
Preclinical Study in Mouse Thymus and Thymocytes: Effects of Treatment with a Combination of Sodium Dichloroacetate and Sodium Valproate on Infectious Inflammation Pathways
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Investigational Medicinal Product
2.2. The Groups of Mice and Experiments
2.3. Thymus and Thymocyte Preparation
2.4. The Histological and Immunohistochemical Examination of the Thymus
2.5. Extraction of Total RNA from Thymocytes
2.6. Evaluation of the Slc5a8 Expression in Thymocytes
2.7. Next-Generation Sequencing
2.8. Statistical Analysis
2.9. Bioinformatic Analysis
3. Results and Discussion
3.1. Effects of DCA–VPA Treatment on Body Weight, Thymus Structure, and Number of Hassall’s Corpuscles in the Medulla of the Thymus
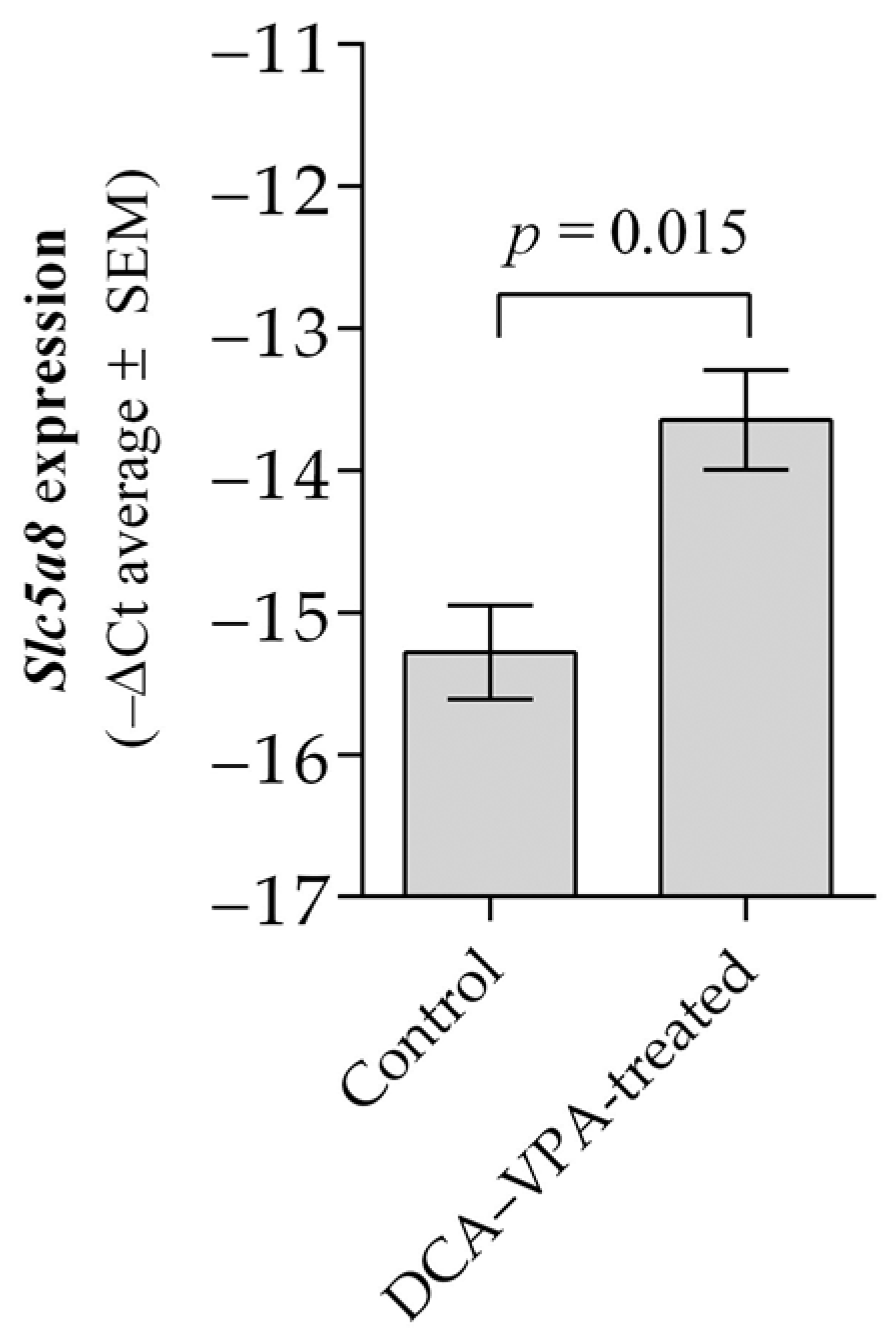
3.2. The Effect of DCA–VPA Treatment on Thymocytes’ Slc5a8 Gene Expression
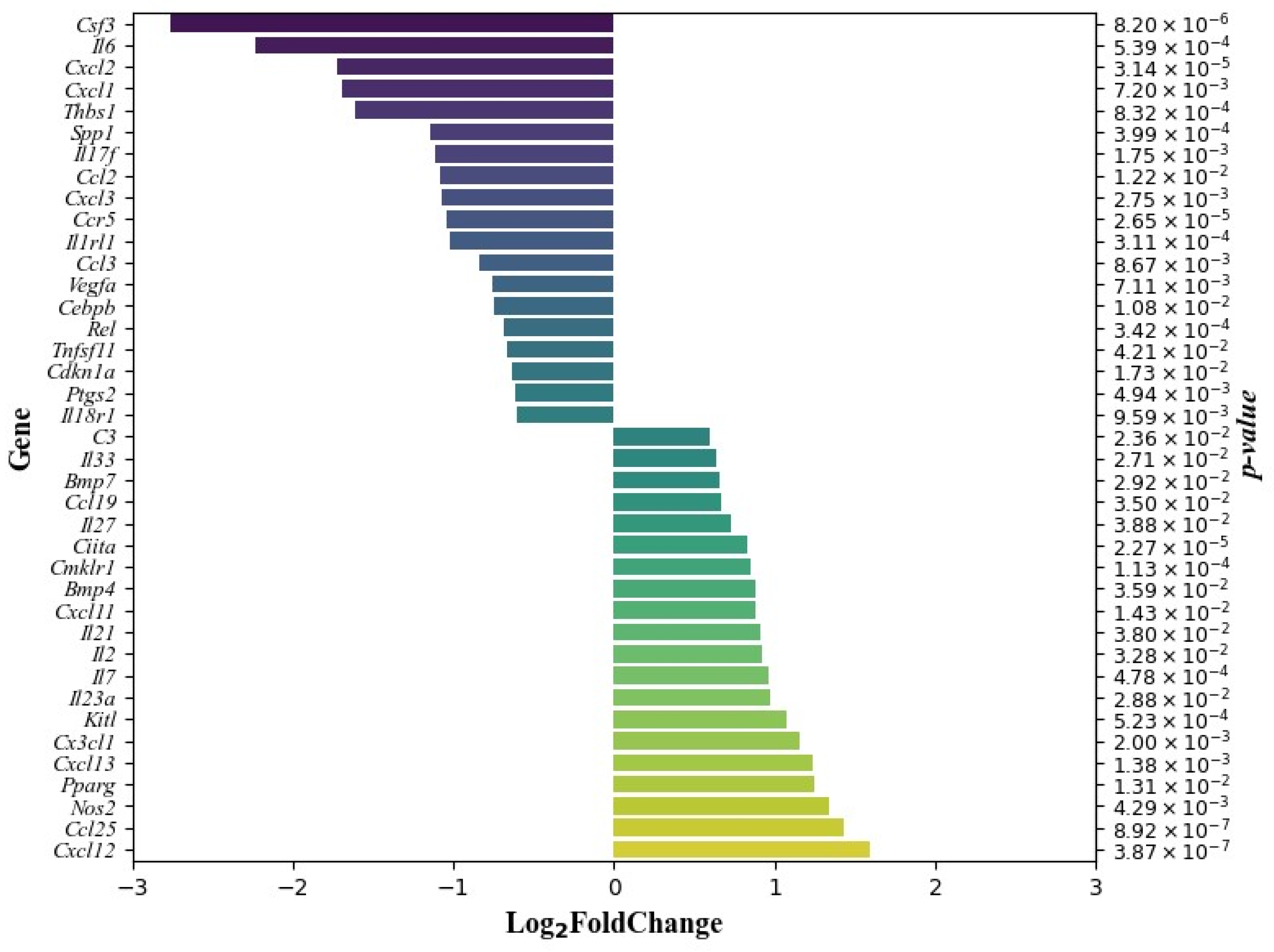
3.3. The Effect of DCA–VPA on Related Inflammation and Immune Response Genes’ Expression in Mice Thymocytes
3.3.1. A Possible DCA–VPA Treatment Effect on Thymus Function
3.3.2. Effects of DCA–VPA Treatment on Cytokine Activity Pathway, Inflammatory Response Pathway, and Il17 Signaling Pathway Genes in Male Balb/c Mice Thymocytes
DCA–VPA Effects on the Cytokine Activity Pathway
DCA–VPA Effects on the Inflammatory Response Pathway
DCA–VPA Effects on the Il17 Signaling Pathway
4. Conclusions
5. Patent
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Correction Statement
References
- Fleischmann, C.; Scherag, A.; Adhikari, N.K.J.; Hartog, C.S.; Tsaganos, T.; Schlattmann, P.; Angus, D.C.; Reinhart, K. Assessment of Global Incidence and Mortality of Hospital-Treated Sepsis Current Estimates and Limitations. Am. J. Respir. Crit. Care Med. 2016, 193, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Vincent, J.L.; Bakker, J. Blood Lactate Levels in Sepsis: In 8 Questions. Curr. Opin. Crit. Care 2021, 27, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Langley, R.J.; Tsalik, E.L.; Van Velkinburgh, J.C.; Glickman, S.W.; Rice, B.J.; Wang, C.; Chen, B.; Carin, L.; Suarez, A.; Mohney, R.P.; et al. Sepsis: An Integrated Clinico-Metabolomic Model Improves Prediction of Death in Sepsis. Sci. Transl. Med. 2013, 5, 195ra95. [Google Scholar] [CrossRef] [PubMed]
- Kotas, M.E.; Medzhitov, R. Homeostasis, Inflammation, and Disease Susceptibility. Cell 2015, 160, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.P.; Teixeira, L.; Moita, L.F. Disease Tolerance and Immunity in Host Protection against Infection. Nat. Rev. Immunol. 2017, 17, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Shoraka, S.; Samarasinghe, A.E.; Ghaemi, A.; Mohebbi, S.R. Host Mitochondria: More than an Organelle in SARS-CoV-2 Infection. Front. Cell. Infect. Microbiol. 2023, 13, 1228275. [Google Scholar] [CrossRef] [PubMed]
- Stakišaitis, D.; Kapočius, L.; Valančiūte, A.; Balnyte, I.; Tamošuitis, T.; Vaitkevičiūs, A.; Sužiedelis, K.; Urbonienė, D.; Tatarūnas, V.; Urbonienė, E.; et al. SARS-CoV-2 Infection, Sex-Related Differences, and a Possible Personalized Treatment Approach with Valproic Acid: A Review. Biomedicines 2022, 10, 962. [Google Scholar] [CrossRef]
- Mainali, R.; Zabalawi, M.; Long, D.; Buechler, N.; Quillen, E.; Key, C.C.; Zhu, X.; Parks, J.S.; Furdui, C.; Stacpoole, P.W.; et al. Dichloroacetate Reverses Sepsis-Induced Hepatic Metabolic Dysfunction. eLife 2021, 10, e64611. [Google Scholar] [CrossRef]
- Patel, M.S.; Nemeria, N.S.; Furey, W.; Jordan, F. The Pyruvate Dehydrogenase Complexes: Structure-Based Function and Regulation. J. Biol. Chem. 2014, 289, 16615–16623. [Google Scholar] [CrossRef]
- James, M.O.; Jahn, S.C.; Zhong, G.; Smeltz, M.G.; Hu, Z.; Stacpoole, P.W. Therapeutic Applications of Dichloroacetate and the Role of Glutathione Transferase Zeta-1. Pharmacol. Ther. 2017, 170, 166–180. [Google Scholar] [CrossRef]
- El Sayed, S.M.; Baghdadi, H.; Ahmed, N.S.; Almaramhy, H.H.; Mahmoud, A.A.A.; El-Sawy, S.A.; Ayat, M.; Elshazley, M.; Abdel-Aziz, W.; Abdel-Latif, H.M.; et al. Dichloroacetate Is an Antimetabolite That Antagonizes Acetate and Deprives Cancer Cells from Its Benefits: A Novel Evidence-Based Medical Hypothesis. Med. Hypotheses 2019, 122, 206–209. [Google Scholar] [CrossRef]
- Zeng, Z.; Huang, Q.; Mao, L.; Wu, J.; An, S.; Chen, Z.; Zhang, W. The Pyruvate Dehydrogenase Complex in Sepsis: Metabolic Regulation and Targeted Therapy. Front. Nutr. 2021, 8, 783164. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Jeon, J.H.; Min, B.K.; Ha, C.M.; Thoudam, T.; Park, B.Y.; Lee, I.K. Role of the Pyruvate Dehydrogenase Complex in Metabolic Remodeling: Differential Pyruvate Dehydrogenase Complex Functions in Metabolism. Diabetes Metab. J. 2018, 42, 270–281. [Google Scholar] [CrossRef] [PubMed]
