
Synergistic Activity and Mechanism of Sanguinarine with Polymyxin B against Gram-Negative Bacterial Infections
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. Bacterial Strains and Growth Conditions
2.3. Minimal Inhibitory Concentration (MIC) Determination
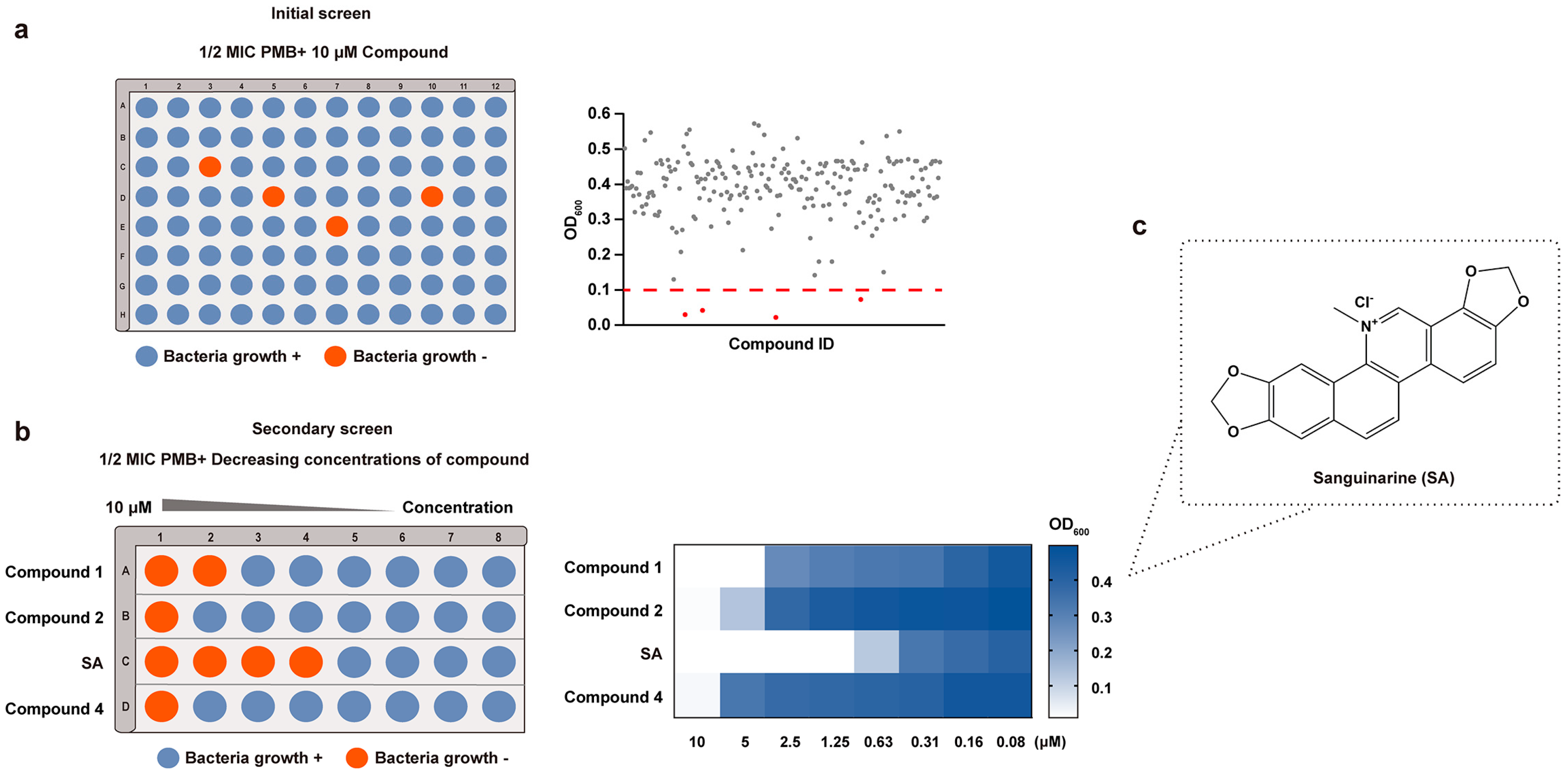
2.4. Natural Product Library Screening
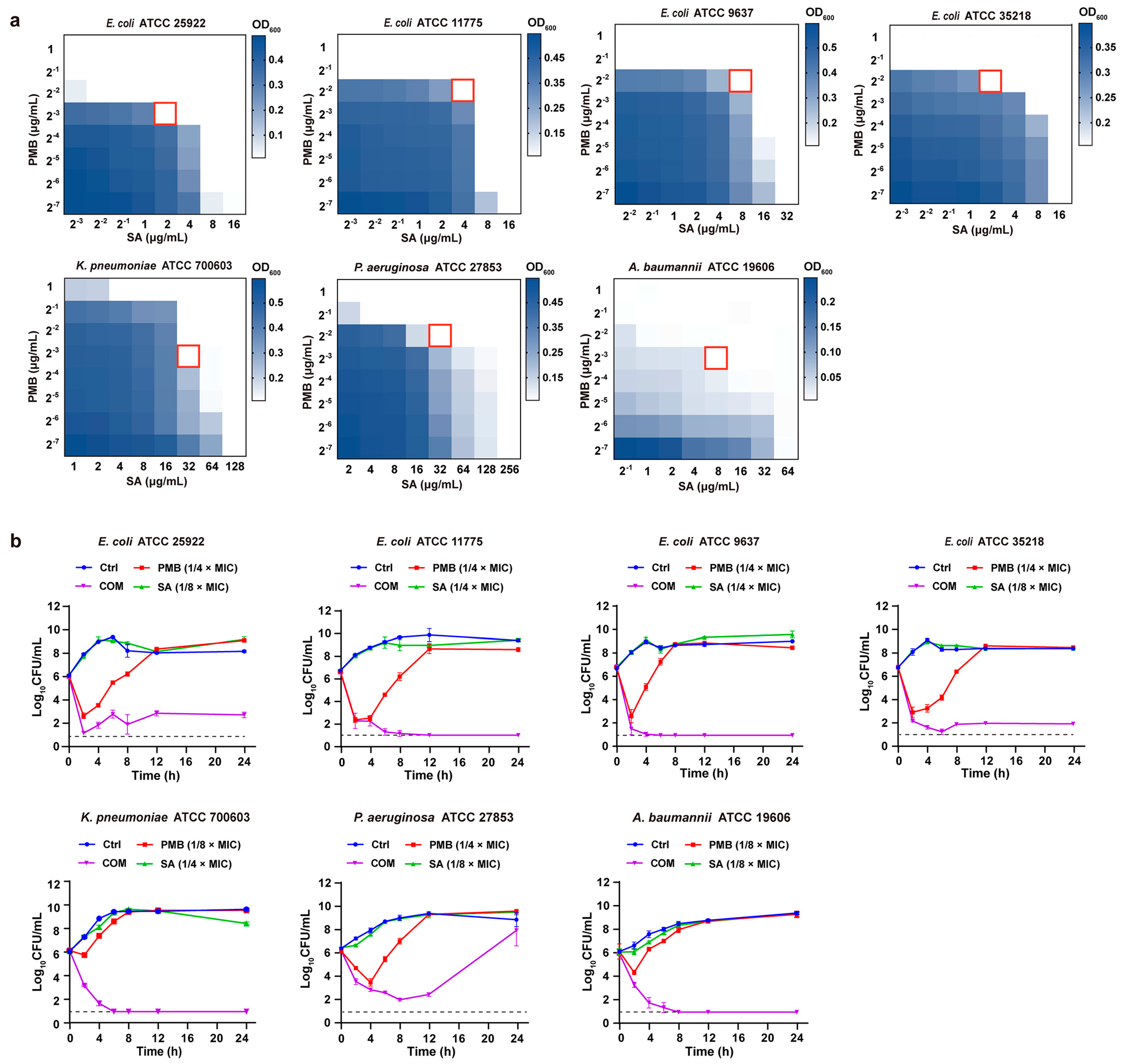
2.5. Checkerboard Assay
2.6. Time–Kill Curves
2.7. E. coli-Induced Peritonitis Model
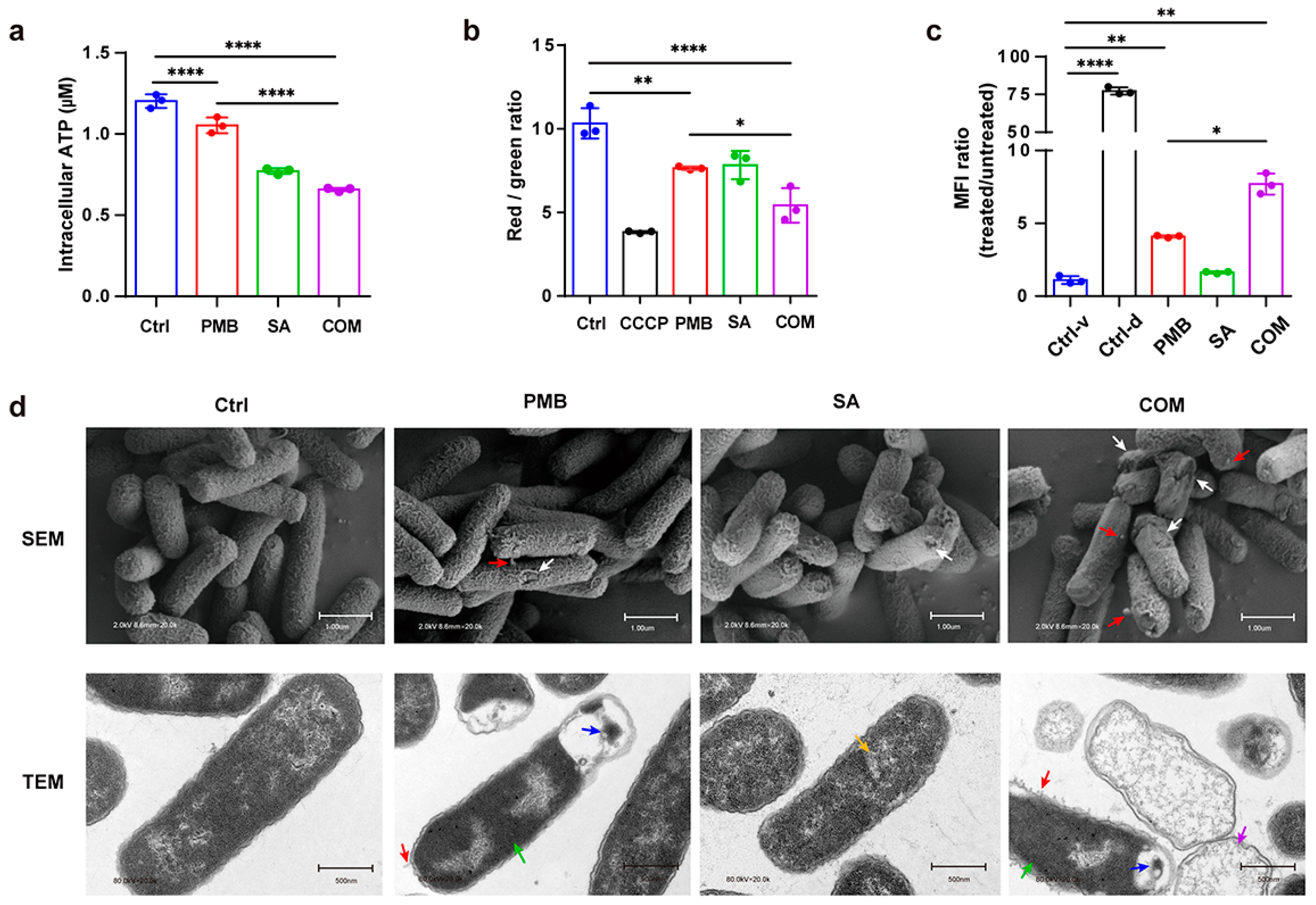
2.8. Intracellular ATP Level Detection
2.9. Determination of Membrane Potential
2.10. Determination of Membrane Integrity
2.11. Electron Microscopy
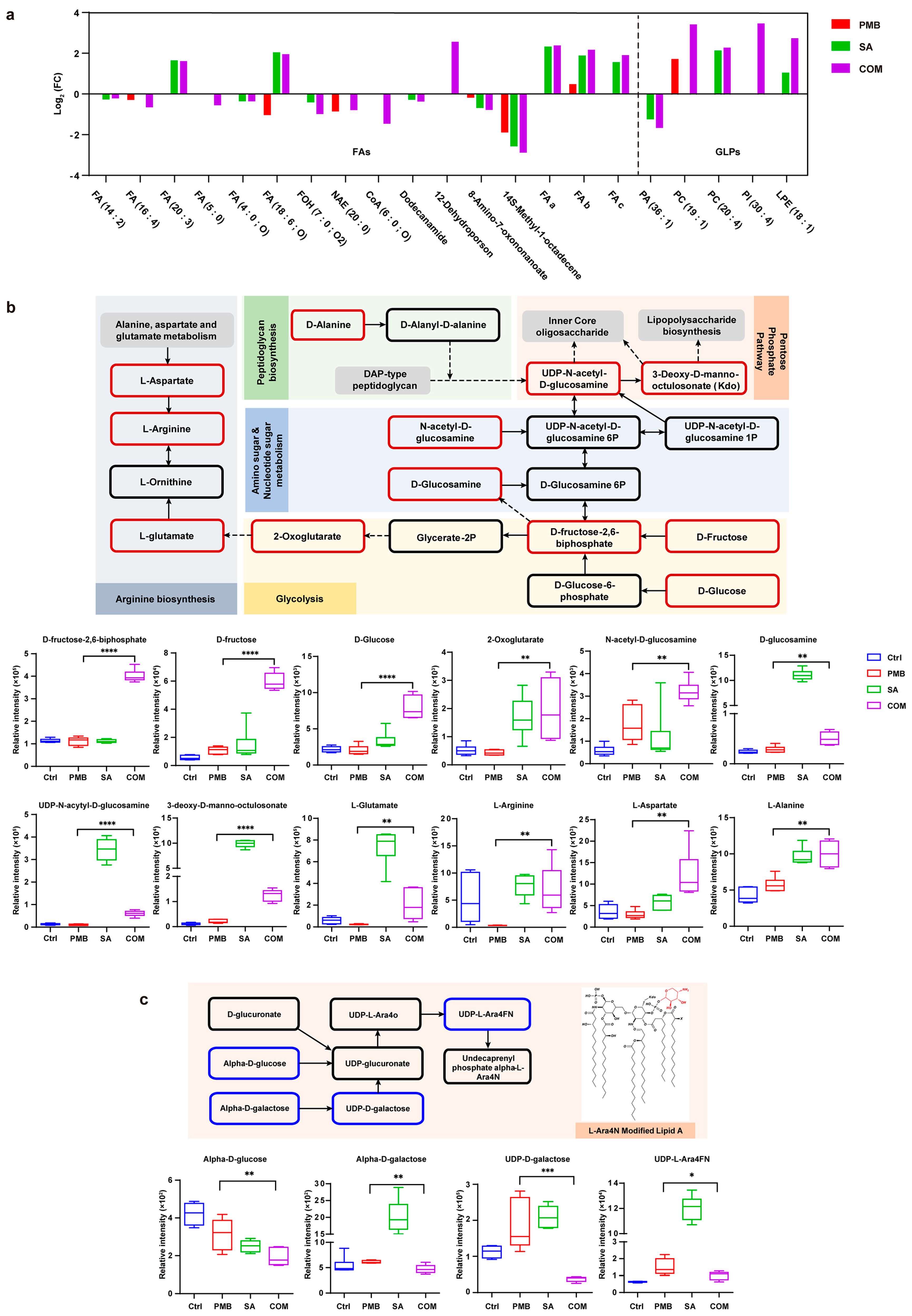
2.12. Metabolomic Analysis
2.13. Statistical Analysis
3. Results
3.1. Screening for Compounds Potentiating the Activity of PMB against E. coli
3.2. In Vitro and In Vivo Antibacterial Activity Assays
3.2.1. In Vitro Antibacterial Activity of PMB-SA Combinations
3.2.2. In Vivo Antibacterial Activity of PMB-SA Combination in E. coli-Induced Peritonitis Model
3.3. Exploring the Synergic Mechanism of SA with PMB
3.3.1. Determination of Intracellular ATP
3.3.2. Fluctuation of Bacterial Membrane Potential
3.3.3. Changes in Bacterial Membrane Permeability
3.3.4. Observation of Cell Morphology Using Electron Microscopy
3.3.5. Metabolomic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| GNB | Gram-negative bacteria |
| PMB | Polymyxin B |
| SA | Sanguinarine |
| ICU | Intensive care unit |
| WHO | World Health Organization |
| OM | Outer membrane |
| LPS | Lipopolysaccharides |
| MDR | Multidrug-resistant |
| RSDs | Relative standard deviations |
| PCA | Principal component analysis |
| FAs | Fatty acids |
| GLPs | Glycerophospholipids |
| MRSA | Methicillin-resistant staphylococcus aureus |
| ELISA | Enzyme-linked immunosorbent assay |
| E. coli | Escherichia coli |
| K. pneumoniae | Klebsiella Pneumoniae |
| P. aeruginosa | Pseudomonas aeruginosa |
| A. baumannii | Acinetobacter baumannii |
| MIC | Minimal inhibitory concentration |
| CLSI | Clinical and Laboratory Standards Institute |
| FICI | Fractional inhibitory concentration index |
| RT-qPCR | Quantitative reverse transcriptase PCR |
| SEM | Scanning electron microscopy |
| TEM | Transmission electron microscopy |
| ESI | Electrospray ionization |
References
- Mushtaq, A.; Kazi, F. New guidelines to treat bacterial infections. Lancet Infect. Dis. 2020, 20, 1239. [Google Scholar] [CrossRef] [PubMed]
