
Controlling the Evolution of Selective Vancomycin Resistance through Successful Ophthalmic Eye-Drop Preparation of Vancomycin-Loaded Nanoliposomes Using the Active-Loading Method

 , ,
, ,
Abstract
:1. Introduction
2. Materials
3. Methodology
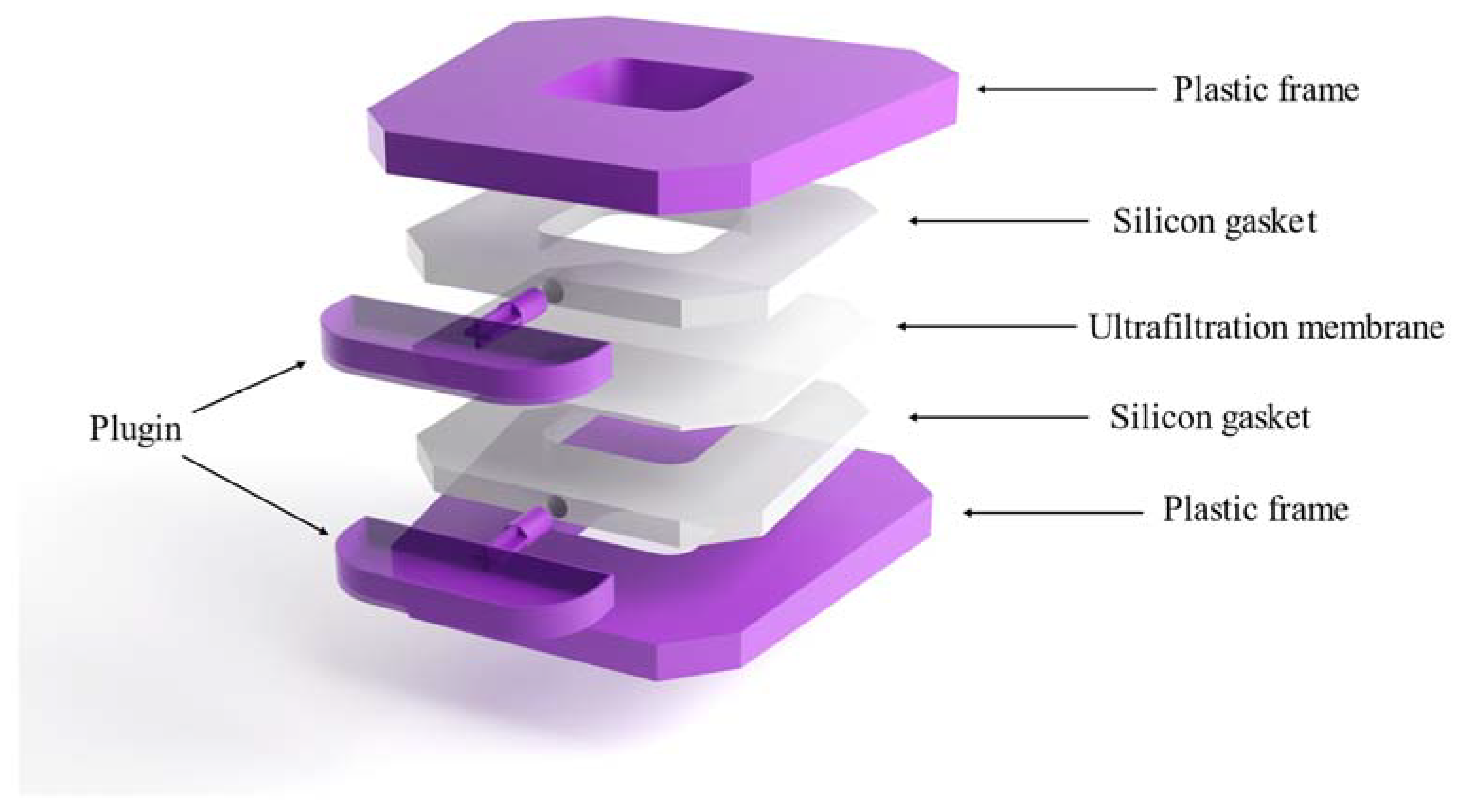
3.1. Cassette Construction
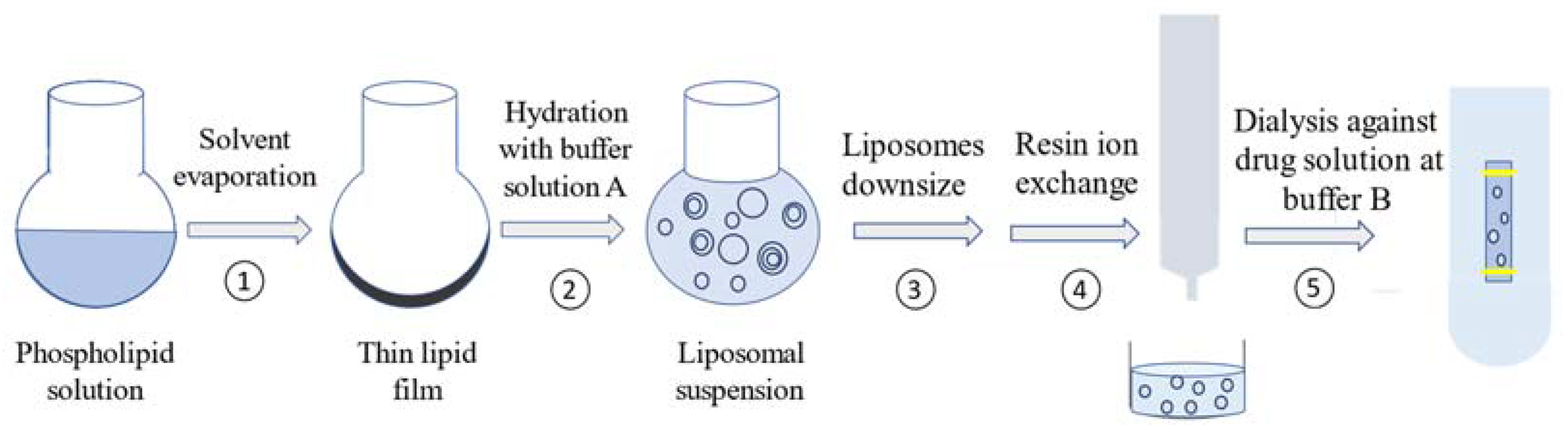
3.2. Liposome Preparation
3.2.1. Preparation of Drug-Free Liposomes
3.2.2. Liposome Downsizing
3.2.3. Gel Filtration and Ion Exchange
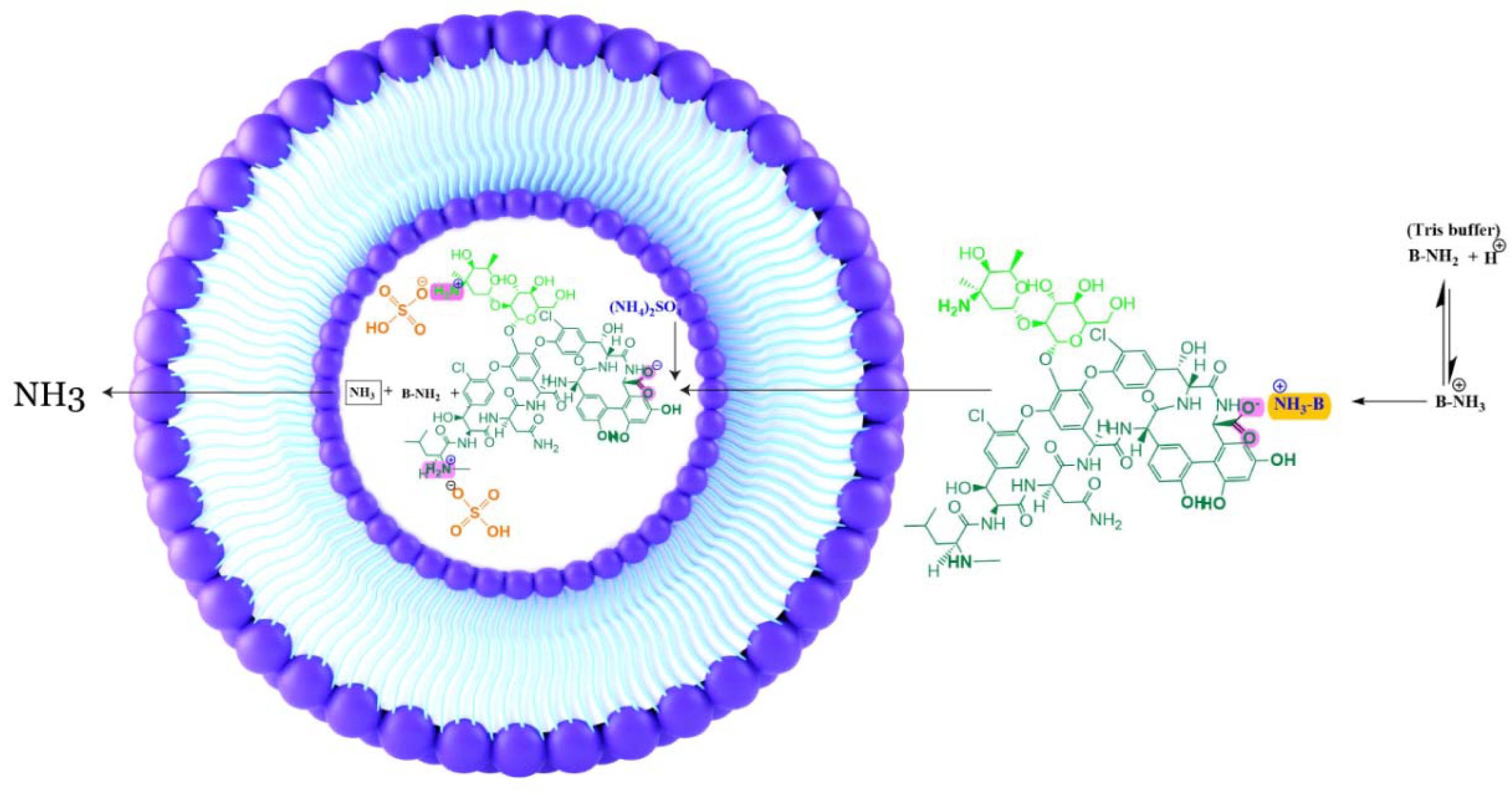
3.2.4. Drug Loading
3.3. Evaluation of Vancomycin-Loaded Liposomes
3.3.1. Particle Size and Zeta-Potential Determination
3.3.2. Entrapment Efficiency
3.3.3. Morphological Characterization
3.3.4. Differential Scanning Colorimetry (DSC)
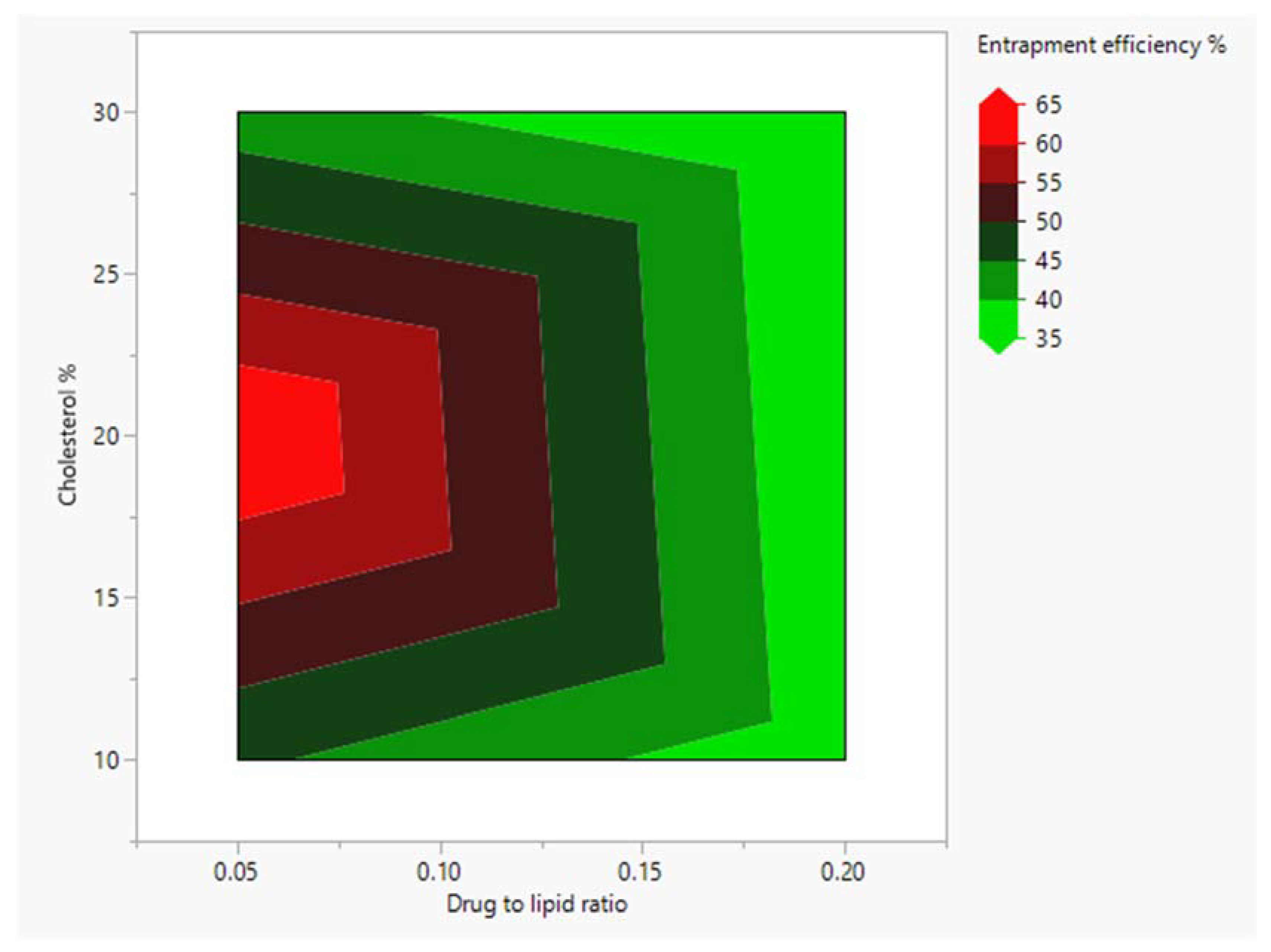
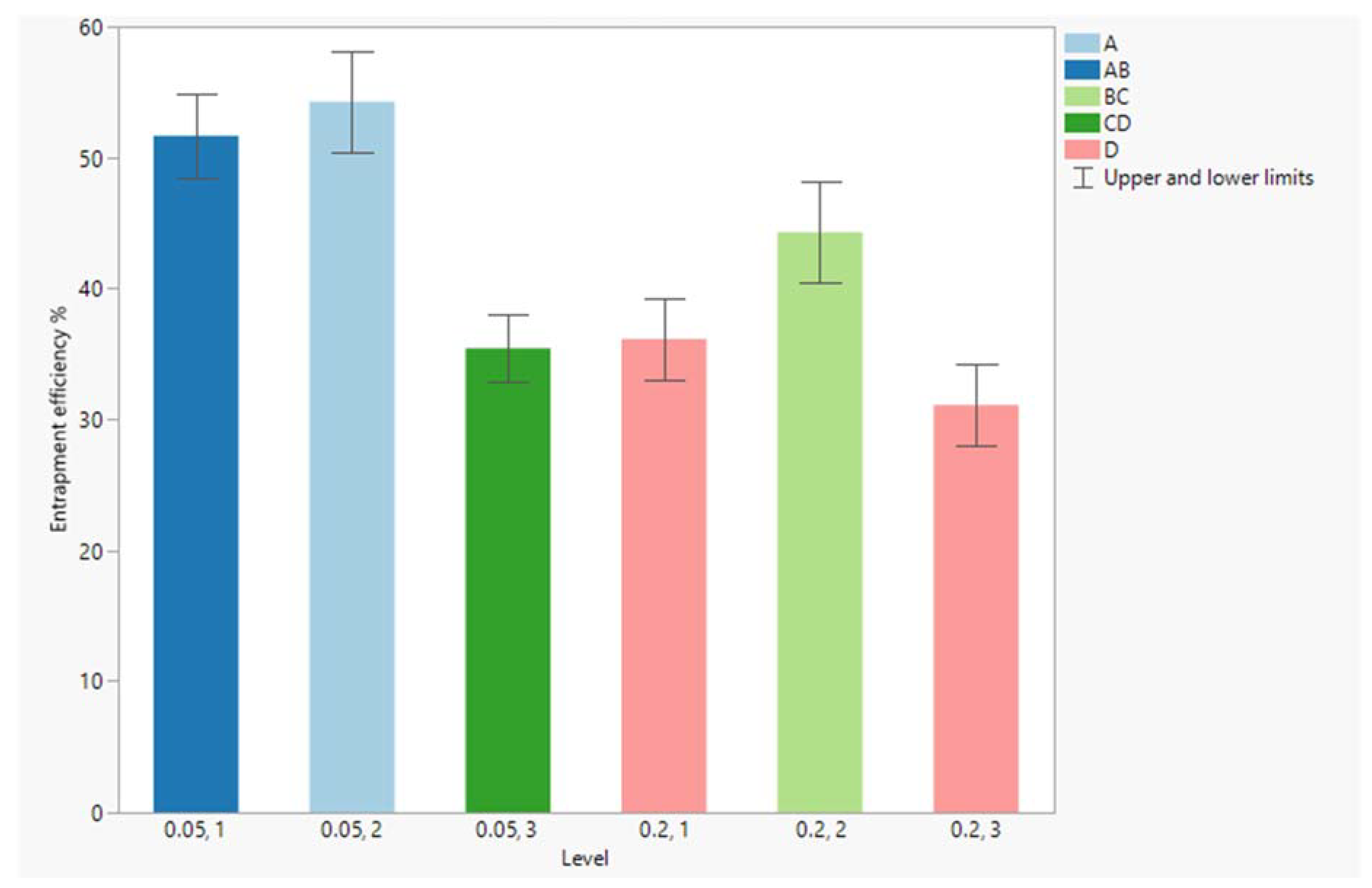
3.4. Experimental Design and Statistical Analysis
3.5. Antimicrobial Activity
3.5.1. Antibacterial-Activity MIC and MBC
3.5.2. Effect of the Optimized Vancomycin-Loaded Liposomes on the Antibiofilm Activity of Vancomycin
Biofilm Inhibition Assay
Biofilm Eradication Assay
3.5.3. In Vitro Resistance Study
3.6. In Vitro Eye-Safety Test
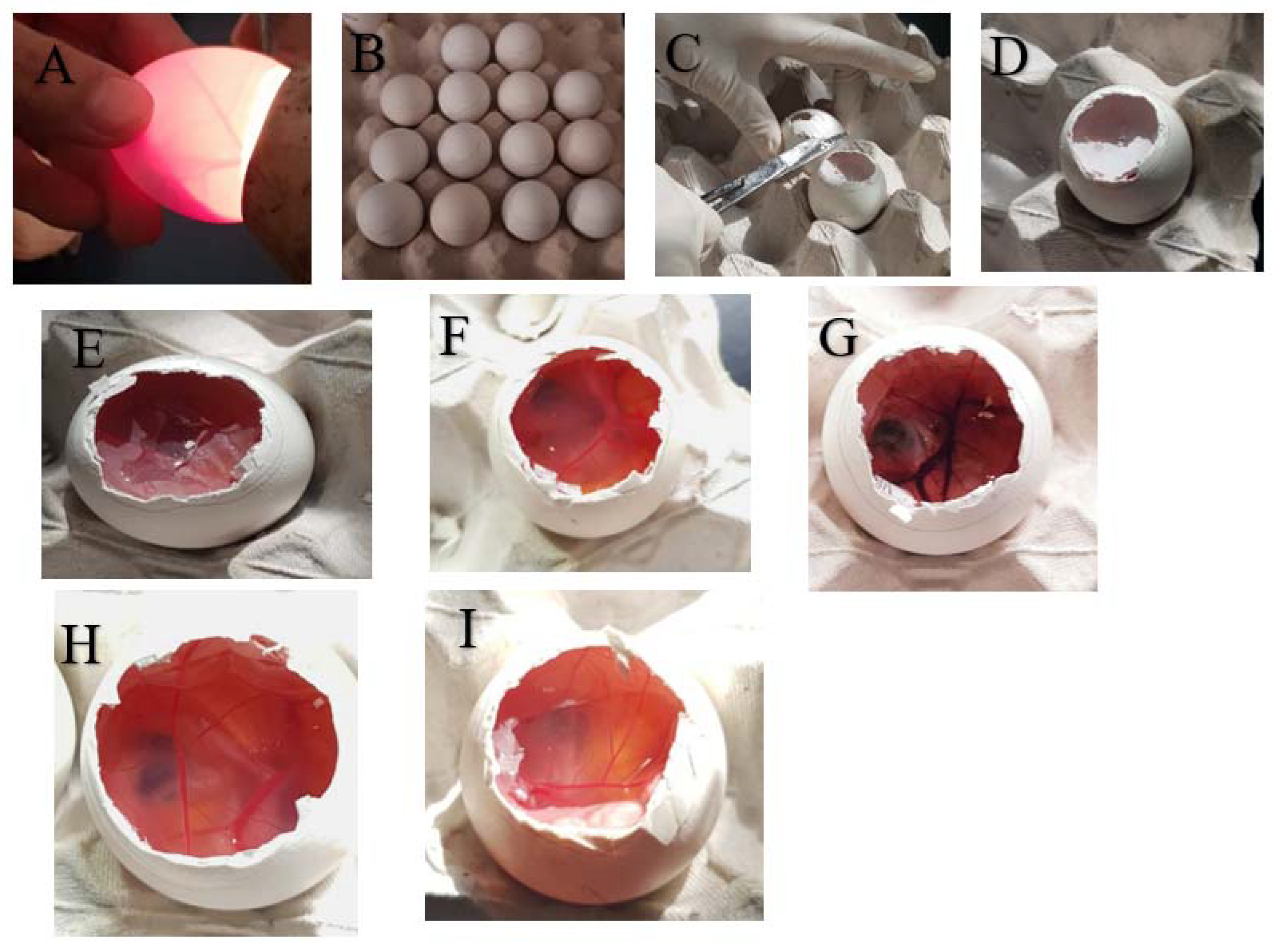
3.6.1. Preparing the Eggs
3.6.2. Application of Test Materials
3.6.3. Visual Analysis
4. Results and Discussion
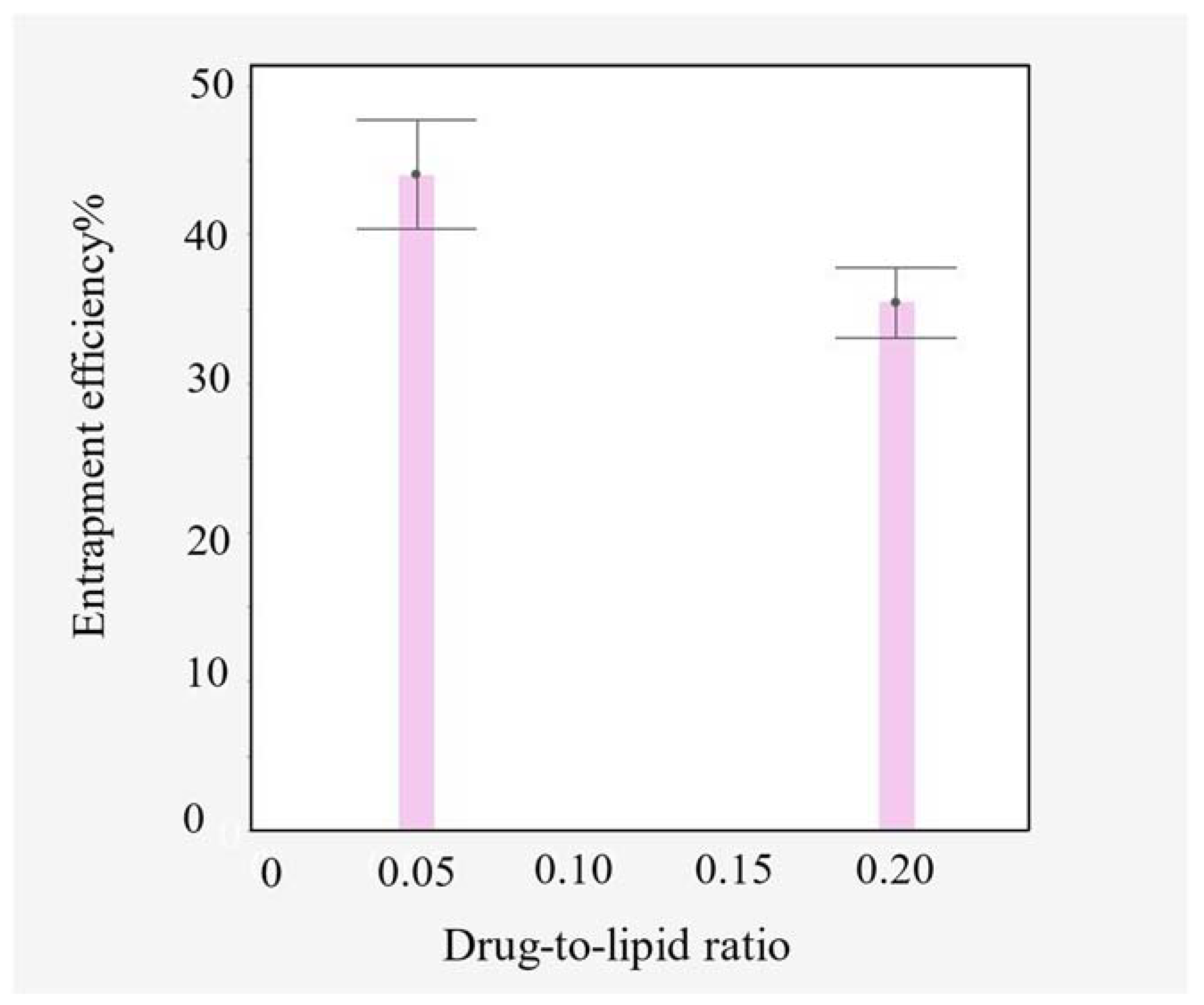
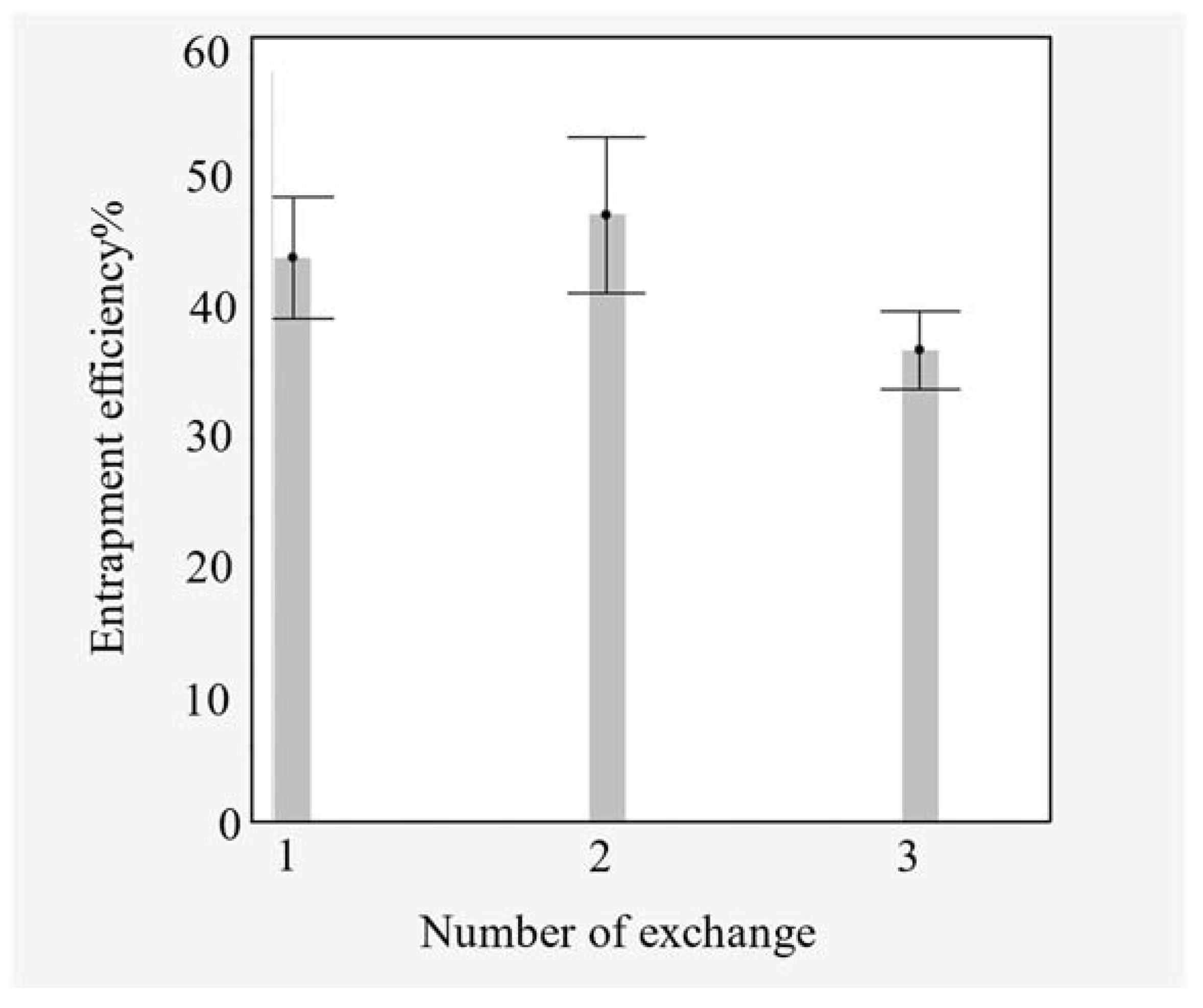
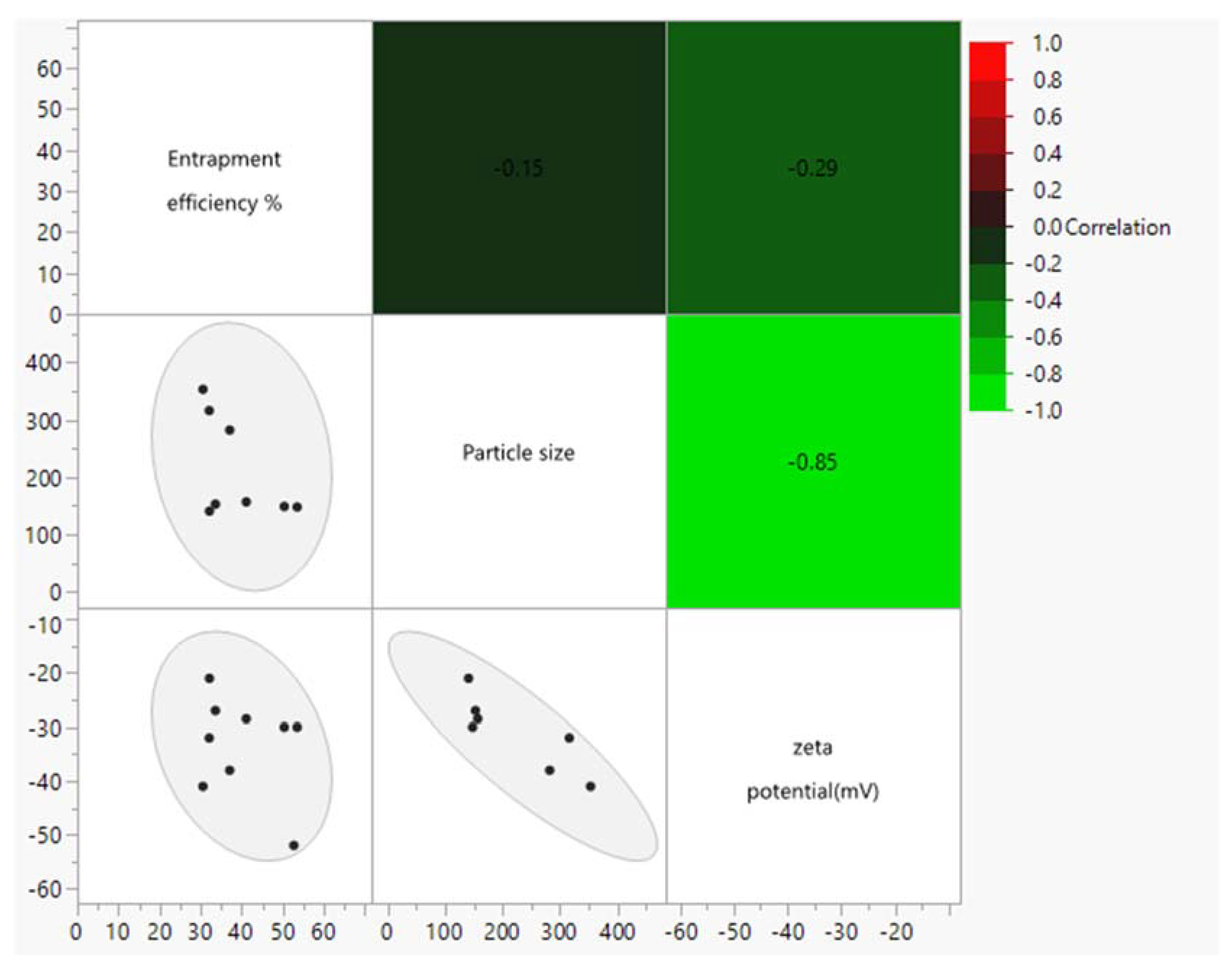
4.1. Entrapment Efficiency
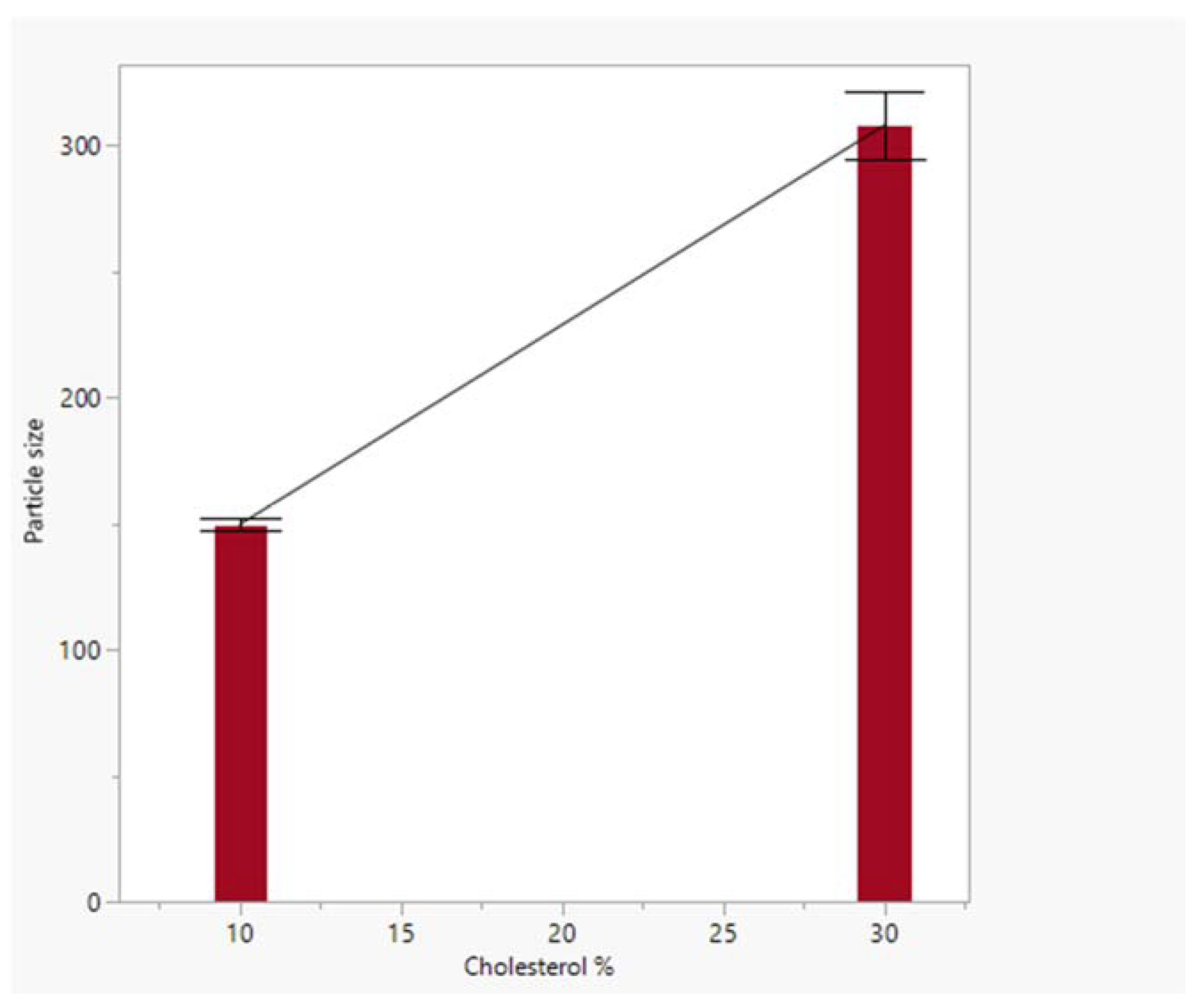
4.2. Particle Size
Hydrodynamic Sizes and Zeta Potential of Liposomes as Measured with Dynamic Light Scattering (DLS)
4.3. Zeta Potential
4.4. Physicochemical Characterization of Optimized Vancomycin-Loaded Nanoliposomes
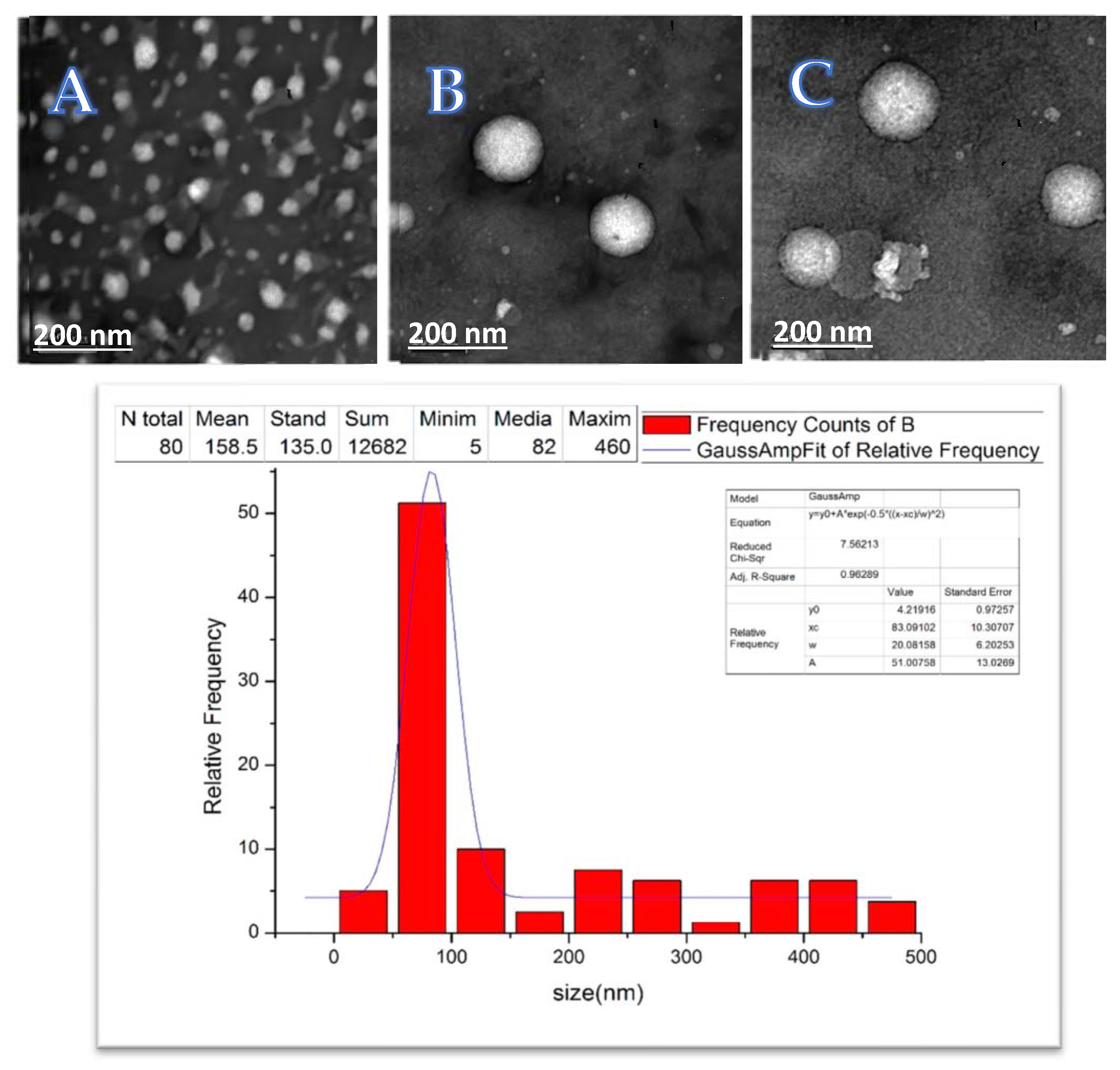
4.4.1. Liposome Size and Morphology Measurements with Transmission Electron Microscopy (TEM) Imaging
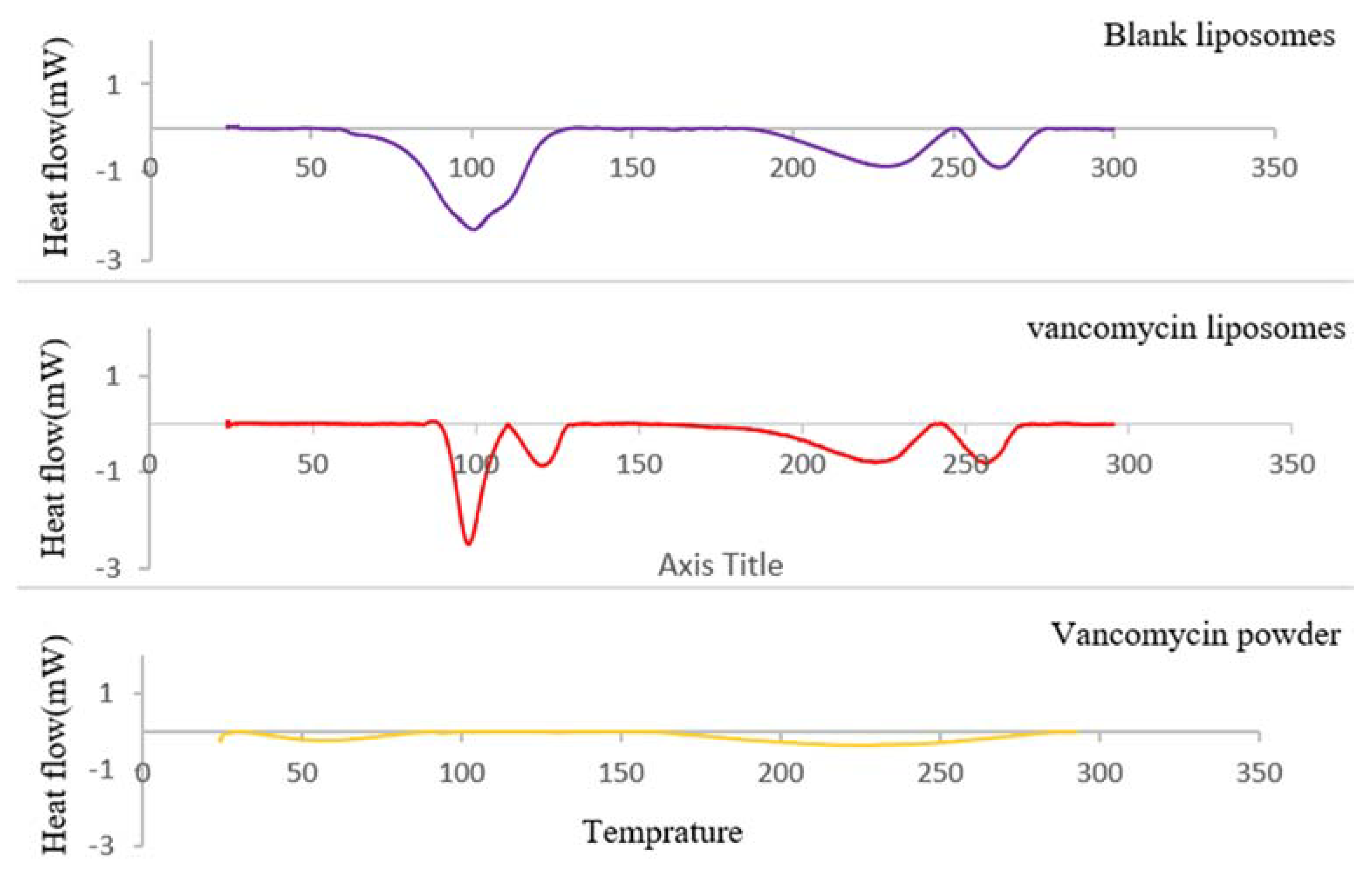
4.4.2. Physicochemical Thermodynamic Stability Determination Using Differential Scanning Calorimetry (DSC)
4.5. Antibacterial Activity
4.5.1. Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
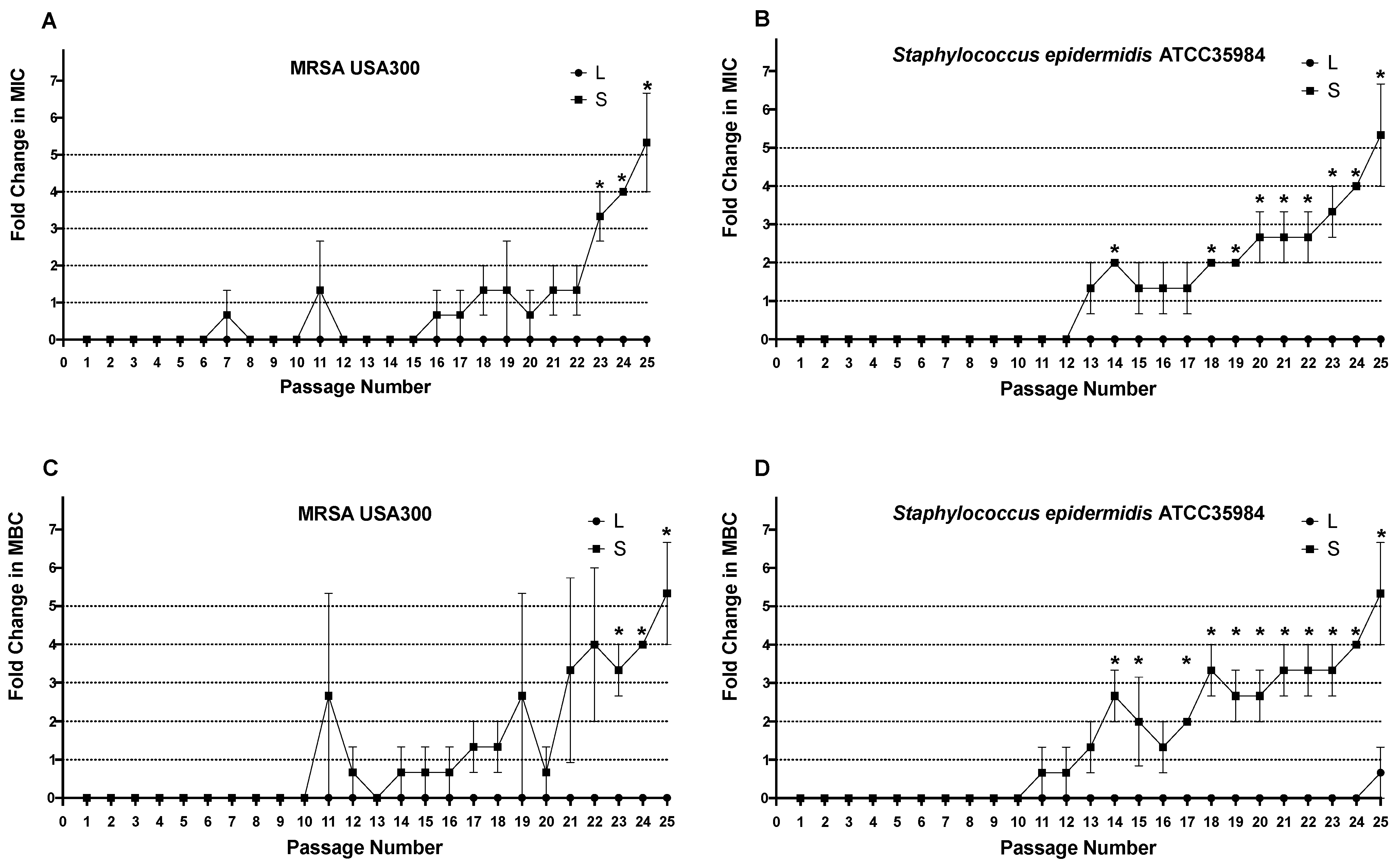
4.5.2. In Vitro Resistance Study
4.5.3. Effect of the Optimized Formula (Liposomes) on the Antibiofilm Activity of Vancomycin
4.6. In Vitro Eye Irritation Test
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bourne, R.R.A.; Steinmetz, J.D.; Saylan, M.; Mersha, A.M.; Weldemariam, A.H.; Wondmeneh, T.G.; Sreeramareddy, C.T.; Pinheiro, M.; Yaseri, M.; Yu, C.; et al. Causes of blindness and vision impairment in 2020 and trends over 30 years, and prevalence of avoidable blindness in relation to VISION 2020: The Right to Sight: An analysis for the Global Burden of Disease Study. Lancet Glob. Health 2021, 9, e144–e160. [Google Scholar] [CrossRef]
- Burton, M.J.; Ramke, J.; Marques, A.P.; A Bourne, R.R.; Congdon, N.; Jones, I.; Tong, B.A.M.A.; Arunga, S.; Bachani, D.; Bascaran, C.; et al. The Lancet Global Health Commission on Global Eye Health: Vision beyond 2020. Lancet Glob. Health 2021, 9, e489–e551. [Google Scholar] [CrossRef] [PubMed]
- Ung, L.; Bispo, P.J.; Shanbhag, S.S.; Gilmore, M.S.; Chodosh, J. The persistent dilemma of microbial keratitis: Global burden, diagnosis, and antimicrobial resistance. Surv. Ophthalmol. 2018, 64, 255–271. [Google Scholar] [CrossRef] [PubMed]
- Grzybowski, A.; Brona, P.; Kim, S.J. Microbial flora and resistance in ophthalmology: A review. Graefe’s Arch. Clin. Exp. Ophthalmol. 2017, 255, 851–862. [Google Scholar] [CrossRef] [PubMed]
