
Differential Expression of ABC Transporter Genes in Brain Vessels vs. Peripheral Tissues and Vessels from Human, Mouse and Rat
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human and Rodent Tissues
2.2. Isolation of Microvessels and Capillaries from Tissues
2.3. RNA Isolation
2.4. RNA-Seq
2.5. Public Datasets and Analysis
2.6. Automated Western Blot Analysis (WesTM)
2.7. Statistical Analysis
3. Results
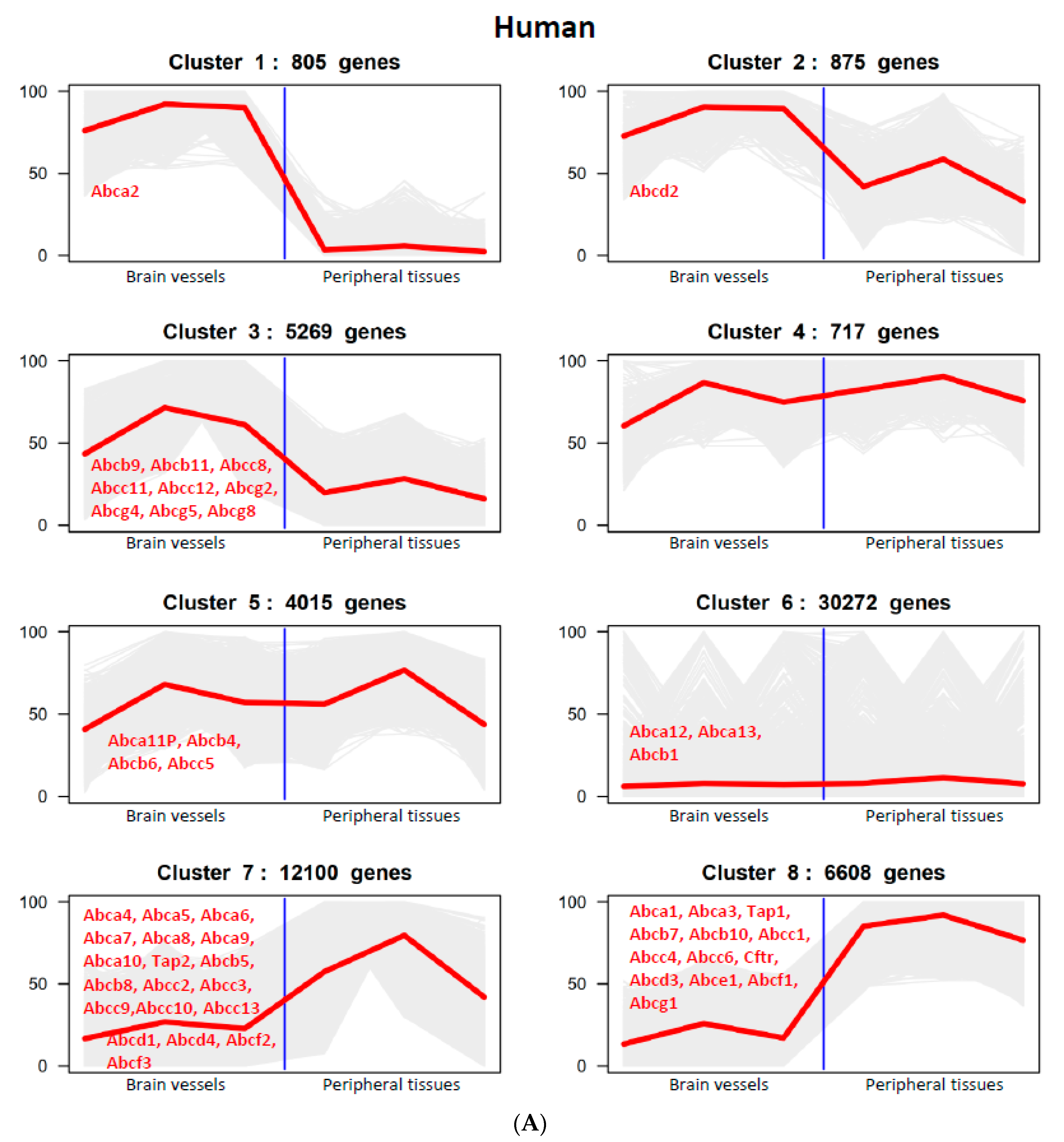
3.1. RNA-Seq Datasets: Quality, Comparability and Validation
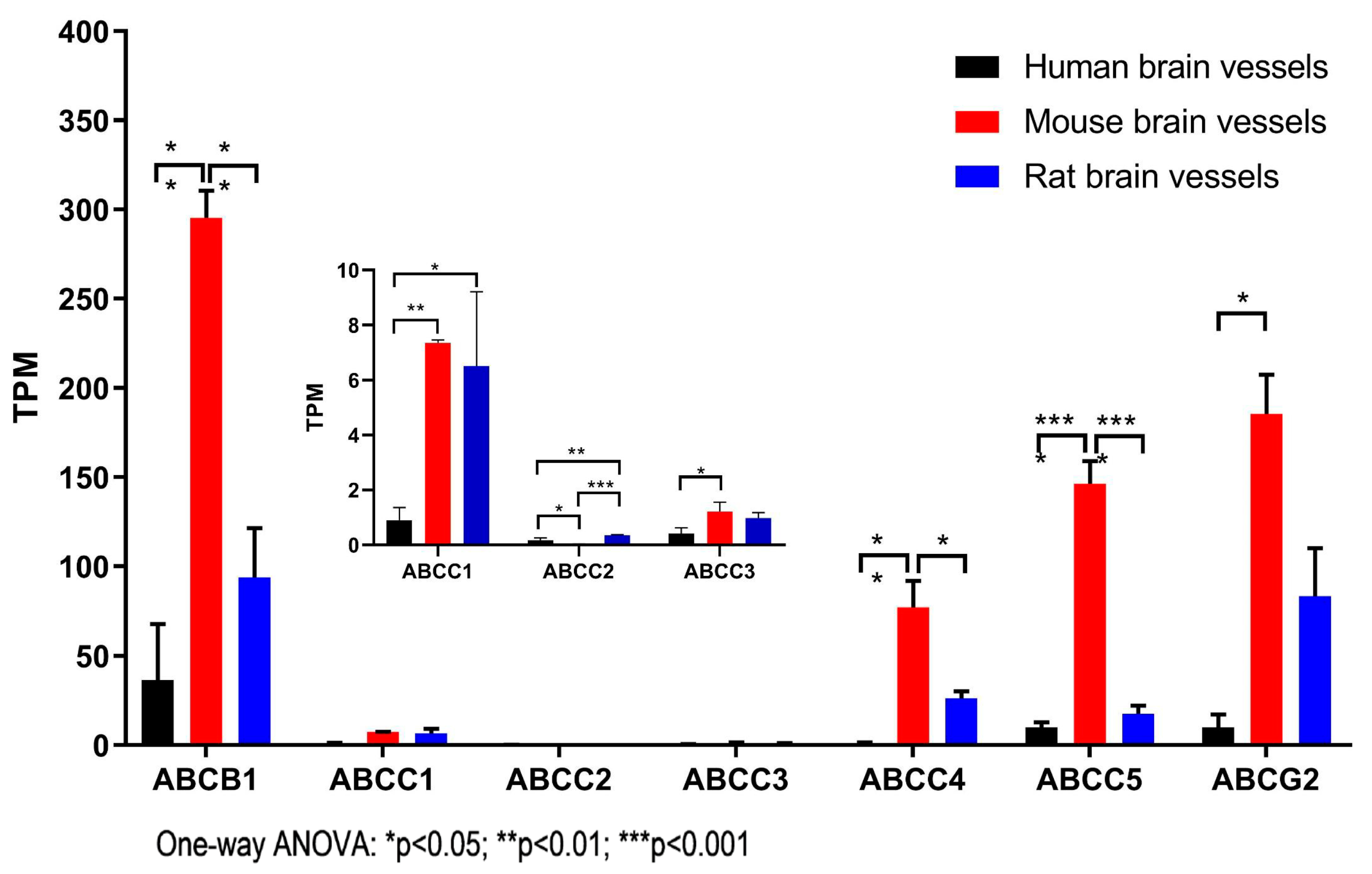
3.2. The Expression of ABC Transporter Genes in Brain Vessels across Species
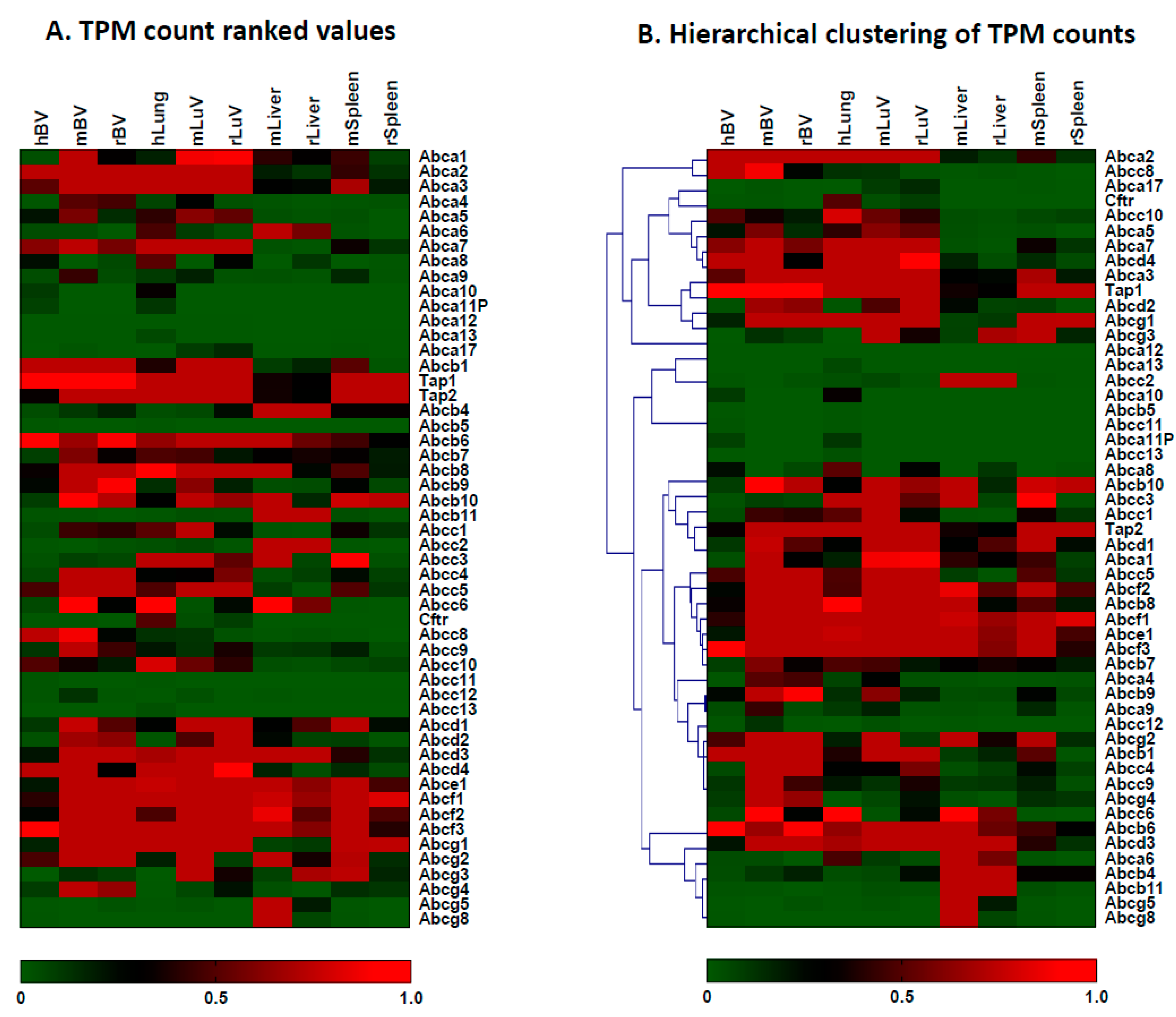
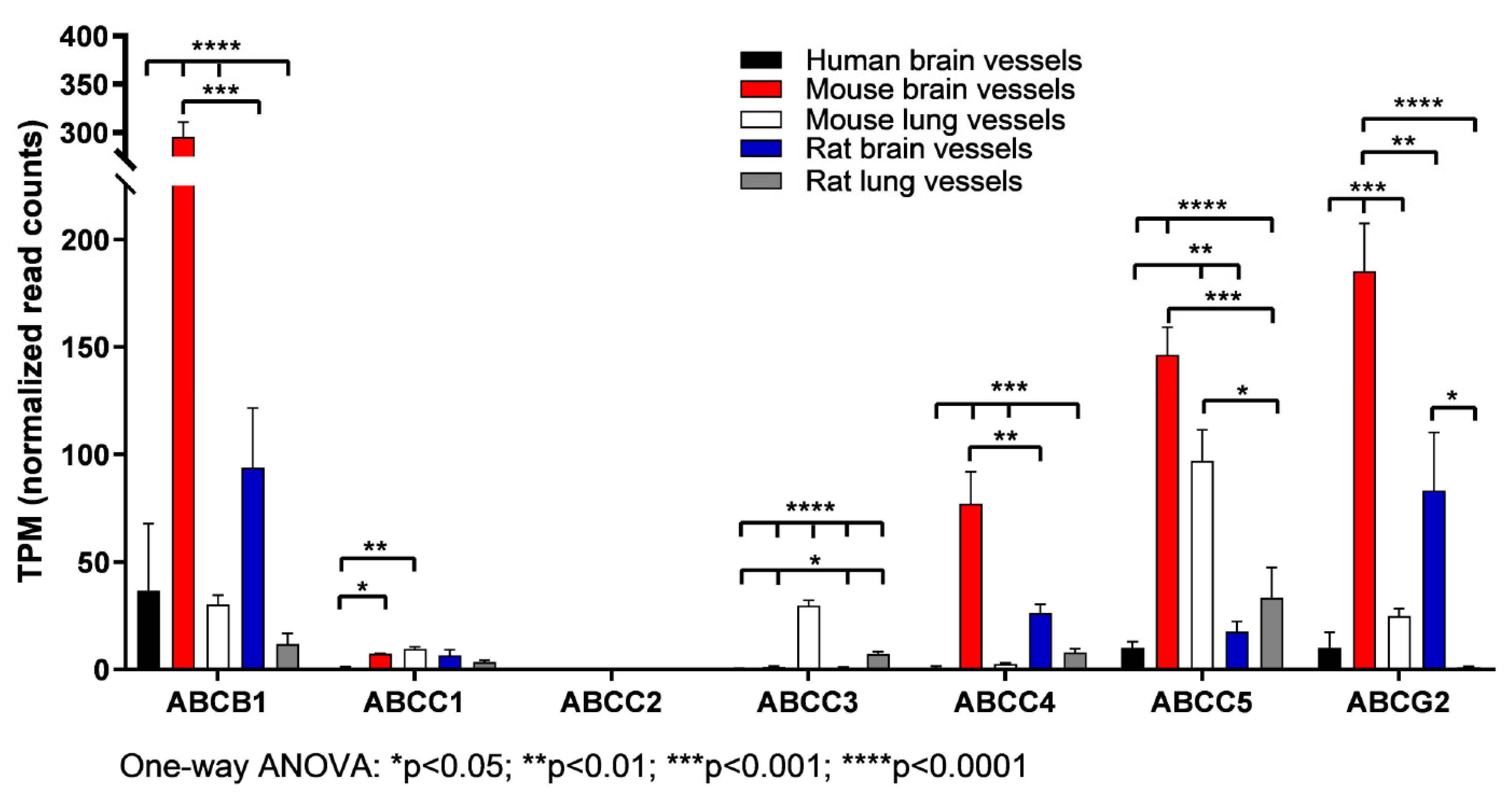
3.3. The Expression of ABC Transporter Genes in Brain Vessels vs. Lung Vessels, Liver and Spleen
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dean, M.; Rzhetsky, A.; Allikmets, R. The human ATP-binding cassette (ABC) transporter superfamily. Genome Res. 2001, 11, 1156–1166. [Google Scholar] [CrossRef] [PubMed]
- Dean, M.; Moitra, K.; Allikmets, R. The human ATP-binding cassette (ABC) transporter superfamily. Hum. Mutat. 2022, 43, 1162–1182. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Bamji-Mirza, M.; Chang, N.; Haqqani, A.; Stanimirovic, D.B. Expression and Function of ABC Transporters at the Blood-Brain Barrier. In The Blood-Brain Barrier in Health and Disease; Dorovini-Zis, K., Ed.; CRC press/Taylor & Francis Group: New York, NY, USA, 2015; Volume one, pp. 172–214. [Google Scholar]
- Dezi, M.; Fribourg, P.F.; Di Cicco, A.; Arnaud, O.; Marco, S.; Falson, P.; Di Pietro, A.; Lévy, D. The multidrug resistance half-transporter ABCG2 is purified as a tetramer upon selective extraction from membranes. Biochim. Biophys. Acta 2010, 1798, 2094–2101. [Google Scholar] [CrossRef] [PubMed]
- Shimabuku, A.M.; Nishimoto, T.; Ueda, K.; Komano, T. P-glycoprotein. ATP hydrolysis by the N-terminal nucleotide-binding domain. J. Biol. Chem. 1992, 267, 4308–4311. [Google Scholar]
- Shen, S.; Zhang, W. ABC transporters and drug efflux at the blood-brain barrier. Rev. Neurosci. 2010, 21, 29–53. [Google Scholar] [CrossRef]
- Redzic, Z. Molecular biology of the blood-brain and the blood-cerebrospinal fluid barriers: Similarities and differences. Fluids Barriers CNS 2011, 8, 3. [Google Scholar] [CrossRef]
- Miller, D.S. Regulation of ABC transporters at the blood-brain barrier. Clin. Pharmacol. Ther. 2015, 97, 395–403. [Google Scholar] [CrossRef]
- Zhang, W.; Stanimirovic, D.B. The transport systems of the blood-brain barrier. In The Blood-Brain Barrier and Its Microenvironment: Basic Physiology to Neurological Disease; De Vries, E., Prat, A., Eds.; Taylor & Francis Group: New York, NY, USA, 2005; pp. 103–142. [Google Scholar]
- Han, L.W.; Gao, C.; Mao, Q. An update on expression and function of P-gp/ABCB1 and BCRP/ABCG2 in the placenta and fetus. Expert Opin. Drug Metab. Toxicol. 2018, 14, 817–829. [Google Scholar] [CrossRef]
- Abbott, N.J.; Patabendige, A.A.; Dolman, D.E.; Yusof, S.R.; Begley, D.J. Structure and function of the blood-brain barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef]
- Miller, D.S. Regulation of P-glycoprotein and other ABC drug transporters at the blood-brain barrier. Trends Pharmacol. Sci. 2010, 31, 246–254. [Google Scholar] [CrossRef]
- Terstappen, G.; Meyer, A.; Bell, R.; Zhang, W. Strategies for delivering central nervous system therapeutics across the blood-brain barrier. Nat. Rev. Drug Discov. 2021, 20, 362–383. [Google Scholar] [CrossRef] [PubMed]
- Schinkel, A.H.; Mayer, U.; Wagenaar, E.; Mol, C.A.; van Deemter, L.; Smit, J.J.; van der Valk, M.A.; Voordouw, A.C.; Spits, H.; van Tellingen, O.; et al. Normal viability and altered pharmacokinetics in mice lacking mdr1-type (drug-transporting) P-glycoproteins. Proc. Natl. Acad. Sci. USA 1997, 94, 4028–4033. [Google Scholar] [CrossRef] [PubMed]