- McCall, C.E.; Zabalawi, M.; Liu, T.; Martin, A.; Long, D.L.; Buechler, N.L.; Arts, R.J.W.; Netea, M.; Yoza, B.K.; Stacpoole, P.W.; et al. Pyruvate Dehydrogenase Complex Stimulation Promotes Immunometabolic Homeostasis and Sepsis Survival. JCI Insight 2018, 3, e99292. [Google Scholar] [CrossRef] [PubMed]
- Kelly, B.; O’Neill, L.A.J. Metabolic Reprogramming in Macrophages and Dendritic Cells in Innate Immunity. Cell Res. 2015, 25, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Eyenga, P.; Roussel, D.; Morel, J.; Rey, B.; Romestaing, C.; Gueguen-Chaignon, V.; Sheu, S.S.; Viale, J.P. Time Course of Liver Mitochondrial Function and Intrinsic Changes in Oxidative Phosphorylation in a Rat Model of Sepsis. Intensive Care Med. Exp. 2018, 6, 31. [Google Scholar] [CrossRef] [PubMed]
- Nuzzo, E.; Berg, K.M.; Andersen, L.W.; Balkema, J.; Montissol, S.; Cocchi, M.N.; Liu, X.; Donnino, M.W. Pyruvate Dehydrogenase Activity Is Decreased in the Peripheral Blood Mononuclear Cells of Patients with Sepsis: A Prospective Observational Trial. Ann. Am. Thorac. Soc. 2015, 12, 1662–1666. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Fan, M.; Wang, X.; Xu, J.; Wang, Y.; Tu, F.; Gill, P.S.; Ha, T.; Liu, L.; Williams, D.L.; et al. Lactate Promotes Macrophage HMGB1 Lactylation, Acetylation, and Exosomal Release in Polymicrobial Sepsis. Cell Death Differ. 2022, 29, 133–146. [Google Scholar] [CrossRef]
- Alamdari, N.; Constantin-Teodosiu, D.; Murton, A.J.; Gardiner, S.M.; Bennett, T.; Layfield, R.; Greenhaff, P.L. Temporal Changes in the Involvement of Pyruvate Dehydrogenase Complex in Muscle Lactate Accumulation during Lipopolysaccharide Infusion in Rats. J. Physiol. 2008, 586, 1767–1775. [Google Scholar] [CrossRef]
- Singer, P.M.; De Santis, V.; Vitale, D.; Jeffcoate, W. Multiorgan Failure Is an Adaptive, Endocrine-Mediated, Metabolic Response to Overwhelming Systemic Inaflammation. Lancet 2004, 364, 545–548. [Google Scholar] [CrossRef]
- Colegio, O.R.; Chu, N.Q.; Szabo, A.L.; Chu, T.; Rhebergen, A.M.; Jairam, V.; Cyrus, N.; Brokowski, C.E.; Eisenbarth, S.C.; Phillips, G.M.; et al. Functional Polarization of Tumour-Associated Macrophages by Tumour-Derived Lactic Acid. Nature 2014, 513, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Samuvel, D.J.; Sundararaj, K.P.; Nareika, A.; Lopes-Virella, M.F.; Huang, Y. Lactate Boosts TLR4 Signaling and NF-ΚB Pathway-Mediated Gene Transcription in Macrophages via Monocarboxylate Transporters and MD-2 Up-Regulation. J. Immunol. 2009, 184, 2476–2484. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Meng, Z.; Li, Y.; Zhao, J.; Wu, S.; Gou, S.; Wu, H. Prognostic Accuracy of the Serum Lactate Level, the SOFA Score and the QSOFA Score for Mortality among Adults with Sepsis. Scand. J. Trauma. Resusc. Emerg. Med. 2019, 27, 51. [Google Scholar] [CrossRef]
- Planche, T. Malaria and Fluids—Balancing Acts. Trends Parasitol. 2005, 21, 562–567. [Google Scholar] [CrossRef] [PubMed]
- An, S.; Yao, Y.; Hu, H.; Wu, J.; Li, J.; Li, L.; Wu, J.; Sun, M.; Deng, Z.; Zhang, Y.; et al. PDHA1 Hyperacetylation-Mediated Lactate Overproduction Promotes Sepsis-Induced Acute Kidney Injury via Fis1 Lactylation. Cell Death Dis. 2023, 14, 457. [Google Scholar] [CrossRef] [PubMed]
- Zemlin, A.E.; Sigwadhi, L.N.; Wiese, O.J.; Jalavu, T.P.; Chapanduka, Z.C.; Allwood, B.W.; Tamuzi, J.L.; Koegelenberg, C.F.; Irusen, E.M.; Lalla, U.; et al. The Association between Acid–base Status and Clinical Outcome in Critically Ill COVID-19 Patients Admitted to Intensive Care Unit with an Emphasis on High Anion Gap Metabolic Acidosis. Ann. Clin. Biochem. 2023, 60, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Long, D.; Zabalawi, M.; Ingram, B.; Yoza, B.K.; Stacpoole, P.W.; McCall, C.E. Stimulating Pyruvate Dehydrogenase Complex Reduces Itaconate Levels and Enhances TCA Cycle Anabolic Bioenergetics in Acutely Inflamed Monocytes. J. Leukoc. Biol. 2020, 107, 467–484. [Google Scholar] [CrossRef]
- McCall, C.E.; Zhu, X.; Zabalawi, M.; Long, D.; Quinn, M.A.; Yoza, B.K.; Stacpoole, P.W.; Vachharajani, V. Sepsis, Pyruvate, and Mitochondria Energy Supply Chain Shortage. J. Leukoc. Biol. 2022, 112, 1509–1514. [Google Scholar] [CrossRef]
- Perucca, E. Pharmacological and Therapeutic Properties of Valproate: A Summary after 35 Years of Clinical Experience. CNS Drugs 2002, 16, 695–714. [Google Scholar] [CrossRef]
- Michaelis, M.; Doerr, H.; Cinatl, J., Jr. Valproic Acid as Anti-Cancer Drug. Curr. Pharm. Des. 2007, 13, 3378–3393. [Google Scholar] [CrossRef]
- Andreu, S.; Ripa, I.; Bello-Morales, R.; López-Guerrero, J.A. Valproic Acid and Its Amidic Derivatives as New Antivirals against Alphaherpesviruses. Viruses 2020, 12, 1356. [Google Scholar] [CrossRef]
- Phiel, C.J.; Zhang, F.; Huang, E.Y.; Guenther, M.G.; Lazar, M.A.; Klein, P.S. Histone Deacetylase Is a Direct Target of Valproic Acid, a Potent Anticonvulsant, Mood Stabilizer, and Teratogen. J. Biol. Chem. 2001, 276, 36734–36741. [Google Scholar] [CrossRef] [PubMed]
