

Exosomes: Potential Next-Generation Nanocarriers for the Therapy of Inflammatory Diseases

Abstract
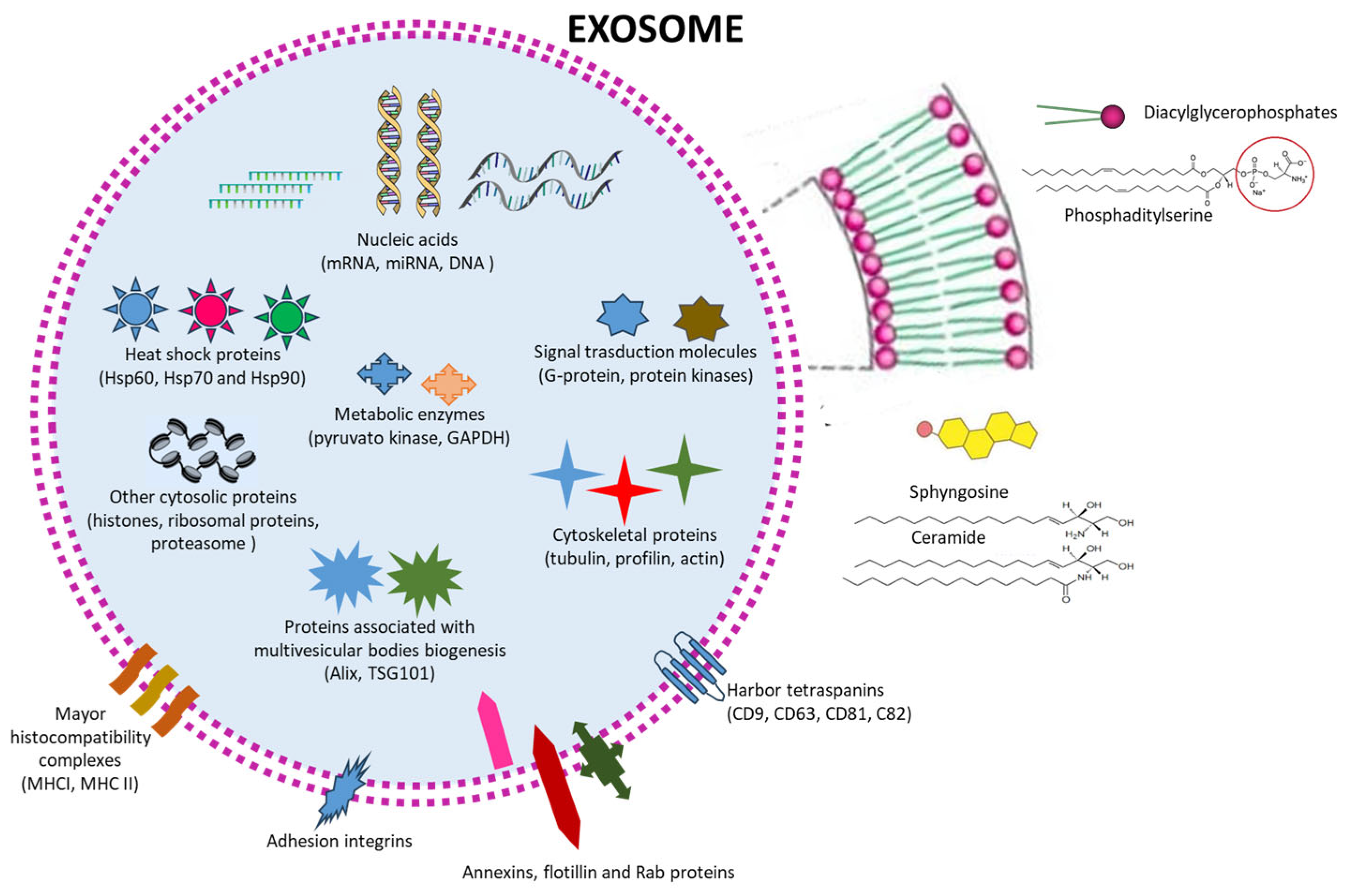
:1. Introduction
2. Classification and Characteristics of the Different EVs
3. Aim of the Review
4. Studies of EXs in Inflammatory Diseases
4.1. Neurological Disorders
4.1.1. Alzheimer’s Disease
4.1.2. Parkinson’s Disease
4.1.3. Ischemic Stroke
4.1.4. Multiple Sclerosis and Autoimmune Encephalomyelitis
4.1.5. Spinal Cord Injury
4.1.6. Traumatic Brain Injury
4.2. Rheumatoid Arthritis and Osteoarthritis
4.3. Lung Injuries
4.4. Liver Injury
4.5. Kidney Injuries
4.6. Intestinal Bowel Disease (IBD)
5. Discussion
6. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Halwani, A.A. Development of Pharmaceutical Nanomedicines: From the Bench to the Market. Pharmaceutics 2022, 14, 106. [Google Scholar] [CrossRef] [PubMed]
- Bilia, A.R.; Piazzini, V.; Risaliti, L.; Vanti, G.; Casamonti, M.; Wang, M.; Bergonzi, M.C. Nanocarriers: A Successful Tool to Increase Solubility, Stability and Optimise Bioefficacy of Natural Constituents. Curr. Med. Chem. 2019, 26, 4631–4656. [Google Scholar] [CrossRef] [PubMed]
- Brusini, R.; Varna, M.; Couvreur, P. Advanced nanomedicines for the treatment of inflammatory diseases. Adv. Drug Deliv. Rev. 2020, 157, 161–178. [Google Scholar] [CrossRef]
- Shan, X.; Gong, X.; Li, J.; Wen, J.; Li, Y.; Zhang, Z. Current approaches of nanomedicines in the market and various stage of clinical translation. Acta Pharm. Sin. B 2022, 12, 3028–3048. [Google Scholar] [CrossRef]
- Bilia, A.R.; Bergonzi, M.C.; Guccione, C.; Manconi, M.; Fadda, A.M.; Sinico, C. Vesicles and micelles: Two versatile vectors for the delivery of natural products. J. Drug Deliv. Sci. Technol. 2016, 32, 241–255. [Google Scholar] [CrossRef]
- Alshawwa, S.Z.; Kassem, A.A.; Farid, R.M.; Mostafa, S.K.; Labib, G.S. Nanocarrier Drug Delivery Systems: Characterization, Limitations, Future Perspectives and Implementation of Artificial Intelligence. Pharmaceutics 2022, 14, 883. [Google Scholar] [CrossRef]
- Herrmann, I.K.; Wood, M.J.A.; Fuhrmann, G. Extracellular vesicles as a next-generation drug delivery platform. Nat. Nanotechnol. 2021, 16, 748–759. [Google Scholar] [CrossRef]
- Busatto, S.; Morad, G.; Guo, P.; Moses, M.A. The role of extracellular vesicles in the physiological and pathological regulation of the blood-brain barrier. FASEB BioAdv. 2021, 3, 665–675. [Google Scholar] [CrossRef]
- Rezaie, J.; Feghhi, M.; Etemadi, T. A review on exosomes application in clinical trials: Perspective, questions, and challenges. Cell Commun. Signal 2022, 20, 145. [Google Scholar] [CrossRef]
- Zeng, H.; Guo, S.; Ren, X.; Wu, Z.; Liu, S.; Yao, X. Current Strategies for Exosome Cargo Loading and Targeting Delivery. Cells 2023, 12, 1416. [Google Scholar] [CrossRef]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef]
- Bonsergent, E.; Grisard, E.; Buchrieser, J.; Schwartz, O.; Théry, C.; Lavieu, G. Quantitative characterization of extracellular vesicle uptake and content delivery within mammalian cells. Nat. Commun. 2021, 12, 1864. [Google Scholar] [CrossRef] [PubMed]
- Battistelli, M.; Falcieri, E. Apoptotic Bodies: Particular Extracellular Vesicles Involved in Intercellular Communication. Biology 2020, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Lai, Y.; Hua, Z.C. Apoptosis and apoptotic body: Disease message and therapeutic target potentials. Biosci. Rep. 2019, 39, BSR20180992. [Google Scholar] [CrossRef] [PubMed]
- Clancy, J.W.; Schmidtmann, M.; D’Souza-Schorey, C. The ins and outs of microvesicles. FASEB BioAdv. 2021, 3, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.Y.; Kee, L.T.; Al-Masawa, M.E.; Lee, Q.H.; Subramaniam, T.; Kok, D.; Ng, M.H.; Law, J.X. Scalable Production of Extracellular Vesicles and Its Therapeutic Values: A Review. Int. J. Mol. Sci. 2022, 23, 7986. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Skryabin, G.O.; Komelkov, A.V.; Savelyeva, E.E.; Tchevkina, E.M. Lipid Rafts in Exosome Biogenesis. Biochem. Biokhimiia 2020, 85, 177–191. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, biologic function and clinical potential. Cell Biosci. 2019, 9, 19. [Google Scholar] [CrossRef]
- Gurung, S.; Perocheau, D.; Touramanidou, L.; Baruteau, J. The exosome journey: From biogenesis to uptake and intracellular signalling. Cell Commun. Signal 2021, 19, 47. [Google Scholar] [CrossRef]
- Gonda, A.; Kabagwira, J.; Senthil, G.N.; Wall, N.R. Internalization of Exosomes through Receptor-Mediated Endocytosis. Mol. Cancer Res. 2019, 17, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Gurunathan, S.; Kang, M.H.; Jeyaraj, M.; Qasim, M.; Kim, J.H. Review of the Isolation, Characterization, Biological Function, and Multifarious Therapeutic Approaches of Exosomes. Cells 2019, 8, 307. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://clinicaltrials.gov/search?cond=exosomes&aggFilters=status:com (accessed on 29 July 2023).