- Wijnholds, J.; deLange, E.C.; Scheffer, G.L.; van den Berg, D.J.; Mol, C.A.; van der Valk, M.; Schinkel, A.H.; Scheper, R.J.; Breimer, D.D.; Borst, P. Multidrug resistance protein 1 protects the choroid plexus epithelium and contributes to the blood-cerebrospinal fluid barrier. J. Clin. Investig. 2000, 105, 279–285. [Google Scholar] [CrossRef]
- Zhang, W.; Xiong, H.; Callaghan, D.; Jones, A.; Pei, K.; Stanimirovic, D. Blood-brain barrier transport of amyloid beta peptides in efflux pump knock-out animals evaluated by in vivo optical imaging. Fluids Barriers CNS 2013, 10, 13. [Google Scholar] [CrossRef]
- Wang, J.; Gan, C.; Retmana, I.A.; Sparidans, R.W.; Li, W.; Lebre, M.C.; Beijnen, J.H.; Schinkel, A.H. P-glycoprotein (MDR1/ABCB1) and Breast Cancer Resistance Protein (BCRP/ABCG2) limit brain accumulation of the FLT3 inhibitor quizartinib in mice. Int. J. Pharm. 2019, 556, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Mak, I.W.; Evaniew, N.; Ghert, M. Lost in translation: Animal models and clinical trials in cancer treatment. Am. J. Transl. Res. 2014, 6, 114–118. [Google Scholar]
- Morris, M.E.; Rodriguez-Cruz, V.; Felmlee, M.A. SLC and ABC Transporters: Expression, Localization, and Species Differences at the Blood-Brain and the Blood-Cerebrospinal Fluid Barriers. AAPS J. 2017, 19, 1317–1331. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, Q.Y.; Haqqani, A.S.; Leclerc, S.; Baumann, E.; Ly, D.; Delaney, C.E.; Liu, Z.-Y.; Star, A.T.; Brunette, E.; et al. Differential expression of receptors mediating receptor-mediated transcytosis (RMT) in brain microvessels, brain parenchyma and peripheral tissues of mouse and human. Fluids Barriers CNS 2020, 17, 47. [Google Scholar] [CrossRef]
- Stanimirovic, D.B.; Bani-Yaghoub, M.; Perkins, M.; Haqqani, A.S. Blood–brain barrier models: In vitro to in vivo translation in preclinical development of CNS-targeting biotherapeutics. Expert Opin. Drug Discov. 2015, 10, 141–155. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Leinonen, R.; Sugawara, H.; Shumway, M. International nucleotide sequence database collaboration. The sequence read archive. Nucleic Acids Res. 2011, 39, D19–D21. [Google Scholar] [CrossRef] [PubMed]
- Grossman, R.L.; Heath, A.; Murphy, M.; Patterson, M.; Wells, W. A case for data commons: Toward data science as a service. Comput. Sci. Eng. 2016, 18, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Zepeda, D.; Taghi, M.; Scherrmann, J.M.; Decleves, X.; Menet, M.C. ABC Transporters at the Blood-Brain Interfaces, Their Study Models, and Drug Delivery Implications in Gliomas. Pharmaceutics 2019, 12, 20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Mojsilovic-Petrovic, J.; Andrade, M.; Zhang, H.; Ball, M.; Stanimirovic, D. Expression and Functional Characterization of ABCG2 in Brain Endothelial Cells and Vessels. FASEB J. 2003, 17, 2085–2087. [Google Scholar] [CrossRef]
- Hirsch-Reinshagen, V.; Zhou, S.; Burgess, B.L.; Bernier, L.; McIsaac, S.A.; Chan, J.Y.; Tansley, G.; Cohn, J.S.; Hayden, M.R.; Wellington, C.L. Deficiency of ABCA1 impairs apolipoprotein E metabolism in brain. J. Biol. Chem. 2004, 279, 41197–41207. [Google Scholar] [CrossRef] [PubMed]
- Karasinska, J.M.; de Haan, W.; Franciosi, S.; Ruddle, P.; Fan, J.; Kruit, J.K.; Stukas, S.; Lütjohann, D.; Gutmann, D.H.; Wellington, C.L.; et al. ABCA1 influences neuroinflammation and neuronal death. Neurobiol. Dis. 2013, 54, 445–455. [Google Scholar] [CrossRef]
- Davis, W., Jr.; Tew, K.D. ATP-binding cassette transporter-2 (ABCA2) as a therapeutic target. Biochem. Pharmacol. 2018, 151, 188–200. [Google Scholar] [CrossRef]
- Stahlman, M.T.; Besnard, V.; Wert, S.E.; Weaver, T.E.; Dingle, S.; Xu, Y.; von Zychlin, K.; Olson, S.J.; Whitsett, J.A. Expression of ABCA3 in developing lung and other tissues. J. Histochem. Cytochem. 2007, 55, 71–83. [Google Scholar] [CrossRef]