- Paul, M.; Carrara, E.; Retamar, P.; Tängdén, T.; Bitterman, R.; Bonomo, R.A.; de Waele, J.; Daikos, G.L.; Akova, M.; Harbarth, S.; et al. European Society of Clinical Microbiology and Infectious Diseases (ESCMID) guidelines for the treatment of infections caused by multidrug-resistant Gram-negative bacilli (endorsed by European society of intensive care medicine). Clin. Microbiol. Infect. 2022, 28, 521–547. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, R.; Xiao, X.; Wang, Z. Antibiotic adjuvants: An alternative approach to overcome multi-drug resistant Gram-negative bacteria. Crit. Rev. Microbiol. 2019, 45, 301–314. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H. Preventing drug access to targets: Cell surface permeability barriers and active efflux in bacteria. Semin. Cell Dev. Biol. 2001, 12, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Simpson, B.W.; Trent, M.S. Pushing the envelope: LPS modifications and their consequences. Nat. Rev. Microbiol. 2019, 17, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Z.; Plésiat, P.; Nikaido, H. The challenge of efflux-mediated antibiotic resistance in Gram-negative bacteria. Clin. Microbiol. Rev. 2015, 28, 337–418. [Google Scholar] [CrossRef]
- Doi, Y. Treatment Options for Carbapenem-resistant Gram-negative Bacterial Infections. Clin. Infect. Dis. 2019, 69, s565–s575. [Google Scholar] [CrossRef]
- Voulgaris, G.L.; Voulgari, M.L.; Falagas, M.E. Developments on antibiotics for multidrug resistant bacterial Gram-negative infections. Expert Rev. Anti Infect. Ther. 2019, 17, 387–401. [Google Scholar] [CrossRef]
- Lin, Y.W.; Han, M.L.; Zhao, J.; Zhu, Y.; Rao, G.; Forrest, A.; Song, J.; Kaye, K.S.; Hertzog, P.; Purcell, A.; et al. Synergistic Combination of Polymyxin B and Enrofloxacin Induced Metabolic Perturbations in Extensive Drug-Resistant Pseudomonas aeruginosa. Front. Pharmacol. 2019, 10, 1146. [Google Scholar] [CrossRef]
- Nang, S.C.; Azad, M.A.K.; Velkov, T.; Zhou, Q.T.; Li, J. Rescuing the Last-Line Polymyxins: Achievements and Challenges. Pharmacol. Rev. 2021, 73, 679–728. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Lu, J.; Han, M.L.; Jiang, X.; Azad, M.A.K.; Patil, N.A.; Lin, Y.W.; Zhao, J.; Hu, Y.; Yu, H.H.; et al. Polymyxins Bind to the Cell Surface of Unculturable Acinetobacter baumannii and Cause Unique Dependent Resistance. Adv. Sci. 2020, 7, 2000704. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Shen, Y.; Walsh, T.R.; Wang, Y.; Hu, F. Use of polymyxins in Chinese hospitals. Lancet Infect. Dis. 2020, 20, 1125–1126. