.; Bell, M.R.; Marcy, J.; Taramangalam, B.; Kim, K.M.; Lin, P.J.C.; Tam, Y.K.; et al. Lipid Nanoparticles (LNP) Induce Activation and Maturation of Antigen Presenting Cells in Young and Aged Individuals. Commun. Biol. 2023, 6, 188. [Google Scholar] [CrossRef] [PubMed]
- Bevers, S.; Kooijmans, S.A.A.; Van de Velde, E.; Evers, M.J.W.; Seghers, S.; Gitz-Francois, J.J.J.M.; van Kronenburg, N.C.H.; Fens, M.H.A.M.; Mastrobattista, E.; Hassler, L.; et al. MRNA-LNP Vaccines Tuned for Systemic Immunization Induce Strong Antitumor Immunity by Engaging Splenic Immune Cells. Mol. Ther. 2022, 30, 3078–3094. [Google Scholar] [CrossRef]
- Oyama, R.; Ishigame, H.; Tanaka, H.; Tateshita, N.; Itazawa, M.; Imai, R.; Nishiumi, N.; Kishikawa, J.; Kato, T.; Anindita, J.; et al. An Ionizable Lipid Material with a Vitamin E Scaffold as an MRNA Vaccine Platform for Efficient Cytotoxic T Cell Responses. ACS Nano 2023, 17, 18758–18774. [Google Scholar] [CrossRef]
- Hsu, L.Y.; Nordman, C.E. Phase Transition and Crystal Structure of the 37 Degrees C Form of Cholesterol. Science 1983, 220, 604–606. [Google Scholar] [CrossRef] [PubMed]
- Shieh, H.S.; Hoard, L.G.; Nordman, C.E. Crystal Structure of Anhydrous Cholesterol. Nature 1977, 267, 287–289. [Google Scholar] [CrossRef]
- Craven, B.M. Crystal Structure of Cholesterol Monohydrate. Nature 1976, 260, 727–729. [Google Scholar] [CrossRef] [PubMed]
- Bakthavatchalam, M.; Venkataraman, J.; Ramana, R.J.; Jain, M.; Singh, B.; Thanigai, A.K.; Velyoudam, V.; Manickam Neethirajan, S.; Tiwari, M.K.; Agarwal, A.K.; et al. Morphological and Elemental Mapping of Gallstones Using Synchrotron Microtomography and Synchrotron X-Ray Fluorescence Spectroscopy. JGH Open 2019, 3, 381–387. [Google Scholar] [CrossRef]
- Sebastiani, F.; Yanez Arteta, M.; Lerche, M.; Porcar, L.; Lang, C.; Bragg, R.A.; Elmore, C.S.; Krishnamurthy, V.R.; Russell, R.A.; Darwish, T.; et al. Apolipoprotein E Binding Drives Structural and Compositional Rearrangement of MRNA-Containing Lipid Nanoparticles. ACS Nano 2021, 15, 6709–6722. [Google Scholar] [CrossRef] [PubMed]
- Eygeris, Y.; Patel, S.; Jozic, A.; Sahay, G. Deconvoluting Lipid Nanoparticle Structure for Messenger RNA Delivery. Nano Lett. 2020, 20, 4543–4549. [Google Scholar] [CrossRef]
- Shu, F.; Chen, J.; Ma, X.; Fan, Y.; Yu, L.; Zheng, W.; Amrein, M.W.; Xia, T.; Shi, Y. Cholesterol Crystal-Mediated Inflammation Is Driven by Plasma Membrane Destabilization. Front. Immunol. 2018, 9, 1163. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Particle | Composition (%) | Size (d.nm) | PdI | ZP (mV) | |||