- Adcock, I.M. HDAC Inhibitors as Anti-Inflammatory Agents. Br. J. Pharmacol. 2007, 150, 829–831. [Google Scholar] [CrossRef] [PubMed]
- Van Beneden, K.; Geers, C.; Pauwels, M.; Mannaerts, I.; Verbeelen, D.; Van Grunsven, L.A.; Van Den Branden, C. Valproic Acid Attenuates Proteinuria and Kidney Injury. J. Am. Soc. Nephrol. 2011, 22, 1863–1875. [Google Scholar] [CrossRef] [PubMed]
- Roda, M.A.; Sadik, M.; Gaggar, A.; Hardison, M.T.; Jablonsky, M.J.; Braber, S.; Blalock, J.E.; Redegeld, F.A.; Folkerts, G.; Jackson, P.L. Targeting Prolyl Endopeptidase with Valproic Acid as a Potential Modulator of Neutrophilic Inflammation. PLoS ONE 2014, 9, e97594. [Google Scholar]
- Roger, T.; Lugrin, J.; Le Roy, D.; Goy, G.; Mombelli, M.; Koessler, T.; Ding, X.C.; Chanson, A.L.; Reymond, M.K.; Miconnet, I.; et al. Histone Deacetylase Inhibitors Impair Innate Immune Responses to Toll-like Receptor Agonists and to Infection. Blood 2011, 117, 1205–1217. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Guo, X. Valproic Acid Ameliorates Coxsackievirus-B3-Induced Viral Myocarditis by Modulating Th17/Treg Imbalance. Virol. J. 2016, 13, 168. [Google Scholar] [CrossRef]
- Ximenes, J.C.M.; De Oliveira Gonçalves, D.; Siqueira, R.M.P.; Neves, K.R.T.; Santos Cerqueira, G.; Correia, A.O.; Félix, F.H.C.; Leal, L.K.A.M.; De Castro Brito, G.A.; Da Graça Naffah-Mazzacorati, M.; et al. Valproic Acid: An Anticonvulsant Drug with Potent Antinociceptive and Anti-Inflammatory Properties. Naunyn. Schmiedeberg’s Arch. Pharmacol. 2013, 386, 575–587. [Google Scholar] [CrossRef]
- Shyer, J.A.; Flavell, R.A.; Bailis, W. Metabolic Signaling in T Cells. Cell Res. 2020, 30, 649–659. [Google Scholar] [CrossRef]
- Sinclair, L.V.; Rolf, J.; Emslie, E.; Shi, Y.B.; Taylor, P.M.; Cantrell, D.A. Control of Amino-Acid Transport by Antigen Receptors Coordinates the Metabolic Reprogramming Essential for T Cell Differentiation. Nat. Immunol. 2013, 14, 500–508. [Google Scholar] [CrossRef]
- Kidani, Y.; Elsaesser, H.; Hock, M.B.; Vergnes, L.; Williams, K.J.; Argus, J.P.; Marbois, B.N.; Komisopoulou, E.; Wilson, E.B.; Osborne, T.F.; et al. Sterol Regulatory Element-Binding Proteins Are Essential for the Metabolic Programming of Effector T Cells and Adaptive Immunity. Nat. Immunol. 2013, 14, 489–499. [Google Scholar] [CrossRef]
- Codo, A.C.; Davanzo, G.G.; de Brito Monteiro, L.; de Souza, G.F.; Muraro, S.P.; Virgilio-da-Silva, J.V.; Prodonoff, J.S.; Carregari, V.C.; de Biagi Junior, C.A.O.; Crunfli, F.; et al. Erratum: Elevated Glucose Levels Favor SARS-CoV-2 Infection and Monocyte Response through a HIF-1α/Glycolysis-Dependent Axis. Cell Metab. 2020, 32, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Luan, Y.; Luan, Y.; He, H.; Jue, B.; Yang, Y.; Qin, B.; Ren, K. Glucose Metabolism Disorder: A Potential Accomplice of SARS-CoV-2. Int. J. Obes. 2023, 47, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Jena, G. Valproic Acid Improves Glucose Homeostasis by Increasing Beta-Cell Proliferation, Function, and Reducing Its Apoptosis through HDAC Inhibition in Juvenile Diabetic Rat. J. Biochem. Mol. Toxicol. 2016, 30, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Rakitin, A.; Kõks, S.; Haldre, S. Valproate Modulates Glucose Metabolism in Patients with Epilepsy after First Exposure. Epilepsia 2015, 56, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Schoenmann, N.; Tannenbaum, N.; Hodgeman, R.M.; Raju, R.P. Regulating Mitochondrial Metabolism by Targeting Pyruvate Dehydrogenase with Dichloroacetate, a Metabolic Messenger. Biochim. Biophys. Acta-Mol. Basis Dis. 2023, 1869, 166769. [Google Scholar] [CrossRef] [PubMed]
- Milutinovic, S.; Detich, N.; Szyf, M. Valproate Induces Widespread Epigenetic Reprogramming Which Involves Demethylation of Specific Genes. Carcinogenesis 2007, 28, 560–571. [Google Scholar] [CrossRef]
- Veronezi, G.M.B.; Felisbino, M.B.; Gatti, M.S.V.; Mello, M.L.S.; De Vidal, B.C. DNA Methylation Changes in Valproic Acid-Treated HeLa Cells as Assessed by Image Analysis, Immunofluorescence and Vibrational Microspectroscopy. PLoS ONE 2017, 12, e0170740. [Google Scholar] [CrossRef]
- Thangaraju, M.; Ananth, S.; Martin, P.M.; Roon, P.; Smith, S.B.; Sterneck, E.; Prasad, P.D.; Ganapathy, V. C/Ebpδ Null Mouse as a Model for the Double Knock-out of Slc5a8 and Slc5a12 in Kidney. J. Biol. Chem. 2006, 281, 26769–26773. [Google Scholar] [CrossRef]
- Frank, H.; Gröger, N.; Diener, M.; Becker, C.; Braun, T.; Boettger, T. Lactaturia and Loss of Sodium-Dependent Lactate Uptake in the Colon of SLC5A8-Deficient Mice. J. Biol. Chem. 2008, 283, 24729–24737. [Google Scholar] [CrossRef]
- Singh, N.; Thangaraju, M.; Prasad, P.D.; Martin, P.M.; Lambert, N.A.; Boettger, T.; Offermanns, S.; Ganapathy, V. Blockade of Dendritic Cell Development by Bacterial Fermentation Products Butyrate and Propionate through a Transporter (Slc5a8)-Dependent Inhibition of Histone Deacetylases. J. Biol. Chem. 2010, 285, 27601–27608. [Google Scholar] [CrossRef]