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, B.T.; Pogue, J.M.; Zavascki, A.P.; Paul, M.; Daikos, G.L.; Forrest, A.; Giacobbe, D.R.; Viscoli, C.; Giamarellou, H.; Karaiskos, I.; et al. International Consensus Guidelines for the Optimal Use of the Polymyxins: Endorsed by the American College of Clinical Pharmacy (ACCP), European Society of Clinical Microbiology and Infectious Diseases (ESCMID), Infectious Diseases Society of America (IDSA), International Society for Anti-infective Pharmacology (ISAP), Society of Critical Care Medicine (SCCM), and Society of Infectious Diseases Pharmacists (SIDP). Pharmacotherapy 2019, 39, 10–39. [Google Scholar] [PubMed]
- Tian, Y.; Zhang, Q.; Wen, L.; Chen, J. Combined effect of Polymyxin B and Tigecycline to overcome Heteroresistance in Carbapenem-Resistant Klebsiella pneumoniae. Microbiol. Spectr. 2021, 9, e0015221. [Google Scholar] [CrossRef] [PubMed]
- Lenhard, J.R.; Gall, J.S.; Bulitta, J.B.; Thamlikitkul, V.; Landersdorfer, C.B.; Forrest, A.; Nation, R.L.; Li, J.; Tsuji, B.T. Comparative pharmacodynamics of four different carbapenems in combination with polymyxin B against carbapenem-resistant Acinetobacter baumannii. Int. J. Antimicrob. Agents 2016, 48, 719–724. [Google Scholar] [CrossRef] [PubMed]
- Rosso-Fernández, C.; Garnacho-Montero, J.; Antonelli, M.; Dimopoulos, G.; Cisneros, J.M. Safety and efficacy of colistin versus meropenem in the empirical treatment of ventilator-associated pneumonia as part of a macro-project funded by the Seventh Framework Program of the European Commission studying off-patent antibiotics: Study protocol for a randomized controlled trial. Trials 2015, 16, 102. [Google Scholar]
- Qu, J.; Qi, T.T.; Qu, Q.; Long, W.M.; Chen, Y.; Luo, Y.; Wang, Y. Polymyxin B-Based Regimens for Patients Infected with Carbapenem-Resistant Gram-Negative Bacteria: Clinical and Microbiological Efficacy, Mortality, and Safety. Infect. Drug Resist. 2022, 15, 1205–1218. [Google Scholar] [CrossRef]
- John, J.F.; Falci, D.R.; Rigatto, M.H.; Oliveira, R.D.; Kremer, T.G.; Zavascki, A.P. Severe Infusion-Related Adverse Events and Renal Failure in Patients Receiving High-Dose Intravenous Polymyxin B. Antimicrob. Agents Chemother. 2018, 62, e01617-17. [Google Scholar] [CrossRef]
- Beuria, T.K.; Santra, M.K.; Panda, D. Sanguinarine blocks cytokinesis in bacteria by inhibiting FtsZ assembly and bundling. Biochemistry 2005, 44, 16584–16593. [Google Scholar] [CrossRef]
- Falchi, F.A.; Borlotti, G.; Ferretti, F.; Pellegrino, G.; Raneri, M.; Schiavoni, M.; Caselli, A.; Briani, F. Sanguinarine Inhibits the 2-Ketogluconate Pathway of Glucose Utilization in Pseudomonas aeruginosa. Front. Microbiol. 2021, 12, 744458. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Zhang, N.; Kou, S.; Gao, L.; Peng, B.; Dai, Y.; Zheng, J. Sanguinarine synergistically potentiates aminoglycoside-mediated bacterial killing. Microb. Biotechnol. 2022, 15, 2055–2070. [Google Scholar] [CrossRef] [PubMed]