- Steinbach, D.; Gillet, J.P.; Sauerbrey, A.; Gruhn, B.; Dawczynski, K.; Bertholet, V.; de Longueville, F.; Zintl, F.; Remacle, J.; Efferth, T. ABCA3 as a possible cause of drug resistance in childhood acute myeloid leukemia. Clin. Cancer Res. 2006, 12 Pt 1, 4357–4363. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Lan, D.; Gerbod-Giannone, M.; Linsel-Nitschke, P.; Jehle, A.W.; Chen, W.; Martinez, L.O.; Tall, A.R. ATP-binding cassette transporter A7 (ABCA7) binds apolipoprotein A-I and mediates cellular phospholipid but not cholesterol efflux. J. Biol. Chem. 2003, 278, 42906–42912. [Google Scholar] [CrossRef] [PubMed]
- Trigueros-Motos, L.; van Capelleveen, J.C.; Torta, F.; Castaño, D.; Zhang, L.H.; Chai, E.C.; Kang, M.; Dimova, L.G.; Schimmel, A.W.M.; Tietjen, I.; et al. ABCA8 Regulates Cholesterol Efflux and High-Density Lipoprotein Cholesterol Levels. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 2147–2155. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, J.; Suarez, M.D.; Yadav, P.K.; Walsh, M.T.; Li, Y.; Wu, Y.; Huang, Z.; James, A.W.; Escobar, V.; Mokbe, A.; et al. ATP-binding cassette protein ABCA7 deficiency impairs sphingomyelin synthesis, cognitive discrimination, and synaptic plasticity in the entorhinal cortex. J. Biol. Chem. 2022, 298, 102411. [Google Scholar] [CrossRef] [PubMed]
- Bamji-Mirza, M.; Li, Y.; Najem, D.; Liu, Q.-Y.; Stupak, J.; Chen, K.; Li, J.; Walker, D.; Lue, L.-F.; Yang, Z.; et al. Genetic variations in ABCA7 can increase secreted levels of Aβ1-42 and ABCA7 transcription in cell culture models. J. Alzheimer’s Dis. 2016, 53, 875–892, Erratum in J. Alzheimer’s Dis. 2018, 66, 853–854. [Google Scholar] [CrossRef]
- Ma, F.C.; Wang, H.F.; Cao, X.P.; Tan, C.C.; Tan, L.; Yu, J.T. Meta-Analysis of the Association between Variants in ABCA7 and Alzheimer’s Disease. J. Alzheimer’s Dis. 2018, 63, 1261–1267. [Google Scholar] [CrossRef]
- Sakae, N.; Liu, C.C.; Shinohara, M.; Frisch-Daiello, J.; Ma, L.; Yamazaki, Y.; Tachibana, M.; Younkin, L.; Kurti, A.; Carrasquillo, M.M.; et al. ABCA7 Deficiency Accelerates Amyloid-β Generation and Alzheimer’s Neuronal Pathology. J. Neurosci. 2016, 36, 3848–3859. [Google Scholar] [CrossRef]
- Abe-Dohmae, S.; Yokoyama, S. ABCA7 links sterol metabolism to the host defense system: Molecular background for potential management measure of Alzheimer’s disease. Gene 2021, 768, 145316. [Google Scholar] [CrossRef]
- Ichikawa, Y.; Bayeva, M.; Ghanefar, M.; Potini, V.; Sun, L.; Mutharasan, R.K.; Wu, R.; Khechaduri, A.; Jairaj Naik, T.; Ardehali, H. Disruption of ATP-binding cassette B8 in mice leads to cardiomyopathy through a decrease in mitochondrial iron export. Proc. Natl. Acad. Sci. USA 2012, 109, 4152–4157. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, W.; Liu, L.; Fisher, C.L.; Hui, D.; Childs, S.; Dorovini-Zis, K.; Ling, V. Characterization of ABCB9, an ATP binding cassette protein associated with lysosomes. J. Biol. Chem. 2000, 275, 23287–23294. [Google Scholar] [CrossRef]
- Park, J.G.; Kim, S.; Jang, E.; Choi, S.H.; Han, H.; Ju, S.; Kim, J.W.; Min, D.S.; Jin, M.S. The lysosomal transporter TAPL has a dual role as peptide translocator and phosphatidylserine floppase. Nat. Commun. 2022, 13, 5851. [Google Scholar] [CrossRef] [PubMed]
- Bryan, J.; Muñoz, A.; Zhang, X.; Düfer, M.; Drews, G.; Krippeit-Drews, P.; Aguilar-Bryan, L. ABCC8 and ABCC9: ABC transporters that regulate K+ channels. Pflug. Arch. 2007, 453, 703–718. [Google Scholar] [CrossRef] [PubMed]
- Tansley, G.H.; Burgess, B.L.; Bryan, M.T.; Su, Y.; Hirsch-Reinshagen, V.; Pearce, J.; Chan, J.Y.; Wilkinson, A.; Evans, J.; Naus, K.E.; et al. The cholesterol transporter ABCG1 modulates the subcellular distribution and proteolytic processing of beta-amyloid precursor protein. J. Lipid Res. 2007, 48, 1022–1034. [Google Scholar] [CrossRef]
- Wang, N.; Yvan-Charvet, L.; Lütjohann, D.; Mulder, M.; Vanmierlo, T.; Kim, T.W.; Tall, A.R. ATP-binding cassette transporters G1 and G4 mediate cholesterol and desmosterol efflux to HDL and regulate sterol accumulation in the brain. FASEB J. 2008, 22, 1073–1082. [Google Scholar] [CrossRef] [PubMed]
- Mickley, L.; Jain, P.; Miyake, K.; Schriml, L.M.; Rao, K.; Fojo, T.; Bates, S.; Dean, M. An ATP-binding cassette gene (ABCG3) closely related to the multidrug transporter ABCG2 (MXR/ABCP) has an unusual ATP-binding domain. Mamm. Genome 2001, 12, 86–88. [Google Scholar] [CrossRef]