- Zimmerman, M.A.; Singh, N.; Martin, P.M.; Thangaraju, M.; Ganapathy, V.; Waller, J.L.; Shi, H.; Robertson, K.D.; Munn, D.H.; Liu, K. Butyrate Suppresses Colonic Inflammation through HDAC1-Dependent Fas Upregulation and Fas-Mediated Apoptosis of T Cells. Am. J. Physiol.-Gastrointest. Liver Physiol. 2012, 302, 1405–1415. [Google Scholar] [CrossRef]
- Savino, W.; Durães, J.; Maldonado-Galdeano, C.; Perdigon, G.; Mendes-da-Cruz, D.A.; Cuervo, P. Thymus, Undernutrition, and Infection: Approaching Cellular and Molecular Interactions. Front. Nutr. 2022, 9, 948488. [Google Scholar] [CrossRef]
- Stanevičiūtė, J.; Juknevičienė, M.; Balnyte, I.; Valančiūte, A.; Lesauskaite, V.; Fadejeva, J.; Stakauskas, R.; Stakišaitis, D. Gender-Related Effect of Sodium Dichloroacetate on the Number of Hassall’s Corpuscles and RNA NKCC1 Expression in Rat Thymus. Biomed Res. Int. 2019, 2019, 1602895. [Google Scholar] [CrossRef] [PubMed]
- Juknevičienė, M.; Balnytė, I.; Valančiūtė, A.; Lesauskaitė, V.; Stanevičiūtė, J.; Curkūnavičiūtė, R.; Stakišaitis, D. Valproic Acid Inhibits NA-K-2CL Cotransporter RNA Expression in Male but Not in Female Rat Thymocytes. Dose-Response 2019, 17, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize Analysis Results for Multiple Tools and Samples in a Single Report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Cunningham, F.; Allen, J.E.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Bennett, R.; et al. Ensembl 2022. Nucleic Acids Res. 2022, 50, 988–995. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A Web Server for Functional Enrichment Analysis and Functional Annotation of Gene Lists (2021 Update). Nucleic Acids Res. 2022, 50, 216–221. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. ClusterProfiler 4.0: A Universal Enrichment Tool for Interpreting Omics Data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef] [PubMed]
- Ciofani, M.; Zúñiga-Pflücker, J.C. The Thymus as an Inductive Site for T Lymphopoiesis. Annu. Rev. Cell Dev. Biol. 2007, 23, 463–493. [Google Scholar] [CrossRef]
- Savino, W.; Mendes-Da-Cruz, D.A.; Lepletier, A.; Dardenne, M. Hormonal Control of T-Cell Development in Health and Disease. Nat. Rev. Endocrinol. 2016, 12, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Valančiute, A.; Mozuraite, R.; Balnyte, I.; Didžiapetriene, J.; Matusevičius, P.; Stakišaitis, D. Sodium Valproate Effect on the Structure of Rat Glandule Thymus: Gender-Related Differences. Exp. Toxicol. Pathol. 2015, 67, 399–406. [Google Scholar] [CrossRef]
- Sebastianelli, M.; Forte, C.; Galarini, R.; Gobbi, M.; Pistidda, E.; Moncada, C.; Cannizzo, F.T.; Pezzolato, M.; Bozzetta, E.; Cenci-Goga, B.T.; et al. LC-MS/MS Analyses of Bile and Histological Analyses of Thymus as Diagnostic Tools to Detect Low Dose Dexamethasone Illicit Treatment in Beef Cattle at Slaughterhouse. Steroids 2020, 160, 108671. [Google Scholar] [CrossRef]
- Asghar, A.; Syed, Y.M.; Nafis, F.A. Polymorphism of Hassall′s Corpuscles in Thymus of Human Fetuses. Int. J. Appl. Basic Med. Res. 2012, 2, 7–10. [Google Scholar] [CrossRef]
- Hale, L.P.; Markert, M.L. Corticosteroids Regulate Epithelial Cell Differentiation and Hassall Body Formation in the Human Thymus. J. Immunol. 2004, 172, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Kadouri, N.; Nevo, S.; Goldfarb, Y.; Abramson, J. Thymic Epithelial Cell Heterogeneity: TEC by TEC. Nat. Rev. Immunol. 2020, 20, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Laan, M.; Bichele, R.; Kisand, K.; Scott, H.S.; Peterson, P. Post-Aire Maturation of Thymic Medullary Epithelial Cells Involves Selective Expression of Keratinocyte-Specific Autoantigens. Front. Immunol. 2012, 3, 19. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Ossa-Gomez, L.J. A Quantitative Histologic Comparison of the Thymus in 100 Healthy and Diseased Adults. Am. J. Clin. Pathol. 1981, 76, 657–665. [Google Scholar] [CrossRef]
- Skogberg, G.; Lundberg, V.; Lindgren, S.; Gudmundsdottir, J.; Sandström, K.; Kämpe, O.; Annerén, G.; Gustafsson, J.; Sunnegårdh, J.; van der Post, S.; et al. Altered Expression of Autoimmune Regulator in Infant Down Syndrome Thymus, a Possible Contributor to an Autoimmune Phenotype. J. Immunol. 2014, 193, 2187–2195. [Google Scholar] [CrossRef]
- Wang, J.; Sekai, M.; Matsui, T.; Fujii, Y.; Matsumoto, M.; Takeuchi, O.; Minato, N.; Hamazaki, Y. Hassall’s Corpuscles with Cellular-Senescence Features Maintain IFNα Production through Neutrophils and PDC Activation in the Thymus. Int. Immunol. 2019, 31, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Yano, M.; Kuroda, N.; Han, H.; Meguro-Horike, M.; Nishikawa, Y.; Kiyonari, H.; Maemura, K.; Yanagawa, Y.; Obata, K.; Takahashi, S.; et al. Aire Controls the Differentiation Program of Thymic Epithelial Cells in the Medulla for the Establishment of Self-Tolerance. J. Exp. Med. 2008, 205, 2827–2838. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, N.; Wang, Y.H.; Lee, H.K.; Ito, T.; Wang, Y.H.; Cao, W.; Liu, Y.J. Hassall’s Corpuscles Instruct Dendritic Cells to Induce CD4+ CD25+ Regulatory T Cells in Human Thymus. Nature 2005, 436, 1181–1185. [Google Scholar] [CrossRef]