- Humphries, R.; Bobenchik, A.M.; Hindler, J.A.; Schuetz, A.N. Overview of Changes to the Clinical and Laboratory Standards Institute Performance Standards for Antimicrobial Susceptibility Testing, M100, 31st Edition. J. Clin. Microbiol. 2021, 59, e0021321. [Google Scholar] [CrossRef] [PubMed]
- Odds, F.C. Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 2003, 52, 1. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Hudson, M.A.; Siegele, D.A.; Lockless, S.W. Use of a Fluorescence-Based Assay To Measure Escherichia coli Membrane Potential Changes in High Throughput. Antimicrob. Agents Chemother. 2020, 64, e00910–e00920. [Google Scholar] [CrossRef]
- Kerstens, M.; Boulet, G.; Tritsmans, C.; Horemans, T.; Hellings, M.; Delputte, P.; Maes, L.; Cos, P. Flow cytometric enumeration of bacteria using TO-PRO-3 iodide as a single-stain viability dye. J. Lab. Autom. 2014, 19, 555–561. [Google Scholar] [CrossRef]
- Pan, Y.; Li, J.; Xia, X.; Wang, J.; Jiang, Q.; Yang, J.; Dou, H.; Liang, H.; Li, K.; Hou, Y. β-glucan-coupled superparamagnetic iron oxide nanoparticles induce trained immunity to protect mice against sepsis. Theranostics 2022, 12, 675–688. [Google Scholar] [CrossRef]
- Ueda, T.; Fukunaga, K.; Seki, H.; Miyata, J.; Arita, M.; Miyasho, T.; Obata, T.; Asano, K.; Betsuyaku, T.; Takeda, J. Combination therapy of 15-epi-lipoxin A4 with antibiotics protects mice from Escherichia coli-induced sepsis. Crit. Care Med. 2014, 42, e288–e295. [Google Scholar] [CrossRef]
- Michels, K.R.; Lambrecht, N.J.; Carson, W.F.; Schaller, M.A.; Lukacs, N.W.; Bermick, J.R. The Role of Iron in the Susceptibility of Neonatal Mice to Escherichia coli K1 Sepsis. J. Infect. Dis. 2019, 220, 1219–1229. [Google Scholar] [CrossRef]
- Zhao, J.; Han, M.L.; Zhu, Y.; Lin, Y.W.; Wang, Y.W.; Lu, J.; Hu, Y.; Tony Zhou, Q.; Velkov, T.; Li, J. Comparative metabolomics reveals key pathways associated with the synergistic activity of polymyxin B and rifampicin combination against multidrug-resistant Acinetobacter baumannii. Biochem. Pharmacol. 2021, 184, 114400. [Google Scholar] [CrossRef] [PubMed]
- Olaitan, A.O.; Morand, S.; Rolain, J.M. Mechanisms of polymyxin resistance: Acquired and intrinsic resistance in bacteria. Front. Microbiol. 2014, 5, 643. [Google Scholar] [CrossRef] [PubMed]
