
Naïve or Engineered Extracellular Vesicles from Different Cell Sources: Therapeutic Tools for Kidney Diseases

 ,
,  and
and
Abstract
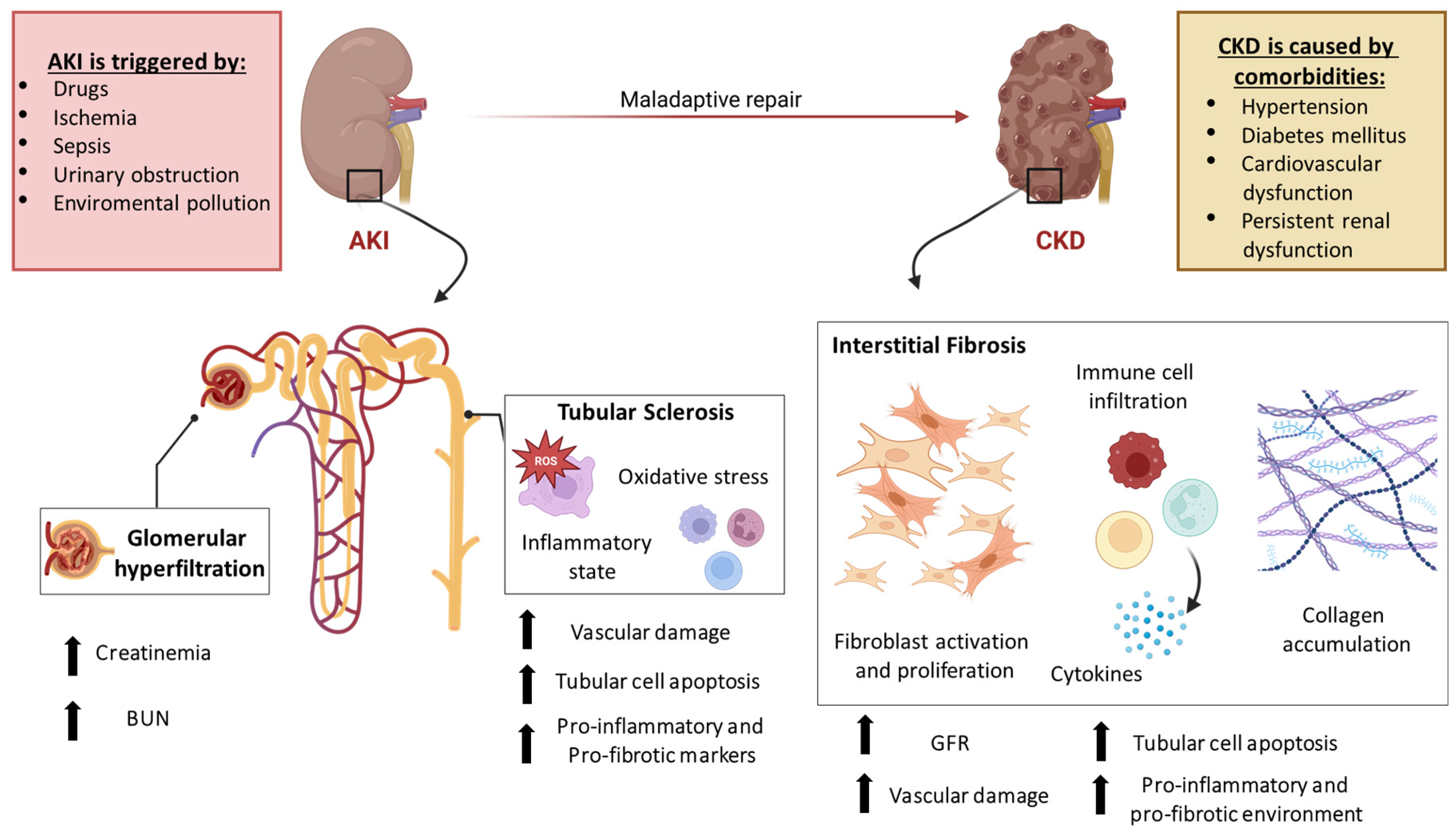
:1. Introduction
2. EVs of Different Origins for the Treatment of Kidney Diseases
3. Extracellular Vesicles Applications in AKI
3.1. MSC-Derived EVs in AKI

{kind=link}
{kind=link}
{kind=link}
| In Vivo Model | EV-Source | EV Isolation Method | EV Route of Administration | EV Biological Effect/ Mechanism of Action | References |
|---|---|---|---|---|---|
| Glycerol mouse model Intra-muscular glycerol injection into inferior hind limbs (8 mL/kg) | BM-MSCs (15 µg/dose) | UC | Single intravenous injection 3 days after the damage |
| [19] |
| Cisplatin mouse model Subcutaneous cisplatin injection (12 mg/kg) | BM-MSCs (100 µg/dose) | UC | Single injection 8 h after the damage |
| [20] |
| IRI rat model Ligation of the left renal artery and vein for 45 min with contralateral nephrectomy | BM-MSCs (30 μg/dose) | UC | Single intravenous injection immediately after surgery |
| [21] |
| IRI mouse model Renal pedicle clamping for 30 min | h-UC-MSCs (100 µg/dose) | UC | Two intravenous injections after IRI |
| [23] |
| IRI mouse model Renal pedicle clamping for 45 min | BM-MSCs (5 × 1010 EV/dose) | UC | Single tail intravenous injection 1 h before IRI |
| [24] |
| Cisplatin rat model Single intraperitoneal cisplatin injection (6 mg/kg) | UC-MSCs (200 µg/dose) | UC | Single injection in both the renal capsules 24 h damage |
| [25] |
| Sepsis mouse model Cecal ligation and puncture with CLP | UC-MSCs (120 μg/dose) | UC | Tail intravenous injection 3 h after CLP injury |
| [27] |
| IRI porcine model Unilateral ischemia for 120 min and contralateral nephrectomy | UC-MSCs (1 × 109 EV/dose) | UC and sequential 100 K Amicon centrifugal filter | Intravenous injection during the ischemia |
| [28] |
| IRI rat model Left renal ischemia for 60 min | UC-MSCs (100 μg EV/dose) | UC | Caudal intravenous injection immediately after reperfusion |
| [29] |
| Sepsis mouse model Cecal ligation and puncture with CLP | A-MSCs (100 μg EV/dose) | UC | Tail intravenous injection |
| [30] |
| LPS rat model Intraperitoneal injection of LPS (7.5 mg/kg) | BM-MSCs and A-MSCs (5 × 105 EV/dose) | UC | Single Injection 30 min before LPS injection |
| [31] |
| Unilateral IRI mouse model Renal artery clamping for 45 min | UC-MSCs (100 μg EV/dose) | UC | Caudal vein injection immediately after reperfusion |
| [32] |
| IRI mouse model Bilateral renal pedicle clamping for 30 min | BM-MSCs and UC-MSCs (6.96 × 1010 EV/dose) | UC | Single-tail intravenous injection |
| [33] |
| Glycerol mouse model Intramuscular glycerol injection into inferior hind limbs (8 mL/kg) | P-MSCs (80 µg EV/dose) | UC | Single tail intravenous injection 3 days after damage |
| [34] |
| Rat kidney DCD ex vivo model | BM-MSCs | UC | Ex vivo EV perfusion |
| [35] |
| IRI mouse model 35 min Left renal pedicle clamping for 35 min and contralateral nephrectomy | Gl-MSCs (400 × 106 EV/dose) | UC | Single tail intravenous the administration immediately after surgery |
| [36] |
3.2. Human Liver Stem Cell (HLSC)-Derived EVs in AKI
3.3. Non-MSC-Derived EVs in AKI
4. Extracellular Vesicles Application in CKD
4.1. MSC-Derived EVs in CKD
| In Vivo Model | EV-Source | EV Isolation Method | EV Route of Administration | EV Biological Effect/ Mechanism of Action | References |
|---|---|---|---|---|---|
| IRI rat model 45 min IRI and contralateral nephrectomy | h-BM-MSCs (30 µg EV/dose) | UC | Single intravenous injection immediately after IRI |
| [21] |
| Cisplatin mouse model Subcutaneous cisplatin injection (12 mg/kg) | BM-MSCs (100 µg EV/dose; 50 µg EV/dose) | UC | First (100 µg/dose) intravenous injection 8 h after the damage. Multiple (50 µg/dose) injections 2, 6, 10, 14, and 18 days after the damage |
| [20] |
| STZ-DN mouse model Intraperitoneal injection of STZ (37 mg/kg) for 4 consecutive days | h-BM-MSCs (1 × 1010 EV/dose) | UC | Multiple intravenous injections once a week for 4 weeks starting from day 30 after diabetes onset |
| [46] |
| AAN mouse model Intraperitoneal injection of 4 mg/kg of AA on a weekly basis for 4 weeks | h-BM-MSCs (1 × 1010 EV/dose) | UC | Multiple intravenous injections on a weekly basis starting from 3 days after AA administration |