- Besin, G.; Gaudreau, S.; Menard, M.; Guindi, C.; Dupuis, G.; Amrani, A. Thymic Stromal Lymphopoietin and Thymic Stromal Lymphopoietin-Conditioned Dendritic Cells Induce Regulatory T-Cell Differentiation and Protection of NOD Mice against Diabetes. Diabetes 2008, 57, 2107–2117. [Google Scholar] [CrossRef]
- Laan, M.; Salumets, A.; Klein, A.; Reintamm, K.; Bichele, R.; Peterson, H.; Peterson, P. Post-Aire Medullary Thymic Epithelial Cells and Hassall’s Corpuscles as Inducers of Tonic Pro-Inflammatory Microenvironment. Front. Immunol. 2021, 12, 635569. [Google Scholar] [CrossRef]
- Bhutia, Y.D.; Babu, E.; Ramachandran, S.; Yang, S.; Thangaraju, M.; Ganapathy, V. SLC Transporters as a Novel Class of Tumour Suppressors: Identity, Function and Molecular Mechanisms. Biochem. J. 2016, 473, 1113–1124. [Google Scholar] [CrossRef]
- Babu, E.; Ramachandran, S.; Coothankandaswamy, V.; Elangovan, S.; Prasad, P.D.; Ganapathy, V.; Thangaraju, M. Role of SLC5A8, a Plasma Membrane Transporter and a Tumor Suppressor, in the Antitumor Activity of Dichloroacetate. Oncogene 2011, 30, 4026–4037. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhao, S.; Cheng, Z.; Ren, Y.; Shi, X.; Mu, J.; Ge, X.; Dai, Y.; Li, L.; Zhang, Z. Akkermansia Muciniphila Protects against Antibiotic-Associated Diarrhea in Mice. Probiot. Antimicrob. Proteins 2023, 15, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Damanskienė, E.; Balnytė, I.; Valančiūtė, A.; Alonso, M.M.; Stakišaitis, D. Different Effects of Valproic Acid on SLC12A2, SLC12A5 and SLC5A8 Gene Expression in Pediatric Glioblastoma Cells as an Approach to Personalised Therapy. Biomedicines 2022, 10, 968. [Google Scholar] [CrossRef] [PubMed]
- Colantonio, A.D.; Epeldegui, M.; Jesiak, M.; Jachimowski, L.; Blom, B.; Uittenbogaart, C.H. IFN-α Is Constitutively Expressed in the Human Thymus, but Not in Peripheral Lymphoid Organs. PLoS ONE 2011, 6, e24252. [Google Scholar] [CrossRef]
- Lienenklaus, S.; Cornitescu, M.; Ziętara, N.; Łyszkiewicz, M.; Gekara, N.; Jabłońska, J.; Edenhofer, F.; Rajewsky, K.; Bruder, D.; Hafner, M.; et al. Novel Reporter Mouse Reveals Constitutive and Inflammatory Expression of IFN-β In Vivo. J. Immunol. 2009, 183, 3229–3236. [Google Scholar] [CrossRef]
- Trampont, P.C.; Tosello-Trampont, A.C.; Shen, Y.; Duley, A.K.; Sutherland, A.E.; Bender, T.P.; Littman, D.R.; Ravichandran, K.S. CXCR4 Acts as a Costimulator during Thymic Β-Selection. Nat. Immunol. 2010, 11, 162–170. [Google Scholar] [CrossRef]
- Annunziato, F.; Romagnani, P.; Cosmi, L.; Lazzeri, E.; Romagnani, S. Chemokines and Lymphopoiesis in Human Thymus. Trends Immunol. 2001, 22, 277–281. [Google Scholar] [CrossRef]
- Campbell, J.J.; Pan, J.; Butcher, E.C. Cutting Edge: Developmental Switches in Chemokine Responses during T Cell Maturation. J. Immunol. 1999, 163, 2353–2357. [Google Scholar] [CrossRef]
- Leite-de-Moraes, M.C.; Hontebeyrie-Joskowicz, M.; Dardenne, M.; Savino, W. Modulation of Thymocyte Subsets during Acute and Chronic Phases of Experimental Trypanosoma Cruzi Infection. Immunology 1992, 77, 95–98. [Google Scholar]
- Korn, T.; Hiltensperger, M. Role of IL-6 in the Commitment of T Cell Subsets. Cytokine 2021, 146, 155654. [Google Scholar] [CrossRef] [PubMed]
- Balcells, F.; Martínez Monteros, M.J.; Gómez, A.L.; Cazorla, S.I.; Perdigón, G.; Maldonado-Galdeano, C. Probiotic Consumption Boosts Thymus in Obesity and Senescence Mouse Models. Nutrients 2022, 14, 616. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, H.; Rothenberg, E.V. Cytokines, Transcription Factors, and the Initiation of T-Cell Development. Cold Spring Harb. Perspect. Biol. 2018, 10, a028621. [Google Scholar] [CrossRef] [PubMed]
- Palomino, D.C.; Marti, L.C. Chemokines and Immunity. Einstein 2015, 13, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.; Zhang, J.; Dai, S.; Wang, C.; Chen, Z.; Zhang, S.; Xu, J.; Qin, X.; Liu, F. CX3CR1 Deficiency Exacerbates Immune-Mediated Hepatitis by Increasing NF-ΚB-Mediated Cytokine Production in Macrophage and T Cell. Exp. Biol. Med. 2023, 248, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Griffith, J.W.; Sokol, C.L.; Luster, A.D. Chemokines and Chemokine Receptors: Positioning Cells for Host Defense and Immunity. Annu. Rev. Immunol. 2014, 32, 659–702. [Google Scholar] [CrossRef] [PubMed]
- Gaffen, S.L. Structure and Signalling in the IL-17 Receptor Family. Nat. Rev. Immunol. 2009, 9, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Brocker, C.; Thompson, D.; Matsumoto, A.; Nebert, D.W.; Vasiliou, V. Evolutionary Divergence and Functions of the Human Interleukin (IL) Gene Family. Hum. Genomics 2010, 5, 30–55. [Google Scholar] [CrossRef]