- O’neill, E.C.; Yeoh, J.; A Fabinyi, D.C.; Cassidy, D.; Vajpayee, R.B.; Allen, P.; Connell, P.P. Risk factors, microbial profiles and prognosis of microbial keratitis-associated endophthalmitis in high-risk eyes. Graefe’s Arch. Clin. Exp. Ophthalmol. 2014, 252, 1457–1462. [Google Scholar] [CrossRef]
- Ting, D.S.J.; Settle, C.; Morgan, S.J.; Baylis, O.; Ghosh, S. A 10-year analysis of microbiological profiles of microbial keratitis: The North East England Study. Eye 2018, 32, 1416–1417. [Google Scholar] [CrossRef]
- Asbell, P.A.; Sanfilippo, C.M.; Pillar, C.M.; DeCory, H.H.; Sahm, D.F.; Morris, T.W. Antibiotic resistance among ocular pathogens in the United States five-year results from the Antibiotic Resistance Monitoring in Ocular Microorganisms (ARMOR) surveillance study. JAMA Ophthalmol. 2015, 133, 1445–1454. [Google Scholar] [CrossRef]
- Lin, A.; Rhee, M.K.; Akpek, E.K.; Amescua, G.; Farid, M.; Garcia-Ferrer, F.J.; Varu, D.M.; Musch, D.; Dunn, S.P.; Mah, F.S.; et al. Bacterial Keratitis Preferred Practice Pattern®. Ophthalmology 2019, 126, P1–P55. [Google Scholar] [CrossRef]
- Chiquet, C.; Maurin, M.; Altayrac, J.; Aptel, F.; Boisset, S.; Vandenesch, F.; Cornut, P.; Romanet, J.; Gain, P.; Carricajo, A. Correlation between clinical data and antibiotic resistance in coagulase-negative Staphylococcus species isolated from 68 patients with acute post-cataract endophthalmitis. Clin. Microbiol. Infect. 2015, 21, 592.e1–592.e8. [Google Scholar] [CrossRef]
- Tena, D.; Rodríguez, N.; Toribio, L.; González-Praetorius, A. Infectious Keratitis: Microbiological Review of 297 Cases. Jpn. J. Infect. Dis. 2019, 72, 121–123. [Google Scholar] [CrossRef]
- Saillard, J.; Spiesser-Robelet, L.; Gohier, P.; Briot, T. Bacterial keratitis treated by strengthened antibiotic eye drops: An 18 months review of clinical cases and antibiotic susceptibilities. Ann. Pharm. Françaises 2018, 76, 107–113. [Google Scholar] [CrossRef]
- Hussain, S.; Joo, J.; Kang, J.; Kim, B.; Braun, G.B.; She, Z.-G.; Kim, D.; Mann, A.P.; Mölder, T.; Teesalu, T.; et al. Antibiotic-loaded nanoparticles targeted to the site of infection enhance antibacterial efficacy. Nat. Biomed. Eng. 2018, 2, 95–103. [Google Scholar] [CrossRef]
- Lakshminarayanan, R.; Ye, E.; Young, D.J.; Li, Z.; Loh, X.J. Recent Advances in the Development of Antimicrobial Nanoparticles for Combating Resistant Pathogens. Adv. Healthc. Mater. 2018, 7, e1701400. [Google Scholar] [CrossRef]
- Mi, G.J.; Shi, D.; Wang, M.; Webster, T.J. Reducing Bacterial Infections and Biofilm Formation Using Nanoparticles and Nanostructured Antibacterial Surfaces. Adv. Healtc. Mater. 2018, 7, e1800103. [Google Scholar] [CrossRef]
- Aminzadeh, Z.; Yadegarynia, D.; Fatemi, A.; Tahmasebian Dehkordi, E.; Azad Armaki, S. Vancomycin Minimum Inhibitory Concentration for Methicillin-Resistant Staphylococcus aureus Infections; Is There Difference in Mortality Between Patients? Jundishapur J. Microbiol. 2014, 7, e12831. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Chen, D.-Y.; Zhu, Y.-W.; Jiang, Z.-L.; Cui, K.; Zhang, S.; Chen, L.-H. Population pharmacokinetic modeling and clinical application of vancomycin in Chinese patients hospitalized in intensive care units. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rybak, M.J. The Pharmacokinetic and Pharmacodynamic Properties of Vancomycin. Clin. Infect. Dis. 2006, 42, S35–S39. [Google Scholar] [CrossRef]