- Uchida, Y.; Ohtsuki, S.; Katsukura, Y.; Ikeda, C.; Suzuki, T.; Kamiie, J.; Terasaki, T. Quantitative targeted absolute proteomics of human blood-brain barrier transporters and receptors. J. Neurochem. 2011, 117, 333–345. [Google Scholar] [CrossRef]
- Uchida, Y.; Zhang, Z.; Tachikawa, M.; Terasaki, T. Quantitative targeted absolute proteomics of rat blood-cerebrospinal fluid barrier transporters: Comparison with a human specimen. J. Neurochem. 2015, 134, 1104–1115. [Google Scholar] [CrossRef]
- Ito, K.; Uchida, Y.; Ohtsuki, S.; Aizawa, S.; Kawakami, H.; Katsukura, Y.; Kamiie, J.; Terasaki, T. Quantitative membrane protein expression at the blood-brain barrier of adult and younger cynomolgus monkeys. J. Pharm. Sci. 2011, 100, 3939–3950. [Google Scholar] [CrossRef]
- Wang, L.; Prasad, B.; Salphati, L.; Chu, X.; Gupta, A.; Hop, C.E.; Evers, R.; Unadkat, J.D. Interspecies variability in expression of hepatobiliary transporters across human, dog, monkey, and rat as determined by quantitative proteomics. Drug Metab. Dispos. 2015, 43, 367–374. [Google Scholar] [CrossRef]
- Fallon, J.K.; Smith, P.C.; Xia, C.Q.; Kim, M.S. Quantification of Four Efflux Drug Transporters in Liver and Kidney across Species Using Targeted Quantitative Proteomics by Isotope Dilution NanoLC-MS/MS. Pharm. Res. 2016, 33, 2280–2288. [Google Scholar] [CrossRef]
- Qosa, H.; Miller, D.S.; Pasinelli, P.; Trotti, D. Regulation of ABC efflux transporters at blood-brain barrier in health and neurological disorders. Brain Res. 2015, 1628 Pt B, 298–316. [Google Scholar] [CrossRef]
- Al Feteisi, H.; Al-Majdoub, Z.M.; Achour, B.; Couto, N.; Rostami-Hodjegan, A.; Barber, J. Identification and quantification of blood-brain barrier transporters in isolated rat brain microvessels. J. Neurochem. 2018, 146, 670–685. [Google Scholar] [CrossRef] [PubMed]
- Verscheijden, L.F.M.; Koenderink, J.B.; de Wildt, S.N.; Russel, F.G.M. Differences in P-glycoprotein activity in human and rodent blood-brain barrier assessed by mechanistic modelling. Arch. Toxicol. 2021, 95, 3015–3029. [Google Scholar] [CrossRef] [PubMed]
- Mazur, C.S.; Marchitti, S.A.; Dimova, M.; Kenneke, J.F.; Lumen, A.; Fisher, J. Human and rat ABC transporter efflux of bisphenol a and bisphenol a glucuronide: Interspecies comparison and implications for pharmacokinetic assessment. Toxicol. Sci. 2012, 128, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Baltes, S.; Gastens, A.M.; Fedrowitz, M.; Potschka, H.; Kaever, V.; Löscher, W. Differences in the transport of the antiepileptic drugs phenytoin, levetiracetam and carbamazepine by human and mouse P-glycoprotein. Neuropharmacology 2007, 52, 333–346. [Google Scholar] [CrossRef] [PubMed]
- Van Norman, G.A. Limitations of Animal Studies for Predicting Toxicity in Clinical Trials: Is it Time to Rethink Our Current Approach? JACC Basic Transl. Sci. 2019, 4, 845–854. [Google Scholar] [CrossRef]
- Van Norman, G.A. Limitations of Animal Studies for Predicting Toxicity in Clinical Trials: Part 2: Potential Alternatives to the Use of Animals in Preclinical Trials. JACC Basic Transl. Sci. 2020, 5, 387–397. [Google Scholar] [CrossRef]
- Atkins, J.T.; George, G.C.; Hess, K.; Marcelo-Lewis, K.L.; Yuan, Y.; Borthakur, G.; Khozin, S.; LoRusso, P.; Hong, D.S. Pre-clinical animal models are poor predictors of human toxicities in phase 1 oncology clinical trials. Br. J. Cancer 2020, 123, 1496–1501. [Google Scholar] [CrossRef]
- Rider, C.F.; Newton, R.; Bear, C.; Carlsten, C.; Hirota, J.A. The ATP Binding Cassette C4 (ABCC4) Transporter Regulates Extracellular cAMP Transport, Intracellular PKA Activity, and CFTR Channel Activity in Human Airway Epithelial Cells. Am. J. Respir. Crit. Care Med. 2016, 193, A5557. [Google Scholar]
- Mares, L.; Vilchis, F.; Chávez, B.; Ramos, L. Expression and regulation of ABCG2/BCRP1 by sex steroids in the Harderian gland of the Syrian hamster (Mesocricetus auratus). Comptes Rendus Biol. 2019, 342, 279–289. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