- Matsubara, E.; Yano, H.; Pan, C.; Komohara, Y.; Fujiwara, Y.; Zhao, S.; Shinchi, Y.; Kurotaki, D.; Suzuki, M. The Significance of SPP1 in Lung Cancers and Its Impact as a Marker for Protumor Tumor-Associated Macrophages. Cancers 2023, 15, 2250. [Google Scholar] [CrossRef]
- MacDonald, L.; Alivernini, S.; Tolusso, B.; Elmesmari, A.; Somma, D.; Perniola, S.; Paglionico, A.; Petricca, L.; Bosello, S.L.; Carfì, A.; et al. COVID-19 and RA Share an SPP1 Myeloid Pathway That Drives PD-L1+ Neutrophils and CD14+ Monocytes. JCI Insight 2021, 6, e147413. [Google Scholar] [CrossRef]
- Naito, T.; Ise, M.; Tanaka, Y.; Kohwi-Shigematsu, T.; Kondo, M. Crucial Roles of SATB1 in Regulation of Thymocyte Migration after Positive Selection. J. Immunol. 2023, 211, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Kondělková, K.; Vokurková, D.; Krejsek, J.; Borská, L.; Fiala, Z.; Ctirad, A. Regulatory T Cells (TREG) and Their Roles in Immune System with Respect to Immunopathological Disorders. Acta Medica 2010, 53, 73–77. [Google Scholar] [CrossRef]
- Alkharsah, K.R. VEGF Upregulation in Viral Infections and Its Possible Therapeutic Implications. Int. J. Mol. Sci. 2018, 19, 1642. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.Q.; Li, J.M. G-CSF and Its Receptor in Hematonosis. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2015, 23, 871–877. [Google Scholar] [PubMed]
- Dharra, R.; Kumar Sharma, A.; Datta, S. Emerging Aspects of Cytokine Storm in COVID-19: The Role of Proinflammatory Cytokines and Therapeutic Prospects. Cytokine 2023, 169, 156287. [Google Scholar] [CrossRef]
- Wang, H.; FitzPatrick, M.; Wilson, N.J.; Anthony, D.; Reading, P.C.; Satzke, C.; Dunne, E.M.; Licciardi, P.V.; Seow, H.J.; Nichol, K.; et al. CSF3R/CD114 Mediates Infection-Dependent Transition to Severe Asthma. J. Allergy Clin. Immunol. 2019, 143, 785–788. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Aloe, C.; Wilson, N.; Bozinovski, S. G-CSFR Antagonism Reduces Neutrophilic Inflammation during Pneumococcal and Influenza Respiratory Infections without Compromising Clearance. Sci. Rep. 2019, 9, 17732. [Google Scholar] [CrossRef] [PubMed]
- Takatsuka, H.; Takemoto, Y.; Mori, A.; Okamoto, T.; Kanamaru, A.; Kakishita, E. Common Features in the Onset of ARDS after Administration of Granulocyte Colony-Stimulating Factor. Chest 2002, 121, 1716–1720. [Google Scholar] [CrossRef] [PubMed]
- Sameni, M.; Mirmotalebisohi, S.A.; Dadashkhan, S.; Ghani, S.; Abbasi, M.; Noori, E.; Zali, H. COVID-19: A Novel Holistic Systems Biology Approach to Predict Its Molecular Mechanisms (In Vitro) and Repurpose Drugs. DARU J. Pharm. Sci. 2023, 31, 155–171. [Google Scholar] [CrossRef]
- Gould, S.E.; Day, M.; Jones, S.S.; Doral, H. Bmp-7 Regulates Chemokine, Cytokine, and Hemodynamic Gene Expression in Proximal Tubule Cells. Kidney Int. 2002, 61, 51–60. [Google Scholar] [CrossRef]
- Singla, D.K.; Singla, R.; Wang, J. BMP-7 Treatment Increases M2 Macrophage Differentiation and Reduces Inflammation and Plaque Formation in Apo E−/− Mice. PLoS ONE 2016, 11, e0147897. [Google Scholar] [CrossRef] [PubMed]
- Alves, N.L.; Arosa, F.A.; van Lier, R.A.W. Common γ Chain Cytokines: Dissidence in the Details. Immunol. Lett. 2007, 108, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Rochman, Y.; Spolski, R.; Leonard, W.J. New Insights into the Regulation of T Cells by Γc Family Cytokines. Nat. Rev. Immunol. 2009, 9, 480–490. [Google Scholar] [CrossRef]
- Collison, L.W.; Vignali, D.A.A. Interleukin-35: Odd One out or Part of the Family? Immunol. Rev. 2008, 226, 248–262. [Google Scholar] [CrossRef] [PubMed]
- Kinsella, S.; Dudakov, J.A. When the Damage Is Done: Injury and Repair in Thymus Function. Front. Immunol. 2020, 11, 1745. [Google Scholar] [CrossRef]
- Oshio, T.; Komine, M.; Tsuda, H.; Tominaga, S.I.; Saito, H.; Nakae, S.; Ohtsuki, M. Nuclear Expression of IL-33 in Epidermal Keratinocytes Promotes Wound Healing in Mice. J. Dermatol. Sci. 2017, 85, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Mencarelli, A.; Cipriani, S.; Francisci, D.; Santucci, L.; Baldelli, F.; Distrutti, E.; Fiorucci, S. Highly Specific Blockade of CCR5 Inhibits Leukocyte Trafficking and Reduces Mucosal Inflammation in Murine Colitis. Sci. Rep. 2016, 6, 30802. [Google Scholar] [CrossRef]
- Cayrol, C.; Girard, J.P. Interleukin-33 (IL-33): A Critical Review of Its Biology and the Mechanisms Involved in Its Release as a Potent Extracellular Cytokine. Cytokine 2022, 156, 155891. [Google Scholar] [CrossRef]
- Koh, S.S.; Ooi, S.C.Y.; Lui, N.M.Y.; Qiong, C.; Ho, L.T.Y.; Cheah, I.K.M.; Halliwell, B.; Herr, D.R.; Ong, W.Y. Effect of Ergothioneine on 7-Ketocholesterol-Induced Endothelial Injury. Neuro Mol. Med. 2021, 23, 184–198. [Google Scholar] [CrossRef]
