
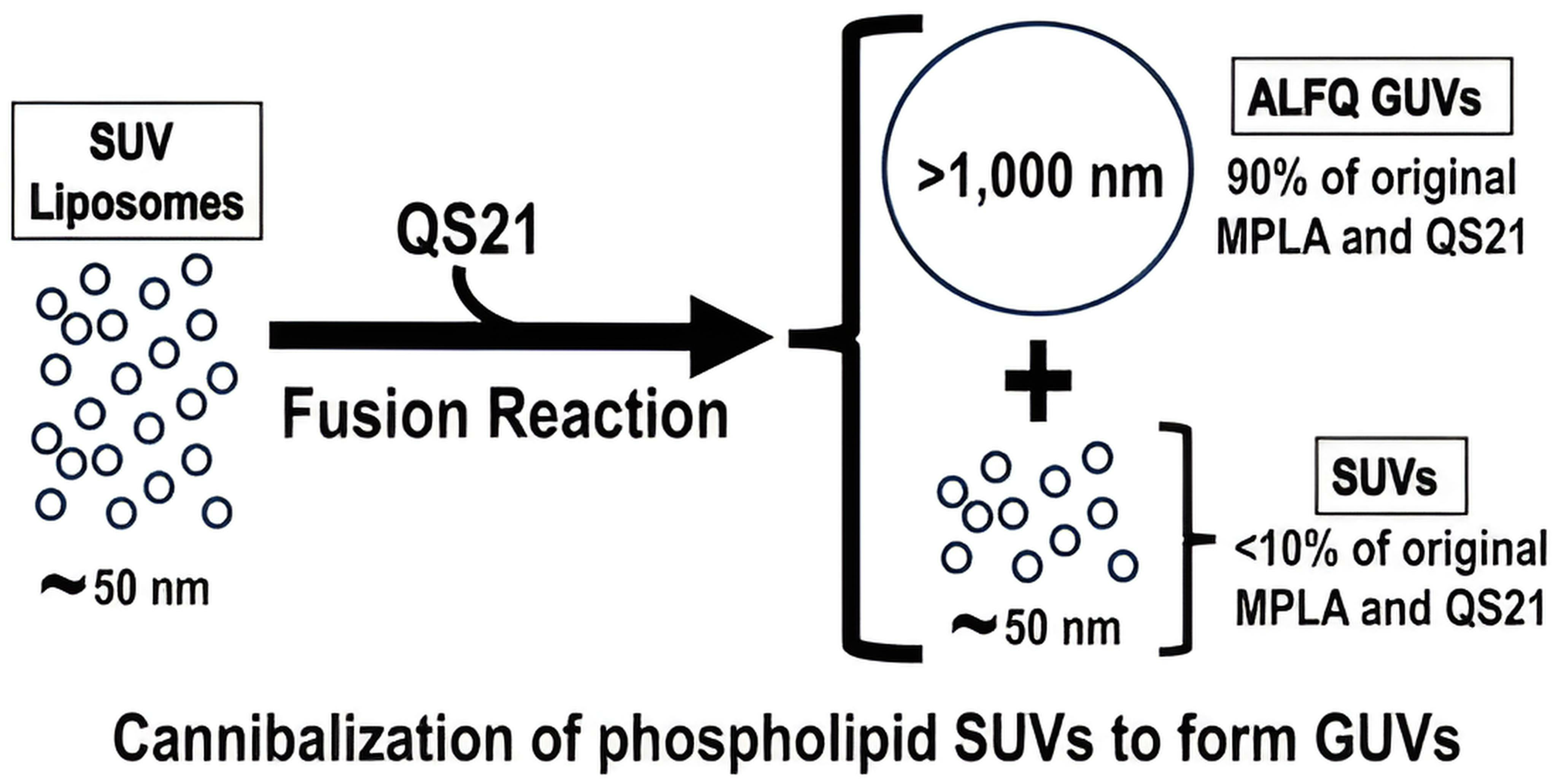
QS21-Initiated Fusion of Liposomal Small Unilamellar Vesicles to Form ALFQ Results in Concentration of Most of the Monophosphoryl Lipid A, QS21, and Cholesterol in Giant Unilamellar Vesicles

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. ALFQ
2.3. AS01-Like Liposomes
2.4. Purification of GUVs
2.5. Microscopy
2.6. Dynamic Light Scattering for Size and Zeta Potential
2.7. Quantitative Analysis of Lipid Components by UPLC–MS/MS
2.8. Data Analysis
3. Results
3.1. Separation of GUVs from SUVs and LUVs by Centrifugation
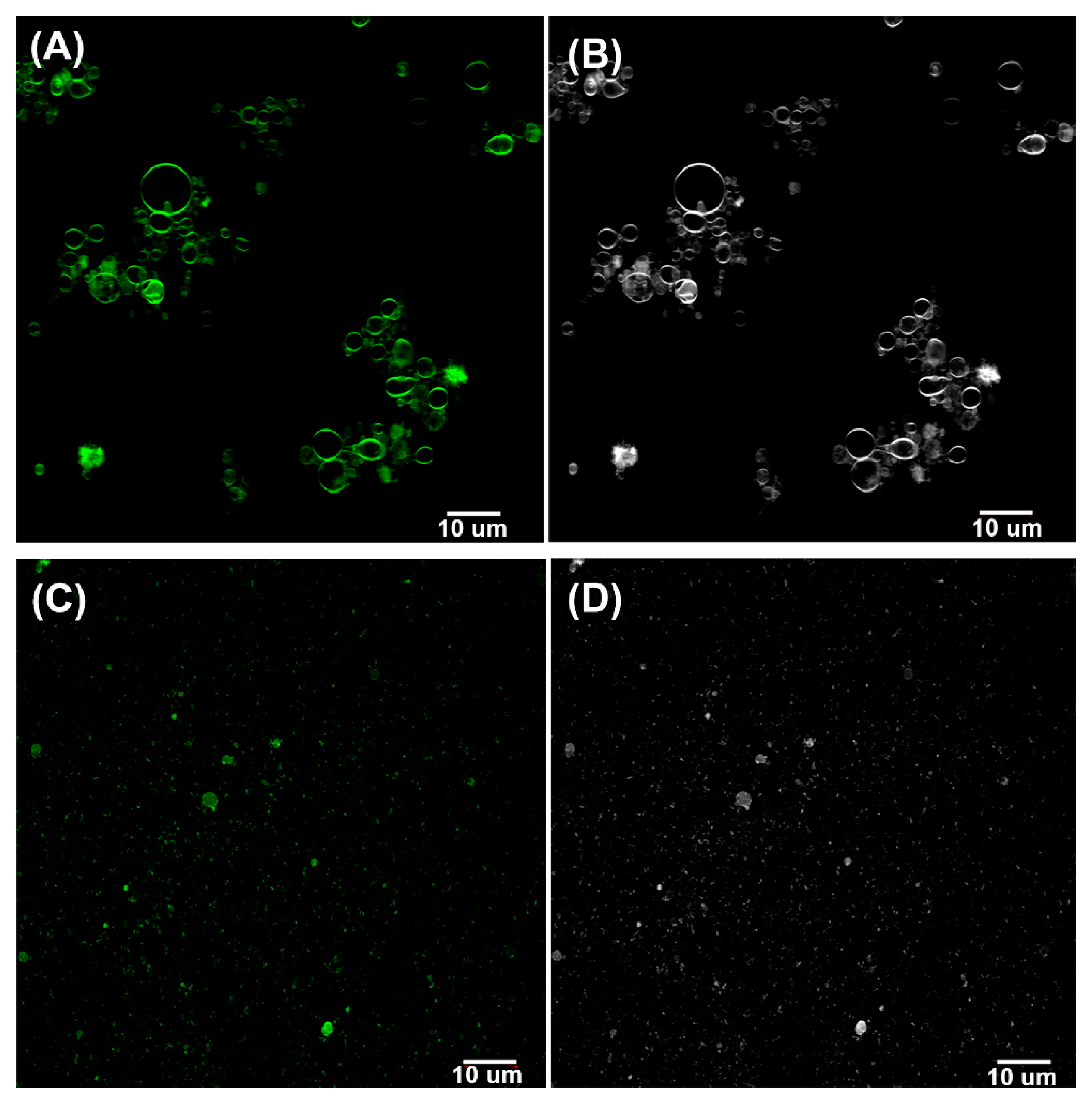
3.2. Fluorescence Analysis of GUVs
3.3. Total Lipid Recovery by Weight after Centrifugation
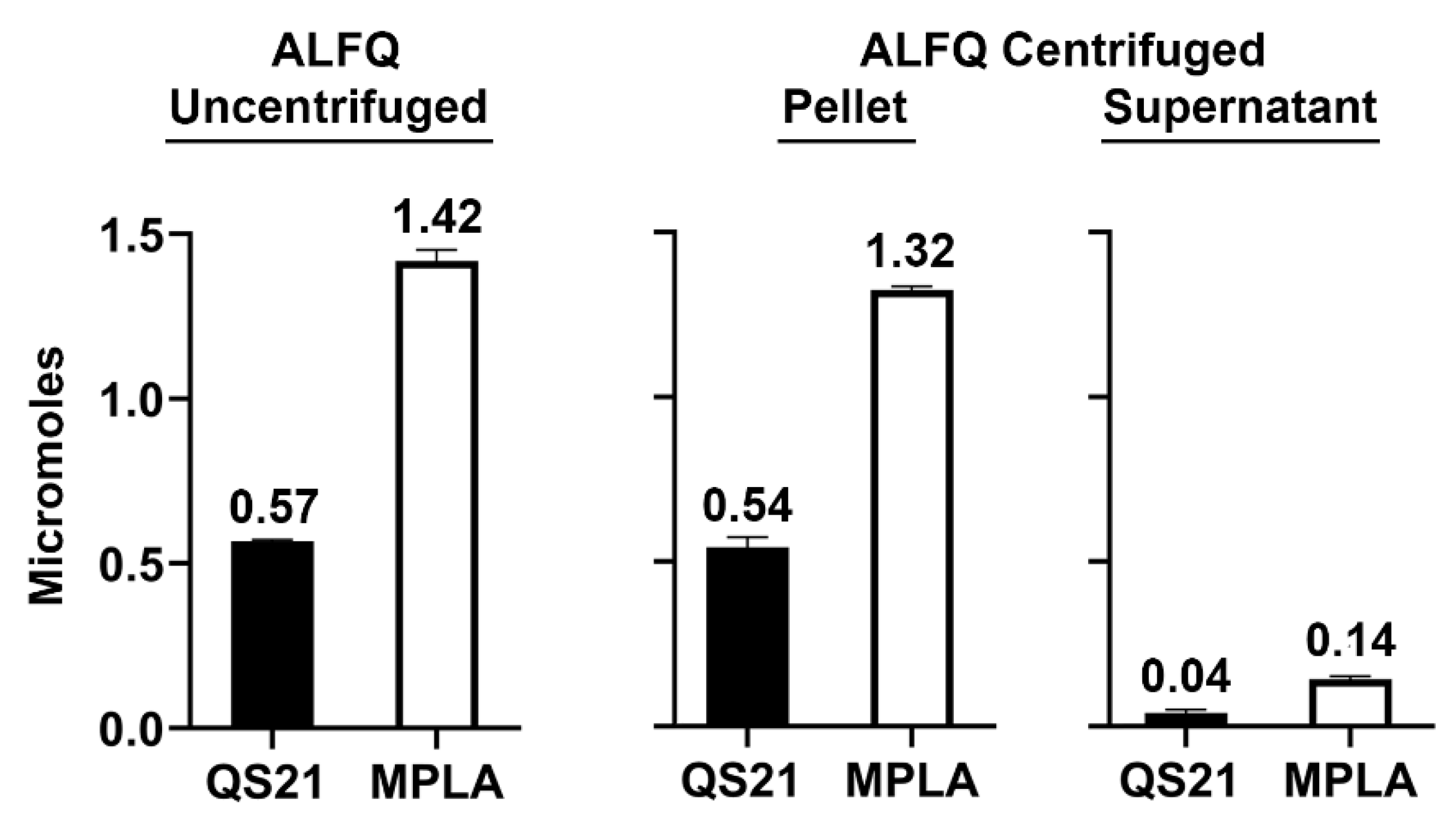
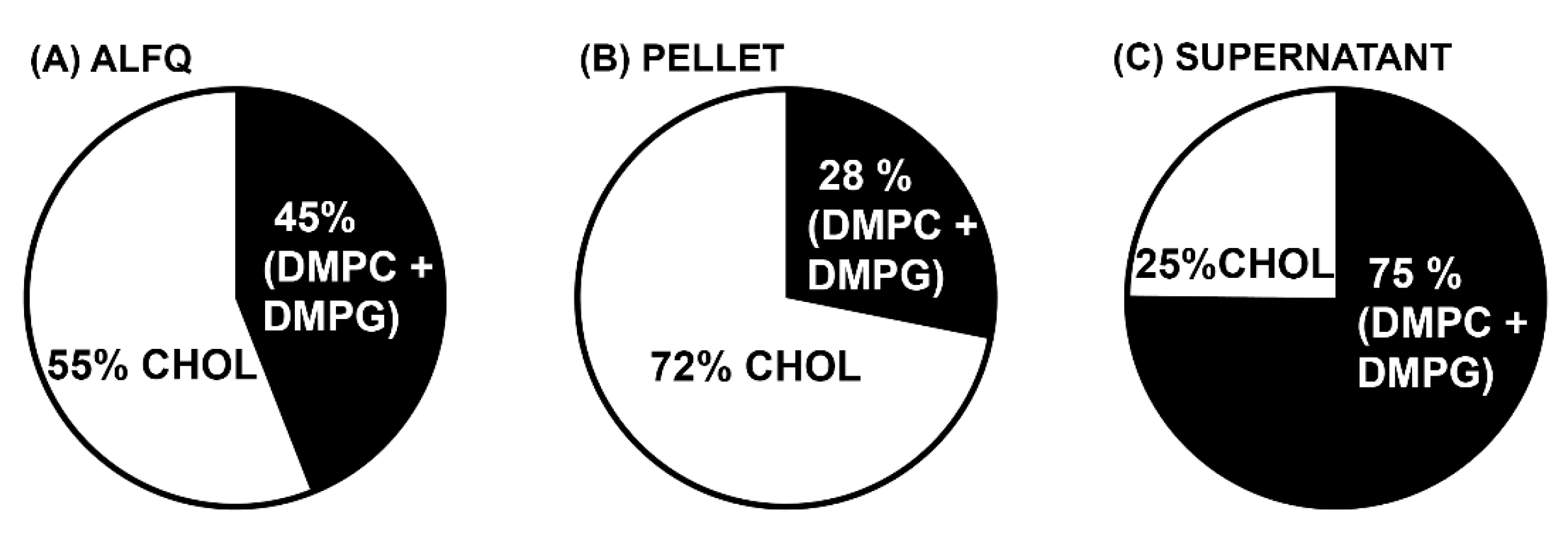
3.4. Molar Amounts of Individual Lipids in the Pellet and Supernatant after Centrifugation
3.5. Zeta Potentials of Particles
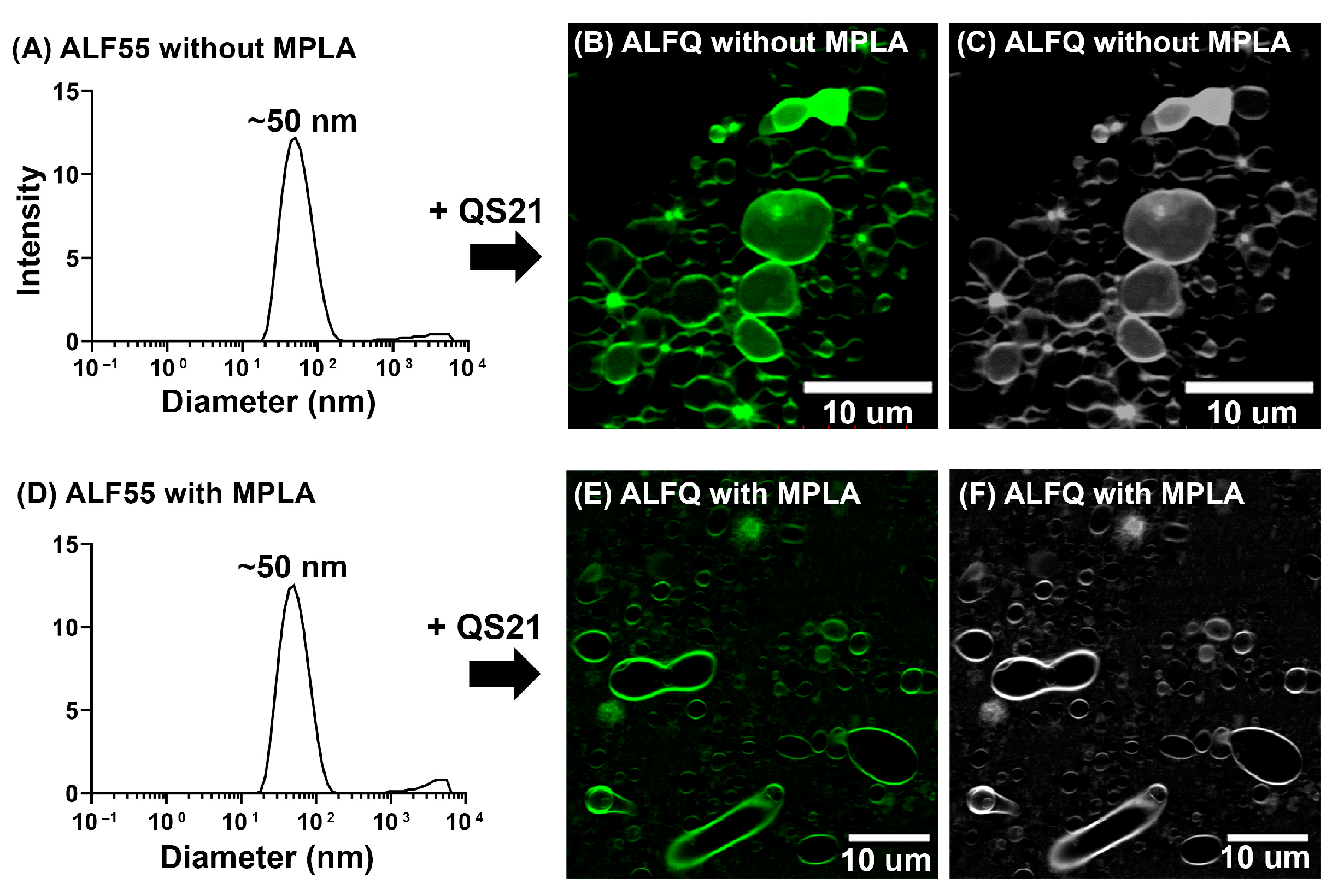
3.6. Is MPLA Required for GUV Formation?