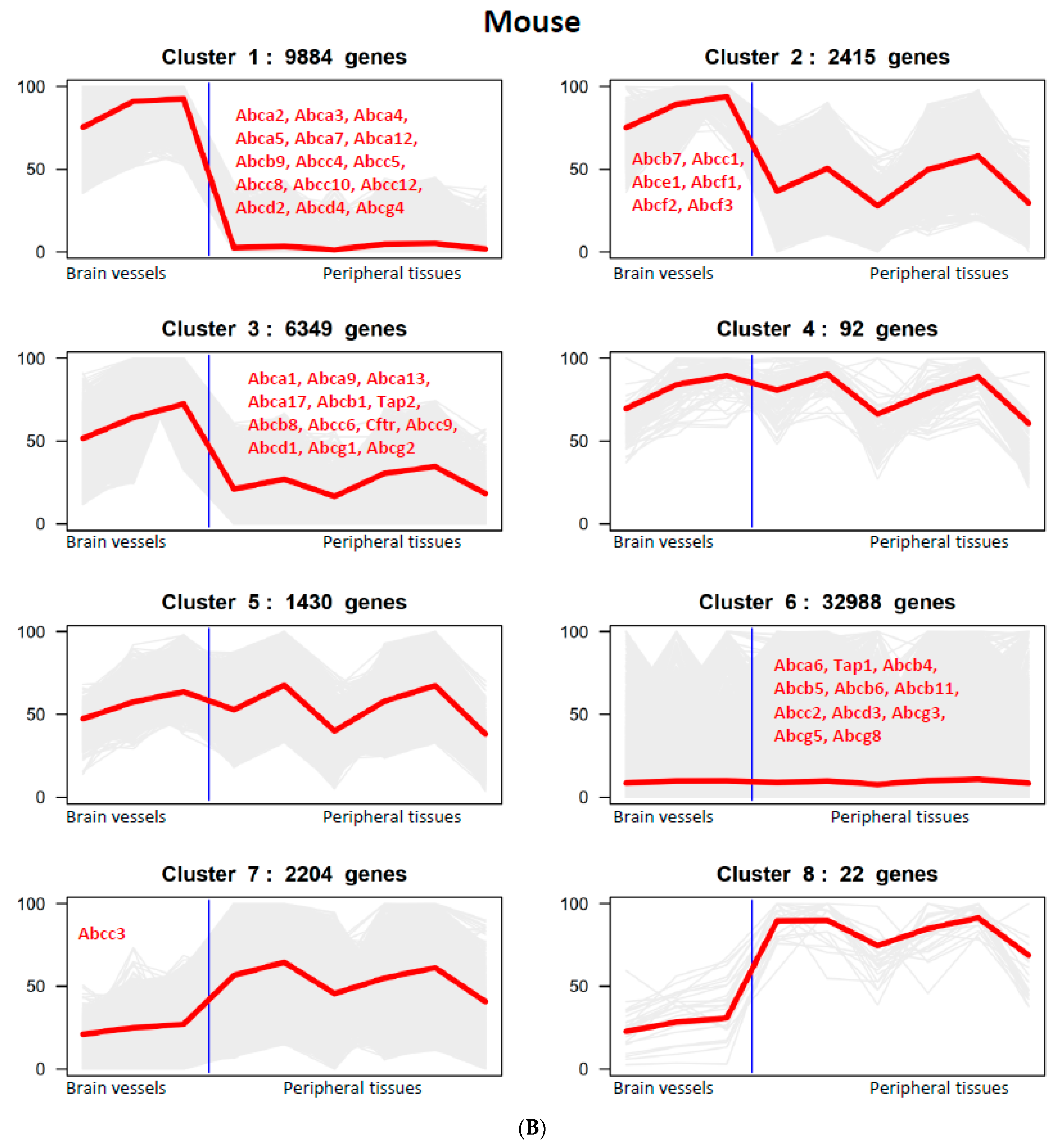{kind=link}
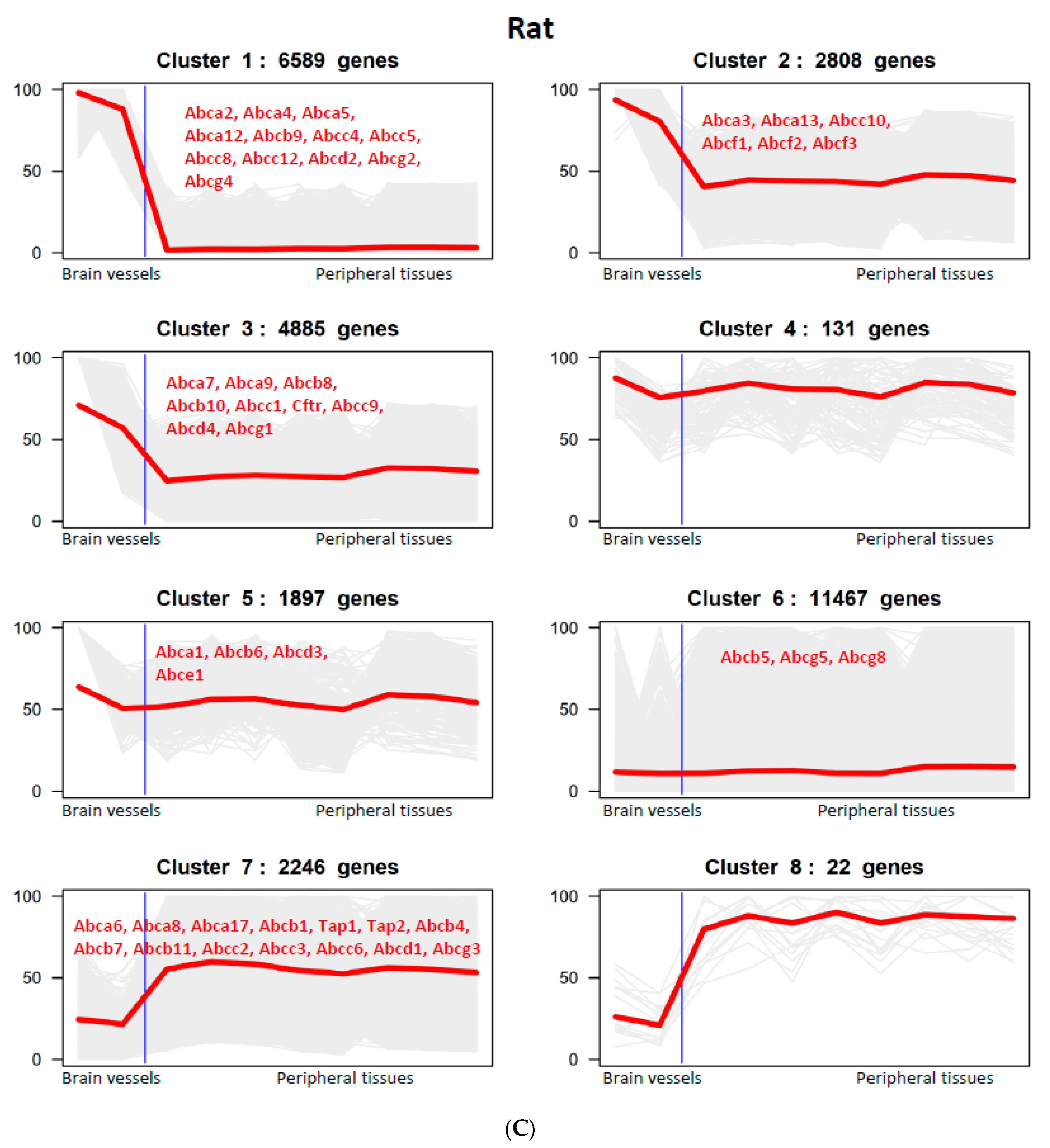{kind=link}
| ABC Transporter Genes | Human (Mean ± SD) | Mouse (Mean ± SD) | Rat (Mean ± SD) |
|---|---|---|---|
| ABCA1 | 0.69 ± 0.66 | 19.45 ± 1.72 h**; r*** | 5.90 ± 1.47 h*** |
| ABCA2 | 208.09 ± 70.27 r* | 107.57 ± 15.42 | 71.63 ± 25.12 |
| ABCA3 | 10.85 ± 6.45 | 35.98 ± 2.22 h** | 26.49 ± 7.73 h* |
| ABCA4 | 0.19 ± 0.15 | 8.22 ± 1.19 h* | 7.85 ± 4.64 h* |
| ABCA5 | 4.51 ± 1.34 | 9.77 ± 1.16 r***; h** | 2.62 ± 0.72 |
| ABCA6 | 0.88 ± 0.08 m*; r**** | 0.67 ± 0.10 r*** | 0.03 ± 0.01 |
| ABCA7 | 13.04 ± 3.68 | 19.29 ± 1.78 | 9.86 ± 4.96 |
| ABCA8 | 4.84 ± 1.54 | 13.82 ± 3.03 h* | 13.52 ± 2.97 h* |
| ABCA9 | 0.81 ± 0.91 | 7.12 ± 1.23 r***; h*** | 0.78 ± 0.26 |
| ABCA10 | 1.93 ± 1.26 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| ABCA12 | 0.01 ± 0.00 | 0.03 ± 0.01 | 0.05 ± 0.02 |
| ABCA13 | 0.02 ± 0.01 | 0.03 ± 0.01 | 0.05 ± 0.02 |
| Abca14 | Not present | 0.00 ± 0.01 | 0.00 ± 0.00 |
| Abca15 | Not present | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Abca16 | Not present | 0.00 ± 0.00 | 0.04 ± 0.04 |
| Abca17 | Not present | 0.20 ± 0.13 | 0.04 ± 0.02 |
| Abcb1a | Homologous to ABCB1 | 290.67 ± 26.70 r## | 93.50 ± 47.78 |
| Abcb1b | Homologous to ABCB1 | 4.67 ± 0.45 r### | 0.28 ± 0.12 |
| ABCB1 (Abcb1a + b1b) | 36.62 ± 54.00 | 295.34 ± 26.58 h***; r** | 93.78 ± 47.90 |
| ABCB2/TAP1 | 19.84 ± 19.30 | 14.98 ± 1.72 | 18.94 ± 5.33 |
| ABCB3/TAP2 | 6.99 ± 8.53 | 47.64 ± 4.68 r*; h* | 12.64 ± 20.52 |
| ABCB4 | 0.84 ± 0.33 | 1.71 ± 0.04 | 3.21 ± 1.00 h** |
| ABCB5 | 0.08 ± 0.03 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| ABCB6 | 19.99 ± 7.05 | 10.93 ± 1.61 | 17.39 ± 2.98 |
| ABCB7 | 1.72 ± 0.41 | 9.95 ± 1.09 h**; r* | 5.79 ± 2.54 h* |
| ABCB8 | 7.08 ± 3.07 | 33.10 ± 9.46 h** | 40.43 ± 5.68 h** |
| ABCB9 | 5.26 ± 2.20 | 21.47 ± 4.52 h** | 17.09 ± 4.28 h* |
| ABCB10 | 1.88 ± 1.37 | 16.11 ± 1.24 h** | 14.00 ± 2.80 h** |
| ABCB11 | 0.31 ± 0.25 | 0.14 ± 0.21 | 0.03 ± 0.01 |
| ABCC1 | 0.90 ± 0.80 | 7.35 ± 0.19 h** | 6.52 ± 4.65 h* |
| ABCC2 | 0.18 ± 0.11 m* | 0.03 ± 0.01 | 0.35 ± 0.05 m***; h** |
| ABCC3 | 0.41 ± 0.37 | 1.21 ± 0.59 h* | 0.98 ± 0.34 |
| ABCC4 | 1.00 ± 1.02 | 77.02 ± 25.83 h**; r* | 26.29 ± 6.88 |
| ABCC5 | 10.00 ± 5.01 | 146.28 ± 22.30 h****; r**** | 17.69 ± 7.92 |
| ABCC6 | 1.04 ± 0.74 | 15.72 ± 4.27 h**; r* | 4.69 ± 3.00 |
| ABCC7 | 0.13 ± 0.07 | 0.14 ± 0.02 | 0.04 ± 0.04 |
| ABCC8 | 43.31 ± 37.99 | 14.67 ± 2.43 | 4.95 ± 1.34 |
| ABCC9 | 2.40 ± 1.95 | 26.48 ± 10.26 h**; r* | 8.14 ± 2.74 |
| ABCC10 | 10.30 ± 4.29 r* | 5.89 ± 0.27 | 3.48 ± 1.13 |
| ABCC11 | 0.23 ± 0.10 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| ABCC12 | 0.24 ± 0.20 | 1.99 ± 0.39 h***; r*** | 0.11 ± 0.02 |
| ABCC13 | 0.01 ± 0.01 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| ABCD1 | 2.19 ± 1.04 | 12.84 ± 1.51 h***; r** | 9.39 ± 2.51 |
| ABCD2 | 0.53 ± 0.11 | 11.41 ± 1.92 h** | 11.61 ± 3.44 h** |
| ABCD3 | 4.49 ± 1.70 | 39.19 ± 4.07 h**** | 49.20 ± 3.48 h****; m* |
| ABCD4 | 21.42 ± 11.18 | 18.38 ± 1.36 | 6.57 ± 0.60 |
| ABCE1 | 4.30 ± 1.77 | 33.30 ± 0.94 h****; r* | 22.97 ± 4.96 h*** |
| ABCF1 | 8.61 ± 3.62 | 60.72 ± 1.79 h**** | 63.48 ± 5.19 h**** |
| ABCF2 | 5.54 ± 2.98 | 54.18 ± 6.08 h** | 55.27 ± 21.99 h** |
| ABCF3 | 19.77 ± 9.53 | 63.67 ± 4.50 h***; r* | 42.97 ± 2.88 h** |
| ABCG1 | 3.57 ± 2.96 | 29.25 ± 5.94 h** | 23.23 ± 6.08 h** |
| ABCG2 | 10.01 ± 12.63 | 185.35 ± 38.42 h**; r* | 83.22 ± 46.64 |
| Abcg3 | Not present | 2.24 ± 1.14 | 18.27 ± 12.99 |
| ABCG4 | 1.98 ± 1.49 | 30.69 ± 7.74 h**; r* | 12.07 ± 4.17 |
| ABCG5 | 0.08 ± 0.10 | 0.02 ± 0.02 | 0.42 ± 0.05 h**; m*** |
| ABCG8 | 0.15 ± 0.08 m*; r* | 0.01 ± 0.02 | 0.01 ± 0.02 |
| ABC Transporter Genes | Human Brain Vessels (Mean ± SD) | Human Lung Tissues (Mean ± SD) |
|---|---|---|
| ABCA1 | 0.69 ± 0.66 | 2.40 ± 0.32 # |
| ABCA2 | 208.09 ± 70.27 ## | 12.32 ± 5.17 |
| ABCA3 | 10.85 ± 6.45 | 95.24 ± 49.02 # |
| ABCA4 | 0.19 ± 0.15 | 0.75 ± 0.56 |