- Gutierrez, L.S.; Lopez-Dee, Z.; Pidcock, K. Thrombospondin-1: Multiple Paths to Inflammation. Mediat. Inflamm. 2011, 2011, 296069. [Google Scholar]
- Hamldar, S.; Kiani, S.J.; Khoshmirsafa, M.; Nahand, J.S.; Mirzaei, H.; Khatami, A.R.; Kahyesh-Esfandiary, R.; Khanaliha, K.; Tavakoli, A.; Babakhaniyan, K.; et al. Expression Profiling of Inflammation-Related Genes Including IFI-16, NOTCH2, CXCL8, THBS1 in COVID-19 Patients. Biologicals 2022, 80, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, Q.; Wang, H.; Feng, P.; Yang, G.; Yang, L. Effects of Early Pregnancy on the Complement System in the Ovine Thymus. Vet. Res. Commun. 2022, 46, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, Y.; Kim, S.-I.; Kim, E.H.; Lambris, J.D.; Sandor, M.; Suresh, M. C3 Promotes Expansion of CD8+ and CD4+ T Cells in a Listeria Monocytogenes Infection. J. Immunol. 2009, 183, 2921–2931. [Google Scholar] [CrossRef] [PubMed]
- Sahu, S.K.; Ozantürk, A.N.; Kulkarni, D.H.; Ma, L.; Barve, R.A.; Dannull, L.; Lu, A.; Starick, M.; McPhatter, J.; Garnica, L.; et al. Lung Epithelial Cell–derived C3 Protects against Pneumonia-Induced Lung Injury. Sci. Immunol. 2023, 8, eabp9547. [Google Scholar] [CrossRef]
- Nagasubramanian, K.; Jha, S.; Rathore, A.S.; Gupta, K. Identification of Small Molecule Modulators of Class II Transactivator-I Using Computational Approaches. J. Biomol. Struct. Dyn. 2023, 41, 8349–8361. [Google Scholar] [CrossRef] [PubMed]
- Forlani, G.; Shallak, M.; Gatta, A.; Shaik, A.K.B.; Accolla, R.S. The NLR Member CIITA: Master Controller of Adaptive and Intrinsic Immunity and Unexpected Tool in Cancer Immunotherapy. Biomed. J. 2023, 46, 100631. [Google Scholar] [CrossRef] [PubMed]
- Provoost, S.; De Grove, K.C.; Fraser, G.L.; Lannoy, V.J.; Tournoy, K.G.; Brusselle, G.G.; Maes, T.; Joos, G.F. Pro- and Anti-Inflammatory Role of ChemR23 Signaling in Pollutant-Induced Inflammatory Lung Responses. J. Immunol. 2016, 196, 1882–1890. [Google Scholar] [CrossRef]
- Mannes, P.Z.; Barnes, C.E.; Biermann, J.; Latoche, J.D.; Day, K.E.; Zhu, Q.; Tabary, M.; Xiong, Z.; Nedrow, J.R.; Izar, B.; et al. Molecular Imaging of Chemokine-like Receptor 1 (CMKLR1) in Experimental Acute Lung Injury. Proc. Natl. Acad. Sci. USA 2023, 120, e2216458120. [Google Scholar] [CrossRef]
- Bondue, B.; Vosters, O.; de Nadai, P.; Glineur, S.; de Henau, O.; Luangsay, S.; van Gool, F.; Communi, D.; de Vuyst, P.; Desmecht, D.; et al. ChemR23 Dampens Lung Inflammation and Enhances Anti-Viral Immunity in a Mouse Model of Acute Viral Pneumonia. PLoS Pathog. 2011, 7, e1002358. [Google Scholar] [CrossRef]
- Luangsay, S.; Wittamer, V.; Bondue, B.; De Henau, O.; Rouger, L.; Brait, M.; Franssen, J.-D.; de Nadai, P.; Huaux, F.; Parmentier, M. Mouse ChemR23 Is Expressed in Dendritic Cell Subsets and Macrophages, and Mediates an Anti-Inflammatory Activity of Chemerin in a Lung Disease Model. J. Immunol. 2009, 183, 6489–6499. [Google Scholar] [CrossRef]
- Zheng, S.Y.; Shao, X.; Qi, Z.; Yan, M.; Tao, M.H.; Wu, X.M.; Zhang, L.; Ma, J.; Li, A.; Chang, M.X. Zebrafish Nos2a Benefits Bacterial Proliferation via Suppressing ROS and Inducing NO Production to Impair the Expressions of Inflammatory Cytokines and Antibacterial Genes. Fish Shellfish Immunol. 2023, 142, 109178. [Google Scholar] [CrossRef] [PubMed]
- Andelova, N.; Waczulikova, I.; Kunstek, L.; Talian, I.; Ravingerova, T.; Jasova, M.; Suty, S.; Ferko, M. Dichloroacetate as a Metabolic Modulator of Heart Mitochondrial Proteome under Conditions of Reduced Oxygen Utilization. Sci. Rep. 2022, 12, 16348. [Google Scholar] [CrossRef] [PubMed]
- Uzel, G.; Oylumlu, E.; Durmus, L.; Ciraci, C. Duality of Valproic Acid Effects on Inflammation, Oxidative Stress and Autophagy in Human Eosinophilic Cells. Int. J. Mol. Sci. 2023, 24, 13446. [Google Scholar] [CrossRef] [PubMed]
- Ahmadian, M.; Suh, J.M.; Hah, N.; Liddle, C.; Atkins, A.R.; Downes, M.; Evans, R.M. Pparγ Signaling and Metabolism: The Good, the Bad and the Future. Nat. Med. 2013, 19, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Mikacenic, C.; Hansen, E.E.; Radella, F.; Gharib, S.A.; Stapleton, R.D.; Wurfel, M.M. Interleukin-17A Is Associated with Alveolar Inflammation and Poor Outcomes in Acute Respiratory Distress Syndrome. Crit. Care Med. 2016, 44, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Righetti, R.F.; Dos Santos, T.M.; Camargo, L.D.N.; Barbosa Aristóteles, L.R.C.R.; Fukuzaki, S.; De Souza, F.C.R.; Santana, F.P.R.; De Agrela, M.V.R.; Cruz, M.M.; Alonso-Vale, M.I.C.; et al. Protective Effects of Anti-IL17 on Acute Lung Injury Induced by LPS in Mice. Front. Pharmacol. 2018, 9, 1021. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological Findings of COVID-19 Associated with Acute Respiratory Distress Syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Groups Studied | n | Ct Mean | ΔCt Mean ± SEM | ΔΔCT | |
|---|---|---|---|---|---|
| Slc5a8 | Gapdh | ||||
| Control | 6 | 34.77 | 19.49 | 15.27 ± 0.329 | −1.63 |