- Butler, M.S.; Hansford, K.A.; Blaskovich, M.A.T.; Halai, R.; Cooper, M.A. Glycopeptide antibiotics: Back to the future. J. Antibiot. 2014, 67, 631–644. [Google Scholar] [CrossRef]
- Courvalin, P. Vancomycin Resistance in Gram-Positive Cocci. Clin. Infect. Dis. 2006, 42 (Suppl. 1), S25–S34. [Google Scholar] [CrossRef]
- Cui, L.; Ma, X.; Sato, K.; Okuma, K.; Tenover, F.C.; Mamizuka, E.M.; Gemmell, C.G.; Kim, M.-N.; Ploy, M.-C.; El Solh, N.; et al. Cell Wall Thickening Is a Common Feature of Vancomycin Resistance in Staphylococcus aureus. J. Clin. Microbiol. 2003, 41, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, U.H.; Devnarain, N.; Omolo, C.A.; Mocktar, C.; Govender, T. Biomimetic pH/lipase dual responsive vitamin-based solid lipid nanoparticles for on-demand delivery of vancomycin. Int. J. Pharm. 2021, 607, 120960. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Preet, S.; Kumar, V.; Kumar, R.; Kumar, R. Synergetic effect of vancomycin loaded silver nanoparticles for enhanced antibacterial activity. Colloids Surf. B Biointerfaces 2018, 176, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Dizaji, A.N.; Ozek, N.S.; Aysin, F.; Calis, A.; Yilmaz, A.; Yilmaz, M. Combining vancomycin-modified gold nanorod arrays and colloidal nanoparticles as a sandwich model for the discrimination of Gram-positive bacteria and their detection via surface-enhanced Raman spectroscopy (SERS). Analyst 2021, 146, 3642–3653. [Google Scholar] [CrossRef] [PubMed]
- Sande, L.; Sanchez, M.; Montes, J.; Wolf, A.J.; Morgan, M.A.; Omri, A.; Liu, G.Y. Liposomal encapsulation of vancomycin improves killing of methicillin-resistant Staphylococcus aureus in a murine infection model. J. Antimicrob. Chemother. 2012, 67, 2191–2194. [Google Scholar] [CrossRef]
- Singh, S.; Aldawsari, H.M.; Alam, A.; Alqarni, M.H.S.; Ranjan, S.; Kesharwani, P. Synthesis and antimicrobial activity of vancomycin–conjugated zinc coordination polymer nanoparticles against methicillin-resistant staphylococcus aureus. J. Drug Deliv. Sci. Technol. 2022, 70, 103255. [Google Scholar] [CrossRef]
- Zhang, W.; Taheri-Ledari, R.; Hajizadeh, Z.; Zolfaghari, E.; Ahghari, M.R.; Maleki, A.; Hamblin, M.R.; Tian, Y. Enhanced activity of vancomycin by encapsulation in hybrid magnetic nanoparticles conjugated to a cell-penetrating peptide. Nanoscale 2020, 12, 3855–3870. [Google Scholar] [CrossRef]
- Akbarzadeh, A.; Rezaei-Sadabady, R.; Davaran, S.; Joo, S.W.; Zarghami, N.; Hanifehpour, Y.; Samiei, M.; Kouhi, M.; Nejati-Koshki, K. Liposome: Classification, preparation, and applications. Nanoscale Res. Lett. 2013, 8, 102. [Google Scholar] [CrossRef]
- Bozzuto, G.; Molinari, A. Liposomes as nanomedical devices. Int. J. Nanomed. 2015, 10, 975–999. [Google Scholar] [CrossRef]
- Gao, A.; Hu, X.; Saeed, M.; Chen, B.; Li, Y.; Yu, H. Overview of recent advances in liposomal nanoparticle-based cancer immunotherapy. Acta Pharmacol. Sin. 2019, 40, 1129–1137. [Google Scholar] [CrossRef]
- Torchilin, V.P. Recent advances with liposomes as pharmaceutical carriers. Nat. Rev. Drug Discov. 2005, 4, 145–160. [Google Scholar] [CrossRef]
- Tasharrofi, N.; Nourozi, M.; Marzban, A. How liposomes pave the way for ocular drug delivery after topical administration. J. Drug Deliv. Sci. Technol. 2021, 67, 103045. [Google Scholar] [CrossRef]