- Baron, S.; Hadjadj, L.; Rolain, J.M.; Olaitan, A.O. Molecular mechanisms of polymyxin resistance: Knowns and unknowns. Int. J. Antimicrob. Agents 2016, 48, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Du, G.F.; Zheng, Y.D.; Chen, J.; He, Q.Y.; Sun, X. Novel Mechanistic Insights into Bacterial Fluoroquinolone Resistance. J. Proteome Res. 2019, 18, 3955–3966. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Huang, Z.; Ruan, B.; Wang, H.; Chen, M.; Rehman, S.; Wu, P. Quantitative proteomic analysis reveals the mechanisms of polymyxin B toxicity to Escherichia coli. Chemosphere 2020, 259, 127449. [Google Scholar] [CrossRef] [PubMed]
- Göpel, Y.; Görke, B. Interaction of lipoprotein QseG with sensor kinase QseE in the periplasm controls the phosphorylation state of the two-component system QseE/QseF in Escherichia coli. PLoS Genet. 2018, 14, e1007547. [Google Scholar] [CrossRef] [PubMed]
- Hussein, M.; Schneider-Futschik, E.K.; Paulin, O.K.A.; Allobawi, R.; Crawford, S.; Zhou, Q.T.; Hanif, A.; Baker, M.; Zhu, Y.; Li, J.; et al. Effective Strategy Targeting Polymyxin-Resistant Gram-Negative Pathogens: Polymyxin B in Combination with the Selective Serotonin Reuptake Inhibitor Sertraline. ACS Infect. Dis. 2020, 6, 1436–1450. [Google Scholar] [CrossRef]
- Abdul Rahim, N.; Zhu, Y.; Cheah, S.E.; Johnson, M.D.; Yu, H.H.; Sidjabat, H.E.; Butler, M.S.; Cooper, M.A.; Fu, J.; Paterson, D.L.; et al. Synergy of the Polymyxin-Chloramphenicol Combination against New Delhi Metallo-β-Lactamase-Producing Klebsiella pneumoniae Is Predominately Driven by Chloramphenicol. ACS Infect. Dis. 2021, 7, 1584–1595. [Google Scholar] [CrossRef]
- Pallan, P.S.; Kreutz, C.; Bosio, S.; Micura, R.; Egli, M. Effects of N2,N2-dimethylguanosine on RNA structure and stability: Crystal structure of an RNA duplex with tandem m2 2G: A pairs. RNA 2008, 14, 2125–2135. [Google Scholar] [CrossRef]
- Barth, N.; Ribeiro, V.B.; Zavascki, A.P. In vitro activity of polymyxin B plus imipenem, meropenem, or tigecycline against KPC-2-producing Enterobacteriaceae with high MICs for these antimicrobials. Antimicrob. Agents Chemother. 2015, 59, 3596–3597. [Google Scholar] [CrossRef]
- Vestergaard, M.; Paulander, W.; Marvig, R.L.; Clasen, J.; Jochumsen, N.; Molin, S.; Jelsbak, L.; Ingmer, H.; Folkesson, A. Antibiotic combination therapy can select for broad-spectrum multidrug resistance in Pseudomonas aeruginosa. Int. J. Antimicrob. Agents 2016, 47, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Koh Jing Jie, A.; Hussein, M.; Rao, G.G.; Li, J.; Velkov, T. Drug Repurposing Approaches towards Defeating Multidrug-Resistant Gram-Negative Pathogens: Novel Polymyxin/Non-Antibiotic Combinations. Pathogens 2022, 11, 1420. [Google Scholar] [CrossRef] [PubMed]