| [49] |
| UUO rat model Ligation of the left ureter and the right the ureter was not ligated | Rat-BM-MSCs (3 × 105 EV/dose) | UC and exoEasy Maxi Kit (Qiagen, Germany) | Single tail vein injection 3 days before UUO surgery |
| [50] |
| UUO rat model Ligation of the left ureter and the right the ureter was not ligated | Rat-BM-MSCs (0.5 mg/kg EV/dose) | UC | Single intravenous injection 1 day after UUO surgery |
| [45] |
| UUO mouse model Ligation of the left ureter and the right the ureter was not ligated | m-BM-MSCs (30 mg EV/dose) | UC | Single caudal vein injection 2 days after the UUO surgery |
| [51] |
| UUO rat model Ligation of the left ureter and the right the ureter was not ligated | h-UC-MSCs (200 µg EV/dose) | UC | Not specified |
| [52] |
| PNx mouse model Resection of the upper and lower pole of the left kidney. Contralateral nephrectomy one week after | m-BM-MSCs (30 mg EV/dose) | UC | Multiple caudal vein injections 2, 3, and 5 days after the surgery |
| [54] |
| PNx rat model Resection of the upper and lower pole of the left kidney. Contralateral nephrectomy one week after | r-BM-MSCs (1 × 107 EV/dose) | UC | Multiple caudal vein injections on days 30 and 45 post surgery |
| [55] |
| DOCA rat model Nephrectomy of the left kidney. One week after the animals started a high-Na+ diet with subcutaneous DOCA | h-ASCs (1.5 × 109 EV/dose + DMSO 0.2 mg/~300 g) | UC | Weekly intravenous injection starting from 2 weeks after nephrectomy |
| [56] |
4.2. HLSC-Derived EVs in CKD
4.3. Non-MSC-Derived EVs in CKD
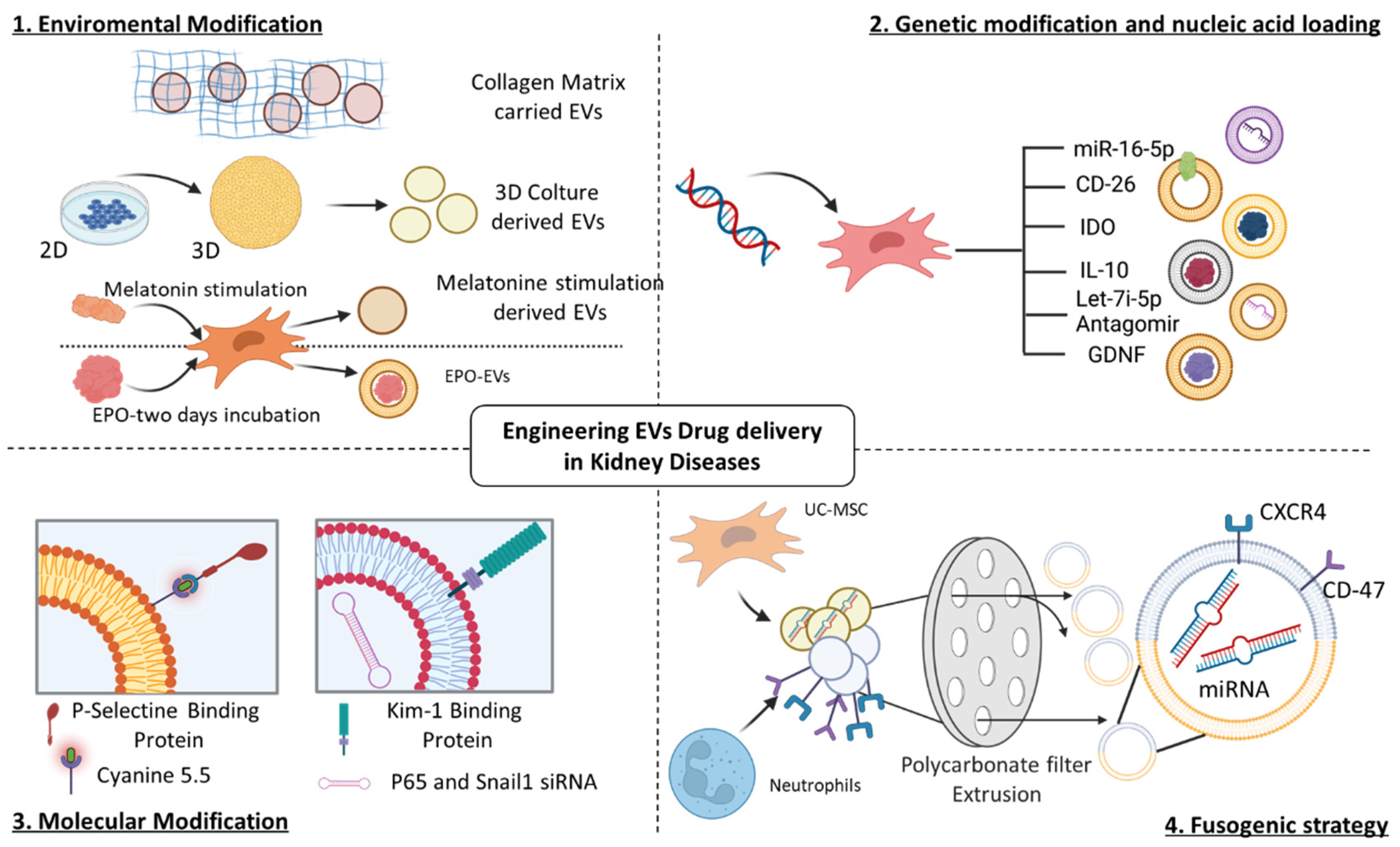
5. Engineering Strategies to Enhance the Capabilities of Native EVs
5.1. Engineering EVs for AKI Treatment
5.2. Engineering EVs for CKD Treatment
| In Vivo Model | EV-Source | EV Isolation Method | EV Route of Administration | EV Biological Effect | References |
|---|---|---|---|---|---|
| UUO mouse model Ligation of the left ureter and right ureter was not ligated | m-BM-MSCs (30 µg EV/dose) Tail intravenous injection 1 day after the surgery | UC | Two days of EPO incubation with m-BM-MSC and subsequent EPO-EVs isolation |
| [70] |
| UUO mouse model Ligation of the left ureter and right ureter was not ligated | h-BM-MSCs (50 µg EV/dose) Intravenous injection twice a week for 4 weeks | GETTM Exosome isolation kit | h-BM-MSC transfected with Let-7i-5p antagomir in presence of lipofectamine |
| [71] |
| UUO mouse model Ligation of the left ureter and right ureter was not ligated | h-A-MSCs (1000 µg EV/dose) Tail intravenous injection | UC | h-A-MSC lentiviral transfection with a vector plasmid system for GDNF gene |
| [73] |
| CKD mouse model Mice were fed with 0.25% adenine-containing diet | h-A-MSCs (50-100 µg EV/dose) Tail intravenous injection for 2 weeks | Exosome isolation kit | h-A-MSCs were treated with melatonin (1µM/mL) and subsequentially Exocue were isolated |
| [75] |
| STZ-DN rat model Intraperitoneal injection of STZ (65 mg/kg) | h-USCs (100 μg EV/dose) Multiple intravenous injections once a week for 12 consecutive weeks | Exo Quick-TC | Lentiviral transfection of h-USCs |
| [76] |
6. Future Perspectives
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Elsharkasy, O.M.; Nordin, J.Z.; Hagey, D.W.; De Jong, O.G.; Schiffelers, R.M.; Andaloussi, S.E.; Vader, P. Extracellular vesicles as drug delivery systems: Why and how? Adv. Drug Deliv. Rev. 2020, 159, 332–343. [Google Scholar] [CrossRef]
- Herrmann, I.K.; Wood, M.J.A.; Fuhrmann, G. Extracellular vesicles as a next-generation drug delivery platform. Nat. Nanotechnol. 2021, 16, 748–759. [Google Scholar] [CrossRef] [PubMed]
- Bunggulawa, E.J.; Wang, W.; Yin, T.; Wang, N.; Durkan, C.; Wang, Y.; Wang, G. Recent advancements in the use of exosomes as drug delivery systems. J. Nanobiotechnol. 2018, 16, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrêa, R.R.; Juncosa, E.M.; Masereeuw, R.; Lindoso, R.S. Extracellular Vesicles as a Therapeutic Tool for Kidney Disease: Current Advances and Perspectives. Int. J. Mol. Sci. 2021, 22, 5787. [Google Scholar] [CrossRef]
- Tang, T.-T.; Wang, B.; Lv, L.-L.; Dong, Z.; Liu, B.-C. Extracellular vesicles for renal therapeutics: State of the art and future perspective. J. Control. Release 2022, 349, 32–50. [Google Scholar] [CrossRef]