- Wei, W.; Ao, Q.; Wang, X.; Cao, Y.; Liu, Y.; Zheng, S.G.; Tian, X. Mesenchymal Stem Cell-Derived Exosomes: A Promising Biological Tool in Nanomedicine. Front. Pharmacol. 2021, 11, 590470. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.D.; Wong, W.Y.; Lee, M.M.; Cho, W.C.; Yee, B.K.; Kwan, Y.W.; Tai, W.C. Exosomes in Inflammation and Inflammatory Disease. Proteomics 2019, 19, e1800149. [Google Scholar] [CrossRef]
- Yu, B.; Zhang, X.; Li, X. Exosomes derived from mesenchymal stem cells. Int. J. Mol. Sci. 2014, 15, 4142–4157. [Google Scholar] [CrossRef]
- Leñero, C.; Kaplan, L.D.; Best, T.M.; Kouroupis, D. CD146+ Endometrial-Derived Mesenchymal Stem/Stromal Cell Subpopulation Possesses Exosomal Secretomes with Strong Immunomodulatory miRNA Attributes. Cells 2022, 11, 4002. [Google Scholar] [CrossRef]
- Kouroupis, D.; Kaplan, L.D.; Best, T.M. Human infrapatellar fat pad mesenchymal stem cells show immunomodulatory exosomal signatures. Sci. Rep. 2022, 12, 3609. [Google Scholar] [CrossRef]
- Gupta, A.; Pulliam, L. Exosomes as mediators of neuroinflammation. J. Neuroinflamm. 2014, 11, 68. [Google Scholar] [CrossRef]
- Wang, S.; Cesca, F.; Loers, G.; Schweizer, M.; Buck, F.; Benfenati, F.; Schachner, M.; Kleene, R. Synapsin I is an oligomannose-carrying glycoprotein, acts as an oligomannose-binding lectin, and promotes neurite outgrowth and neuronal survival when released via glia-derived exosomes. J. Neurosci. 2011, 31, 7275–7290. [Google Scholar] [CrossRef]
- Taylor, A.R.; Robinson, M.B.; Gifondorwa, D.J.; Tytell, M.; Milligan, C.E. Regulation of heat shock protein 70 release in astrocytes: Role of signaling kinases. Dev. Neurobiol. 2007, 67, 1815–1829. [Google Scholar] [CrossRef]
- Tytell, M. Release of heat shock proteins (Hsps) and the effects of extracellular Hsps on neural cells and tissues. Int. J. Hyperth. 2005, 21, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Verrilli, M.A.; Picou, F.; Court, F.A. Schwann cell-derived exosomes enhance axonal regeneration in the peripheral nervous system. Glia 2013, 61, 1795–1806. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Wang, Y.L.; Sun, H.F.; Wang, Z.Y.; Zhang, Q.; Fan, F.Y.; Ma, Y.C.; Liu, F.X.; Zhang, Y.K. Potential regulatory effects of stem cell exosomes on inflammatory response in ischemic stroke treatment. World J. Stem Cells 2023, 15, 561–575. [Google Scholar] [CrossRef]
- Perets, N.; Hertz, S.; London, M.; Offen, D. Intranasal administration of exosomes derived from mesenchymal stem cells ameliorates autistic-like behaviors of BTBR mice. Mol. Autism 2018, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.M.; Dennahy, I.S.; Bhatti, U.F.; Halaweish, I.; Xiong, Y.; Chang, P.; Nikolian, V.C.; Chtraklin, K.; Brown, J.; Zhang, Y.; et al. Mesenchymal Stem Cell-Derived Exosomes Provide Neuroprotection and Improve Long-Term Neurologic Outcomes in a Swine Model of Traumatic Brain Injury and Hemorrhagic Shock. J. Neurotrauma 2019, 36, 54–60. [Google Scholar] [CrossRef]
- Giovannelli, L.; Bari, E.; Jommi, C.; Tartara, F.; Armocida, D.; Garbossa, D.; Cofano, F.; Torre, M.L.; Segale, L. Mesenchymal stem cell secretome and extracellular vesicles for neurodegenerative diseases: Risk-benefit profile and next steps for the market access. Bioact. Mater. 2023, 29, 16–35. [Google Scholar] [CrossRef]
- Arabpour, M.; Saghazadeh, A.; Rezaei, N. Anti-inflammatory and M2 macrophage polarization-promoting effect of mesenchymal stem cell-derived exosomes. Int. Immunopharmacol. 2021, 97, 107823. [Google Scholar] [CrossRef]
- Andjus, P.; Kosanović, M.; Milićević, K.; Gautam, M.; Vainio, S.J.; Jagečić, D.; Kozlova, E.N.; Pivoriūnas, A.; Chachques, J.C.; Sakaj, M.; et al. Extracellular Vesicles as Innovative Tool for Diagnosis, Regeneration and Protection against Neurological Damage. Int. J. Mol. Sci. 2020, 21, 6859. [Google Scholar] [CrossRef]
- Zhang, Z.G.; Buller, B.; Chopp, M. Exosomes—Beyond stem cells for restorative therapy in stroke and neurological injury. Nat. Rev. Neurol. 2019, 15, 193–203. [Google Scholar] [CrossRef]
- Zhai, Y.; Wang, Q.; Zhu, Z.; Hao, Y.; Han, F.; Hong, J.; Zheng, W.; Ma, S.; Yang, L.; Cheng, G. High-efficiency brain-targeted intranasal delivery of BDNF mediated by engineered exosomes to promote remyelination. Biomater. Sci. 2022, 10, 5707–5718. [Google Scholar] [CrossRef]
- Upadhya, R.; Madhu, L.N.; Attaluri, S.; Gitaí, D.L.G.; Pinson, M.R.; Kodali, M.; Shetty, G.; Zanirati, G.; Kumar, S.; Shuai, B.; et al. Extracellular vesicles from human iPSC-derived neural stem cells: miRNA and protein signatures, and anti-inflammatory and neurogenic properties. J. Extracell. Vesicles 2020, 9, 1809064. [Google Scholar] [CrossRef]
- Upadhya, R.; Zingg, W.; Shetty, S.; Shetty, A.K. Astrocyte-derived extracellular vesicles: Neuroreparative properties and role in the pathogenesis of neurodegenerative disorders. J. Control Release 2020, 323, 225–239. [Google Scholar] [CrossRef]
- Guo, M.; Hao, Y.; Feng, Y.; Li, H.; Mao, Y.; Dong, Q.; Cui, M. Microglial Exosomes in Neurodegenerative Disease. Front. Mol. Neurosci. 2021, 14, 630808. [Google Scholar] [CrossRef]
- Krämer-Albers, E.M.; Bretz, N.; Tenzer, S.; Winterstein, C.; Möbius, W.; Berger, H.; Nave, K.A.; Schild, H.; Trotter, J. Oligodendrocytes secrete exosomes containing major myelin and stress-protective proteins: Trophic support for axons? Proteomics Clin. Appl. 2007, 1, 1446–1461. [Google Scholar] [CrossRef]
- Shaimardanova, A.A.; Solovyeva, V.V.; Chulpanova, D.S.; James, V.; Kitaeva, K.V.; Rizvanov, A.A. Extracellular vesicles in the diagnosis and treatment of central nervous system diseases. Neural Regen. Res. 2020, 15, 586–596. [Google Scholar]
- Elia, C.A.; Losurdo, M.; Malosio, M.L.; Coco, S. Extracellular Vesicles from Mesenchymal Stem Cells Exert Pleiotropic Effects on Amyloid-β, Inflammation, and Regeneration: A Spark of Hope for Alzheimer’s Disease from Tiny Structures? BioEssays News Rev. Mol. Cell Dev. Biol. 2019, 41, e1800199. [Google Scholar] [CrossRef]
- de Godoy, M.A.; Saraiva, L.M.; de Carvalho, L.R.P.; Vasconcelos-Dos-Santos, A.; Beiral, H.J.V.; Ramos, A.B.; Silva, L.R.P.; Leal, R.B.; Monteiro, V.H.S.; Braga, C.V.; et al. Mesenchymal stem cells and cell-derived extracellular vesicles protect hippocampal neurons from oxidative stress and synapse damage induced by amyloid-β oligomers. J. Biol. Chem. 2018, 293, 1957–1975. [Google Scholar] [CrossRef]