| ABCA5 | 4.51 ± 1.34 | 5.50 ± 0.23 |
| ABCA6 | 0.88 ± 0.08 | 6.12 ± 1.13 ## |
| ABCA7 | 13.04 ± 3.68 | 19.18 ± 6.67 |
| ABCA8 | 4.84 ± 1.54 | 6.8 ± 0.46 |
| ABCA9 | 0.81 ± 0.91 | 1.24 ± 0.22 |
| ABCA10 | 1.93 ± 1.26 | 3.17 ± 2.47 |
| ABCA12 | 0.01 ± 0.00 | 0.03 ± 0.04 |
| ABCA13 | 0.02 ± 0.01 | 0.54 ± 0.67 |
| ABCB1 | 36.62 ± 54.00 | 4.35 ± 2.84 |
| ABCB2/TAP1 | 19.84 ± 19.30 | 48.58 ± 16.21 |
| ABCB3/TAP2 | 6.99 ± 8.53 | 20.72 ± 4.66 |
| ABCB4 | 0.84 ± 0.33 | 0.38 ± 0.35 |
| ABCB5 | 0.08 ± 0.03 | 0.08 ± 0.02 |
| ABCB6 | 19.99 ± 7.05 | 7.10 ± 5.68 |
| ABCB7 | 1.72 ± 0.41 | 6.94 ± 1.81 ## |
| ABCB8 | 7.08 ± 3.07 | 11.87 ± 1.11 |
| ABCB9 | 5.26 ± 2.20 # | 1.70 ± 0.30 |
| ABCB10 | 1.88 ± 1.37 | 4.42 ± 2.15 |
| ABCB11 | 0.31 ± 0.25 | 0.10 ± 0.04 |
| ABCC1 | 0.90 ± 0.80 | 7.28 ± 1.60 ## |
| ABCC2 | 0.18 ± 0.11 | 0.32 ± 0.20 |
| ABCC3 | 0.41 ± 0.37 | 21.07 ± 3.14 ### |
| ABCC4 | 1.00 ± 1.02 | 4.44 ± 2.86 |
| ABCC5 | 10.00 ± 5.01 | 5.91 ± 2.24 |
| ABCC6 | 1.04 ± 0.74 | 13.82 ± 1.98 ### |
| ABCC7 | 0.13 ± 0.07 | 8.30 ± 6.06 |
| ABCC8 | 43.31 ± 37.99 | 1.78 ± 1.56 |
| ABCC9 | 2.40 ± 1.95 | 2.74 ± 0.16 |
| ABCC10 | 10.30 ± 4.29 | 9.85 ± 3.57 |
| ABCC11 | 0.23 ± 0.10 | 0.06 ± 0.07 |
| ABCC12 | 0.24 ± 0.20 | 0.03 ± 0.03 |
| ABCC13 | 0.01 ± 0.01 | 0.00 ± 0.00 |
| ABCD1 | 2.19 ± 1.04 | 3.95 ± 0.33 # |
| ABCD2 | 0.53 ± 0.11 ## | 0.14 ± 0.07 |
| ABCD3 | 4.49 ± 1.70 | 10.13 ± 3.67 |
| ABCD4 | 21.42 ± 11.18 | 24.41 ± 4.06 |
| ABCE1 | 4.30 ± 1.77 | 11.34 ± 4.15 |
| ABCF1 | 8.61 ± 3.62 | 21.40 ± 3.42 # |
| ABCF2 | 5.54 ± 2.98 | 6.56 ± 0.97 |
| ABCF3 | 19.77 ± 9.53 | 28.26 ± 2.61 |
| ABCG1 | 3.57 ± 2.96 | 31.01 ± 16.02 # |
| ABCG2 | 10.01 ± 12.63 | 0.03 ± 0.02 |
| ABCG4 | 1.98 ± 1.49 | 0.04 ± 0.03 |
| ABCG5 | 0.08 ± 0.10 | 0.02 ± 0.01 |
| ABCG8 | 0.15 ± 0.08 | 0.00 ± 0.00 |
| ABC Transporter Genes | Brain Vessels (Mean ± SD) | Liver Tissue (Mean ± SD) | Lung Vessels (Mean ± SD) | Spleen Tissues (Mean ± SD) |
|---|---|---|---|---|
| Abca1 | 19.45 ± 1.72 LT, LV, ST** | 4.29 ± 4.03 | 7.18 ± 3.59 | 5.02 ± 2.74 |
| Abca2 | 107.57 ± 15.42 LT, LV, ST*** | 2.02 ± 1.40 | 15.62 ± 5.47 | 4.81 ± 2.73 |
| Abca3 | 35.98 ± 2.22 LT, ST*** | 3.04 ± 3.04 | 37.80 ± 8.16 LT, ST*** | 7.90 ± 4.63 |
| Abca4 | 8.22 ± 1.19 LT, LV, ST**** | 0.19 ± 0.27 | 2.54 ± 0.63 LT, ST* | 0.14 ± 0.09 |
| Abca5 | 9.77 ± 1.16 LT, ST**; LV* | 0.32 ± 0.21 | 5.08 ± 2.65 LT, ST* | 0.29 ± 0.11 |
| Abca6 | 0.67 ± 0.10 | 18.76 ± 9.39 BV, LV, ST** | 0.76 ± 0.24 | 0.16 ± 0.14 |
| Abca7 | 19.29 ± 1.78 LT, ST** | 0.27 ± 0.12 | 24.35 ± 7.14 LT, ST*** | 4.22 ± 1.23 |
| Abca8 | 13.82 ± 3.03 | 14.47 ± 5.09 | 82.39 ± 16.70 BV, LT, ST**** | 3.44 ± 0.96 |
| Abca9 | 7.12 ± 1.23 LT****; LV, ST*** | 0.24 ± 0.19 | 1.47 ± 0.46 | 1.70 ± 1.16 |
| Abca10 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Abca12 | 0.03 ± 0.01 | 0.00 ± 0.00 | 0.02 ± 0.02 | 0.00 ± 0.00 |
| Abca13 | 0.03 ± 0.01 | 0.00 ± 0.00 | 0.01 ± 0.02 | 0.01 ± 0.01 |
| Abca14 | 0.00 ± 0.01 | 0.06 ± 0.04 | 2.02 ± 1.30 LT, ST* | 0.03 ± 0.03 |
| Abca15 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Abca16 | 0.00 ± 0.00 | 0.00 ± 0. 00 | 0.01 ± 0.02 | 0.00 ± 0.00 |
| Abca17 | 0.20 ± 0.13 | 0.02 ± 0.04 | 0.81 ± 0.44 | 0.10 ± 0.08 |
| Abcb1a | 290.67 ± 26.70 LT, LV, ST**** | 0.27 ± 0.07 | 26.81 ± 7.34 | 3.37 ± 0.99 |
| Abcb1b | 4.67 ± 0.45 LT*** | 0.80 ± 0.13 | 3.39 ± 1.12 LT* | 2.85 ± 0.81 LT* |
| Abca1a + b1b | 295.34 ± 26.58 LT, LV, ST**** | 1.06 ± 0.09 | 30.19 ± 7.63 | 6.21 ± 0.13 |
| Abcb2/Tap1 | 14.98 ± 1.72 | 4.12 ± 1.59 | 81.16 ± 24.53 BV, LT** | 58.37 ± 19.49 BV, LT* |
| Abcb3/Tap2 | 47.64 ± 4.68 LT** | 4.00 ± 2.28 | 123.39 ± 20.66 LT****; BV, ST*** | 58.10 ± 8.97 LT** |
| Abcb4 | 1.71 ± 0.04 | 26.94 ± 18.47 BV, LV* | 0.45 ± 0.13 | 4.10 ± 0.87 |
| Abcb5 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Abcb6 | 10.93 ± 1.61 | 34.06 ± 9.07 BV, LV, ST**** | 20.95 ± 2.10 BV***; ST**** | 6.76 ± 0.87 |
| Abcb7 | 9.95 ± 1.09 LT, LV, ST** | 3.46 ± 2.07 | 3.94 ± 0.81 | 3.99 ± 0.61 |
| Abcb8 | 33.10 ± 9.46 ST* | 12.75 ± 2.54 | 28.88 ± 13.28 ST* | 6.89 ± 1.83 |
