1. Introduction
Nucleic-acid-based vaccines are based on the use of plasmid DNA (pDNA) or messenger RNA (mRNA) as a source of antigens. Upon the administration of such a vaccine, these genetic materials need to be delivered into the nucleus/cytoplasm, their site of action, where they will be transcribed/translated into antigen proteins. Since the mRNA can function in the cytoplasm and nuclear localization is not required, the mRNA can provide efficient and robust gene transfer, even in non-dividing cells [
1,
2,
3]. On the other hand, since mRNA is highly susceptible to enzymatic degradation in extracellular fluid, the development of suitable delivery technology is a prerequisite for its clinical application [
3,
4]. One of the more promising carriers for RNA delivery is lipid nanoparticles (LNPs), which contain an ionizable lipid as the main component. In 2018, ONPATTRO
® (Alnylam Pharmaceutical, Cambridge, MA, USA), an LNP-based therapeutic using small interfering RNA (siRNA), became the first ever approved RNA interference (RNAi) therapeutic for the treatment of hereditary transthyretin-mediated amyloidosis [
5,
6,
7,
8,
9]. Moreover, the approval of RNA vaccines (mRNA-LNPs) against severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) [
10,
11,
12,
13,
14] further demonstrated the versatility of the LNP as an mRNA carrier.
We have been developing LNPs that are composed of a series of ionizable lipids, referred to as SS-cleavable and pH-activated lipid-like materials (ssPalm). The ssPalm(s) respond to the difference in extracellular and intracellular environments based on their dual-sensing motifs: tertiary amines and a disulfide bond. After cellular uptake, the neutrally charged LNPs
ssPalm are sorted into endosomes. When arriving in the endosome, the LNPs are exposed to the acidic environment in the endosomal compartment (pH 6.5–5.5) and develop positive charges on their surface through the protonation of tertiary amines. The positively charged LNPs then cause endosomal membrane disruption/destabilization, which results in the endosomal escape of their cargo. Following this endosomal escape event, the disulfide bond in LNPs
ssPalm will be cleaved by glutathione in the reductive environment of the cytoplasm. This then triggers the decapsulation of the mRNA cargoes, releasing them into the cytoplasm [
15,
16,
17].
As additional modifications of the ionizable lipids, we have focused on the hydrophobic scaffold and the linker between the amine and the scaffolds. Derivatives of ssPalms can be developed by changing their hydrophobic scaffolds. Changing the hydrophobic scaffolds from fatty acids such as myristic acid (ssPalmM) to retinoic acid (vitamin A; ssPalmA) allowed the intracellular trafficking of the nucleic acid cargo to be controlled [
15]. Types of ssPalm with vitamin E scaffolds (ssPalmE) have become a focus of vaccine development since they are equipped with both gene expression activity and immune activation properties [
16]. The vitamin E scaffolds were assumed to be suitable for cancer vaccines because they can efficiently activate cell-mediated immunity via the type I interferon pathway [
18]. In addition to the modification of hydrophobic scaffolds, the incorporation of a self-degradable phenyl ester moiety can be used to improve the gene transfer activity of the mRNA-LNP [
15,
17]. The cleavage of disulfide bonds by reducing agents in the cytoplasm elevates the concentration of hydrophobic thiols in the particle. Within those limited intraparticle spaces, the concentrated thiols attack the phenyl ester and trigger a nucleophilic substitution reaction. This self-degradation reaction is irreversible, leading to the more efficient collapse of particles, which further enhances the release of the nucleic acid cargo [
15,
17]. In this study, a phenyl ester moiety (for achieving high transfection activity) and vitamin E scaffolds (for promoting immune activation) were combined to design ssPalmE-Phe-P4C2 for RNA vaccine development.
LNPs are generally prepared using the alcohol dilution method. In this method, the lipids and nucleic acids spontaneously form a complex due to the electrostatic interactions between the positively charged amine groups and the negatively charged nucleic acids, as well as the hydrophobic interactions between the hydrophobic moieties of lipids [
19]. However, we recently reported that the post-encapsulation of mRNA into a pre-formed LNP is also applicable for use in preparing an mRNA-LNP [
20]. Therefore, it is possible to encapsulate a desired mRNA into an mRNA-LNP by using pre-formed ready-to-use (RtoU) types of LNPs. Since the LNP(RtoU) can be used simply by rehydration and incubation, only a typical temperature-controlling device (i.e., block incubator, water bath, or thermal cycler) is needed for use. The development of an LNP(RtoU) using ssPalmE-Phe-P4C2 (LNP
ssPalmE-Phe(RtoU)) would produce a convenient form of an mRNA vaccine that offers handling practicality and vaccination efficacy. In this study, we investigated the potency of ssPalmE-Phe-P4C2 as an RNA vaccine and optimized the conditions needed for the preparation of the LNP
ssPalmE-Phe(RtoU). The ability to induce cytotoxic T lymphocyte (CTL) activation and antitumor activity against an E.G7-OVA tumor model was also investigated.