- Quaglia, M.; Merlotti, G.; Colombatto, A.; Bruno, S.; Stasi, A.; Franzin, R.; Castellano, G.; Grossini, E.; Fanelli, V.; Cantaluppi, V. Stem Cell-Derived Extracellular Vesicles as Potential Therapeutic Approach for Acute Kidney Injury. Front. Immunol. 2022, 13, 849891. [Google Scholar] [CrossRef]
- Gowen, A.; Shahjin, F.; Chand, S.; Odegaard, K.E.; Yelamanchili, S.V. Mesenchymal Stem Cell-Derived Extracellular Vesicles: Challenges in Clinical Applications. Front. Cell Dev. Biol. 2020, 8, 149. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Wang, L.; Zhang, M.; Chen, Z. Mesenchymal Stem Cell-Derived Small Extracellular Vesicles: A Novel Approach for Kidney Disease Treatment. Int. J. Nanomed. 2022, 17, 3603–3618. [Google Scholar] [CrossRef] [PubMed]
- Grange, C.; Skovronova, R.; Marabese, F.; Bussolati, B. Stem Cell-Derived Extracellular Vesicles and Kidney Regeneration. Cells 2019, 8, 1240. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Yang, L. Mesenchymal stem cells and extracellular vesicles in therapy against kidney diseases. Stem Cell Res. Ther. 2021, 12, 219. [Google Scholar] [CrossRef]
- Grange, C.; Bussolati, B. Extracellular vesicles in kidney disease. Nat. Rev. Nephrol. 2022, 18, 499–513. [Google Scholar] [CrossRef]
- Bento, G.; Shafigullina, A.K.; Rizvanov, A.A.; Sardão, V.A.; Macedo, M.P.; Oliveira, P.J. Urine-Derived Stem Cells: Applications in Regenerative and Predictive Medicine. Cells 2020, 9, 573. [Google Scholar] [CrossRef] [PubMed]
- Herrera, M.B.; Bruno, S.; Buttiglieri, S.; Tetta, C.; Gatti, S.; Deregibus, M.C.; Bussolati, B.; Camussi, G. Isolation and Characterization of a Stem Cell Population from Adult Human Liver. Stem Cells 2006, 24, 2840–2850. [Google Scholar] [CrossRef]
- Bruno, S.; Sanchez, M.B.H.; Chiabotto, G.; Fonsato, V.; Navarro-Tableros, V.; Pasquino, C.; Tapparo, M.; Camussi, G. Human Liver Stem Cells: A Liver-Derived Mesenchymal Stromal Cell-Like Population With Pro-regenerative Properties. Front. Cell Dev. Biol. 2021, 9, 644088. [Google Scholar] [CrossRef] [PubMed]
- Kosanović, M.; Milutinovic, B.; Glamočlija, S.; Morlans, I.M.; Ortiz, A.; Bozic, M. Extracellular Vesicles and Acute Kidney Injury: Potential Therapeutic Avenue for Renal Repair and Regeneration. Int. J. Mol. Sci. 2022, 23, 3792. [Google Scholar] [CrossRef]
- Bonventre, J.V.; Yang, L. Cellular pathophysiology of ischemic acute kidney injury. J. Clin. Investig. 2011, 121, 4210–4221. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-Y.; Kim, J.-S.; Jeong, K.-H.; Kim, S.-K. Acute Kidney Injury: Biomarker-Guided Diagnosis and Management. Medicina 2022, 58, 340. [Google Scholar] [CrossRef]
- Pickkers, P.; Darmon, M.; Hoste, E.; Joannidis, M.; Legrand, M.; Ostermann, M.; Prowle, J.R.; Schneider, A.; Schetz, M. Acute kidney injury in the critically ill: An updated review on pathophysiology and management. Intensive Care Med. 2021, 47, 835–850. [Google Scholar] [CrossRef]
- Bruno, S.; Grange, C.; Deregibus, M.C.; Calogero, R.A.; Saviozzi, S.; Collino, F.; Morando, L.; Busca, A.; Falda, M.; Bussolati, B.; et al. Mesenchymal stem cell-derived microvesicles protect against acute tubular injury. J. Am. Soc. Nephrol. 2009, 20, 1053–1067. [Google Scholar] [CrossRef] [Green Version]
- Bruno, S.; Grange, C.; Collino, F.; Deregibus, M.C.; Cantaluppi, V.; Biancone, L.; Tetta, C.; Camussi, G. Microvesicles derived from mesenchymal stem cells enhance survival in a lethal model of acute kidney injury. PLoS ONE 2012, 7, e33115. [Google Scholar] [CrossRef]
- Gatti, S.; Bruno, S.; Deregibus, M.C.; Sordi, A.; Cantaluppi, V.; Tetta, C.; Camussi, G. Microvesicles derived from human adult mesenchymal stem cells protect against ischaemia-reperfusion-induced acute and chronic kidney injury. Nephrol. Dial. Transplant. 2011, 26, 1474–1483. [Google Scholar] [CrossRef] [Green Version]
- Collino, F.; Bruno, S.; Incarnato, D.; Dettori, D.; Neri, F.; Provero, P.; Pomatto, M.; Oliviero, S.; Tetta, C.; Quesenberry, P.J.; et al. AKI recovery induced by mesenchymal stromal cell-derived extracellular vesicles carrying microRNAs. J. Am. Soc. Nephrol. 2015, 26, 2349–2360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.-Y.; Wang, B.; Tang, T.-T.; Wen, Y.; Li, Z.-L.; Feng, S.-T.; Wu, M.; Liu, D.; Yin, D.; Ma, K.-L.; et al. Exosomal miR-125b-5p deriving from mesenchymal stem cells promotes tubular repair by suppression of p53 in ischemic acute kidney injury. Theranostics 2021, 11, 5248–5266. [Google Scholar] [CrossRef]