- Otto, R.G.; van Gorp, E.; Kloezen, W.; Meletiadis, J.; van den Berg, S.; Mouton, J.W. An alternative strategy for combination therapy: Interactions between polymyxin B and non-antibiotics. Int. J. Antimicrob. Agents 2019, 53, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Betts, J.W.; Sharili, A.S.; La Ragione, R.M.; Wareham, D.W. In Vitro Antibacterial Activity of Curcumin-Polymyxin B Combinations against Multidrug-Resistant Bacteria Associated with Traumatic Wound Infections. J. Nat. Prod. 2016, 79, 1702–1706. [Google Scholar] [CrossRef] [PubMed]
- Yi, K.; Liu, S.; Liu, P.; Luo, X.; Zhao, J.; Yan, F.; Pan, Y.; Liu, J.; Zhai, Y.; Hu, G. Synergistic antibacterial activity of tetrandrine combined with colistin against MCR-mediated colistin-resistant Salmonella. Biomed. Pharmacother. 2022, 149, 112873. [Google Scholar] [CrossRef] [PubMed]
- Hussein, M.; Allobawi, R.; Levou, I.; Blaskovich, M.A.T.; Rao, G.G.; Li, J.; Velkov, T. Mechanisms Underlying Synergistic Killing of Polymyxin B in Combination with Cannabidiol against Acinetobacter baumannii: A Metabolomic Study. Pharmaceutics 2022, 14, 786. [Google Scholar] [CrossRef]
- Burgeiro, A.; Bento, A.C.; Gajate, C.; Oliveira, P.J.; Mollinedo, F. Rapid human melanoma cell death induced by sanguinarine through oxidative stress. Eur. J. Pharmacol. 2013, 705, 109–118. [Google Scholar] [CrossRef]
- Marasco, D.; Vicidomini, C.; Krupa, P.; Cioffi, F.; Huy, P.D.Q.; Li, M.S.; Florio, D.; Broersen, K.; De Pandis, M.F.; Roviello, G.N. Plant isoquinoline alkaloids as potential neurodrugs: A comparative study of the effects of benzo[c]phenanthridine and berberine-based compounds on β-amyloid aggregation. Chem. Biol. Interact. 2021, 334, 109300. [Google Scholar] [CrossRef]
- Shu, D.; Zhu, Y.; Lu, M.; He, A.D.; Chen, J.B.; Ye, D.S.; Liu, Y.; Zeng, X.B.; Ma, R.; Ming, Z.Y. Sanguinarine Attenuates Collagen-Induced Platelet Activation and Thrombus Formation. Biomedicines 2021, 9, 444. [Google Scholar] [CrossRef]
- Zhang, Q.; Lyu, Y.; Huang, J.; Zhang, X.; Yu, N.; Wen, Z.; Chen, S. Antibacterial activity and mechanism of sanguinarine against Providencia rettgeri in vitro. PeerJ 2020, 8, e9543. [Google Scholar] [CrossRef]
- Zhong, H.; Hu, D.D.; Hu, G.H.; Su, J.; Bi, S.; Zhang, Z.E.; Wang, Z.; Zhang, R.L.; Xu, Z.; Jiang, Y.Y.; et al. Activity of Sanguinarine against Candida albicans Biofilms. Antimicrob. Agents Chemother. 2017, 61, e02259-16. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Luo, Y.; Fang, Z.; Teng, W.; Yu, Y.; Tian, J.; Guo, P.; Xu, R.; Wu, J.; Li, Y. Mechanism of Sanguinarine in Inhibiting Macrophages to Promote Metastasis and Proliferation of Lung Cancer via Modulating the Exosomes in A549 Cells. Onco Targets Ther. 2020, 13, 8989–9003. [Google Scholar] [CrossRef] [PubMed]