2. Materials and Methods
2.1. Animals
C57BL/6J mice (female, 6 weeks) were purchased from Japan SLC, Inc. (Shizuoka, Japan). Protocols for the animal experiments were reviewed and approved by the Chiba University Animal Care Committee following the “Guide for Care and Use of Laboratory Animals”.
2.2. Materials
A detailed list of supplier information, including the item numbers of all the reagents used in this study, is listed in Supporting Information (
Table S1). The ssPalmE-Phe-P4C2, ssPalmE-P4C2 (Product# COATSOME
® SS-EC), 1,2-dioleoyl-
sn-glycero-3-phosphatidylcholine (DOPC, Product# COATSOME
® MC-8181), and 1-(Monomethoxy polyethyleneglycol2000)2,3-dimyristoylglycerol (DMG-PEG
2000, Product# SUNBRIGHT
® GM-020) were supplied by NOF CORPORATION (Tokyo, Japan). The 1,2-dioleoyl-
sn-glycero-3-phosphoethanolamine (DOPE, Product# 18:1 (△9-Cis) PE (DOPE)) was purchased from Avanti
® Polar Lipids (Alabaster, AL, USA). Cholesterol was purchased from SIGMA Aldrich (St. Louis, MO, USA). The mRNA-encoding gene (Firefly Luciferase or Ovalbumin (OVA)) was prepared through the in vitro transcription reaction described in the following section. The Quant-IT™ RiboGreen
® RNA reagent was purchased from Thermo Fisher Scientific (Waltham, MA, USA). All other reagents and chemicals were commercially available and used without further purification.
2.3. Cell Culture
Bone-marrow-derived dendritic cell (BMDC): Murine BMDCs were isolated from the femurs of C57BL/6J mice. Bone marrow cells were cultured over 6 h in Roswell Park Memorial Institute (RPMI)-1640 medium (#R8758, SIGMA Aldrich, St. Louis, MO, USA) supplemented with 10% (
v/
v) Fetal Calf Serum (FCS, #SH30910.03, Hyclone, Logan, UT, USA), 50 µM of 2-mercaptoethanol (#21985023, Thermo Fisher Scientific, Waltham, MA, USA), 10 mM of HEPES buffer (#17557-94, nacalai tesque, Kyoto, Japan), 1 mM of sodium pyruvate (#06977-34, nacalai tesque, Kyoto, Japan), and 100 U/mL of penicillin/streptomycin (#26253-84, nacalai tesque, Kyoto, Japan). The non-adherent cells were harvested and cultured in another dish with the same medium with an additional 10 mg/mL of Recombinant Mouse GM-CSF (#415-ML-050, R&D Systems, Minneapolis, MN, USA). Non-adherent cells were further removed on Days 2 and 4. The remaining adherent cells were cultured in a fresh culture medium containing 10 mg/mL of GM-CSF. On Day 6, the non-adherent cells were used in the experiments as immature BMDCs [
16].
RAW 264.7 cell: The RAW 264.7 cells, a murine macrophage cell line, were cultured in RPMI-1640 medium (#R8758, SIGMA Aldrich, St. Louis, MO, USA) supplemented with 10% (v/v) Fetal Bovine Serum (FBS, #10270, Gibco, New York, USA) and 100 U/mL of penicillin/streptomycin (#26253-84, nacalai tesque, Kyoto, Japan). The adherent cells were peeled from the bottom of the dish with 0.25% Trypsin/EDTA (#32777-15, nacalai tesque, Kyoto, Japan) and cultured in another dish with a fresh medium. These cells were passaged every 2 days. The cells were used in the experiments after the third cell passage.
E.G7-OVA cell: E.G7-OVA cells, a murine lymphoma cell line EL4-expressing OVA, were purchased from the American Type Culture Collection (Manassas, VA, USA). E.G7-OVA cells were cultured in RPMI-1640 medium (#R8758, SIGMA Aldrich, St. Louis, MO, USA) supplemented with 10% (
v/
v) FCS (#SH30910.03, Hyclone, Logan, UT, USA), 50 µM of 2-mercaptoethanol (#21985023, Thermo Fisher Scientific, Waltham, MA, USA), 10 mM of HEPES buffer (#17557-94, nacalai tesque, Kyoto, Japan), 1 mM of sodium pyruvate (#06977-34, nacalai tesque, Kyoto Japan), 400 µg/mL of G418 Sulfate (#074-05963, FUJIFILM Wako Pure Chemical Corporation, Tokyo, Japan), and 100 U/mL of penicillin/streptomycin (#26253-84, nacalai tesque, Kyoto, Japan). The cells were collected and cultured in another dish with a fresh medium every 2 days (cell passage). The cells were used in the experiments after the third cell passage [
16].