- Kadry, A.A.; Al-Suwayeh, S.A.; Abd-Allah, A.; Bayomi, M.A. Treatment of experimental osteomyelitis by liposomal antibiotics. J. Antimicrob. Chemother. 2004, 54, 1103–1108. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, Z.; Li, F.; Gao, J.; Wang, L.; Huang, G. Liposomes for systematic delivery of vancomycin hydrochloride to decrease nephrotoxicity: Characterization and evaluation. Asian J. Pharm. Sci. 2015, 10, 212–222. [Google Scholar] [CrossRef]
- Scriboni, A.B.; Couto, V.M.; Ribeiro, L.N.D.M.; Freires, I.A.; Groppo, F.C.; De Paula, E.; Franz-Montan, M.; Cogo-Müller, K. Fusogenic Liposomes Increase the Antimicrobial Activity of Vancomycin Against Staphylococcus aureus Biofilm. Front. Pharmacol. 2019, 10, 1401. [Google Scholar] [CrossRef] [PubMed]
- Uhl, P.; Sauter, M.; Hertlein, T.; Witzigmann, D.; Laffleur, F.; Hofhaus, G.; Fidelj, V.; Tursch, A.; Özbek, S.; Hopke, E.; et al. Overcoming the Mucosal Barrier: Tetraether Lipid-Stabilized Liposomal Nanocarriers Decorated with Cell-Penetrating Peptides Enable Oral Delivery of Vancomycin. Adv. Ther. 2021, 4, 2000247. [Google Scholar] [CrossRef]
- Muppidi, K.; Pumerantz, A.S.; Wang, J.; Betageri, G. Development and Stability Studies of Novel Liposomal Vancomycin Formulations. ISRN 2012, 2012, 1–8. [Google Scholar] [CrossRef]
- Bolotin, E.M.; Cohen, R.; Bar, L.K.; Emanuel, N.; Ninio, S.; Barenholz, Y.; Lasic, D.D. Ammonium Sulfate Gradients for Efficient and Stable Remote Loading of Amphipathic Weak Bases into Liposomes and Ligandoliposomes. J. Liposome Res. 1994, 4, 455–479. [Google Scholar] [CrossRef]
- Li, S.; Goins, B.; Phillips, W.T.; Bao, A. Remote-loading labeling of liposomes with99mTc-BMEDA and its stability evaluation: Effects of lipid formulation and pH/chemical gradient. J. Liposome Res. 2010, 21, 17–27. [Google Scholar] [CrossRef]
- Odeh, F.; Nsairat, H.; Alshaer, W.; Alsotari, S.; Buqaien, R.; Ismail, S.; Awidi, A.; Al Bawab, A. Remote loading of curcumin-in-modified β-cyclodextrins into liposomes using a transmembrane pH gradient. RSC Adv. 2019, 9, 37148–37161. [Google Scholar] [CrossRef]
- Alyane, M.; Barratt, G.; Lahouel, M. Remote loading of doxorubicin into liposomes by transmembrane pH gradient to reduce toxicity toward H9c2 cells. Saudi Pharm. J. 2015, 24, 165–175. [Google Scholar] [CrossRef]
- Gubernator, J. Active methods of drug loading into liposomes: Recent strategies for stable drug entrapment and increased in vivo activity. Expert Opin. Drug Deliv. 2011, 8, 565–580. [Google Scholar] [CrossRef] [PubMed]
- Haran, G.; Cohen, R.; Bar, L.K.; Barenholz, Y. Transmembrane ammonium sulfate gradients in liposomes produce efficient and stable entrapment of amphipathic weak bases. Biochim. Biophys. Acta (BBA)-Biomembr. 1993, 1151, 201–215. [Google Scholar] [CrossRef]
- Hwang, S.; Maitani, Y.; Qi, X.-R.; Takayama, K.; Nagai, T. Remote loading of diclofenac, insulin and fluorescein isothiocyanate labeled insulin into liposomes by pH and acetate gradient methods. Int. J. Pharm. 1999, 179, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H. Thin-film hydration followed by extrusion method for liposome preparation. Methods Mol. Biol. 2017, 1522, 17–22. [Google Scholar] [CrossRef]
- Chu, Y.H.; Whitesides, G.M. Affinity capillary electrophoresis can simultaneously measure binding constants of multiple peptides to vancomycin. J. Org. Chem. 1992, 57, 3524–3525. [Google Scholar] [CrossRef]
- Takács-Novák, K.; Noszál, B.; Tókés-Kövesdi, M.; Szász, G. Acid-base properties and proton-speciation of vancomycin. Int. J. Pharm. 1993, 89, 261–263. [Google Scholar] [CrossRef]