| Abcb9 | 21.47 ± 4.52 LT, ST****; LV*** | 0.42 ± 0.20 | 5.32 ± 0.57 | 3.24 ± 1.17 |
| Abcb10 | 16.11 ± 1.24 | 12.51 ± 4.56 | 15.02 ± 1.10 | 9.96 ± 1.91 |
| Abcb11 | 0.14 ± 0.21 | 89.50 ± 23.92 BV, LV, ST**** | 0.28 ± 0.48 | 0.36 ± 0.53 |
| Abcc1 | 7.35 ± 0.19 LT*** | 0.19 ± 0.10 | 9.54 ± 1.78 LT***; ST** | 4.00 ± 2.05 LT* |
| Abcc2 | 0.03 ± 0.01 | 15.76 ± 13.17 | 0.43 ± 0.18 | 0.06 ± 0.09 |
| Abcc3 | 1.21 ± 0.59 | 16.62 ± 8.84 BV* | 29.72 ± 4.05 BV***; LT*; ST** | 11.97 ± 0.21 |
| Abcc4 | 77.02 ± 25.83 LT, LV, ST*** | 0.36 ± 0.27 | 2.60 ± 0.97 | 2.76 ± 1.38 |
| Abcc5 | 146.28 ± 22.30 LT, ST****; LV* | 1.10 ± 0.54 | 96.94 ± 25.04 LT, ST*** | 5.96 ± 1.90 |
| Abcc6 | 15.72 ± 4.27 LV, ST*** | 10.60 ± 3.70 LV, ST** | 0.19 ± 0.17 | 0.05 ± 0.07 |
| Abcc7 | 0.14 ± 0.02 | 0.02 ± 0.00 | 0.25 ± 0.13 LT, ST* | 0.02 ± 0.02 |
| Abcc8 | 14.67 ± 2.43 LT, LV, ST**** | 0.05 ± 0.09 | 1.02 ± 0.28 | 0.05 ± 0.07 |
| Abcc9 | 26.48 ± 10.26 LT, LV** | 1.03 ± 0.72 | 1.02 ± 0.40 | 2.07 ± 1.19 |
| Abcc10 | 5.89 ± 0.27 LT, ST** | 0.36 ± 0.17 | 4.64 ± 2.13 LT, ST** | 0.65 ± 0.19 |
| Abcc11 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Abcc12 | 1.99 ± 0.39 LT, LV, ST**** | 0.06 ± 0.04 | 0.20 ± 0.17 | 0.01 ± 0.01 |
| Abcc13 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Abcd1 | 12.84 ± 1.51 | 3.18 ± 2.39 | 26.75 ± 2.48 BV, ST*; LT*** | 17.06 ± 6.80 |
| Abcd2 | 11.41 ± 1.92 LT, ST***; LV** | 2.70 ± 2.12 | 4.23 ± 0.74 | 1.01 ± 0.12 |
| Abcd3 | 39.19 ± 4.07 LV*; ST**** | 100.43 ± 65.85 ST* | 26.40 ± 4.68 ST*** | 4.73 ± 1.73 |
| Abcd4 | 18.38 ± 1.36 LT, ST** | 2.05 ± 0.63 | 19.81 ± 6.34 LT, ST*** | 1.86 ± 0.28 |
| Abce1 | 33.30 ± 0.94 LT, ST** | 13.44 ± 6.11 | 26.15 ± 5.40 LT* | 16.58 ± 3.04 |
| Abcf1 | 60.72 ± 1.79 LT****; ST** | 8.65 ± 4.83 | 65.19 ± 7.33 LT****; ST*** | 34.83 ± 4.98 LT** |
| Abcf2 | 54.18 ± 6.08 LT****; LV**; ST*** | 9.84 ± 4.21 | 30.66 ± 3.54 LT** | 22.49 ± 3.93 LT* |
| Abcf3 | 63.67 ± 4.50 LT, ST**** | 16.19 ± 5.45 | 59.31 ± 9.07 LT****; ST*** | 19.70 ± 0.78 |
| Abcg1 | 29.25 ± 5.94 LT** | 0.80 ± 0.45 | 27.65 ± 2.70 LT** | 19.93 ± 9.74 LT* |
| Abcg2 | 185.35 ± 38.42 LT, LV, ST**** | 28.53 ± 15.63 | 24.84 ± 5.87 | 9.38 ± 1.22 |
| Abcg3 | 2.24 ± 1.14 | 0.84 ± 0.21 | 9.07 ± 1.59 LT* | 25.01 ± 5.93 BV, LT****; LV** |
| Abcg4 | 30.69 ± 7.74 LT, LV, ST**** | 0.33 ± 0.35 | 0.45 ± 0.23 | 1.64 ± 0.34 |
| Abcg5 | 0.02 ± 0.02 | 13.96 ± 8.20 LT, LV, ST* | 0.15 ± 0.19 | 0.09 ± 0.10 |
| Abcg8 | 0.01 ± 0.02 | 10.87± 8.46 | 0.01 ± 0.02 | 0.12 ± 0.14 |
| ABC Transporter Genes | Brain Vessels (Mean ± SD) | Liver Tissues (Mean ± SD) | Lung Vessels (Mean ± SD) | Spleen Tissues (Mean ± SD) |
|---|---|---|---|---|
| Abca1 | 5.90 ± 1.47 | 11.81 ± 2.86 | 13.32 ± 6.25 ST* | 3.92 ± 0.90 |
| Abca2 | 71.63 ± 25.12 LT***; LV, ST** | 4.19 ± 1.92 | 16.56 ± 5.44 | 5.97 ± 0.30 |
| Abca3 | 26.49 ± 7.73 LT, ST* | 9.58 ± 2.57 | 113.33 ± 6.40 BV, LT, ST**** | 9.57 ± 0.29 |
| Abca4 | 7.85 ± 4.64 LT, LV, ST* | 0.20 ± 0.04 | 0.36 ± 0.08 | 0.97 ± 0.21 |
| Abca5 | 2.60 ± 0.72 LT* | 0.54 ± 0.05 | 7.89 ± 2.98 LT, ST** | 0.11 ± 0.03 |
| Abca6 | 0.03 ± 0.01 | 21.79 ± 4.20 BV, LV, ST**** | 0.45 ± 0.07 | 0.03 ± 0.01 |
| Abca7 | 9.86 ± 4.96 | 0.30 ± 0.11 | 19.32 ± 7.94 LT**; ST* | 5.72 ± 0.17 |
| Abca8 | 13.52 ± 2.10 | 21.57 ± 2.51 ST** | 21.59 ± 9.15 ST** | 3.28 ± 0.50 |
| Abca9 | 0.78 ± 0.26 | 0.48 ± 0.25 | 0.24 ± 0.18 | 0.50 ± 0.07 |
| Abca10 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Abca12 | 0.05 ± 0.02 | 0.00 ± 0.00 | 0.00 ± 0.01 | 0.00 ± 0.00 |
| Abca13 | 0.05 ± 0.02 | 0.00 ± 0.00 | 0.00 ± 0.01 | 0.05 ± 0.01 |
| Abca14 | 0.00 ± 0.00 | 0.01 ± 0.02 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Abca15 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00± 0.00 | 0.00 ± 0.00 |
| Abca16 | 0.04 ± 0.04 | 0.69 ± 0.33 BV, LV, ST** | 0.03 ± 0.04 | 0.04 ± 0.01 |
| Abca17 | 0.04 ± 0.02 | 0.07 ± 0.04 | 2.12 ± 1.04 BV, LT, ST** | 0.06 ± 0.01 |
| Abcb1a | 93.50 ± 47.78 LT, LV, ST** | 4.18 ± 1.07 | 2.43 ± 0.90 | 0.21 ± 0.08 |
| Abcb1b | 0.28 ± 0.12 | 1.97 ± 1.39 | 9.37 ± 7.72 | 0.42 ± 0.05 |