- Yuyama, K.; Sun, H.; Sakai, S.; Mitsutake, S.; Okada, M.; Tahara, H.; Furukawa, J.; Fujitani, N.; Shinohara, Y.; Igarashi, Y. Decreased amyloid-β pathologies by intracerebral loading of glycosphingolipid-enriched exosomes in Alzheimer model mice. J. Biol. Chem. 2014, 289, 24488–24498. [Google Scholar] [CrossRef]
- Cui, G.H.; Wu, J.; Mou, F.F.; Xie, W.H.; Wang, F.B.; Wang, Q.L.; Fang, J.; Xu, Y.W.; Dong, Y.R.; Liu, J.R.; et al. Exosomes derived from hypoxia-preconditioned mesenchymal stromal cells ameliorate cognitive decline by rescuing synaptic dysfunction and regulating inflammatory responses in APP/PS1 mice. FASEB J. 2018, 32, 654–668. [Google Scholar] [CrossRef]
- Yang, L.; Zhai, Y.; Hao, Y.; Zhu, Z.; Cheng, G. The Regulatory Functionality of Exosomes Derived from hUMSCs in 3D Culture for Alzheimer’s Disease Therapy. Small Weinh. Bergstr. Ger. 2020, 16, e1906273. [Google Scholar] [CrossRef]
- Cone, A.S.; Yuan, X.; Sun, L.; Duke, L.C.; Vreones, M.P.; Carrier, A.N.; Kenyon, S.M.; Carver, S.R.; Benthem, S.D.; Stimmell, A.C.; et al. Mesenchymal stem cell-derived extracellular vesicles ameliorate Alzheimer’s disease-like phenotypes in a preclinical mouse model. Theranostics 2021, 11, 8129–8142. [Google Scholar] [CrossRef]
- Liew, L.C.; Katsuda, T.; Gailhouste, L.; Nakagama, H.; Ochiya, T. Mesenchymal stem cell-derived extracellular vesicles: A glimmer of hope in treating Alzheimer’s disease. Int. Immunol. 2017, 29, 11–19. [Google Scholar] [CrossRef]
- Fatima, F.; Ekstrom, K.; Nazarenko, I.; Maugeri, M.; Valadi, H.; Hill, A.F.; Camussi, G.; Nawaz, M. Non-coding RNAs in Mesenchymal Stem Cell-Derived Extracellular Vesicles: Deciphering Regulatory Roles in Stem Cell Potency, Inflammatory Resolve, and Tissue Regeneration. Front. Genet. 2017, 8, 161. [Google Scholar] [CrossRef]
- Wang, S.S.; Jia, J.; Wang, Z. Mesenchymal Stem Cell-Derived Extracellular Vesicles Suppresses iNOS Expression and Ameliorates Neural Impairment in Alzheimer’s Disease Mice. J. Alzheimers Dis. 2018, 61, 1005–1013. [Google Scholar] [CrossRef]
- Haney, M.J.; Klyachko, N.L.; Zhao, Y.; Gupta, R.; Plotnikova, E.G.; He, Z.; Patel, T.; Piroyan, A.; Sokolsky, M.; Kabanov, A.V.; et al. Exosomes as drug delivery vehicles for Parkinson’s disease therapy. J. Control Release 2015, 207, 18–30. [Google Scholar] [CrossRef]
- Kojima, R.; Bojar, D.; Rizzi, G.; Hamri, G.C.; El-Baba, M.D.; Saxena, P.; Ausländer, S.; Tan, K.R.; Fussenegger, M. Designer exosomes produced by implanted cells intracerebrally deliver therapeutic cargo for Parkinson’s disease treatment. Nat. Commun. 2018, 9, 1305. [Google Scholar] [CrossRef]
- Peng, H.; Li, Y.; Ji, W.; Zhao, R.; Lu, Z.; Shen, J.; Wu, Y.; Wang, J.; Hao, Q.; Wang, J.; et al. Intranasal Administration of Self-Oriented Nanocarriers Based on Therapeutic Exosomes for Synergistic Treatment of Parkinson’s Disease. ACS Nano 2022, 16, 869–884. [Google Scholar] [CrossRef]
- Yang, H.; Tu, Z.; Yang, D.; Hu, M.; Zhou, L.; Li, Q.; Yu, B.; Hou, S. Exosomes from hypoxic pre-treated ADSCs attenuate acute ischemic stroke-induced brain injury via delivery of circ-Rps5 and promote M2 microglia/macrophage polarization. Neurosci. Lett. 2022, 769, 136389. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Sun, K.; Liu, Y.; Yin, X.; Zhu, H.; Yu, F.; Zhao, W. Resveratrol-loaded macrophage exosomes alleviate multiple sclerosis through targeting microglia. J. Control Release 2023, 353, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Riazifar, M.; Mohammadi, M.R.; Pone, E.J.; Yeri, A.; Lässer, C.; Segaliny, A.I.; McIntyre, L.L.; Shelke, G.V.; Hutchins, E.; Hamamoto, A.; et al. Stem Cell-Derived Exosomes as Nanotherapeutics for Autoimmune and Neurodegenerative Disorders. ACS Nano 2019, 13, 6670–6688. [Google Scholar] [CrossRef]
- Ma, K.; Xu, H.; Zhang, J.; Zhao, F.; Liang, H.; Sun, H.; Li, P.; Zhang, S.; Wang, R.; Chen, X. Insulin-like growth factor-1 enhances neuroprotective effects of neural stem cell exosomes after spinal cord injury via an miR-219a-2-3p/YY1 mechanism. Aging 2019, 11, 12278–12294. [Google Scholar] [CrossRef]
- Liu, W.; Rong, Y.; Wang, J.; Zhou, Z.; Ge, X.; Ji, C.; Jiang, D.; Gong, F.; Li, L.; Chen, J.; et al. Exosome-shuttled miR-216a-5p from hypoxic preconditioned mesenchymal stem cells repair traumatic spinal cord injury by shifting microglial M1/M2 polarization. J. Neuroinflamm. 2020, 17, 47. [Google Scholar] [CrossRef]
- Zhou, W.; Silva, M.; Feng, C.; Zhao, S.; Liu, L.; Li, S.; Zhong, J.; Zheng, W. Exosomes derived from human placental mesenchymal stem cells enhanced the recovery of spinal cord injury by activating endogenous neurogenesis. Stem Cell Res. Ther. 2021, 12, 174. [Google Scholar] [CrossRef]
- Zhang, W.; Hong, J.; Zhang, H.; Zheng, W.; Yang, Y. Astrocyte-derived exosomes protect hippocampal neurons after traumatic brain injury by suppressing mitochondrial oxidative stress and apoptosis. Aging 2021, 13, 21642–21658. [Google Scholar] [CrossRef]
- Wen, L.; Wang, Y.D.; Shen, D.F.; Zheng, P.D.; Tu, M.D.; You, W.D.; Zhu, Y.R.; Wang, H.; Feng, J.F.; Yang, X.F. Exosomes derived from bone marrow mesenchymal stem cells inhibit neuroinflammation after traumatic brain injury. Neural Regen. Res. 2022, 17, 2717–2724. [Google Scholar]
- Chen, Y.; Li, J.; Ma, B.; Li, N.; Wang, S.; Sun, Z.; Xue, C.; Han, Q.; Wei, J.; Zhao, R.C. MSC-derived exosomes promote recovery from traumatic brain injury via microglia/macrophages in rat. Aging 2020, 12, 18274–18296. [Google Scholar] [CrossRef]
- Wang, S.; Lei, B.; Zhang, E.; Gong, P.; Gu, J.; He, L.; Han, L.; Yuan, Z. Targeted Therapy for Inflammatory Diseases with Mesenchymal Stem Cells and Their Derived Exosomes: From Basic to Clinics. Int. J. Nanomed. 2022, 17, 1757–1781. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, H.; Xia, Y.; Yan, F.; Lu, Y. Therapeutic Potential of Mesenchymal Cell-Derived miRNA-150-5p-Expressing Exosomes in Rheumatoid Arthritis Mediated by the Modulation of MMP14 and VEGF. J. Immunol. 2018, 201, 2472–2482. [Google Scholar] [CrossRef]
- You, D.G.; Lim, G.T.; Kwon, S.; Um, W.; Oh, B.H.; Song, S.H.; Lee, J.; Jo, D.G.; Cho, Y.W.; Park, J.H. Metabolically engineered stem cell-derived exosomes to regulate macrophage heterogeneity in rheumatoid arthritis. Sci. Adv. 2021, 7, eabe0083. [Google Scholar] [CrossRef]