- Yang, Z.; Vorpagel, E.R.; Laskin, J. Influence of the Charge State on the Structures and Interactions of Vancomycin Antibiotics with Cell-Wall Analogue Peptides: Experimental and Theoretical Studies. Chem. A Eur. J. 2009, 15, 2081–2090. [Google Scholar] [CrossRef]
- Adamala, K.; Engelhart, A.E.; Kamat, N.P.; Jin, L.; Szostak, J.W. Construction of a Liposome Dialyzer for preparation of high-value, small-volume liposome formulations. Nat. Protoc. 2015, 10, 927. [Google Scholar] [CrossRef]
- Has, C.; Sunthar, P. A comprehensive review on recent preparation techniques of liposomes. J. Liposome Res. 2019, 30, 336–365. [Google Scholar] [CrossRef]
- Chen, P.S.; Toribara, T.Y.; Warner, H. Microdetermination of Phosphorus. Anal. Chem. 1956, 28, 1756–1758. [Google Scholar] [CrossRef]
- Deepa, B.; Balasubramanian, N.; Nagaraja, K.S. Spectrophotometric Determination of Cyanide Based on Berthelot Reaction. Anal. Lett. 2003, 36, 2865–2874. [Google Scholar] [CrossRef]
- Zou, Z.; Sun, J.; Li, Q.; Pu, Y.; Liu, J.; Sun, R.; Wang, L.; Jiang, T. Vancomycin modified copper sulfide nanoparticles for photokilling of vancomycin-resistant enterococci bacteria. Colloids Surf. B Biointerfaces 2020, 189, 110875. [Google Scholar] [CrossRef]
- Cao, M.; Feng, Y.; Zhang, Y.; Kang, W.; Lian, K.; Ai, L. Studies on the metabolism and degradation of vancomycin in simulated in vitro and aquatic environment by UHPLC-Triple-TOF-MS/MS. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Sedeek, M.S.; Afifi, S.M.; Mansour, M.K.; Hassan, M.; Mehaya, F.M.; Naguib, I.A.; Abourehab, M.A.; Farag, M.A. Unveiling Antimicrobial and Antioxidant Compositional Differences between Dukkah and Za’atar via SPME-GCMS and HPLC-DAD. Molecules 2022, 27, 6471. [Google Scholar] [CrossRef] [PubMed]
- Humphries, R.M.; Ambler, J.; Mitchell, S.L.; Castanheira, M.; Dingle, T.; Hindler, J.A.; Koeth, L.; Sei, K.; on behalf of the CLSI Methods Development and Standardization Working Group of the Subcommittee on Antimicrobial Susceptibility Testing. CLSI Methods Development and Standardization Working Group Best Practices for Evaluation of Antimicrobial Susceptibility Tests. J. Clin. Microbiol. 2018, 56, e01934-17. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, N.M.; Fahim, S.H.; Hassan, M.; Farag, A.E.; Georgey, H.H. Design and synthesis of ciprofloxacin-sulfonamide hybrids to manipulate ciprofloxacin pharmacological qualities: Potency and side effects. Eur. J. Med. Chem. 2021, 228, 114021. [Google Scholar] [CrossRef] [PubMed]
- Salem, M.A.; El-Shiekh, R.A.; Hashem, R.A.; Hassan, M. In vivo antibacterial activity of star anise (Illicium verum hook.) extract using murine mrsa skin infection model in relation to its metabolite profile. Infect. Drug Resist. 2021, 14, 33–48. [Google Scholar] [CrossRef] [PubMed]
- Albash, R.; Abdellatif, M.M.; Hassan, M.; Badawi, N.M. Tailoring Terpesomes and Leciplex for the Effective Ocular Conveyance of Moxifloxacin Hydrochloride (Comparative Assessment): In-vitro, Ex-vivo, and In-vivo Evaluation. Int. J. Nanomed. 2021, 16, 5247–5263. [Google Scholar] [CrossRef]
- Farrell, D.J.; Robbins, M.; Rhys-Williams, W.; Love, W.G. Investigation of the potential for mutational resistance to XF-73, retapamulin, mupirocin, fusidic acid, daptomycin, and vancomycin in methicillin-resistant Staphylococcus aureus isolates during a 55-passage study. Antimicrob. Agents Chemother. 2011, 55. [Google Scholar] [CrossRef]