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Mayer, L.D.; Bally, M.B.; Hope, M.J.; Cullis, P.R. Techniques for encapsulating bioactive agents into liposomes. Chem. Phys. Lipids 1986, 40, 333–345. [Google Scholar] [CrossRef]
- Szoka, F., Jr.; Papahadjopoulos, D. Procedure for preparation of liposomes with large internal aqueous space and high capture by reverse-phase evaporation. Proc. Natl. Acad. Sci. USA 1978, 75, 4194–4198. [Google Scholar] [CrossRef]
- Antimisiaris, S.G.; Jayasekera, P.; Gregoriadis, G. Liposomes as vaccine carriers Incorporation of soluble and particulate antigens in giant vesicles. J. Immunol. Methods 1993, 166, 271–280. [Google Scholar] [CrossRef]
- Henriksen-Lacey, M.; Korsholm, K.S.; Andersen, P.; Perrie, Y.; Christensen, D. Liposomal vaccine delivery systems. Expert Opin. Drug Deliv. 2011, 8, 505–519. [Google Scholar] [CrossRef]
- Reeves, J.P.; Dowben, R.M. Formation and properties of thin-walled phospholipid vesicles. J. Cell. Physiol. 1969, 73, 49–60. [Google Scholar] [CrossRef]
- Kim, S.; Martin, G.M. Preparation of cell-size unilamellar liposomes with high captured volume and defined size distribution. Biochem. Biophys. Acta 1981, 646, 1–9. [Google Scholar] [CrossRef]
- Lasic, D.D. The mechanism of vesicle formation. Biochem. J. 1988, 256, 1–11. [Google Scholar] [CrossRef]
- Akashi, K.; Miyata, H.; Itoh, H.; Kinosita, K., Jr. Formation of giant liposomes promoted by divalent cations: Critical role of electrostatic repulsion. Biophys. J. 1998, 74, 2973–2982. [Google Scholar] [CrossRef]
- Stein, H.; Spindler, S.; Bonakdar, N.; Wang, C.; Sandoghdar, V. Production of isolated giant unilamellar vesicles under high salt concentrations. Front. Physiol. 2017, 8, 63. [Google Scholar] [CrossRef]
- Knorr, R.L.; Steinkühler, J.; Dimova, R. Micron-sized domains in quasi single-component giant vesicles. Biochim. Biophys. Acta Biomembr. 2018, 1860, 1957–1964. [Google Scholar] [CrossRef]
- Omidvar, R.; Römer, W. Glycan-decorated protocells: Novel features for rebuilding cellular processes. Interface Focus 2019, 9, 20180084. [Google Scholar] [CrossRef]