- Zhu, G.; Pei, L.; Lin, F.; Yin, H.; Li, X.; He, W.; Liu, N.; Gou, X. Exosomes from human-bone-marrow-derived mesenchymal stem cells protect against renal ischemia/reperfusion injury via transferring miR-199a-3p. J. Cell. Physiol. 2019, 234, 23736–23749. [Google Scholar] [CrossRef]
- Zhou, Y.; Xu, H.; Xu, W.; Wang, B.; Wu, H.; Tao, Y.; Zhang, B.; Wang, M.; Mao, F.; Yan, Y.; et al. Exosomes released by human umbilical cord mesenchymal stem cells protect against cisplatin-induced renal oxidative stress and apoptosis in vivo and in vitro. Stem Cell Res. Ther. 2013, 4, 34. [Google Scholar] [CrossRef] [Green Version]
- Pomatto, M.; Gai, C.; Negro, F.; Cedrino, M.; Grange, C.; Ceccotti, E.; Togliatto, G.; Collino, F.; Tapparo, M.; Figliolini, F.; et al. Differential therapeutic effect of extracellular vesicles derived by bone marrow and adipose mesenchymal stem cells on wound healing of diabetic ulcers and correlation to their cargoes. Int. J. Mol. Sci. 2021, 22, 3851. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhu, Y.; Li, Y.; Liu, W.; Yin, L.; Yin, S.; Ji, C.; Hu, Y.; Wang, Q.; Zhou, X.; et al. Human umbilical cord mesenchymal stem cell exosomes alleviate sepsis-associated acute kidney injury via regulating microRNA-146b expression. Biotechnol. Lett. 2020, 42, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Cao, H.; Cui, B.; Ma, X.; Gao, L.; Yu, C.; Shen, F.; Yang, X.; Liu, N.; Qiu, A.; et al. Mesenchymal Stem Cells-Derived Exosomes Ameliorate Ischemia/Reperfusion Induced Acute Kidney Injury in a Porcine Model. Front. Cell Dev. Biol. 2022, 10, 899869. [Google Scholar] [CrossRef]
- Zou, X.; Zhang, G.; Cheng, Z.; Yin, D.; Du, T.; Ju, G.; Miao, S.; Liu, G.; Lu, M.; Zhu, Y. Microvesicles derived from human Wharton’s Jelly mesenchymal stromal cells ameliorate renal ischemia-reperfusion injury in rats by suppressing CX3CL1. Stem Cell Res. Ther. 2014, 5, 40. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Zuo, B.; Wang, Y.; Li, S.; Yang, J.; Sun, D. Protective function of exosomes from adipose tissue-derived mesenchymal stem cells in acute kidney injury through SIRT1 pathway. Life Sci. 2020, 255, 117719. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, J.; Huang, H. Exosomes from adipose-derived stem cells inhibit inflammation and oxidative stress in LPS-acute kidney injury. Exp. Cell Res. 2022, 420, 113332. [Google Scholar] [CrossRef]
- Zou, X.; Gu, D.; Xing, X.; Cheng, Z.; Gong, D.; Zhang, G.; Zhu, Y. Human mesenchymal stromal cell-derived extracellular vesicles alleviate renal ischemic reperfusion injury and enhance angiogenesis in rats. Am. J. Transl. Res. 2016, 8, 4289–4299. [Google Scholar] [PubMed]
- Zhao, M.; Liu, S.; Wang, C.; Wang, Y.; Wan, M.; Liu, F.; Gong, M.; Yuan, Y.; Chen, Y.; Cheng, J.; et al. Mesenchymal Stem Cell-Derived Extracellular Vesicles Attenuate Mitochondrial Damage and Inflammation by Stabilizing Mitochondrial DNA. ACS Nano 2021, 15, 1519–1538. [Google Scholar] [CrossRef]
- Cao, H.; Cheng, Y.; Gao, H.; Zhuang, J.; Zhang, W.; Bian, Q.; Wang, F.; Du, Y.; Li, Z.; Kong, D.; et al. In Vivo Tracking of Mesenchymal Stem Cell-Derived Extracellular Vesicles Improving Mitochondrial Function in Renal Ischemia-Reperfusion Injury. ACS Nano 2020, 14, 4014–4026. [Google Scholar] [CrossRef]
- Gregorini, M.; Corradetti, V.; Pattonieri, E.F.; Rocca, C.; Milanesi, S.; Peloso, A.; Canevari, S.; De Cecco, L.; Dugo, M.; Avanzini, M.A.; et al. Perfusion of isolated rat kidney with Mesenchymal Stromal Cells/Extracellular Vesicles prevents ischaemic injury. J. Cell. Mol. Med. 2017, 21, 3381–3393. [Google Scholar] [CrossRef] [PubMed]
- Ranghino, A.; Bruno, S.; Bussolati, B.; Moggio, A.; DiMuccio, V.; Tapparo, M.; Biancone, L.; Gontero, P.; Frea, B.; Camussi, G. The effects of glomerular and tubular renal progenitors and derived extracellular vesicles on recovery from acute kidney injury. Stem Cell Res. Ther. 2017, 8, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rampino, T.; Gregorini, M.; Germinario, G.; Pattonieri, E.F.; Erasmi, F.; Grignano, M.A.; Bruno, S.; Alomari, E.; Bettati, S.; Asti, A.; et al. Extracellular Vesicles Derived from Mesenchymal Stromal Cells Delivered during Hypothermic Oxygenated Machine Perfusion Repair Ischemic/Reperfusion Damage of Kidneys from Extended Criteria Donors. Biology 2022, 11, 350. [Google Scholar] [CrossRef]