- Mohammad, H.; Younis, W.; Ezzat, H.G.; Peters, C.E.; AbdelKhalek, A.; Cooper, B.; Pogliano, K.; Pogliano, J.; Mayhoub, A.S.; Seleem, M.N. Bacteriological profiling of diphenylureas as a novel class of antibiotics against methicillin-resistant Staphylococcus aureus. PLoS ONE 2017, 12, e0182821. [Google Scholar] [CrossRef]
- Budai, P.; Kormos, É.; Buda, I.; Somody, G.; Lehel, J. Comparative evaluation of HET-CAM and ICE methods for objective assessment of ocular irritation caused by selected pesticide products. Toxicol. In Vitro 2021, 74, 105150. [Google Scholar] [CrossRef]
- McKenzie, B.; Kay, G.; Matthews, K.H.; Knott, R.M.; Cairns, D. The hen’s egg chorioallantoic membrane (HET-CAM) test to predict the ophthalmic irritation potential of a cysteamine-containing gel: Quantification using Photoshop® and ImageJ. Int. J. Pharm. 2015, 490, 1–8. [Google Scholar] [CrossRef]
- Valadares, M.C.; De Oliveira, G.A.R.; De Ávila, R.I.; Da Silva, A.C.G. Strategy Combining Nonanimal Methods for Ocular Toxicity Evaluation. Toxic. Assess. Methods Protoc. 2021, 2240, 175–195. [Google Scholar] [CrossRef]
- Gilleron, L.; Coecke, S.; Sysmans, M.; Hansen, E.; Van Oproy, S.; Marzin, D.; Van Cauteren, H.; Vanparys, P. Evaluation of a modified HET-CAM assay as a screening test for eye irritancy. Toxicol. Vitr. 1996, 10, 431–446. [Google Scholar] [CrossRef]
- Rivero, M.N.; Lenze, M.; Izaguirre, M.; Damonte, S.H.P.; Aguilar, A.; Wikinski, S.; Gutiérrez, M.L. Comparison between HET-CAM protocols and a product use clinical study for eye irritation evaluation of personal care products including cosmetics according to their surfactant composition. Food Chem. Toxicol. 2021, 153, 112229. [Google Scholar] [CrossRef]
- Over, B.; Matsson, P.; Tyrchan, C.; Artursson, P.; Doak, B.C.; A Foley, M.; Hilgendorf, C.; E Johnston, S.; Lee, M.D.; Lewis, R.J.; et al. Structural and conformational determinants of macrocycle cell permeability. Nat. Chem. Biol. 2016, 12, 1065–1074. [Google Scholar] [CrossRef]
- Kamenik, A.S.; Kraml, J.; Hofer, F.; Waibl, F.; Quoika, P.K.; Kahler, U.; Schauperl, M.; Liedl, K.R. Macrocycle Cell Permeability Measured by Solvation Free Energies in Polar and Apolar Environments. J. Chem. Inf. Model. 2020, 60, 3508–3517. [Google Scholar] [CrossRef] [PubMed]
- Bockus, A.T.; Lexa, K.W.; Pye, C.R.; Kalgutkar, A.S.; Gardner, J.W.; Hund, K.C.R.; Hewitt, W.M.; Schwochert, J.A.; Glassey, E.; Price, D.A.; et al. Probing the Physicochemical Boundaries of Cell Permeability and Oral Bioavailability in Lipophilic Macrocycles Inspired by Natural Products. J. Med. Chem. 2015, 58, 4581–4589. [Google Scholar] [CrossRef] [PubMed]
- Begnini, F.; Poongavanam, V.; Atilaw, Y.; Erdelyi, M.; Schiesser, S.; Kihlberg, J. Cell Permeability of Isomeric Macrocycles: Predictions and NMR Studies. ACS Med. Chem. Lett. 2021, 12, 983–990. [Google Scholar] [CrossRef] [PubMed]
- Phillips-Jones, M.K.; Lithgo, R.; Dinu, V.; Gillis, R.B.; Harding, J.E.; Adams, G.G.; Harding, S.E. Full hydrodynamic reversibility of the weak dimerization of vancomycin and elucidation of its interaction with VanS monomers at clinical concentration. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Jia, Z.; O’Mara, M.L.; Zuegg, J.; Cooper, M.A.; Mark, A.E. Vancomycin: Ligand recognition, dimerization and super-complex formation. FEBS J. 2013, 280, 1294–1307. [Google Scholar] [CrossRef]