2.4. In Vitro mRNA Transcription (IVT-mRNA)
The pcDNA3.1 vector was used as a coding template for Luciferase (Luc) and ovalbumin (OVA). The pcDNA3.1-Luc or pcDNA3.1-OVA were linearized with the restriction enzymes BspEI (#R0540S, New England Biolabs, Ipswich, MA, USA) or XhoI (#R0146S, New England Biolabs, Ipswich, MA, USA), respectively. After phenol-chloroform extraction and ethanol precipitation, the linearized pDNA was transcribed into mRNA with a mMESSAGE mMACHINE T7 ULTRA Transcription kit (#AM1345, Invitrogen, Waltham, MA, USA) following the manufacturer’s instructions. The transcribed mRNA was dissolved in THE RNA Storage Solution (#AM7000, Invitrogen, Waltham, MA, USA) and stored at −80 °C. The concentrations of linearized pDNA and transcribed mRNA were measured with a microvolume UV-Vis spectrometer (NanoDrop™ One, Thermo Fisher Scientific, Waltham, MA, USA).
2.5. Preparation of Microfluidic LNPsssPalm
The lipid composition was ssPalmE-P4C2 (or ssPalmE-Phe-P4C2)/DOPE (or DOPC)/Chol = 60/30/10 with additional DMG-PEG2000 (3 mol% of total lipid). The total lipid amount was 394.5 nmol to encapsulate 3 µg of mRNA (lipid/mRNA: 131.5 nmol/µg). The lipid mixtures were dissolved in 99.5% ethanol to a concentration of 1.32 mM. The mLuc or mOVA were diluted in 20 mM of Malic acid buffer (30 mM NaCl, pH 3.0) to a concentration of 0.0033 µg/µL. The lipid mixtures in the ethanol and mRNA solutions were set to their respective syringes in the NanoAssemblr™ Benchtop device (Precision Nanosystems, Vancouver, Canada with the settings of flow rate = 2.5 mL/min, flow rate ratio (buffer:lipid) = 3:1, and total volume = 1.5 mL. The LNPs were recovered and diluted with 20 mM of MES buffer (pH 5.5). The external solution of the LNPs was replaced with D-PBS (−) through ultrafiltration using Amicon Ultra-4-100K Centrifugal Units (#UFC810096, # UFC910096, Merck, Rahway, NJ, USA). The LNPs were then diluted to an adequate volume with D-PBS (−).
2.6. Preparation of LNPsssPalmE-Phe (RtoU)
The lipid composition was ssPalmE-Phe-P4C2/DOPE (or DOPC)/Chol = 52.5/7.5/40 with additional DMG-PEG
2000 (3 mol% of total lipid) [
17]. The amount of total lipid was 100 nmol, encapsulating 0.5 µg of mRNA (lipid/mRNA: 200 nmol/µg). The lipid mixtures were dissolved in 90% t-butanol to a concentration of 8.0 mM. The lipid mixtures in t-butanol and buffer solution, containing 40 mM MES buffer (pH 5.0, salt-free) with a 744 mg/mL solution of sucrose (in equal volume), were set to their respective syringes in the NanoAssemblr™ Benchtop device (Precision Nanosystems, Vancouver, Canada) with the settings of flow rate = 1.0 mL/min, flow rate ratio (buffer:lipid) = 3:1, and total volume = 0.52 mL. An equal volume of 320 mg/mL of sucrose solution (#30404-45, nacalai tesque, Kyoto, Japan) was added (final concentration of sucrose in the LNP solution = 320 mg/mL). The LNP solution was frozen in liquid nitrogen and transferred to the freeze-dryer (FDU-2110, EYELA, Tokyo, Japan). The drying sequence of the freeze-dryer was set to 9 segments: −40 °C (±7 °C, 40 min), −40 °C (vacuum start, 20 min), −30 °C (1 h), −20 °C (1 h), −10 °C (1 h), 0 °C (1 h), 10 °C (1 h), 20 °C (1 h), and 30 °C (3 h). The freeze-dried LNPs were recovered, and their physical appearances were inspected for visible cracks, dents, or collapses in front of a black background [
21]. The dry LNPs were reconstituted with a water solution containing 0.5 µg of either mLuc or mOVA (0.0025 µg/µL). The solution was mixed well, followed by heating at 95 °C for 5 min, then allowed to cool to room temperature (RT) for approximately 10 min before use in the experiments/analysis.