- Sanchez, M.B.H.; Bruno, S.; Grange, C.; Tapparo, M.; Cantaluppi, V.; Tetta, C.; Camussi, G. Human liver stem cells and derived extracellular vesicles improve recovery in a murine model of acute kidney injury. Stem Cell Res. Ther. 2014, 5, 124. [Google Scholar] [CrossRef] [Green Version]
- Collino, F.; Lopes, J.A.; Tapparo, M.; Tortelote, G.G.; Kasai-Brunswick, T.H.; Lopes, G.M.; Almeida, D.B.; Skovronova, R.; Wendt, C.H.C.; de Miranda, K.R.; et al. Extracellular Vesicles Derived from Induced Pluripotent Stem Cells Promote Renoprotection in Acute Kidney Injury Model. Cells 2020, 9, 453. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liao, J.; Su, X.; Li, W.; Bi, Z.; Wang, J.; Su, Q.; Huang, H.; Wei, Y.; Gao, Y.; et al. Human urine-derived stem cells protect against renal ischemia/reperfusion injury in a rat model via exosomal miR-146a-5p which targets IRAK1. Theranostics 2020, 10, 9561–9578. [Google Scholar] [CrossRef]
- Sun, Z.; Wu, J.; Bi, Q.; Wang, W. Exosomal lncRNA TUG1 derived from human urine-derived stem cells attenuates renal ischemia/reperfusion injury by interacting with SRSF1 to regulate ASCL4-mediated ferroptosis. Stem Cell Res. Ther. 2022, 13, 297. [Google Scholar] [CrossRef]
- Grange, C.; Papadimitriou, E.; Dimuccio, V.; Pastorino, C.; Molina, J.; O’kelly, R.; Niedernhofer, L.J.; Robbins, P.D.; Camussi, G.; Bussolati, B. Urinary Extracellular Vesicles Carrying Klotho Improve the Recovery of Renal Function in an Acute Tubular Injury Model. Mol. Ther. 2020, 28, 490–502. [Google Scholar] [CrossRef] [Green Version]
- Tetta, C.; Deregibus, M.C.; Camussi, G. Stem cells and stem cell-derived extracellular vesicles in acute and chronic kidney diseases: Mechanisms of repair. Ann. Transl. Med. 2020, 8, 570. [Google Scholar] [CrossRef]
- Bruno, S.; Chiabotto, G.; Cedrino, M.; Ceccotti, E.; Pasquino, C.; De Rosa, S.; Grange, C.; Tritta, S.; Camussi, G. Extracellular Vesicles Derived from Human Liver Stem Cells Attenuate Chronic Kidney Disease Development in an In Vivo Experimental Model of Renal Ischemia and Reperfusion Injury. Int. J. Mol. Sci. 2022, 23, 1485. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Wang, Q.; Zhang, Y.; Jiang, D. Extracellular vesicles produced by bone marrow mesenchymal stem cells attenuate renal fibrosis, in part by inhibiting the RhoA/ROCK pathway, in a UUO rat model. Stem Cell Res. Ther. 2020, 11, 253. [Google Scholar] [CrossRef]
- Grange, C.; Tritta, S.; Tapparo, M.; Cedrino, M.; Tetta, C.; Camussi, G.; Brizzi, M.F. Stem cell-derived extracellular vesicles inhibit and revert fibrosis progression in a mouse model of diabetic nephropathy. Sci. Rep. 2019, 9, 4468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Scarpellini, A.; Funck, M.; Verderio, E.A.; Johnson, T.S. Development of a Chronic Kidney Disease Model in C57BL/6 Mice with Relevance to Human Pathology. Nephron Extra 2013, 3, 12–29. [Google Scholar] [CrossRef] [PubMed]
- Jadot, I.I.; Declèves, A.-E.; Nortier, J.; Caron, N. An integrated view of aristolochic acid nephropathy: Update of the literature. Int. J. Mol. Sci. 2017, 18, 297. [Google Scholar] [CrossRef] [Green Version]
- Kholia, S.; Sanchez, M.B.H.; Cedrino, M.; Papadimitriou, E.; Tapparo, M.; Deregibus, M.C.; Bruno, S.; Antico, F.; Brizzi, M.F.; Quesenberry, P.J.; et al. Mesenchymal Stem Cell Derived Extracellular Vesicles Ameliorate Kidney Injury in Aristolochic Acid Nephropathy. Front. Cell Dev. Biol. 2020, 8, 188. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, Y.F.; Fu, G.P.; Guan, C.; Zhang, X.; Yang, D.G.; Shi, Y.C. Protective effect of miRNA-containing extracellular vesicles derived from mesenchymal stromal cells of old rats on renal function in chronic kidney disease. Stem Cell Res. Ther. 2020, 11, 274. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Wang, Y.; Lu, X.; Zhu, B.; Pei, X.; Wu, J.; Zhao, W. Micro-vesicles derived from bone marrow stem cells protect the kidney both in vivo and in vitro by microRNA-dependent repairing. Nephrology 2015, 20, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Zhang, J.; Zhu, Y.; Shi, H.; Yin, S.; Sun, F.; Wang, Q.; Zhang, L.; Yan, Y.; Zhang, X.; et al. Exosomes derived from hucMSC attenuate renal fibrosis through CK1δ/β-TRCP-mediated YAP degradation. Cell Death Dis. 2020, 11, 327. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.-Z.; Zhong, X.; Li, J.-C.; Zhang, Y.-W.; Yan, Y.; Liao, Y.; Wen, D.; Diao, H.; Wang, L.; Shen, H. An optimized 5/6 nephrectomy mouse model based on unilateral kidney ligation and its application in renal fibrosis research. Ren. Fail. 2019, 41, 555–566. [Google Scholar] [CrossRef]