- Obiang-Obounou, B.W.; Kang, O.H.; Choi, J.G.; Keum, J.H.; Kim, S.B.; Mun, S.H.; Shin, D.W.; Kim, K.W.; Park, C.B.; Kim, Y.G.; et al. The mechanism of action of sanguinarine against methicillin-resistant Staphylococcus aureus. J. Toxicol. Sci. 2011, 36, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.H.; Wang, W.M.; Zhang, Z.; Sun, L.; Wu, S.C. Natural Antibacterial and Antivirulence Alkaloids From Macleaya cordata Against Methicillin-Resistant Staphylococcus aureus. Front. Pharmacol. 2022, 13, 813172. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Liu, W.; Liu, M.; Zhang, J.; Yang, M.; Wang, T.; Qian, W. In vitro anti-biofilm efficacy of sanguinarine against carbapenem-resistant Serratia marcescens. Biofouling 2021, 37, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Obiang-Obounou, B.W.; Kang, O.H.; Choi, J.G.; Keum, J.H.; Kim, S.B.; Mun, S.H.; Shin, D.W.; Park, C.B.; Kim, Y.G.; Han, S.H.; et al. In vitro potentiation of ampicillin, oxacillin, norfloxacin, ciprofloxacin, and vancomycin by sanguinarine against methicillin-resistant Staphylococcus aureus. Foodborne Pathog. Dis. 2011, 8, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Barker, W.T.; Jania, L.A.; Melander, R.J.; Koller, B.H.; Melander, C. Eukaryotic phosphatase inhibitors enhance colistin efficacy in gram-negative bacteria. Chem. Biol. Drug Des. 2020, 96, 1180–1186. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Fan, T.; Li, W.; Xing, W.; Huang, H. The anti-inflammatory effects of sanguinarine and its modulation of inflammatory mediators from peritoneal macrophages. Eur. J. Pharmacol. 2012, 689, 262–269. [Google Scholar] [CrossRef]
- Singh, N.; Sharma, B. Toxicological Effects of Berberine and Sanguinarine. Front. Mol. Biosci. 2018, 5, 21. [Google Scholar] [CrossRef]
- Janovská, M.; Kubala, M.; Simánek, V.; Ulrichová, J. Interaction of sanguinarine and its dihydroderivative with the Na+/K+-ATPase. Complex view on the old problem. Toxicol. Lett. 2010, 196, 56–59. [Google Scholar] [CrossRef]
- Cao, F.J.; Yang, R.; Lv, C.; Ma, Q.; Lei, M.; Geng, H.L.; Zhou, L. Pseudocyanides of sanguinarine and chelerythrine and their series of structurally simple analogues as new anticancer lead compounds: Cytotoxic activity, structure-activity relationship and apoptosis induction. Eur. J. Pharm. Sci. 2015, 67, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Justo, J.A.; Bosso, J.A. Adverse reactions associated with systemic polymyxin therapy. Pharmacotherapy 2015, 35, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Vardakas, K.Z.; Falagas, M.E. Colistin versus polymyxin B for the treatment of patients with multidrug-resistant Gram-negative infections: A systematic review and meta-analysis. Int. J. Antimicrob. Agents 2017, 49, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Ramana, P.; Adams, E.; Augustijns, P.; Van Schepdael, A. Metabonomics and drug development. Methods Mol. Biol. 2015, 1277, 195–207. [Google Scholar] [PubMed]