- Li, H.; Feng, Y.; Zheng, X.; Jia, M.; Mei, Z.; Wang, Y.; Zhang, Z.; Zhou, M.; Li, C. M2-type exosomes nanoparticles for rheumatoid arthritis therapy via macrophage re-polarization. J. Control. Release 2022, 341, 16–30. [Google Scholar] [CrossRef]
- Wu, L.F.; Zhang, Q.; Mo, X.B.; Lin, J.; Wu, Y.L.; Lu, X.; He, P.; Wu, J.; Guo, Y.F.; Wang, M.J.; et al. Identification of novel rheumatoid arthritis-associated MiRNA-204-5p from plasma exosomes. Exp. Mol. Med. 2022, 54, 334–345. [Google Scholar] [CrossRef]
- Li, S.; Stöckl, S.; Lukas, C.; Herrmann, M.; Brochhausen, C.; König, M.A.; Johnstone, B.; Grässel, S. Curcumin-primed human BMSC-derived extracellular vesicles reverse IL-1β-induced catabolic responses of OA chondrocytes by upregulating miR-126-3p. Stem Cell Res. Ther. 2021, 12, 252. [Google Scholar] [CrossRef] [PubMed]
- Xian Bo, S.; Chen, W.; Chang, L.; Hao Ran, Y.; Hui Hui, G.; Ya Kun, Z.; Wu Kun, X.; Hai Tao, F.; Wen Dan, C. The Research Progress of Exosomes in Osteoarthritis, with Particular Emphasis on the Therapeutic Effect. Front. Pharmacol. 2022, 13, 685623. [Google Scholar] [CrossRef]
- Dong, J.; Li, L.; Fang, X.; Zang, M. Exosome-Encapsulated microRNA-127-3p Released from Bone Marrow-Derived Mesenchymal Stem Cells Alleviates Osteoarthritis through Regulating CDH11-Mediated Wnt/β-Catenin Pathway. J. Pain. Res. 2021, 14, 297–310. [Google Scholar] [CrossRef]
- Mao, G.; Hu, S.; Zhang, Z.; Wu, P.; Zhao, X.; Lin, R.; Liao, W.; Kang, Y. Exosomal miR-95-5p regulates chondrogenesis and cartilage degradation via histone deacetylase 2/8. J. Cell Mol. Med. 2018, 22, 5354–5366. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Liu, G.; Wu, X. The umbilical cord mesenchymal stem cell-derived exosomal lncRNA H19 improves osteochondral activity through miR-29b-3p/FoxO3 axis. Clin. Transl. Med. 2021, 11, e255. [Google Scholar] [CrossRef]
- Qiu, M.; Liu, D.; Fu, Q. MiR-129-5p shuttled by human synovial mesenchymal stem cell-derived exosomes relieves IL-1β induced osteoarthritis via targeting HMGB1. Life Sci. 2021, 269, 118987. [Google Scholar] [CrossRef] [PubMed]
- Qi, H.; Liu, D.-P.; Xiao, D.-W.; Tian, D.-C.; Su, Y.-W.; Jin, S.-F. Exosomes derived from mesenchymal stem cells inhibit mitochondrial dysfunction-induced apoptosis of chondrocytes via p38, ERK, and Akt pathways. Vitr. Cell. Dev. Biol. Anim. 2019, 55, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yan, K.; Ge, G.; Zhang, D.; Bai, J.; Guo, X.; Zhou, J.; Xu, T.; Xu, M.; Long, X.; et al. Exosomes derived from miR-155-5p-overexpressing synovial mesenchymal stem cells prevent osteoarthritis via enhancing proliferation and migration, attenuating apoptosis, and modulating extracellular matrix secretion in chondrocytes. Cell Biol. Toxicol. 2021, 37, 85–96. [Google Scholar] [CrossRef]
- Cosenza, S.; Ruiz, M.; Toupet, K.; Jorgensen, C.; Noël, D. Mesenchymal stem cells derived exosomes and microparticles protect cartilage and bone from degradation in osteoarthritis. Sci. Rep. 2017, 7, 16214. [Google Scholar] [CrossRef]
- He, L.; He, T.; Xing, J.; Zhou, Q.; Fan, L.; Liu, C.; Chen, Y.; Wu, D.; Tian, Z.; Liu, B.; et al. Bone marrow mesenchymal stem cell-derived exosomes protect cartilage damage and relieve knee osteoarthritis pain in a rat model of osteoarthritis. Stem Cell Res. Ther. 2020, 11, 276. [Google Scholar] [CrossRef] [PubMed]
- Qiu, B.; Xu, X.; Yi, P.; Hao, Y. Curcumin reinforces MSC-derived exosomes in attenuating osteoarthritis via modulating the miR-124/NF-kB and miR-143/ROCK1/TLR9 signalling pathways. J. Cell Mol. Med. 2020, 24, 10855–10865. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, X.; Chen, J.; Qian, D.; Gao, P.; Qin, T.; Jiang, T.; Yi, J.; Xu, T.; Huang, Y.; et al. Exosomes derived from platelet-rich plasma administration in site mediate cartilage protection in subtalar osteoarthritis. J. Nanobiotechnol. 2022, 20, 56. [Google Scholar] [CrossRef]
- Zhang, S.; Chu, W.C.; Lai, R.C.; Lim, S.K.; Hui, J.H.P.; Toh, W.S. Exosomes derived from human embryonic mesenchymal stem cells promote osteochondral regeneration. Osteoarthr. Cartil. 2016, 24, 2135–2140. [Google Scholar] [CrossRef]
- Tao, S.-C.; Yuan, T.; Zhang, Y.-L.; Yin, W.-J.; Guo, S.-C.; Zhang, C.-Q. Exosomes derived from miR-140-5p-overexpressing human synovial mesenchymal stem cells enhance cartilage tissue regeneration and prevent osteoarthritis of the knee in a rat model. Theranostics 2017, 7, 180–195. [Google Scholar] [CrossRef]
- Qi, X.; Zhang, J.; Yuan, H.; Xu, Z.; Li, Q.; Niu, X.; Hu, B.; Wang, Y.; Li, X. Exosomes Secreted by Human-Induced Pluripotent Stem Cell-Derived Mesenchymal Stem Cells Repair Critical-Sized Bone Defects through Enhanced Angiogenesis and Osteogenesis in Osteoporotic Rats. Int. J. Biol. Sci. 2016, 12, 836–849. [Google Scholar] [CrossRef]
- Zhu, H.; Ji, J.; Fu, T.; Yang, J.; Gu, Z. The effect of exosomes from bone marrow mesenchymal stem cells on osteoarthritis. Ann. Rheum. Dis. 2018, 77, 893. [Google Scholar]
- Mortaz, E.; Alipoor, S.D.; Varahram, M.; Jamaati, H.; Garssen, J.; Mumby, S.E.; Adcock, I.M. Exosomes in Severe Asthma: Update in Their Roles and Potential in Therapy. BioMed Res. Int. 2018, 2018, 2862187. [Google Scholar] [CrossRef]
- Pei, W.; Li, X.; Bi, R.; Zhang, X.; Zhong, M.; Yang, H.; Zhang, Y.; Lv, K. Exosome membrane-modified M2 macrophages targeted nanomedicine: Treatment for allergic asthma. J. Control. Release 2021, 338, 253–267. [Google Scholar] [CrossRef]
- Liu, C.; Xiao, K.; Xie, L. Advances in the use of exosomes for the treatment of ALI/ARDS. Front. Immunol. 2022, 13, 971189. [Google Scholar] [CrossRef]
- Wei, X.; Yi, X.; Lv, H.; Sui, X.; Lu, P.; Li, L.; An, Y.; Yang, Y.; Yi, H.; Chen, G. Microrna-377-3p released by mesenchymal stem cell exosomes ameliorates lipopolysaccharide-induced acute lung injury by targeting RPTOR to induce autophagy. Cell Death Dis. 2020, 11, 657. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, P.; Goodwin, A.J.; Cook, J.A.; Halushka, P.V.; Chang, E.; Zingarelli, B.; Fan, H. Exosomes from endothelial progenitor cells improve outcomes of the lipopolysaccharide-induced acute lung injury. Crit. Care 2019, 23, 44. [Google Scholar] [CrossRef]
- Wu, X.; Liu, Z.; Hu, L.; Gu, W.; Zhu, L. Exosomes derived from endothelial progenitor cells ameliorate acute lung injury by transferring mir-126. Exp. Cell Res. 2018, 370, 13–23. [Google Scholar] [CrossRef]