- He, J.; Wang, Y.; Sun, S.; Yu, M.; Wang, C.; Pei, X.; Zhu, B.; Wu, J.; Zhao, W. Bone marrow stem cells-derived microvesicles protect against renal injury in the mouse remnant kidney model. Nephrology 2012, 17, 493–500. [Google Scholar] [CrossRef]
- Wan, F.; Yang, R.-C.; Tang, Y.-W.; Tang, X.-L.; Ye, T.; Zheng, J.; Zhang, H.-Q.; Lin, Y. BMSC-derived exosomes protect against kidney injury through regulating klotho in 5/6 nephrectomy rats. Eur. J. Med. Res. 2022, 27, 118. [Google Scholar] [CrossRef]
- Lindoso, R.S.; Lopes, J.A.; Binato, R.; Abdelhay, E.; Takiya, C.M.; de Miranda, K.R.; Lara, L.S.; Viola, A.; Bussolati, B.; Vieyra, A.; et al. Adipose Mesenchymal Cells-Derived EVs Alleviate DOCA-Salt-Induced Hypertension by Promoting Cardio-Renal Protection. Mol. Ther. Methods Clin. Dev. 2020, 16, 63–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kholia, S.; Sanchez, M.B.H.; Cedrino, M.; Papadimitriou, E.; Tapparo, M.; Deregibus, M.C.; Brizzi, M.F.; Tetta, C.; Camussi, G. Human liver stem cell-derived extracellular vesicles prevent aristolochic acid-induced kidney fibrosis. Front. Immunol. 2018, 9, 1639. [Google Scholar] [CrossRef] [Green Version]
- Zhong, X.; Tang, T.-T.; Shen, A.-R.; Cao, J.-Y.; Jing, J.; Wang, C.; Zhu, X.-X.; Wen, Y.; Li, Z.-L.; Bin Wang, B.; et al. Tubular epithelial cells-derived small extracellular vesicle-VEGF-A promotes peritubular capillary repair in ischemic kidney injury. npj Regen. Med. 2022, 7, 73. [Google Scholar] [CrossRef]
- Jiang, Z.-Z.; Liu, Y.-M.; Niu, X.; Yin, J.-Y.; Hu, B.; Guo, S.-C.; Fan, Y.; Wang, Y.; Wang, N.-S. Exosomes secreted by human urine-derived stem cells could prevent kidney complications from type I diabetes in rats. Stem Cell Res. Ther. 2016, 7, 24. [Google Scholar] [CrossRef] [Green Version]
- Thakur, A.; Parra, D.C.; Motallebnejad, P.; Brocchi, M.; Chen, H.J. Exosomes: Small vesicles with big roles in cancer, vaccine development, and therapeutics. Bioact. Mater. 2021, 10, 281–294. [Google Scholar] [CrossRef]
- Zhang, K.; Li, R.; Chen, X.; Yan, H.; Li, H.; Zhao, X.; Huang, H.; Chen, S.; Liu, Y.; Wang, K.; et al. Renal Endothelial Cell-Targeted Extracellular Vesicles Protect the Kidney from Ischemic Injury. Adv. Sci. 2022, 10, 2204626. [Google Scholar] [CrossRef]
- Wu, P.; Tang, Y.; Jin, C.; Wang, M.; Li, L.; Liu, Z.; Shi, H.; Sun, Z.; Hou, X.; Chen, W.; et al. Neutrophil membrane engineered HucMSC sEVs alleviate cisplatin-induced AKI by enhancing cellular uptake and targeting. J. Nanobiotechnol. 2022, 20, 353. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Sun, Q.; Wang, Z.; Wang, F.; Chen, F.; Wang, H.; Shang, G.; Chen, X.; Ding, S.; Li, C.; et al. Tubular epithelial cells derived-exosomes containing CD26 protects mice against renal ischemia/reperfusion injury by maintaining proliferation and dissipating inflammation. Biochem. Biophys. Res. Commun. 2021, 553, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Yang, X.; Wu, J.; Tang, S.; Yang, L.; Fei, X.; Wang, M. Exosome from indoleamine 2,3-dioxygenase-overexpressing bone marrow mesenchymal stem cells accelerates repair process of ischemia/reperfusion-induced acute kidney injury by regulating macrophages polarization. Stem Cell Res. Ther. 2022, 13, 367. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.-T.; Wang, B.; Wu, M.; Li, Z.-L.; Feng, Y.; Cao, J.-Y.; Yin, D.; Liu, H.; Tang, R.-N.; Crowley, S.D.; et al. Extracellular vesicle–encapsulated IL-10 as novel nanotherapeutics against ischemic AKI. Sci. Adv. 2020, 6, eaaz0748. [Google Scholar] [CrossRef]
- Tang, T.-T.; Wang, B.; Li, Z.-L.; Wen, Y.; Feng, S.-T.; Wu, M.; Liu, D.; Cao, J.-Y.; Yin, Q.; Yin, D.; et al. Kim-1 targeted extracellular vesicles: A new therapeutic platform for RNAi to treat AKI. J. Am. Soc. Nephrol. 2021, 32, 2467–2483. [Google Scholar] [CrossRef]