- Van de Cauter, L.; Fanalista, F.; van Buren, L.; De Franceschi, N.; Godino, E.; Bouw, S.; Danelon, C.; Dekker, C.; Koenderink, G.H.; Ganzinger, K.A. Optimized cDICE for efficient reconstitution of biological systems in giant unilamellar vesicles. ACS Synth. Biol. 2021, 10, 1690–1702. [Google Scholar] [CrossRef]
- Kramer, K.; Sari, M.; Schulze, K.; Flegel, H.; Stehr, M.; Mey, I.; Janshoff, A.; Steinem, C. From LUVs to GUVs─How to cover micrometer-sized pores with membranes. J. Phys. Chem. B 2022, 126, 8233–8244. [Google Scholar] [CrossRef] [PubMed]
- Cavalcanti, R.R.M.; Lira, R.B.; Ewins, E.J.; Dimova, R.; Riske, K.A. Efficient liposome fusion to phase-separated giant vesicles. Biophys. J. 2022, 122, 1–13. [Google Scholar] [CrossRef]
- Parente, R.A.; Höchli, M.; Lentz, B.R. Morphology and phase behavior of two types of unilamellar vesicles prepared from synthetic phosphatidylcholines studied by freeze-fracture electron microscopy and calorimetry. Biochim. Biophys. Acta 1985, 812, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Litschel, T.; Schwille, P. Protein reconstitution inside giant unilamellar vesicles. Annu. Rev. Biophys. 2021, 50, 525–548. [Google Scholar] [CrossRef]
- Nair, K.S.; Bajaj, H. Advances in giant unilamellar vesicle preparation techniques and applications. Adv. Colloid Interface Sci. 2023, 318, 1–15. [Google Scholar] [CrossRef]
- Beck, Z.; Matyas, G.R.; Jalah, R.; Rao, M.; Polonis, V.R.; Alving, C.R. Differential immune responses to HIV-1 envelope protein induced by liposomal adjuvant formulations containing monophosphoryl lipid A with or without QS21. Vaccine 2015, 33, 5578–5587. [Google Scholar] [CrossRef]
- Alving, C.R.; Rao, M.; Matyas, G.R. Similarities and differences of chemical compositions and physical and functional properties of adjuvant system 01 and army liposome formulation with QS21. Front. Immunol. 2023, 14, 1102524. [Google Scholar] [CrossRef]
- A Trial for the Study of Falciparum Malaria Protein 014 Administered via Intramuscular Injection in Healthy Adults, clinicaltrials.gov ID NCT04296279, U.S. Army Medical Research and Development Command. Available online: https://www.clinicaltrials.gov/search?term=ALFQ%20vaccine%20adjuvant&cond=nct04296279 (accessed on 25 August 2023).
- A Trial for the Study of Falciparum Malaria Protein 013 Administered via Intramuscular Injection in Healthy Adults, clinicaltrials.gov ID NCT04268420, U.S. Army Medical Research and Development Command. Available online: https://www.clinicaltrials.gov/search?term=ALFQ%20vaccine%20adjuvant&cond=NCT04268420 (accessed on 25 August 2023).
- Safety, Tolerability, and Immunogenicity of ALFQ in a HIV Vaccine Containing A244 and B.65321 in Healthy Adults (RV575), clinicaltrials.gov Identifier NCT05423418, U.S. Army Medical Research and Development Command. Available online: https://www.clinicaltrials.gov/search?term=ALFQ%20vaccine%20adjuvant&cond=NCT05423418 (accessed on 25 August 2023).