- Quan, Y.; Wang, Z.; Gong, L.; Peng, X.; Richard, M.A.; Zhang, J.; Fornage, M.; Alcorn, J.L.; Wang, D. Exosome mir-371b-5p promotes proliferation of lung alveolar progenitor type ii cells by using pten to orchestrate the Pi3k/Akt signaling. Stem Cell Res. Ther. 2017, 8, 138. [Google Scholar] [CrossRef]
- Jiang, L.; Ni, J.; Shen, G.; Xia, Z.; Zhang, L.; Xia, S.; Pan, S.; Qu, H.; Li, X. Upregulation of endothelial cell-derived exosomal microrna-125b-5p protects from sepsis-induced acute lung injury by inhibiting topoisomerase ii alpha. Inflamm. Res. 2021, 70, 205–216. [Google Scholar] [CrossRef]
- Yu, Q.; Wang, D.; Wen, X.; Tang, X.; Qi, D.; He, J.; Zhao, Y.; Deng, W.; Zhu, T. Adipose-derived exosomes protect the pulmonary endothelial barrier in ventilator-induced lung injury by inhibiting the Trpv4/Ca2+ signaling pathway. Am. J. Physiol. Lung Cell Mol. Physiol. 2020, 318, L723–L741. [Google Scholar] [CrossRef]
- Kim, G.; Lee, Y.; Ha, J.; Han, S.; Lee, M. Engineering exosomes for pulmonary delivery of peptides and drugs to inflammatory lung cells by inhalation. J. Control. Release 2021, 330, 684–695. [Google Scholar] [CrossRef]
- Zhang, Z.; Long, J.; Meng, L.; Yang, Z.; Zhang, S.; Zhang, L. Immune cells-derived exosomes: A promising strategy for COVID-19 treatment. Clin. Transl. Discov. 2022, 2, e138. [Google Scholar] [CrossRef]
- Mitrani, M.I.; Bellio, M.A.; Sagel, A.; Saylor, M.; Kapp, W.; VanOsdol, K.; Haskell, G.; Stewart, D.; Abdullah, Z.; Santos, I.; et al. Case report: Administration of amniotic fluid-derived nanoparticles in three severely ill COVID-19 patients. Front. Med. 2021, 8, 583842. [Google Scholar] [CrossRef]
- Sengupta, V.; Sengupta, S.; Lazo, A.; Woods, P.; Nolan, A.; Bremer, N. Exosomes Derived from Bone Marrow Mesenchymal Stem Cells as Treatment for Severe COVID-19. Stem Cells Dev. 2020, 29, 747–754. [Google Scholar] [CrossRef]
- Lightner, A.L.; Sengupta, V.; Qian, S.; Ransom, J.T.; Suzuki, S.; Park, D.J.; Melson, T.I.; Williams, B.P.; Walsh, J.J.; Awili, M. Bone Marrow Mesenchymal Stem Cell-Derived Extracellular Vesicle Infusion for the Treatment of Respiratory Failure from COVID-19: A Randomized, Placebo-Controlled Dosing Clinical Trial. Chest 2023, in press. [Google Scholar] [CrossRef] [PubMed]
- Lou, G.; Chen, Z.; Zheng, M.; Liu, Y. Mesenchymal stem cell-derived exosomes as a new therapeutic strategy for liver diseases. Exp. Mol. Med. 2017, 49, e346. [Google Scholar] [CrossRef] [PubMed]
- Bataller, R.; Brenner, D.A. Liver fibrosis. J. Clin. Investig. 2005, 115, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Shang, J.; Yang, Q.; Dai, Z.; Liang, Y.; Lai, C.; Feng, T.; Zhong, D.; Zou, H.; Sun, L.; et al. Exosomes derived from human adipose mesenchymal stem cells ameliorate hepatic fibrosis by inhibiting PI3K/Akt/mTOR pathway and remodeling choline metabolism. J. Nanobiotechnol. 2023, 21, 29. [Google Scholar] [CrossRef]
- Niknam, B.; Baghaei, K.; Mahmoud Hashemi, S.; Hatami, B.; Reza Zali, M.; Amani, D. Human Wharton’s jelly mesenchymal stem cells derived-exosomes enriched by miR-124 promote an anti-fibrotic response in an experimental model of liver fibrosis. Int. Immunopharmacol. 2023, 119, 110294. [Google Scholar] [CrossRef]
- Zhang, Q.; Piao, C.; Xu, J.; Wang, Y.; Liu, T.; Ma, H.; Wang, H. ADSCs-exo attenuates hepatic ischemia-reperfusion injury after hepatectomy by inhibiting endoplasmic reticulum stress and inflammation. J. Cell Physiol. 2023, 238, 659–669. [Google Scholar] [CrossRef]
- Park, S.H.; Lee, E.K.; Yim, J.; Lee, M.H.; Lee, H.; Lee, Y.S.; Seo, W. Exosomes: Nomenclature, Isolation, and Biological Roles in Liver Diseases. Biomol. Ther. 2023, 31, 253–263. [Google Scholar] [CrossRef]
- Malekpour, K.; Hazrati, A.; Soudi, S.; Roshangar, L.; Pourfathollah, A.A.; Ahmadi, M. Combinational administration of mesenchymal stem cell-derived exosomes and metformin reduces inflammatory responses in an in vitro model of insulin resistance in HepG2 cells. Heliyon 2023, 9, e15489. [Google Scholar] [CrossRef]
- Khosrojerdi, A.; Soudi, S.; Hosseini, A.Z.; Khaligh, S.G.; Hashemi, S.M. The combination of mesenchymal stem cell- and hepatocyte-derived exosomes, along with imipenem, ameliorates inflammatory responses and liver damage in a sepsis mouse model. Life Sci. 2023, 326, 121813. [Google Scholar] [CrossRef]
- Cai, J.; Tang, D.; Hao, X.; Liu, E.; Li, W.; Shi, J. Mesenchymal stem cell-derived exosome alleviates sepsis- associated acute liver injury by suppressing MALAT1 through microRNA-26a-5p: An innovative immunopharmacological intervention and therapeutic approach for sepsis. Front. Immunol. 2023, 14, 1157793. [Google Scholar] [CrossRef]
- Ellakany, A.R.; El Baz, H.; Shoheib, Z.S.; Elzallat, M.; Ashour, D.S.; Yassen, N.A. Stem cell-derived exosomes as a potential therapy for schistosomal hepatic fibrosis in experimental animals. Pathog. Glob. Health 2023, 30, 1–21. [Google Scholar] [CrossRef]
- Cao, Q.; Huang, C.; Chen, X.M.; Pollock, C.A. Mesenchymal Stem Cell-Derived Exosomes: Toward Cell-Free Therapeutic Strategies in Chronic Kidney Disease. Front. Med. 2022, 9, 816656. [Google Scholar] [CrossRef]
- Charles, C.; Ferris, A.H. Chronic Kidney Disease. Prim. Care 2020, 47, 585–595. [Google Scholar] [CrossRef]
- Corrêa, R.R.; Juncosa, E.M.; Masereeuw, R.; Lindoso, R.S. Extracellular Vesicles as a Therapeutic Tool for Kidney Disease: Current Advances and Perspectives. Int. J. Mol. Sci. 2021, 22, 5787. [Google Scholar] [CrossRef]
- Nagaishi, K.; Mizue, Y.; Chikenji, T.; Otani, M.; Nakano, M.; Konari, N.; Fujimiya, M. Mesenchymal stem cell therapy ameliorates diabetic nephropathy via the paracrine effect of renal trophic factors including exosomes. Sci. Rep. 2016, 6, 34842. [Google Scholar] [CrossRef]
- Ebrahim, N.; Ahmed, I.A.; Hussien, N.I.; Dessouky, A.A.; Farid, A.S.; Elshazly, A.M.; Mostafa, O.; Gazzar, W.B.E.; Sorour, S.M.; Seleem, Y.; et al. Mesenchymal stem cell-derived exosomes ameliorated diabetic nephropathy by autophagy induction through the mTOR signaling pathway. Cells 2018, 7, 226. [Google Scholar] [CrossRef]
- Jin, J.; Shi, Y.; Gong, J.; Zhao, L.; Li, Y.; He, Q.; Huang, H. Exosome secreted from adipose-derived stem cells attenuates diabetic nephropathy by promoting autophagy flux and inhibiting apoptosis in podocyte. Stem Cell Res. Ther. 2019, 10, 95. [Google Scholar] [CrossRef]