- Liu, Y.; Cui, J.; Wang, H.; Hezam, K.; Zhao, X.; Huang, H.; Chen, S.; Han, Z.; Han, Z.-C.; Guo, Z.; et al. Enhanced therapeutic effects of MSC-derived extracellular vesicles with an injectable collagen matrix for experimental acute kidney injury treatment. Stem Cell Res. Ther. 2020, 11, 161. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Wang, B.; Tang, T.; Lv, L.; Ding, Z.; Li, Z.; Hu, R.; Wei, Q.; Shen, A.; Fu, Y.; et al. Three-dimensional culture of MSCs produces exosomes with improved yield and enhanced therapeutic efficacy for cisplatin-induced acute kidney injury. Stem Cell Res. Ther. 2020, 11, 206. [Google Scholar] [CrossRef]
- Provatopoulou, S.T.; Ziroyiannis, P.N. Clinical use of erythropoietin in chronic kidney disease: Outcomes and future prospects. Hippokratia 2011, 15, 109–115. [Google Scholar]
- Wang, Y.; Lu, X.; He, J.; Zhao, W. Influence of erythropoietin on microvesicles derived from mesenchymal stem cells protecting renal function of chronic kidney disease. Stem Cell Res. Ther. 2015, 6, 100. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Qian, F.; Zheng, D.; He, W.; Gong, J.; He, Q. Mesenchymal stem cells attenuate renal fibrosis via exosomes-mediated delivery of microRNA Let-7i-5p antagomir. Int. J. Nanomed. 2021, 16, 3565–3578. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhao, Y.; Wang, Z.; Wang, J.; Liu, C.; Sun, D. Transplantation of Amniotic Fluid-Derived Stem Cells Preconditioned with Glial Cell Line-Derived Neurotrophic Factor Gene Alleviates Renal Fibrosis. Cell Transplant. 2019, 28, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Wang, Y.; Li, S.; Zuo, B.; Zhang, X.; Wang, F.; Sun, D. Exosomes derived from GDNF-modified human adipose mesenchymal stem cells ameliorate peritubular capillary loss in tubulointerstitial fibrosis by activating the SIRT1/eNOS signaling pathway. Theranostics 2020, 10, 9425–9442. [Google Scholar] [CrossRef]
- Han, Y.-S.; Yoon, Y.M.; Go, G.; Lee, J.H.; Lee, S.H. Melatonin protects human renal proximal tubule epithelial cells against high glucose-mediated fibrosis via the cellular prion protein-TGF-β-Smad signaling axis. Int. J. Med. Sci. 2020, 17, 1235–1245. [Google Scholar] [CrossRef] [PubMed]
- Yea, J.-H.; Yoon, Y.M.; Lee, J.H.; Yun, C.W.; Lee, S.H. Exosomes isolated from melatonin-stimulated mesenchymal stem cells improve kidney function by regulating inflammation and fibrosis in a chronic kidney disease mouse model. J. Tissue Eng. 2021, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Chen, B.; Chen, F.; Yang, S.; Zhu, C.; Ma, Y.; Li, Y.; Shi, J. Exosomal microRNA-16-5p from human urine-derived stem cells ameliorates diabetic nephropathy through protection of podocyte. J. Cell. Mol. Med. 2020, 25, 10798–10813. [Google Scholar] [CrossRef] [Green Version]
- Nassar, W.; El-Ansary, M.; Sabry, D.; Mostafa, M.A.; Fayad, T.; Kotb, E.; Temraz, M.; Saad, A.-N.; Essa, W.; Adel, H. Umbilical cord mesenchymal stem cells derived extracellular vesicles can safely ameliorate the progression of chronic kidney diseases. Biomater. Res. 2016, 20, 21. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-Q.; Liu, J.-F.; Liu, H.; Meng, Y. Extracellular vesicles for ischemia/reperfusion injury-induced acute kidney injury: A systematic review and meta-analysis of data from animal models. Syst. Rev. 2022, 11, 197. [Google Scholar] [CrossRef]



| In Vivo Model | EV-Source | EV Isolation Method | EV Route of Administration | EV Biological Effect/ Mechanism of Action | References |
|---|---|---|---|---|---|
| IRI rat model Bilateral renal arterial clamping for 45 min | h-iPSCs and A-MSCs (1 × 109 EV/dose) | UC | Single sub-capsular injection at the beginning of reperfusion in each kidney |
| [39] |
| IRI rat model Clamping of left renal pedicle for 45 min and contralateral nephrectomy | USCs (20 µg EV/dose) | Exo-Quick (co-precipitation) | Single dorsal penis vein injection |
| [40] |
| IRI mouse model Microvascular clamping for 45 min | USCs (20 μg EV/dose) | UC | Single tail intravenous injection 15 min before renal artery clamping |
| [41] |
| Glycerol mouse model Intra-muscular glycerol injection into inferior hind limbs (8 mL/kg) | Human Urine (2 × 108 EV/dose) | UC | Tail intravenous injection 1 day after the injury |
| [42] |