2.7. Characterization of mRNA-LNPs
The particle size, polydispersity index (PdI), and zeta-potential of the LNPs were measured via dynamic light scattering (Zetasizer Nano ZS, Malvern Panalytical, Malvern, UK). The recovery ratio and encapsulation efficiency of the mRNA were evaluated with RiboGreen® assay. Solutions containing Quant-iT™ RiboGreen® RNA reagent (#R11491, Invitrogen, Waltham, MA, USA) with or without 10% (v/v) TritonX-100 (#168-11805, FUJIFILM Wako Pure Chemical Corporation, Tokyo, Japan), in respective 1:8 ratios were prepared in D-PBS (−): Triton [+] (contained both RiboGreen® and TritonX-100) and Triton [−] (no TritonX-100, only RiboGreen®). The mRNA-LNPs, corresponding to 50 ng of mRNA in 50 µL of D-PBS (−), were prepared twice and each mixed with an equal volume of the Triton [+] and Triton [−] solution in a 96-well black microplate. A calibration curve was prepared through sequential dilution from 0 to 2000 ng/mL of mRNA. The plate was incubated for 5 min with shaking at 500 rpm in a shaking incubator. The fluorescence intensities were analyzed with a plate reader (Infinite M200 PRO, TECAN, Männedorf, Switzerland) set with emission and excitation waves of 484 nm and 535 nm, respectively. The recovery ratio was calculated from the total mRNA (quantified by Triton [+] addition) and mRNA concentration input (standard curve). The encapsulation efficiency was calculated as the ratio of the total mRNA concentration (quantified by Triton [+]) to the non-encapsulated mRNA concentration (quantified by Triton [−] addition).
2.8. Evaluation of In Vitro Gene Expression Efficiency
BMDCs (8 × 10
5 cells/mL) and RAW 264.7 cells (2 × 10
5 cells/mL) seeded in a 12-well plate were transfected with mLuc-LNPs in D-PBS (−) at a dose of 0.8 µg of mRNA. In the case of RAW 264.7 cells, the transfection was demonstrated after pre-incubation for 24 h. After transfection, the BMDCs and RAW 264.7 cells were incubated at 37 °C with 5% CO
2 for 5 h. The BMDCs and RAW 264.7 cell suspensions were collected, washed with D-PBS (−), and then solubilized with Reporter Lysis Buffer (#E397A, Promega, Madison, WI, USA). The cell lysates were added with a Nano-Glo
® Luciferase Assay System (#E1501, Promega), and the luminescence intensities were quantified with a luminometer (GloMax
® 20/20 Luminometer, Promega, Madison, WI, USA). The protein amount in the cell lysates was determined using the Bicinchoninic acid (BCA) Protein Assay kit (#T9300A, Takara Bio Inc., Kusatsu, Shiga, Japan) according to the manufacturer’s instructions. Luciferase activity was represented as a relative light unit (RLU/mg protein), calculated by dividing the luminescence intensity by the protein amount [
16].
2.9. Evaluation of In Vivo Gene Expression Efficiency (IVIS Imaging)
The mLuc-LNPs in D-PBS (−) were administered subcutaneously (back of neck) to C57BL/6J mice at a dose of 1.0 µg of mRNA under anesthetized conditions. The neck region of mice was shaved in advance. Six hours later, D-Luciferin potassium (#126-05116, FUJIFILM Wako Pure Chemical Corporation, Tokyo, Japan) in D-PBS (−) (3 mg/200 µL/mouse) was administered intraperitoneally. After 30 min had passed, the luminescence intensities were measured with an In Vivo Imaging System (IVIS® Lumina II, Caliper Life Sciences, Waltham, MA, USA).