- Loll, P.J.; Miller, R.; Weeks, C.M.; Axelsen, P.H. A ligand-mediated dimerization mode for vancomycin. Chem. Biol. 1998, 5, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Nitanai, Y.; Kikuchi, T.; Kakoi, K.; Hanamaki, S.; Fujisawa, I.; Aoki, K. Crystal Structures of the Complexes between Vancomycin and Cell-Wall Precursor Analogs. J. Mol. Biol. 2008, 385, 1422–1432. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, M.; Schneider, T.R.; Shieldrick, G.M. Crystal structure of vancomycin. Structure 1996, 4, 1509–1515. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.H.; Williamson, M.P.; Butcher, D.W.; Hammond, S.J. Detailed binding sites of the antibiotics vancomycin and ristocetin A: Determination of intermolecular distances in antibiotic/substrate complexes by use of the time-dependent NOE. J. Am. Chem. Soc. 1983, 105, 1332–1339. [Google Scholar] [CrossRef]
- Fan, Q.; Li, L.; Xue, H.; Zhou, H.; Zhao, L.; Liu, J.; Mao, J.; Wu, S.; Zhang, S.; Wu, C.; et al. Precise control over kinetics of molecular assembly: Production of particles with tunable sizes and crystalline forms. Angew. Chem. 2020, 132, 15253–15258. [Google Scholar] [CrossRef]
- Šípová-Jungová, H.; Jurgová, L.; Hemmerová, E.; Homola, J. Interaction of Tris with DNA molecules and carboxylic groups on self-assembled monolayers of alkanethiols measured with surface plasmon resonance. Appl. Surf. Sci. 2021, 546, 148984. [Google Scholar] [CrossRef]
- Dinu, V.; Lu, Y.; Weston, N.; Lithgo, R.; Coupe, H.; Channell, G.; Adams, G.G.; Gómez, A.T.; Sabater, C.; Mackie, A.; et al. The antibiotic vancomycin induces complexation and aggregation of gastrointestinal and submaxillary mucins. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Gallová, J.; Uhríková, D.; Hanulová, M.; Teixeira, J.; Balgavý, P. Bilayer thickness in unilamellar extruded 1,2-dimyristoleoyl and 1,2-dierucoyl phosphatidylcholine vesicles: SANS contrast variation study of cholesterol effect. Colloids Surf. B Biointerfaces 2004, 38, 11–14. [Google Scholar] [CrossRef]
- Hsueh, Y.-W.; Chen, M.-T.; Patty, P.J.; Code, C.; Cheng, J.; Frisken, B.J.; Zuckermann, M.; Thewalt, J. Ergosterol in POPC Membranes: Physical Properties and Comparison with Structurally Similar Sterols. Biophys. J. 2007, 92, 1606–1615. [Google Scholar] [CrossRef]
- Pencer, J.; Nieh, M.P.; Harroun, T.A.; Krueger, S.; Adams, C.; Katsaras, J. Bilayer thickness and thermal response of dimyristoylphosphatidylcholine unilamellar vesicles containing cholesterol, ergosterol and lanosterol: A small-angle neutron scattering study. Biochim. Biophys. Acta (BBA)-Biomembr. 2005, 1720, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Subczynski, W.K.; Pasenkiewicz-Gierula, M.; Widomska, J.; Mainali, L.; Raguz, M. High Cholesterol/Low Cholesterol: Effects in Biological Membranes: A Review. Cell Biochem. Biophys. 2017, 75, 369–385. [Google Scholar] [CrossRef] [PubMed]
- Bot, F.; Cossuta, D.; O’Mahony, J.A. Inter-relationships between composition, physicochemical properties and functionality of lecithin ingredients. Trends Food Sci. Technol. 2021, 111, 261–270. [Google Scholar] [CrossRef]
- List, G.R. Soybean Lecithin: Food, Industrial Uses, and Other Applications. Polar Lipids Biol. Chem. Technol. 2015, 1–33. [Google Scholar] [CrossRef]
- Hafizi, T.; Shahriari, M.H.; Abdouss, M.; Kahdestani, S.A. Synthesis and characterization of vancomycin-loaded chitosan nanoparticles for drug delivery. Polym. Bull. 2022, 80, 5607–5621. [Google Scholar] [CrossRef]