- Liu, B.; Hu, D.; Zhou, Y.; Yu, Y.; Shen, L.; Long, C.; Butnaru, D.; Timashev, P.; He, D.; Lin, T.; et al. Exosomes released by human umbilical cord mesenchymal stem cells protect against renal interstitial fibrosis through ROS-mediated P38MAPK/ERK signaling pathway. Am. J. Transl. Res. 2020, 12, 4998–5014. [Google Scholar]
- Ishiy, C.; Ormanji, M.S.; Maquigussa, E.; Ribeiro, R.S.; da Silva Novaes, A.; Boim, M.A. Comparison of the effects of mesenchymal stem cells with their extracellular vesicles on the treatment of kidney damage induced by chronic renal artery stenosis. Stem Cells Int. 2020, 2020, 8814574. [Google Scholar] [CrossRef]
- Liu, L.; Wu, Y.; Wang, P.; Shi, M.; Wang, J.; Ma, H.; Sun, D. PSC-MSC-derived exosomes protect against kidney fibrosis in vivo and in vitro through the SIRT6/β-catenin signaling pathway. Int. J. Stem Cells 2021, 14, 310–319. [Google Scholar] [CrossRef]
- Hao, Y.; Miao, J.; Liu, W.; Cai, K.; Huang, X.; Peng, L. Mesenchymal stem cell-derived exosomes carry microRNA-125a to protect against diabetic nephropathy by targeting histone deacetylase 1 and downregulating endothelin-1. Diabetes Metab. Syndr. Obes. 2021, 14, 1405–1418. [Google Scholar] [CrossRef]
- Jiang, Z.Z.; Liu, Y.M.; Niu, X.; Yin, J.Y.; Hu, B.; Guo, S.C.; Fan, Y.; Wang, Y.; Wang, N.S. Exosomes secreted by human urine-derived stem cells could prevent kidney complications from type I diabetes in rats. Stem Cell Res. Ther. 2016, 7, 24. [Google Scholar] [CrossRef]
- Duan, Y.R.; Chen, B.P.; Chen, F.; Yang, S.X.; Zhu, C.Y.; Ma, Y.L. Exosomal microRNA-16-5p from human urine-derived stem cells ameliorates diabetic nephropathy through protection of podocyte. J. Cell Mol. Med. 2021, 25, 10798–10813. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Zuo, B.; Wang, Y.; Li, S.; Yang, J.; Sun, D. Protective function of exosomes from adipose tissue-derived mesenchymal stem cells in acute kidney injury through SIRT1 pathway. Life Sci. 2020, 255, 117719. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, Y.; Li, S.; Zuo, B.; Zhang, X.; Wang, F.; Sun, D. Exosomes derived from GDNF-modified human adipose mesenchymal stem cells ameliorate peritubular capillary loss in tubulointerstitial fibrosis by activating the SIRT1/eNOS signaling pathway. Theranostics 2020, 10, 9425–9442. [Google Scholar] [CrossRef] [PubMed]
- Crowley, E.; Muise, A. Inflammatory bowel disease: What very early onset disease teaches us. Gastroenterol. Clin. 2018, 47, 755–772. [Google Scholar] [CrossRef]
- Shapiro, J.M.; Subedi, S.; LeLeiko, N.S. Inflammatory bowel disease. Pediatr. Rev. 2016, 37, 337–347. [Google Scholar] [CrossRef]
- Ocansey, D.K.W.; Zhang, L.; Wang, Y.; Yan, Y.; Qian, H.; Zhang, X.; Xu, W.; Mao, F. Exosome-mediated effects and applications in inflammatory bowel disease. Biol. Rev. 2020, 95, 1287–1307. [Google Scholar] [CrossRef]
- Cai, X.; Zhang, Z.Y.; Yuan, J.T.; Ocansey, D.K.W.; Tu, Q.; Zhang, X.; Qian, H.; Xu, W.R.; Qiu, W.; Mao, F. hucMSC-derived exosomes colitis by regulating macrophage pyroptosis via the miR-378a-5p/ NLRP3 axis. Stem Cell Res. Ther. 2021, 12, 416. [Google Scholar] [CrossRef]
- Wu, X.; Xu, X.; Xiang, Y.; Fan, D.; An, Q.; Yue, G.; Jin, Z.; Ding, J.; Hu, Y.; Du, Q.; et al. Exosome-mediated effects and applications in inflammatory diseases of the digestive system. Eur. J. Med. Res. 2022, 27, 163. [Google Scholar] [CrossRef]
- Mao, F.; Wu, Y.; Tang, X.; Kang, J.; Zhang, B.; Yan, Y.; Qian, H.; Zhang, X.; Xu, W. Exosomes Derived from Human Umbilical Cord Mesenchymal Stem Cells Relieve Inflammatory Bowel Disease in Mice. Biomed. Res. Int. 2017, 2017, 5356760. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yu, Z.; Wan, S.; Wu, F.; Chen, W.; Zhang, B.; Lin, D.; Liu, J.; Xie, H.; Sun, X.; et al. Exosomes Derived from Dendritic Cells Treated with Schistosoma japonicum Soluble Egg Antigen Attenuate DSS-Induced Colitis. Front. Pharmacol. 2017, 8, 651. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Type of Disease/Animal Model | Exosome Origin (Eventual Drug Loaded) | Route of Administration | Effects | Reference |
|---|---|---|---|---|
| Neurological disorders/amyloid-β precursor protein transgenic mice | Neuroblastoma | Intracerebral injection | Marked reductions in Aβ levels and amyloid depositions. | [49] |
| Neurological disorders/transgenic APP/PS1 mice | MSC or hypoxia-preconditioned MSCs | Intravenously through lateral caudal vein | Both EXs, especially those deriving from the hypoxia preconditioned, improved learning and memory functions, reduced plaque deposition, and Aβ levels. | [50] |
| Neurological disorders/transgenic APP/PS1 mice | MSC | Intracerebroventricle injection | Reduction in exogenous Aβ-induced iNOS mRNA and protein expression; improved cognitive behavior. | [55] |
| Neurological disorders/PD model mice obtained with MPTP | MSC (EXs loaded with curcumin) | Intranasal administration | Reduction in α-synuclein aggregates, promoted neuron function recovery, significantly improved the movement and coordination ability. | [58] |
| Neurological disorders/middle cerebral artery occlusion mouse model | Hypoxic pre-treated, adipose-derived stem cells | Tail vein injection | Improvement of cognitive function by reducing neuronal damage in the hippocampus. | [59] |
| Neurological disorders/Experimental Autoimmune Encephalomyelitis | RAW (engineered EXs loaded with resveratrol) | Intranasal administration | Significant improvement of disease severity and significant decrease in multiple inflammatory cytokine responses in the CNS and periphery. Positive effect on the inflammatory infiltration and myelin recovery of MS. | [60] |
| Neurological disorders/Experimental Autoimmune Encephalomyelitis | MSCs (EXs stimulated by IFNγ) | Intravenous injections | Reduction in the mean clinical score and demyelination, decrease in neuroinflammation, and upregulation of CD4 + CD25 + FOXP3 + regulatory T cells. | [61] |
| Neurological disorders/spinal cord injury | Human placental MSCs | Intravenous injections | Significant improvement of the locomotor activity and bladder dysfunction; activation of proliferating endogenous neural stem/progenitor and a significative higher neurogenesis. | [62] |