2.10. In Vivo Cytotoxic T Lymphocyte (CTL) Assay
The in vivo CTL assay was performed as described previously [
16]. The mOVA-LNPs in D-PBS (−) were injected subcutaneously (back of neck) to C57BL/6J mice at a dose of 0.05 or 0.1 µg of mRNA under anesthetized conditions. Seven days after the LNP administration (immunization), the non-treated (NT) mice were sacrificed, and their spleens were collected into a dish containing RPMI-1640 medium (#R8758, SIGMA Aldrich, St. Louis, MO, USA) supplemented with 10% (
v/
v) FCS (#SH30910.03, Hyclone, Logan, UT, USA), 50 µM of 2-mercaptoethanol (#21985023, Thermo Fisher Scientific, Waltham, MA, USA), 10 mM of HEPES buffer (#17557-94, nacalai tesque, Kyoto, Japan), 1 mM of sodium pyruvate (#06977-34, nacalai tesque, Kyoto, Japan), and 100 U/mL of penicillin/streptomycin (#26253-84, nacalai tesque, Kyoto, Japan). The cell suspension of the splenocytes was filtered through a 40 µm cell strainer and resuspended in Red Blood Cell Lysing Buffer (#R7757-100ML, SIGMA Aldrich, St. Louis, MO, USA). The cell suspension was washed and resuspended in a fresh medium, then equally divided into two suspensions: CFSE
high- and CFSE
low-labelled. Each cell suspension was resuspended in a 1.0 × 10
7 cells/mL cell concentration. The OVA H-2K
b cytotoxic T-lymphocyte epitope peptide (SIINFEKL, OVA
257–264) in DMSO was added to the CFSE
high-labelled cells (1/400 of the suspension volume). Both cell suspensions were incubated at 37 °C with 5% CO
2 for 1 h. Each cell suspension was adjusted to a 3.0 × 10
7 cells/mL cell concentration in D-PBS (−). The CFSE
high (5.0 µM) and CFSE
low (0.5 µM) (Cellstain CFSE, #C375, Dojindo Laboratories, Kumamoto, Japan) cells were added to their respective cell suspension tubes and incubated in a 37 °C water bath for 10 min under a light-shielding condition. Each cell suspension was washed repeatedly with a fresh medium and D-PBS (−), then resuspended in 5.0 × 10
7 cells/mL cell concentration in D-PBS (−). The cells were administered intravenously into the immunized mice with equal volumes (ratio 1:1) of CFSE
high- and CFSE
low-labelled splenocytes. Twenty hours after administration, the spleens were collected from the immunized mice, and the splenocytes were suspended into single-cell suspensions in FACS buffer (0.5% Bovine Serum Albumin (#01860-07, nacalai tesque, Kyoto, Japan) and 0.1% NaN
3 (#194-01275, FUJIFILM Wako Pure Chemical Corporation, Tokyo, Japan) in D-PBS (−)). The number of CFSE-labeled cells (CFSE
high and CFSE
low) was quantified with a flow cytometer (NovoCyte Flow Cytometer, Agilent, Santa Clara, CA, USA). The CTL activity was represented by the degree of cell lysis, calculated as the ratio of the number of CFSE
high-labelled cells to CFSE
low-labelled cells.
2.11. Therapeutic Anti-Tumor Response against E.G7-OVA
E.G7-OVA cells (8.0 × 10
7 cells/40 µL) suspended in D-PBS (−) were inoculated subcutaneously on the left flank of mice in anesthetized conditions. After the tumor grew to ≥100 mm
3 (approximately after 7–9 days), the mice were injected subcutaneously (back of neck) with mOVA-LNPs (0.5 µg of mRNA) in D-PBS (−). Tumor sizes were measured at 3-day intervals with the calculation formula: {long axis} × {short axis}
2 × 0.52 [
16]. The endpoint of tumor measurement was set to 1000 mm
3.
4. Discussion
The ionizable lipid ssPalmE-Phe-P4C2 was designed so as to combine two important functions for an RNA vaccine: a vitamin E scaffold for immune activation and a phenyl ester for self-degradability, respectively. Regarding Vitamin E, it has been used as a key component in AS03, a squalene-based oil-in-water emulsion adjuvant that is used in vaccines against the avian influenza virus [
23,
24,
25]. It has been reported that the vitamin E in the AS03-adjuvanted vaccine modulates the production of cytokines and chemokines such as CCL2, CCL3, IL-6, CSF3, and CXCL1 and promotes antigen-loading monocytes as well as the recruitment of granulocytes [
26]. The ssPalm with vitamin E scaffolds (ssPalmE) was initially developed to deliver nucleic acids to the liver; the delivery of siRNA by LNPs
ssPalmE demonstrated gene knockdown efficiency and extensive liver accumulation compared to other ssPalms with myristic acid or retinoic acid [
15]. The molecular tuning of the ssPalmE through attaching tertiary amines to a piperizine ring resulted in the formation of the second-generation ssPalmE-P4C2, which demonstrated an efficient endosomal escape activity, which contributed to the improved gene knockdown efficiency [
15,
17]. However, LNPs prepared with ssPalmE derivatives (with piperazine as the tertiary amine structure) triggered severe inflammatory responses that involved the production of pro-inflammatory cytokines (IL6, TNFα), the intracellular DNA-sensor-related cytokine (IL-1β), and interferon (IFNβ) when they were combined with plasmid DNA. From these observations, we hypothesized that the LNPs formed with the ssPalmE derivative would be applicable for use as an RNA vaccine [
16]. It was revealed that the mRNA LNPs
ssPalmE acted as both a nucleic acid (mRNA) carrier and as an immune adjuvant by inducing the proliferation of antigen-specific CD8+ T cells and their differentiation into effector and memory cells to activate cellular immunity [
18]. However, the mechanism concerning the ssPalmE-triggered immune activation is not clear.