- Mielko, K.A.; Jabłoński, S.J.; Milczewska, J.; Sands, D.; Łukaszewicz, M.; Młynarz, P. Metabolomic studies of Pseudomonas aeruginosa. World J. Microbiol. Biotechnol. 2019, 35, 178. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Xu, G. Mass-spectrometry-based microbial metabolomics: Recent developments and applications. Anal. Bioanal. Chem. 2015, 407, 669–680. [Google Scholar] [CrossRef]
- Kang, M.; Kim, W.; Lee, J.; Jung, H.S.; Jeon, C.O.; Park, W. 6-Bromo-2-naphthol from Silene armeria extract sensitizes Acinetobacter baumannii strains to polymyxin. Sci. Rep. 2022, 12, 8546. [Google Scholar] [CrossRef]
- Huang, W.; Zhang, J.; Liu, S.; Hu, C.; Zhang, M.; Cheng, S.; Yu, H.; Zheng, M.; Wu, J.; Lu, Y.; et al. Disulfiram Enhances the Activity of Polymyxin B Against Klebsiella pneumoniae by Inhibiting Lipid A Modification. Infect. Drug Resist. 2022, 15, 295–306. [Google Scholar] [CrossRef]
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains | PMB (μg/mL) | SA (μg/mL) | FICIs | ||
|---|---|---|---|---|---|
| MIC in Single Use | MIC in Combination | MIC in Single Use | MIC in Combination | ||
| E. coli ATCC 25922 | 0.5 | 0.125 | 16 | 2 | 0.375 |
| E. coli ATCC 11775 | 1 | 0.25 | 16 | 4 | 0.5 |
| E. coli ATCC 9637 | 1 | 0.25 | 32 | 8 | 0.5 |
| E. coli ATCC 35218 | 1 | 0.25 | 16 | 2 | 0.375 |
| K. pneumoniae ATCC 700603 | 2 | 0.25 | 128 | 32 | 0.375 |
| P. aeruginosa ATCC 27853 | 1 | 0.25 | 256 | 32 | 0.375 |
| A. baumannii ATCC 19606 | 1 | 0.125 | 64 | 8 | 0.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiao, L.; Zhang, Y.; Chen, Y.; Chi, X.; Ding, J.; Zhang, H.; Han, Y.; Zhang, B.; Jiang, J.; Lin, Y. Synergistic Activity and Mechanism of Sanguinarine with Polymyxin B against Gram-Negative Bacterial Infections. Pharmaceutics 2024, 16, 70. https://doi.org/10.3390/pharmaceutics16010070
Qiao L, Zhang Y, Chen Y, Chi X, Ding J, Zhang H, Han Y, Zhang B, Jiang J, Lin Y. Synergistic Activity and Mechanism of Sanguinarine with Polymyxin B against Gram-Negative Bacterial Infections. Pharmaceutics. 2024; 16(1):70. https://doi.org/10.3390/pharmaceutics16010070
Chicago/Turabian StyleQiao, Luyao, Yu Zhang, Ying Chen, Xiangyin Chi, Jinwen Ding, Hongjuan Zhang, Yanxing Han, Bo Zhang, Jiandong Jiang, and Yuan Lin. 2024. "Synergistic Activity and Mechanism of Sanguinarine with Polymyxin B against Gram-Negative Bacterial Infections" Pharmaceutics 16, no. 1: 70. https://doi.org/10.3390/pharmaceutics16010070