- HIV Vaccine in HIV-uninfected Adults (RV546), clinicaltrials.gov ID NCT04658667, U.S. Army Medical Research and Development Command. Available online: https://www.clinicaltrials.gov/search?term=ALFQ%20vaccine%20adjuvant&cond=NCT04658667 (accessed on 25 August 2023).
- Safety and Efficacy of Neutralizing Antibodies and Vaccination for Induction of HIV Remission (RV582), clinicaltrials.gov ID NCT05769569, U.S. Army Medical Research and Development Command. Available online: https://www.clinicaltrials.gov/search?term=ALFQ%20vaccine%20adjuvant&cond=NCT05769569 (accessed on 25 August 2023).
- Safety and Immunogenicity of CJCV2 with and without ALFQ, clinicaltrials.gov ID NCT05500417, U.S. Army Medical Research and Development Command. Available online: https://www.clinicaltrials.gov/search?term=ALFQ%20vaccine%20adjuvant&cond=NCT05500417 (accessed on 25 August 2023).
- SARS-COV-2-Spike-Ferritin-Nanoparticle (SpFN) Vaccine with ALFQ Adjuvant for Prevention of COVID-19 in Healthy Adults, clinicaltrials.gov ID NCT04784767, U.S. Army Medical Research and Development Command. Available online: https://www.clinicaltrials.gov/search?term=ALFQ%20vaccine%20adjuvant&cond=NCT04784767 (accessed on 25 August 2023).
- Hutter, J.N.; Robben, P.M.; Lee, C.; Hamer, M.; Moon, J.E.; Merino, K.; Zhu, L.; Galli, H.; Quinn, X.; Brown, D.R.; et al. First-in-human assessment of safety and immunogenicity of low and high doses of Plasmodium falciparum malaria protein 013 (FMP013) administered intramuscularly with ALFQ adjuvant in healthy malaria-naïve adults. Vaccine 2022, 40, 5781–5790. [Google Scholar] [CrossRef] [PubMed]
- Matyas, G.R.; Muderhwa, J.M.; Alving, C.R. Oil-in-water liposomal emulsions for vaccine delivery. Methods Enzymol. 2003, 373, 34–50. [Google Scholar] [CrossRef]
- Singh, P.; Beck, Z.; Matyas, G.R.; Alving, C.R. Saturated phospholipids are required for nano- to micron-size transformation of cholesterol-containing liposomes upon QS21 addition. J. Liposome Res. 2019, 29, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Abucayon, E.G.; Barrientos, R.C.; Torres, O.B.; Sweeney, S.; Whalen, C.; Matyas, G.R. A liquid chromatography high-resolution tandem mass spectrometry method to quantify QS-21 adjuvant and its degradation products in liposomal drug formulations. ACS Omega 2023, 8, 21016–21025. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Tarrant, R.D.; Bolam, E.J.; Angell-Manning, P.; Soegaard, M.; Pattinson, D.J.; Dulal, P.; Silk, S.E.; Marshall, J.M.; Dabbs, R.A.; et al. Production, quality control, stability, and potency of cGMP-produced Plasmodium falciparum RH5.1 protein vaccine expressed in Drosophila S2 cells. NPJ Vaccines 2018, 3, 32. [Google Scholar] [CrossRef] [PubMed]
- Bangham, A.D.; Horne, R.W.; Glauert, A.M.; Dingle, J.T.; Lucy, J.A. Action of saponin on biological cell membranes. Nature 1962, 196, 952–955. [Google Scholar] [CrossRef]
- Seeman, P. Ultrastructure of membrane lesions in immune lysis, osmotic lysis and drug-induced lysis. Fed. Proc. 1974, 33, 2116–2124. [Google Scholar]
- Liu, J.; Harms, M.; Garamus, V.M.; Müller-Goymann, C.C. Reentrant structural phase transition in amphiphiphilic self-assembly. Soft Matter 2013, 9, 6371–6375. [Google Scholar] [CrossRef]