Studies have demonstrated the capability of vaccines and an adjuvant to induce immunogenicity by stimulating the damage-associated molecular patterns (DAMPs) pathway [
27,
28]. Cellular stressors or cell death signaling can trigger the release of DAMPs that are then recognized by other cells via pattern recognition receptors (PRRs). This interaction will result in immunogenic reactions, such as the upregulation of the chemokine/cytokines involved in immune responses. Renowned vaccine adjuvants such as Alum, AS03, and HP-β-cyclodextrin are known to induce the release of DAMP signals, which stimulates and enhances the immunogenicity of vaccines [
27]. Our previous study also reported the detection of dsDNA at the injection site (skin) upon LNP
ssPalmE administration. This finding suggests that the adjuvant effect of LNPs
ssPalmE-Phe(RtoU) could also be dependent on this DAMP pathway [
18]. Further investigation is required into the cells and molecules involved in this response. It is noteworthy that the use of a simple combination of α-tocopherol and α-tocopherol succinates as a component of poorly immune-stimulative LNPs (formed with a myristic acid-scaffold ssPalm) did not enhance cytokine production [
18]. The covalent linkage of α-tocopherol and a tertiary amine into a single molecular structure and/or the dimerization of α-tocopherol could be key factors in the adjuvant effect. Thus, it is plausible that the action of the LNPs
ssPalmE involves a mechanism that differs from that of AS03.
The other molecular design integrated in ssPalmE-P4C2 is the introduction of a phenyl ester. The degradability of ionizable lipids in the body has been investigated as a strategy for reducing adverse side effects caused by the accumulation of lipid-like materials. The introduction of ester bonds and/or disulfide bonds is one of the common strategies for achieving the biodegradation of an LNP [
15]. It was reported that the combination of disulfide bonds and a phenyl ester moiety synergistically enhances the transfection activity of mRNA by inducing an intraparticle self-degradation that occurs within the limited intraparticle spaces. This reaction induces the release of the nucleic acid cargo [
17]. Actually, an siRNA-LNP that contains an oleic-acid-based self-degradable ssPalm (ssPalmO-Phe-P4C2) showed an ED
50 of 0.0044 mg/kg siRNA in the liver when administered via an i.v. injection [
15]. Comparing the knockdown activity of ssPalmE-P4C2 with that of ssPalmO-Phe-P4C2, the findings suggested that the efficiencies of nucleic acid delivery for these two materials were significantly different. From this finding, we concluded that the combination of vitamin E scaffolds and phenyl esters is a strategy that can satisfy both immunostimulatory properties and nucleic acid delivery efficiency. The improvement in the luciferase activity and CTL activity of LNP
ssPalmE-Phe over LNP
ssPalmE suggested that the release of mRNA promoted by self-degradability is an important factor for improving the efficacy of an RNA vaccine. The LNP
ssPalmE-Phe(RtoU) also demonstrated strong CTL activity and a significant suppression of E.G7-OVA tumor growth (
Figure 7). These observations suggest that ssPalmE-Phe-P4C2 would be a reasonable candidate for use in designing an RNA vaccine that focuses on the activation of cellular immunity.
It is noteworthy that the mRNAs used in this study were unmodified/unpurified. However, in the case of ionizable lipids with vitamin E scaffolds, the hydrophobic scaffold contributed significantly to the adjuvant activity of the LNPs [
18]. The contribution of mRNA to the adjuvant activity was negligible. Therefore, ssPalmE-Phe-P4C2, rather than the IVT-mRNA, is considered to be responsible for the immunostimulatory properties of the mRNA-LNPs [
18]. Since the improvement in the quality of mRNA can positively affect the vaccine activity of an RNA vaccine [
29], the combination of chemically modified and purified mRNA with LNPs
ssPalmE-Phe(RtoU) would be a potent strategy for the further development of cancer vaccines.
The ready-to-use (RtoU) formulation, a freeze-drying-based preparation method of mRNA-LNPs, was then applied to the LNPs
ssPalmE-Phe to further develop them as a convenient nucleic acid carrier. Freeze-drying has been extensively used in pharmaceutical industries to improve the stability and shelf life of drug products [
19,
30,
31]. However, the stresses associated with freezing and drying can damage biomolecules unless an appropriate cryoprotectant and lipid composition are used [
32]. In this study, the successful application of an RtoU formulation produced mRNA-LNPs that allow for the post-encapsulation of the desired nucleic acids to be achieved via simple rehydration and incubation. The LNP
ssPalmE-Phe(RtoU) offers handling practicality and a high rate of encapsulation (>90%) with a minimum loss of nucleic acids. The limitation of the LNP
ssPalmE-Phe(RtoU) formulation involves the concentration (1.0 µg mRNA/200 µL) and scale (1.0 µg mRNA/vial) of the resultant mRNA-LNPs. Although this is sufficient for the vaccine experiment in mice, this point should be improved from the viewpoint of other applications.