| Neurological disorders/contusive spinal cord injury | MSCs under hypoxia | Tail vein injection | Promotion of functional behavioral recovery, shifting microglia from the M1 pro-inflammatory phenotype to the M2 anti-inflammatory phenotype by inhibiting TLR4/NF-κB and activating the PI3K/AKT signaling pathway. | [63] |
| Neurological disorders/spinal cord injury with complete transection of the thoracic segment | Human placental MSC | Intravenous injections | Enhanced anatomic and functional recovery; improvement of the locomotor activity and bladder dysfunction. | [64] |
| Neurological disorders/traumatic brain injury | Astrocyte | Tail vein injection | Mitigation of neurobehavioral deficits, cognitive impairment, and brain edema. Reduction in neuronal cell loss and atrophy, augmentation of antioxidant enzymes’ activities. | [65] |
| Neurological disorders/traumatic brain injury | Human adipose MSCs | Intracerebro-ventricular microinjection | Promotion of functional recovery, suppression of neuroinflammation, reduction in neuronal apoptosis, and increase in neurogenesis. | [67] |
| Rheumatoid arthritis and osteoarthritis/collagen-induced arthritis | Bone-marrow-derived MSCs | Intradermal injection | Reduction in hind paw thickness and decrease in clinical arthritis scores. Downregulation of MMP14 and VEGF expression, inhibition of angiogenesis. | [69] |
| Rheumatoid arthritis and osteoarthritis/collagen-induced arthritis | Adipose-derived stem cells (EXs with metabolic glycoengineered surface) | Intradermal injection | Induction of a cascade of anti-inflammatory events via macrophage phenotype regulation. High therapeutic efficacy, increase in T score, an indicator of bone density, and other clinical scores and photographs of inflamed joints. | [70] |
| Rheumatoid arthritis and osteoarthritis/collagen-induced arthritis | M2-type macrophages (EXs loaded with plasmid DNA encoding the anti-inflammatory cytokine interleukin-10) | Intradermal injection | Good accumulation at inflamed joint sites, high anti-inflammatory activity, and potent therapeutic effect. | [71] |
| Rheumatoid arthritis and osteoarthritis/osteoarthritis derived from injection of sodium iodoacetate | Bone marrow MSCs | Injection into the joint cavity | Effective promotion of cartilage repair and extracellular matrix synthesis, as well as alleviation of knee pain. | [82] |
| Rheumatoid arthritis and osteoarthritis/subtalar osteoarthritis by transecting anterior talofibular and calcaneal fibular ligament | Platelet-rich plasma (EXs were formulated in a thermosensitive hydrogel) | Injection in subtalar joint | Cartilage showed mild degeneration, and normal morphology and distribution of chondrocytes. | [84] |
| Rheumatoid arthritis and osteoarthritis/osteoarthritis model with critical-sized osteochondral defects from trochlear grooves of the distal femurs | MSCs derived from HuES9 human embryonic stem cells | Intra-articular injections | Complete restoration of cartilage and subchondral bone, including a hyaline cartilage with good surface regularity, complete bonding to adjacent cartilage, and extracellular matrix deposition. | [85] |
| Rheumatoid arthritis and osteoarthritis/osteoarthritis model from collagenase VII | Bone marrow MSCs | Injection | Inhibition of cartilage degradation and the progression of early osteoarthritis; prevention of the severe damage to knee articular cartilage. | [86] |
| Rheumatoid arthritis and osteoarthritis/osteoarthritis model with critical size bone defects in ovariectomized rats | MSCs from human induced pluripotent stem cells | Injection | Potent stimulation bone regeneration and angiogenesis. | [87] |
| Rheumatoid arthritis and osteoarthritis/osteoarthritis model by destabilization of the medial meniscus surgery | Bone marrow MSCs treated with decellularized extracellular matrix | Intra-articular injection | Better cartilage regeneration, improved anabolism and migration, and inhibiting chondrocyte apoptosis. | [88] |
| Lung injuries/Der f1-induced allergic asthma | M2 macrophages (EXs with coated PLGA@Dnmt3aossmart silencer) | Intravenous injection | Suppression of the development of asthma and marked reduction in inflammation, markedly decrease in the proportion of M2 macrophages and inflammatory cytokines. | [90] |
| Lung injuries/acute lung injury model with lipopolisaccharides | Human fetal lung fibroblast cells | Intratracheal instillation | Suppression of the bronchoalveolar lavage and inflammatory factors; induction of autophagy. | [92] |
| Lung injuries/acute lung injury model with lipopolisaccharides | Endothelial progenitor cell | Intratracheal instillation | Significant reduction in the cell number, protein concentration, and cytokines/chemokines in the bronchoalveolar lavage fluid. Reduction in myeloperoxidase activity, lung injury score, and pulmonary edema. | [93] |
| Lung injuries/acute lung injury model with lipopolisaccharides | Endothelial progenitor cell | Injection | Restoring pulmonary integrity, enhancing the proliferation, migration and tube formation of the endothelial cells. | [94] |
| Lung injuries/septic acute lung injury model via cecal ligation and perforation | Endothelial cells | Tracheal instillation | Promotion of VEGF expression, improvement of pathological changes, and control of lung water content, inflammatory response, protein content. | [96] |
| Lung injuries/ventilator-induced lung injury | Adipose-derived stem cell | Intravenous injection | Suppression of pulmonary endothelial barrier hyperpermeability, repair of the expression of adherens junctions, and alleviation of inflammatory response. | [97] |
| Lung injuries/acute lung injury model with lipopolisaccharides | Human Embryonic Kidney 293 cells (EXs engineered with RAGE-binding peptide, an anti-inflammatory peptide and loaded with curcumin) | Intratracheal instillation | Reduction in the cytokines in tissue lysate according to the levels of TNF-α and IL-1β in the bronchoalveolar lavage fluid. Reduction in hemolysis and infiltration of monocytes. | [98] |
| Liver injuries/carbon-tetrachloride-induced liver fibrosis | Human Wharton’s jelly MSCs (EXs enriched with miR-124-3p) | Intraperitoneal injection | Effective reduction in collagen accumulation and inhibition of inflammation. Phenotype switching of splenic monocytes from inflammatory Ly6Chi to restorative Ly6Clo. | [106] |