|---|---|---|---|---|---|---|---|
| SM-102 | DSPC | Chol | PEG2K | ||||
| Dispersed chol. | 0 | 0 | 98.5 | 1.5 | 147 ± 10.2 | 0.23 ± 0.0 | −4.2 ± 1.2 |
| Liposome_A | 0 | 98.5 | 0 | 1.5 | 183 ± 4.9 | 0.53 ± 0.1 | 0.7 ± 0.4 |
| Liposome_B | 0 | 80 | 18.5 | 1.5 | 178 ± 6.4 | 0.36 ± 0.1 | 0.1 ± 0.1 |
| Liposome_C | 0 | 60 | 38.5 | 1.5 | 137 ± 11.9 | 0.30 ± 0.1 | −3.1 ± 1.5 |
| Liposome_D | 0 | 40 | 58.5 | 1.5 | 108 ± 8.0 | 0.26 ± 0.0 | −6.6 ± 1.1 |
| Sample_set_1 | |||||||
| LNP_A | 88.5 | 10 | 0 | 1.5 | 113 ± 2.0 | 0.11 ± 0.0 | 6.0 ± 0.9 |
| LNP_B | 70 | 10 | 18.5 | 1.5 | 99 ± 1.6 | 0.12 ± 0.0 | 1.8 ± 0.9 |
| LNP_C | 50 | 10 | 38.5 | 1.5 | 89 ± 2.5 | 0.13 ± 0.0 | −0.4 ± 0.5 |
| LNP_D | 30 | 10 | 58.5 | 1.5 | 99 ± 1.8 | 0.16 ± 0.0 | −0.3 ± 0.2 |
| Sample_set_2 | |||||||
| LNP_E | 75 | 15 | 8.5 | 1.5 | 90 ± 6.7 | 0.15 ± 0.0 | 0.5 ± 0.6 |
| LNP_F | 66.7 | 13.3 | 18.5 | 1.5 | 86 ± 4.2 | 0.15 ± 0.0 | −0.5 ± 0.7 |
| LNP_C | 50 | 10 | 38.5 | 1.5 | 89 ± 2.5 | 0.13 ± 0.0 | −0.4 ± 0.5 |
| LNP_G | 33.3 | 6.7 | 58.5 | 1.5 | 91 ± 3.8 | 0.24 ± 0.1 | −2.6 ± 0.9 |
| Sample_set_3 | |||||||
| LNP_H | 74.4 | 14.9 | 8.5 | 2.2 | 92 ± 4.1 | 0.16 ± 0.0 | 5.6 ± 1.4 |
| LNP_I | 66.2 | 13.3 | 18.5 | 2.0 | 93 ± 5.5 | 0.21 ± 0.0 | 0.6 ± 0.4 |
| LNP_C | 50 | 10 | 38.5 | 1.5 | 89 ± 2.5 | 0.13 ± 0.0 | −0.4 ± 0.5 |
| LNP_J | 33.7 | 6.8 | 58.5 | 1.0 | 105 ± 5.2 | 0.24 ± 0.0 | −4.5 ± 1.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anindita, J.; Tanaka, H.; Yamakawa, T.; Sato, Y.; Matsumoto, C.; Ishizaki, K.; Oyama, T.; Suzuki, S.; Ueda, K.; Higashi, K.; et al. The Effect of Cholesterol Content on the Adjuvant Activity of Nucleic-Acid-Free Lipid Nanoparticles. Pharmaceutics 2024, 16, 181. https://doi.org/10.3390/pharmaceutics16020181
Anindita J, Tanaka H, Yamakawa T, Sato Y, Matsumoto C, Ishizaki K, Oyama T, Suzuki S, Ueda K, Higashi K, et al. The Effect of Cholesterol Content on the Adjuvant Activity of Nucleic-Acid-Free Lipid Nanoparticles. Pharmaceutics. 2024; 16(2):181. https://doi.org/10.3390/pharmaceutics16020181
Chicago/Turabian StyleAnindita, Jessica, Hiroki Tanaka, Takuma Yamakawa, Yuka Sato, Chika Matsumoto, Kota Ishizaki, Taiji Oyama, Satoko Suzuki, Keisuke Ueda, Kenjirou Higashi, and et al. 2024. "The Effect of Cholesterol Content on the Adjuvant Activity of Nucleic-Acid-Free Lipid Nanoparticles" Pharmaceutics 16, no. 2: 181. https://doi.org/10.3390/pharmaceutics16020181