Sucrose acts as a cryoprotectant, which helps to maintain the stability and integrity of LNPs during the freeze-drying process. The Pfizer/BioNTech and Moderna COVID-19 mRNA vaccines both used sucrose as a cryoprotectant to protect and stabilize the LNPs while in a deep-frozen state (−80 °C) during distribution [
33,
34,
35,
36]. The Food and Drug Administration authorized the most recent formula of the Pfizer/BioNTech and Moderna COVID-19 mRNA vaccines (2023–2024 Formula), which corresponds to the Omicron variant XBB.1.5 of SARS-CoV-2. In this recent formula, the Pfizer/BioNTech COVID-19 mRNA vaccine used 31 mg of sucrose in each 0.3 mL dose (103.3 mg/mL) [
37]; meanwhile, the Moderna COVID-19 mRNA vaccine used 21.8 mg of sucrose in each 0.25 mL dose (87.2 mg/mL) [
38]. Compared to a frozen vaccine product, such as the COVID-19 mRNA vaccines, a much higher sucrose concentration is needed to stabilize a lyophilized vaccine product. It was mentioned that sugar molecules replace the water between the hydrophilic heads of phospholipids during lyophilization, thus lowering the phase transition temperature and preventing a gel-to-liquid phase transition [
31]. It was also suggested that sugar forms glass matrixes, trapping phospholipids upon the removal of water, which prevents lipid aggregation and damage by ice crystals [
31,
39]. In this study, sucrose was used as a cryoprotectant for the LNPs
ssPalmE-Phe(RtoU) following a previously reported procedure [
20,
40]. The dent or shrinkage appearance of the LNPs
ssPalmE-Phe(RtoU) is probably related to the formulation process, in which the secondary drying process was apparently too rapid [
22]. The amount of sucrose and/or the freeze-drying process should be optimized for each application. In our experiments, the dent appearance of the lyophilized product had no effect on the product’s quality. This is consistent with the general notion that a dent appearance is acceptable [
22]. The pH of the buffer is another important factor in preparing an LNP, especially for the post-encapsulation of mRNA. The electrostatic interaction between ionizable lipids and nucleic acids is probably the main driving force in the post-encapsulation process [
20]. It should be noted here that the buffer pH (5.0) together with a high concentration of sucrose (320 mg/mL) produced preferable particle properties (
Table 2) and a high immune activation activity (
Figure 5).
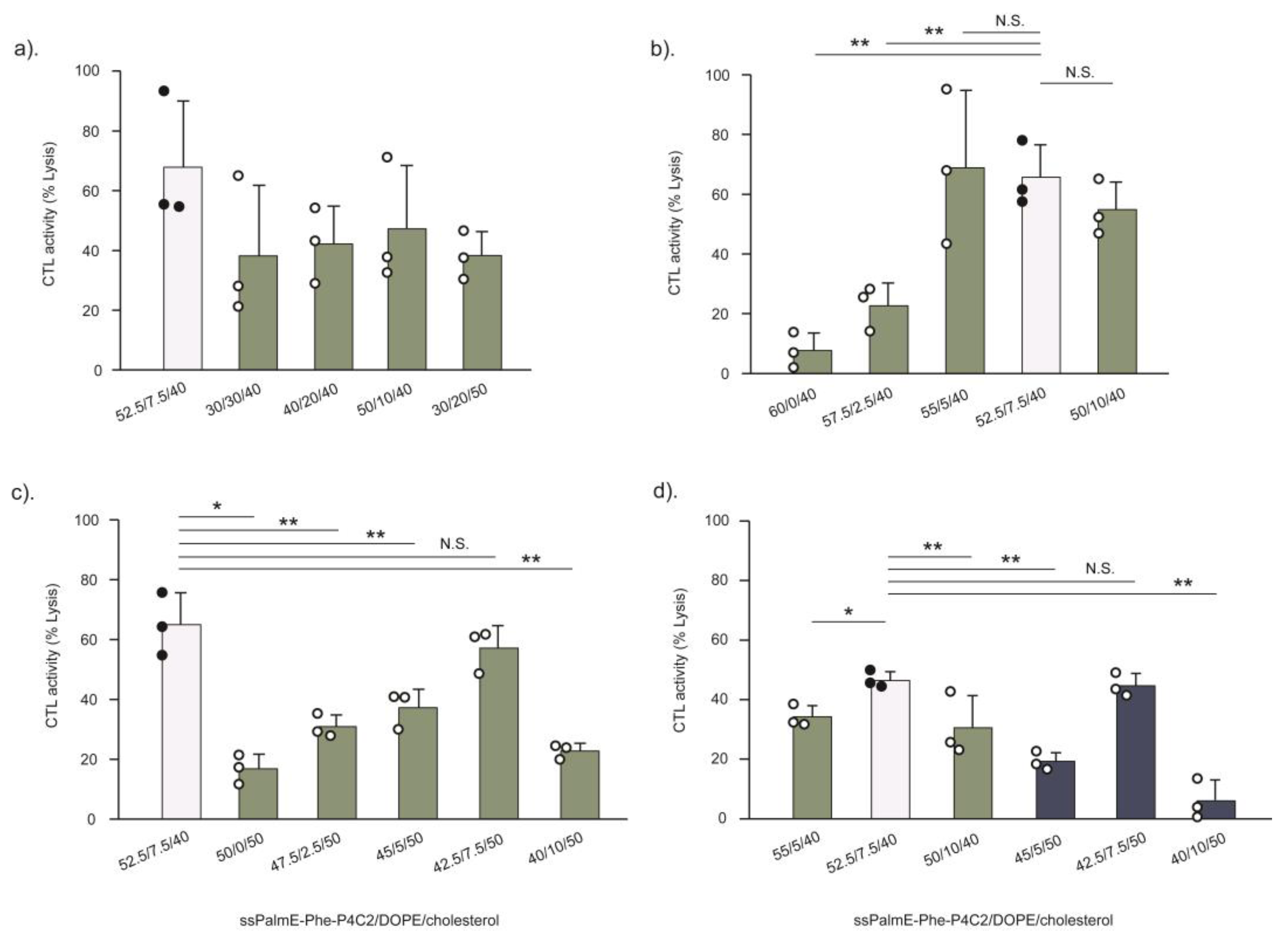
The established RtoU formulation (ssPalm/helper lipid/cholesterol = 52.5/7.5/40) [
20] was found to be the most suitable lipid composition for the LNPs