| Liver injuries/the portal vein and hepatic artery from 70% liver were occluded | Adipose-derived stem cells | Tail vein injection | Significant downregulation of TNF-α, interleukin-1β (IL-1β), and IL-6 levels; upregulation of IL-10 levels. In conclusion, ADSCs-exo protects against hepatic I/R injury after hepatectomy by inhibiting endoplasmic reticulum stress and inflammation. | [107] |
| Liver injuries/induced sepsis by cecal ligation and puncture | MSCs (EXs were co-administered with imipenem) | Imipenem injected subcutaneously plus EXs injected intravenously | Reduction in systemic and local inflammation; maintenance of all T lymphocyte populations, reduction in liver damage, and increase in the survival rate. | [110] |
| Liver injuries/induced sepsis by cecal ligation and puncture and lipopolysaccharide plus D-galactosamine | MSCs | Caudal vein injection | Replenishment of miR-26a-5p protected against hepatocyte death and liver injury caused by sepsis and inhibition of antioxidant system. | [111] |
| Liver injuries/schistosomal hepatic fibrosis | Bone marrow MSCs (EXs were co-administered with praziquantel) | Injection | Significant reduction in the number and diameter of hepatic granulomas, hepatic fibrosis; upregulation of proliferating cell nuclear antigen expression and reduction in NF-κB expression. Antifibrotic and anti-inflammatory effects. | [112] |
| Kidney injuries/diabetic nephropathy induced by streptozocin | MSCs | Injection | Marked improvement of renal function and restoration of renal tissues, with significant increase in LC3 and Beclin-1, and significant decrease in mTOR and fibrotic marker expression in renal tissue. | [117] |
| Kidney injuries/spontaneous diabetes | Adipose-derived stem cells | Tail intravenous injection | Attenuation of spontaneous diabetes by reduction in levels of urine protein, serum creatinine, blood urea nitrogen, and podocyte apoptosis. | [118] |
| Kidney injuries/renal fibrosis of irreversible model of unilateral ureteral obstruction | Human umbilical cord MSCs | Left renal artery injection after total ligation of the left ureter. | Significant decrease in the level of serum creatinine and blood urea nitrogen, of the level of apoptosis and oxidative stress. In addition, the renal tubular injury and tubulointerstitial. | [119] |
| Kidney injuries/renal chronic hypoxia induced by partial clamping of the left renal artery | Adipose-derived MSCs | Tail vein injection | Increase in stromal cell-derived factor-1-alpha (a cytokine) expression; reduction in the expression of hypoxia marker HIF1-α; stabilization of systolic blood pressure. Reduction in the expression of Col I and TGFβ and effective increase in the expression of the anti-inflammatory cytokine IL-10. | [120] |
| Kidney injuries/renal fibrosis from unilateral ureteral obstruction | Pluripotent stem-cell-derived MSCs | Tail vein injection | Reduction in the pathological process of renal fibrosis, of inflammatory reactions and improvement of renal function. Increasing the expression of SIRT6 and decreasing the expression of β-catenin and its downstream products. | [121] |
| Kidney injuries/diabetic nephropathy with streptozocin | Adipose-derived MSCs | Caudal vein injection | Decrease in levels of blood glucose, serum creatinine, 24 h urinary protein, urine albumin-to-creatinine ratio, and kidney weight/body weight. Suppression of mesangial hyperplasia and kidney fibrosis. | [122] |
| Kidney injuries/diabetic nephropathy with streptozocin | Urine-derived stem cells | Tail intravenous injection | Reduction in the urine volume and urinary microalbumin excretion, prevention of podocyte and tubular epithelial cell apoptosis, suppression of caspase-3 overexpression, and enhancement of the glomerular endothelial cell proliferation. | [123] |
| Kidney injuries/sepsis-induced acute kidney injury by cecal ligation and puncture | Adipose-derived MSCs | Tail vein injection | Activation of SIRT1, which reversed inflammation, apoptosis, and microcirculation disorders. In general, renal protective effects. Reduction in mortality. | [125] |
| Kidney injuries/renal fibrosis from unilateral ureteral obstruction | Glial-cell-line-derived neurotrophic factor transfected into human adipose MSCs via a lentiviral transfection system | Tail vein injection | Amelioration of renal fibrosis and significant decrease in peritubular capillary rarefaction Great capability of repair and angiogenesis. | [126] |
| Inflammatory bowel disease/colitis induced by dextran sulfate sodium | Human umbilical cord MSCs | Intravenous injection | Inhibition of NOD-like receptor family, pyrin domain-containing 3- inflammasomes of the colon. Suppression of the secretion of interleukin (IL)-18, IL-1β, and Caspase-1 cleavage, resulting in reduced cell pyroptosis. | [130] |
| Inflammatory bowel disease/colitis induced by dextran sulfate sodium | Human umbilical cord MSCs | Intravenous injection | Decrease in the severity of Inflammatory bowel disease. Increase in the expression of IL-10 gene and decrease in TNF-α, IL-1β, IL-6, iNOS, and IL-7 genes in colon tissues and spleens. Decreased of the infiltration of macrophages into the colon tissues. | [132] |
| Inflammatory bowel disease/acute colitis induced with dextran sulfate sodium | Dendritic cells treated with S. japonicum soluble eggs antigen | Intraperitoneal injection | Decrease in body weight loss and the disease activity index. Improvements of colon lengths and histological scores attenuating the severity of induced colitis in mice. | [133] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mori, T.; Giovannelli, L.; Bilia, A.R.; Margheri, F. Exosomes: Potential Next-Generation Nanocarriers for the Therapy of Inflammatory Diseases. Pharmaceutics 2023, 15, 2276. https://doi.org/10.3390/pharmaceutics15092276
Mori T, Giovannelli L, Bilia AR, Margheri F. Exosomes: Potential Next-Generation Nanocarriers for the Therapy of Inflammatory Diseases. Pharmaceutics. 2023; 15(9):2276. https://doi.org/10.3390/pharmaceutics15092276
Chicago/Turabian StyleMori, Tosca, Lisa Giovannelli, Anna Rita Bilia, and Francesca Margheri. 2023. "Exosomes: Potential Next-Generation Nanocarriers for the Therapy of Inflammatory Diseases" Pharmaceutics 15, no. 9: 2276. https://doi.org/10.3390/pharmaceutics15092276