| DCA–VPA-treated | 6 | 33.40 | 19.76 | 13.64 ± 0.351 a | |
| Gene | Log2 Fold Change | Gene Expression (Average) | Gene Expression (Log2) | p Value | ||
|---|---|---|---|---|---|---|
| Treated | Control | Treated | Control | |||
| Bmp4 | 0.878 | 108.197 | 58.504 | 6.758 | 5.870 | 0.036 |
| Bmp7 | 0.661 | 212.423 | 134.572 | 7.731 | 7.072 | 0.029 |
| C3 | 0.598 | 7367.336 | 4867.781 | 12.847 | 12.249 | 0.024 |
| Ccl19 | 0.667 | 260.248 | 163.851 | 8.024 | 7.356 | 0.035 |
| Ccl25 | 1.427 | 28,348.999 | 10,540.968 | 14.791 | 13.364 | 8.92 × 10−7 |
| Ciita | 0.828 | 1867.537 | 1052.728 | 10.867 | 10.040 | 2.27 × 10−5 |
| Cmklr1 | 0.848 | 653.655 | 363.705 | 9.352 | 8.507 | 0.0001 |
| Cx3cl1 | 1.156 | 708.182 | 317.965 | 9.468 | 8.313 | 0.0020 |
| Cxcl11 | 0.883 | 1184.140 | 641.946 | 10.210 | 9.326 | 0.0143 |
| Cxcl12 | 1.593 | 10,218.765 | 3386.550 | 13.319 | 11.726 | 3.87 × 10−7 |
| Cxcl13 | 1.242 | 1620.704 | 685.156 | 10.662 | 9.420 | 0.0014 |
| Il2 | 0.920 | 63.683 | 33.400 | 5.993 | 5.062 | 0.033 |
| Il7 | 0.966 | 2624.135 | 1343.731 | 11.358 | 10.392 | 0.0005 |
| Il21 | 0.912 | 30.857 | 16.548 | 4.948 | 4.049 | 0.038 |
| Il23a | 0.976 | 118.014 | 59.799 | 6.883 | 5.902 | 0.029 |
| Il27 | 0.727 | 82.518 | 49.879 | 6.367 | 5.640 | 0.039 |
| Il33 | 0.639 | 645.356 | 414.507 | 9.334 | 8.695 | 0.027 |
| Kitl | 1.077 | 566.866 | 268.273 | 9.147 | 8.068 | 0.0005 |
| Nos2 | 1.337 | 136.272 | 54.373 | 7.090 | 5.765 | 0.0043 |
| Pparg | 1.246 | 879.010 | 370.480 | 9.780 | 8.533 | 0.013 |
| Ccl2 | −1.081 | 949.215 | 2007.774 | 9.891 | 10.971 | 0.012 |
| Ccl3 | −0.844 | 1822.248 | 3270.595 | 10.832 | 11.675 | 0.009 |
| Ccr5 | −1.041 | 552.400 | 1135.583 | 9.110 | 10.149 | 2.65 × 10−5 |
| Cdkn1a | −0.639 | 2633.161 | 4101.501 | 11.363 | 12.002 | 0.017 |
| Cebpb | −0.746 | 12,316.158 | 20,655.910 | 13.588 | 14.334 | 0.005 |
| Csf3 | −2.767 | 14.791 | 98.700 | 3.887 | 6.625 | 8.20 × 10−6 |
| Cxcl1 | −1.694 | 1174.780 | 3801.410 | 10.198 | 11.892 | 0.007 |
| Cxcl2 | −1.723 | 719.590 | 2373.510 | 9.491 | 11.213 | 3.14 × 10−5 |
| Cxcl3 | −1.073 | 150.984 | 317.559 | 7.238 | 8.311 | 0.0028 |
| Il1rl1 | −1.021 | 79.044 | 159.541 | 6.305 | 7.318 | 0.0003 |
| Il6 | −2.238 | 678.207 | 3198.631 | 9.406 | 11.643 | 0.0005 |
| Il17f | −1.119 | 49.106 | 106.128 | 5.618 | 6.730 | 0.0017 |
| Il18r1 | −0.608 | 1246.327 | 1898.444 | 10.283 | 10.891 | 0.01 |
| Rel | −0.691 | 3225.337 | 5204.208 | 11.655 | 12.345 | 0.0003 |
| Ptgs2 | −0.611 | 878.529 | 1340.712 | 9.779 | 10.389 | 0.005 |
| Spp1 | −1.142 | 343.025 | 757.012 | 8.422 | 9.564 | 0.0004 |
| Tnfsf11 | −0.669 | 2933.322 | 4664.196 | 11.518 | 12.187 | 0.042 |
| Thbs1 | −1.616 | 309.798 | 948.572 | 8.275 | 9.890 | 0.0008 |
| Vegfa | −0.753 | 3633.337 | 6122.907 | 11.827 | 12.580 | 0.007 |
| Category and Term of Pathway | Count of Genes | p Value | Gene Expression | Fold Enrichment | p Value by Benjamini–Hochberg Method |
|---|---|---|---|---|---|
| GOTERM_ MF_DIRECT—GO:0005125~ cytokine activity | 25 | 7.15 × 10−30 | Decreased: Ccl2, Ccl3, Csf3, Cxcl1, Cxcl2, Cxcl3, Il17f, Il6, Spp1, Tnfsf11, Vegfa Increased: Bmp4, Bmp7, Ccl19, Ccl25, Cx3cl1, Cxcl12, Cxcl13, Il2, Il7, Il21, Il23a, Il27, Il33, Kitl | 31.10 | 1.75 × 10−27 |
| GOTERM_ BP_DIRECT—GO:0006954~ inflammatory response | 25 | 2.01 × 10−27 | Decreased: Ccl2, Ccl3, Ccr5, Cxcl1, Cxcl2, Cxcl3, Il1rl1, Il6, Il17f, Il18r1, Ptgs2, Rel, Thbs1 Increased: C3, Ccl19, Ccl25, Ciita, Cmklr1, Cx3cl1, Cxcl11, Cxcl13, Il23a, Il27, Nos2, Pparg | 20.20 | 2.81 × 10−24 |
| KEGG_PATHWAY—hsa04657: IL-17 signaling pathway | 9 | 7.94 × 10−8 | Decreased: Ccl2, Cebpb, Csf3, Cxcl1, Cxcl2, Cxcl3, Il6, Il17f, Ptgs2 | 15.40 | 2.16 × 10−6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stakišaitis, D.; Kapočius, L.; Kilimaitė, E.; Gečys, D.; Šlekienė, L.; Balnytė, I.; Palubinskienė, J.; Lesauskaitė, V. Preclinical Study in Mouse Thymus and Thymocytes: Effects of Treatment with a Combination of Sodium Dichloroacetate and Sodium Valproate on Infectious Inflammation Pathways. Pharmaceutics 2023, 15, 2715. https://doi.org/10.3390/pharmaceutics15122715
Stakišaitis D, Kapočius L, Kilimaitė E, Gečys D, Šlekienė L, Balnytė I, Palubinskienė J, Lesauskaitė V. Preclinical Study in Mouse Thymus and Thymocytes: Effects of Treatment with a Combination of Sodium Dichloroacetate and Sodium Valproate on Infectious Inflammation Pathways. Pharmaceutics. 2023; 15(12):2715. https://doi.org/10.3390/pharmaceutics15122715
Chicago/Turabian StyleStakišaitis, Donatas, Linas Kapočius, Evelina Kilimaitė, Dovydas Gečys, Lina Šlekienė, Ingrida Balnytė, Jolita Palubinskienė, and Vaiva Lesauskaitė. 2023. "Preclinical Study in Mouse Thymus and Thymocytes: Effects of Treatment with a Combination of Sodium Dichloroacetate and Sodium Valproate on Infectious Inflammation Pathways" Pharmaceutics 15, no. 12: 2715. https://doi.org/10.3390/pharmaceutics15122715