ssPalmE-Phe, which demonstrated stronger immune activation activity compared to the other formulations (
Figure 6). This observation is closely related to the cholesterol composition. Cholesterol is known to have the ability to provide cryoprotection for LNPs during freeze-drying by maintaining the stability of lipid formation and preventing the risk of leakage of contents [
41,
42]. In addition, it was revealed that a cholesterol content of at least 40% in the LNPs
ssPalm was essential for achieving a high transfection activity of the phenyl-type ionizable lipid [
17]. The ONPATTRO
® or Patisiran (Alnylam Pharmaceutical, Cambridge, MA, USA) also utilizes a cholesterol content > 30% (38.5%) in its LNP formulation for siRNA delivery [
33]. Moreover, the current COVID-19 mRNA vaccines by Pfizer/BioNTech and Moderna also utilize a cholesterol content >30% in their LNP formulation, with 42.7% and 38.5% cholesterol, respectively [
33]. Although the exact composition of cholesterol varies depending on the type of ionizable lipid and its delivery purpose, a cholesterol content of at least around 40% in the LNP formulation might be important in terms of exerting its mRNA delivery efficiency. The limitation of the LNP(RtoU) formulation is the concentration (1.0 µg/200 µL) and scale (1.0 µ/vial) of the resultant mRNA-LNPs. Although this is enough for the vaccine experiment in mice, this point should be improved from the viewpoint of other applications.
Phospholipids are also included as helper lipids to provide particle stability and delivery efficiency [
43]. DOPE is a cone-shaped lipid with high fusogenic activity, while DOPC functions to stabilize the LNPs through its overall cylindrical shape. Both lipids were used to form the microfluidic-mixer-type (
Figure 3) or the RtoU-type (
Figure 4) LNPs
ssPalmE-Phe. As a result, the use of DOPE was revealed to be important for achieving higher activity of cytotoxic T cells. The findings indicated that the CTL activity of the LNPs
ssPalmE-Phe(RtoU) tended to increase when the DOPE ratio was reduced. However, the complete removal of the helper lipid was shown to be detrimental to the CTL activity of the LNPs
ssPalmE-Phe(RtoU). This indicates there is a delicate balance between the ionizable lipid, the helper lipid, and the amount of cholesterol needed to achieve an optimum lipid composition. When the cholesterol ratio was fixed, the DOPE ratio was accompanied by an increase in the ssPalmE-Phe-P4C2 ratio. Thus, the amount of ionizable lipid holds priority in the lipid composition, which determines the immune activation activity of the LNPs
ssPalmE-Phe(RtoU).


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}