- El-Shiekh, R.A.; Hassan, M.; Hashem, R.A.; Abdel-Sattar, E. Bioguided Isolation of Antibiofilm and Antibacterial Pregnane Glycosides from Caralluma quadrangula: Disarming Multidrug-Resistant Pathogens. Antibiotics 2021, 10, 811. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formulations | Cholesterol % | Drug:Lipid Ratio | Number Drug Solution Exchange | Entrapment Efficiency % ± SD | Particle Size (nm) ± SD | Zeta Potential (mV) ± SD | PDI |
|---|---|---|---|---|---|---|---|
| F1 | 10 | 0.2 | 3 | 32.14 ± 2.0 | 141 ± 2.7 | −21 ± 1.8 | 0.20 |
| F2 | 30 | 0.05 | 3 | 37.06 ± 1.5 | 282 ± 8.0 | −38 ± 0.7 | 0.37 |
| F3 | 10 | 0.05 | 1 | 50.34 ± 2.4 | 149 ± 9.2 | −30 ± 1.0 | 0.21 |
| F4 | 10 | 0.05 | 3 | 33.58 ± 1.7 | 153 ± 3.2 | −27 ± 0.9 | 0.18 |
| F5 | 10 | 0.2 | 1 | 41.09 ± 2.5 | 157 ± 2.0 | −28.5 ± 1.2 | 0.18 |
| F6 | 30 | 0.2 | 3 | 30.56 ± 1.7 | 353 ± 3.5 | −41 ± 0.5 | 0.42 |
| F7 | 30 | 0.2 | 1 | 32.09 ± 1.5 | 316 ± 10 | −32 ± 2.0 | 0.30 |
| F8 | 30 | 0.2 | 2 | 41.24 ± 8.5 | 307 ± 5.3 | −18.5 ± 1.2 | 0.23 |
| F9 | 10 | 0.05 | 2 | 53.46 ± 2.9 | 148 ± 4.0 | −30 ± 0.7 | 0.18 |
| F10 | 20 | 0.05 | 2 | 65.00 ± 4.0 | 155 ± 5.4 | −34 ± 1.5 | 0.25 |
| Bacteria | Description | Vancomycin Solution | Vancomycin-Loaded Liposomes | ||
|---|---|---|---|---|---|
| MIC (μg/mL) | MBC (μg/mL) | MIC (μg/mL) | MBC (μg/mL) | ||
| Staphylococcus aureus (MRSA USA300) | Methicillin-resistant/Vancomycin senstive | 1.563 ± 0.0 | 2.083 ± 0.9 | 0.338 ± 0.0 | 0.338 ± 0.0 |
| Staphylococcus aureus (hVISA OP933674) | Heteroresistant vancomycin-intermediate Staphylococcus aureus | 2.083 ± 0.9 | 6.25 ± 0.0 | 0.338 ± 0.0 | 0.338 ± 0.0 |
| Staphylococcus epidermidis ATCC 35984 | Biofilm positive | 1.563 ± 0.0 | 1.563 ± 0.0 | 0.338 ± 0.0 | 0.338 ± 0.0 |
| Staphylococcus epidermidis ATCC 12228 | Biofilm negative | 1.563 ± 0.0 | 1.563 ± 0.0 | 0.451 ± 0.2 | 0.451 ± 0.2 |
| Bacteria | Number of Passages | Vancomycin Solution | Vancomyci-Loaded Liposomes | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Start Passage | Last Passage | Start Passage | Last Passage | ||||||
| MIC (μg/mL) | MBC (μg/mL) | MIC (μg/mL) | MBC (μg/mL) | MIC (μg/mL) | MBC (μg/mL) | MIC (μg/mL) | MBC (μg/mL) | ||
| S. aureus (MRSA USA300) | 25 | 1.563 ± 0 | 2.083 ± 0.9 | 8.33 ± 3.61 | 8.33 ± 3.61 | 0.338 ± 0 | 0.338 ± 0 | 0.338 ± 0 | 0.338 ± 0.0 |
| S. epidermidis ATCC 35984 | 25 | 1.563 ± 0 | 1.563 ± 0.0 | 8.33 ± 3.61 | 8.33 ± 3.61 | 0.338 ± 0 | 0.338 ± 0 | 0.338 ± 0 | 0.451 ± 0.195 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, E.T.M.; Hassan, M.; Shamma, R.N.; Makky, A.; Hassan, D.H. Controlling the Evolution of Selective Vancomycin Resistance through Successful Ophthalmic Eye-Drop Preparation of Vancomycin-Loaded Nanoliposomes Using the Active-Loading Method. Pharmaceutics 2023, 15, 1636. https://doi.org/10.3390/pharmaceutics15061636
Ahmed ETM, Hassan M, Shamma RN, Makky A, Hassan DH. Controlling the Evolution of Selective Vancomycin Resistance through Successful Ophthalmic Eye-Drop Preparation of Vancomycin-Loaded Nanoliposomes Using the Active-Loading Method. Pharmaceutics. 2023; 15(6):1636. https://doi.org/10.3390/pharmaceutics15061636
Chicago/Turabian StyleAhmed, El Tahra M., Mariam Hassan, Rehab Nabil Shamma, Amna Makky, and Doaa H. Hassan. 2023. "Controlling the Evolution of Selective Vancomycin Resistance through Successful Ophthalmic Eye-Drop Preparation of Vancomycin-Loaded Nanoliposomes Using the Active-Loading Method" Pharmaceutics 15, no. 6: 1636. https://doi.org/10.3390/pharmaceutics15061636