| Abca1a + b1b | 93.78 ± 47.9078 LT, ST**; LV* | 6.15 ± 2.46 | 11.80 ± 8.56 | 0.63 ± 0.13 |
| Abcb2/Tap1 | 18.94 ± 5.33 | 11.65 ± 3.24 | 140.59 ± 55.46 BV, LV**; ST* | 53.59 ± 8.14 |
| Abcb3/Tap2 | 12.64 ± 20.52 | 11.07 ± 4.69 | 61.88 ± 101.10 | 50.28 ± 3.97 |
| Abcb4 | 3.21 ± 1.00 | 28.83 ± 16.53 BV, LV* | 3.62 ± 3.79 | 15.81 ± 2.31 |
| Abcb5 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Abcb6 | 17.39 ± 2.98 | 20.74 ± 3.91 | 20.01 ± 3.67 | 14.43 ± 1.16 |
| Abcb7 | 5.79 ± 2.54 | 13.74 ± 1.09 BV**; LV***; ST* | 3.01 ± 0.45 | 9.19 ± 1.38 LV** |
| Abcb8 | 40.43 ± 5.68 LT, ST***; LV* | 9.37 ± 4.18 | 25.33 ± 5.35 LT**; ST* | 9.66 ± 0.57 |
| Abcb9 | 17.09 ± 4.28 LT, LV, ST*** | 1.54 ± 0.77 | 2.47 ± 0.41 | 2.77 ± 0.54 |
| Abcb10 | 14.00 ± 2.80 | 5.84 ± 2.34 | 9.59 ± 2.38 | 58.53 ± 6.4328 BV, LT, LV*** |
| Abcb11 | 0.03 ± 0.01 | 103.10 ± 6.5828 BV, LV, ST**** | 0.33 ± 0.50 | 0.59 ± 0.13 |
| Abcc1 | 6.52 ± 4.65 | 0.35 ± 0.20 | 3.50 ± 1.47 | 5.72 ± 0.75 |
| Abcc2 | 0.35 ± 0.05 | 72.54 ± 23.86 BV, LV, ST*** | 0.18 ± 0.02 | 0.06 ± 0.03 |
| Abcc3 | 0.98 ± 0.34 | 2.16 ± 0.56 | 7.17 ± 1.76 BV, LT, ST*** | 1.31 ± 0.02 |
| Abcc4 | 26.29 ± 6.88 LT, ST***; LV** | 3.05 ± 1.41 | 7.92 ± 3.01 | 3.64 ± 0.70 |
| Abcc5 | 17.69 ± 7.92 LT**; ST* | 0.40 ± 0.07 | 33.37 ± 24.38 | 5.19 ± 0.12 |
| Abcc6 | 4.69 ± 3.00 | 21.87 ± 1.95 BV, LV, ST**** | 3.36 ± 2.22 | 0.08 ± 0.01 |
| Abcc7 | 0.04 ± 0.04 | 0.06 ± 0.06 | 1.20 ± 0.46 BV, LV, ST** | 0.01 ± 0.01 |
| Abcc8 | 4.95 ± 1.34 LT**; LV*** | 0.51 ± 0.12 | 0.28 ± 0.04 | 0.00 ± 0.00 |
| Abcc9 | 8.14 ± 2.74 ST* | 4.05 ± 1.50 | 4.53 ± 3.51 | 1.47 ± 0.16 |
| Abcc10 | 3.48 ± 1.13 | 0.84 ± 0.47 | 5.80 ± 1.60 LT** | 3.37 ± 0.16 |
| Abcc11 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Abcc12 | 0.11 ± 0.02 LT, LV, ST*** | 0.01 ± 0.01 | 0.01 ± 0.02 | 0.01 ± 0.01 |
| Abcc13 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Abcd1 | 9.39 ± 2.51 | 18.47 ± 8.20 | 17.72 ± 6.65 | 11.54 ± 0.22 |
| Abcd2 | 11.61 ± 3.44 LT, ST** | 2.64 ± 0.20 | 20.21 ±25.38 | 0.58 ± 0.06 |
| Abcd3 | 49.20 ± 3.48 ST** | 93.03 ± 14.82 BV**; LV***; ST**** | 44.92 ± 8.65 ST** | 6.19 ± 0.64 |
| Abcd4 | 6.57 ± 0.60 | 1.28 ± 0.57 | 12.17 ± 4.39 LT, ST** | 2.18 ± 0.46 |
| Abce1 | 22.97 ± 4.96 | 23.71 ± 0.63 | 19.48 ± 3.47 | 22.87 ± 3.26 |
| Abcf1 | 63.48 ± 5.19 LT**; ST* | 25.22 ± 8.45 | 60.05 ± 13.24 LT** | 40.27 ± 4.01 |
| Abcf2 | 55.27 ± 21.99 LT* | 19.43 ± 3.85 | 39.85 ± 12.28 | 23.77 ± 1.47 |
| Abcf3 | 43.97 ± 2.88 LT*; ST** | 23.21 ± 6.41 | 39.58 ± 9.53 LT*; ST* | 19.44 ± 1.53 |
| Abcg1 | 23.23 ± 6.08 LT* | 3.68 ± 1.21 | 299.66 ± 357.98 | 36.56 ± 3.12 BV*; LT*** |
| Abcg2 | 83.22 ± 46.64 LT, LV, ST* | 13.83 ± 2.39 | 1.17 ± 0.45 | 6.75 ± 0.97 |
| Abcg3 | 18.27 ± 12.99 | 62.98 ± 15.33 BV** | 105.58 ± 126.24 | 35.68 ± 2.39 |
| Abcg4 | 12.07 ± 4.17 LT**; LV* | 0.17 ± 0.04 | 3.34 ± 3.29 | 5.28 ± 0.97 |
| Abcg5 | 0.42 ± 0.05 | 7.38 ± 3.68 BV, LV, ST** | 0.04 ± 0.01 | 0.03 ± 0.01 |
| Abcg8 | 0.01 ± 0.02 | 2.36 ± 2.06 | 0.00 ± 0.01 | 0.02 ± 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Liu, Q.Y.; Haqqani, A.S.; Liu, Z.; Sodja, C.; Leclerc, S.; Baumann, E.; Delaney, C.E.; Brunette, E.; Stanimirovic, D.B. Differential Expression of ABC Transporter Genes in Brain Vessels vs. Peripheral Tissues and Vessels from Human, Mouse and Rat. Pharmaceutics 2023, 15, 1563. https://doi.org/10.3390/pharmaceutics15051563
Zhang W, Liu QY, Haqqani AS, Liu Z, Sodja C, Leclerc S, Baumann E, Delaney CE, Brunette E, Stanimirovic DB. Differential Expression of ABC Transporter Genes in Brain Vessels vs. Peripheral Tissues and Vessels from Human, Mouse and Rat. Pharmaceutics. 2023; 15(5):1563. https://doi.org/10.3390/pharmaceutics15051563
Chicago/Turabian StyleZhang, Wandong, Qing Yan Liu, Arsalan S. Haqqani, Ziying Liu, Caroline Sodja, Sonia Leclerc, Ewa Baumann, Christie E. Delaney, Eric Brunette, and Danica B. Stanimirovic. 2023. "Differential Expression of ABC Transporter Genes in Brain Vessels vs. Peripheral Tissues and Vessels from Human, Mouse and Rat" Pharmaceutics 15, no. 5: 1563. https://doi.org/10.3390/pharmaceutics15051563