| In Vivo Model | EV-Source | EV Isolation Method | EV Route of Administration | EV Biological Effect/ Mechanism of Action | References |
|---|---|---|---|---|---|
| STZ-DN mouse model Intraperitoneal injection of STZ (37 mg/kg) for 4 consecutive days | HLSCs (1 × 1010 EV/dose) | UC | Multiple intravenous injections once a week for 4 weeks starting from day 30 after diabetes onset |
| [46] |
| AAN mouse model Intraperitoneal injection of 4 mg/kg of AA on a weekly basis for 4 weeks | HLSCs (1 × 1010 EV/dose) | UC | Multiple intravenous injections on a weekly basis starting from 3 days after AA administration |
| [57] |
| IRI mouse model 30 min IRI with contralateral nephrectomy | HLSCs (1 × 109 EV/dose) | UC | Two intravenous injections: one immediately after the surgery and one after three days |
| [44] |
| In Vivo Model | EV-Source | EV Isolation Method | EV Route of Administration | EV Biological Effect/ Mechanism of Action | References |
|---|---|---|---|---|---|
| IRI mouse model 35 min IRI and contralateral nephrectomy | m-TECs (200µg EV/dose) | UC | Multiple intravenous injections after IRI and continued every 12 h for seven times |
| [58] |
| STZ-DN rat model Intraperitoneal injection of STZ (65 mg/kg) | h-USCs (100 μg EV/dose) | UC with Ultra-clear tube (Merck-Millipore, Darnstadt, Germany) | Multiple tail intravenous injections once a week |
| [59] |
| In Vivo Model | EV-Source and Route of Administration | EV Isolation Method | EV Engineering Method | EV Biological Effect | References |
|---|---|---|---|---|---|
| IRI mouse model Left renal pedicle clamping for 15, 30, and 45 min | P-MSCs (100 µg EV/dose) Intravenous injection after 12 h from reperfusion | UC | Hydrophobic insertion of P-Selectin Binding Protein (PBP) coupled with Cyanine 5.5 |
| [61] |
| Cisplatin mouse model Subcutaneous cisplatin injection (10 mg/kg) | h-UC-MSCs and neutrophils (1 × 1010 EV/dose) Tail intravenous injection for 2 days | -h-UC-MSC-EVs obtained via UC -Neutrophil-EVs obtained via cold ultrasonication | NEX obtained from ratio 1:1 of h-UC-MSC-EVs and neutrophil-EVs via sequential polycarbonate membrane extrusion |
| [62] |
| IRI mouse model Ligation of left renal artery and vein with contralateral nephrectomy | TECs (100 mg EV/0,5 mL) Single tail intravenous injection 12 h after the surgery | UC | Adenovirus transfection of TECs inducing CD26 overexpression |
| [63] |
| IRI mouse model Renal pedicle clamping for 25 min | h-BM-MSCs (100 µg EV/dose) Intravenous injection 6 h after IRU | UC | Plasmid transfection to overexpress IDO |
| [64] |
| IRI mouse model Renal pedicle clamping for 35 min | RAW macrophages (200 µg EV/dose) Intravenous injection after reperfusion and every 24 h three times | UC | CMV-MCSSV40-Neomycin IL-10 plasmid transfection with Lipofectamine |
| [65] |
| IRI and UUO mouse model IRI: Renal pedicle clamping for 35 min UUO: ligation of the left ureter and right ureter was not ligated | Red blood cells (100–150 µg EV/dose) Multiple injections once a day for five consecutive days | UC | Engineering of EV surface with Kim-1 bound to LTH peptide and EV cargo with siRNA for P65 and Snail65 |
| [66] |
| IRI mouse model Renal pedicle clamping for 40 min | P-MSCs (100 µg EV/dose) Intravenous injection into three sites 5 min after reperfusion | UC | Naïve EV combined with collagen matrix |
| [67] |
| Cisplatin mouse model subcutaneous cisplatin injection (18 mg/kg) | h-UC-MSCs (100 µg EV/dose) Two intravenous injections at 24 and 48 h | UC | Bioreactor 3D |
| [68] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ceccotti, E.; Saccu, G.; Herrera Sanchez, M.B.; Bruno, S. Naïve or Engineered Extracellular Vesicles from Different Cell Sources: Therapeutic Tools for Kidney Diseases. Pharmaceutics 2023, 15, 1715. https://doi.org/10.3390/pharmaceutics15061715
Ceccotti E, Saccu G, Herrera Sanchez MB, Bruno S. Naïve or Engineered Extracellular Vesicles from Different Cell Sources: Therapeutic Tools for Kidney Diseases. Pharmaceutics. 2023; 15(6):1715. https://doi.org/10.3390/pharmaceutics15061715
Chicago/Turabian StyleCeccotti, Elena, Gabriele Saccu, Maria Beatriz Herrera Sanchez, and Stefania Bruno. 2023. "Naïve or Engineered Extracellular Vesicles from Different Cell Sources: Therapeutic Tools for Kidney Diseases" Pharmaceutics 15, no. 6: 1715. https://doi.org/10.3390/pharmaceutics15061715