- Sarmento, M.J.; Hof, M.; Šachl, R. Interleaflet coupling of lipid nanodomains—Insights from in vitro systems. Front. Cell Dev. Biol. 2020, 8, 284. [Google Scholar] [CrossRef]
- Fatouros, D.G.; Klepetsanis, P.; Ioannou, P.V.; Antimisiaris, S.G. The effect of pH on the electrophoretic behaviour of a new class of liposomes: Arsonoliposomes. Int. J. Pharm. 2004, 288, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Berg, J.M.; Romoser, A.; Banerjee, N.; Zebda, R.; Sayes, C.M. The relationship between pH and zeta potential of 30 nm metal oxide nanoparticle suspensions relevant to in vitro toxicological evaluations. Nanotoxicology 2009, 3, 276–283. [Google Scholar] [CrossRef]
- Paepenmüller, T.; Müller-Goymann, C.C. Influence of Quil A on liposomal membranes. Int. J. Pharm. 2014, 475, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Okano, T.; Inoue, K.; Koseki, K.; Suzuki, H. Deformation modes of giant unilamellar vesicles encapsulating biopolymers. ACS Synth. Biol. 2018, 7, 739–747. [Google Scholar] [CrossRef] [PubMed]
- Boks, M.A.; Ambrosini, M.; Bruijns, S.C.; Kalay, H.; van Bloois, L.; Storm, G.; Garcia-Vallejo, J.J.; van Kooyk, Y. MPLA incorporation into DC-targeting glycoliposomes favours anti-tumour T cell responses. J. Control Release 2015, 216, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Manabe, N.; Yamaguchi, Y. 3D structural view of pathogen recognition by mammalian lectin receptors. Front. Mol. Biosci. 2021, 8, 670780. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample * | Weight (mg) | % Recovery |
|---|---|---|
| ALFQ | 139.63 | --- |
| Pellet | 88.29 | 63 |
| Supernatant | 34.60 | 25 |
| Component * | Uncentrifuged ALFQ | Centrifuged ALFQ | |||
|---|---|---|---|---|---|
| Pellet | % Recovery | Supernatant | % Recovery | ||
| DMPC | 71.18 ± 0.68 | 31.07 ± 0.27 | 44 | 25.98 ± 0.24 | 37 |
| DMPG | 8.06 ± 0.08 | 3.66 ± 0.01 | 45 | 2.87 ± 0.04 | 36 |
| DMPC + DMPG | 79.24 ± 0.68 | 34.73 ± 0.26 | 44 | 28.85 ± 0.27 | 36 |
| Cholesterol | 57.08 ± 0.45 | 50.44 ± 2.01 | 88 | 5.45 ± 0.02 | 10 |
| QS21 | 1.13 ± 0.01 | 1.08 ± 0.06 | 96 | 0.08 ± 0.02 | 7 |
| MPLA | 2.18 ± 0.05 | 2.04 ± 0.02 | 94 | 0.22 ± 0.01 | 10 |
| Liposomes | Zeta Potential (mV) |
|---|---|
| ALF55 | −10.50 ± 0.52 |
| ALFQ | −10.60 ± 0.62 |
| Pellet (GUV) | −11.43 ± 1.15 |
| Supernatant (SUV) | −8.38 ± 0.97 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abucayon, E.G.; Rao, M.; Matyas, G.R.; Alving, C.R. QS21-Initiated Fusion of Liposomal Small Unilamellar Vesicles to Form ALFQ Results in Concentration of Most of the Monophosphoryl Lipid A, QS21, and Cholesterol in Giant Unilamellar Vesicles. Pharmaceutics 2023, 15, 2212. https://doi.org/10.3390/pharmaceutics15092212
Abucayon EG, Rao M, Matyas GR, Alving CR. QS21-Initiated Fusion of Liposomal Small Unilamellar Vesicles to Form ALFQ Results in Concentration of Most of the Monophosphoryl Lipid A, QS21, and Cholesterol in Giant Unilamellar Vesicles. Pharmaceutics. 2023; 15(9):2212. https://doi.org/10.3390/pharmaceutics15092212
Chicago/Turabian StyleAbucayon, Erwin G., Mangala Rao, Gary R. Matyas, and Carl R. Alving. 2023. "QS21-Initiated Fusion of Liposomal Small Unilamellar Vesicles to Form ALFQ Results in Concentration of Most of the Monophosphoryl Lipid A, QS21, and Cholesterol in Giant Unilamellar Vesicles" Pharmaceutics 15, no. 9: 2212. https://doi.org/10.3390/pharmaceutics15092212