
Therapeutic and Diagnostic Potential of Exosomes as Drug Delivery Systems in Brain Cancer
 , , and
, , and
Abstract
:
1. Introduction
2. Physicochemical Characteristics of Exosomes
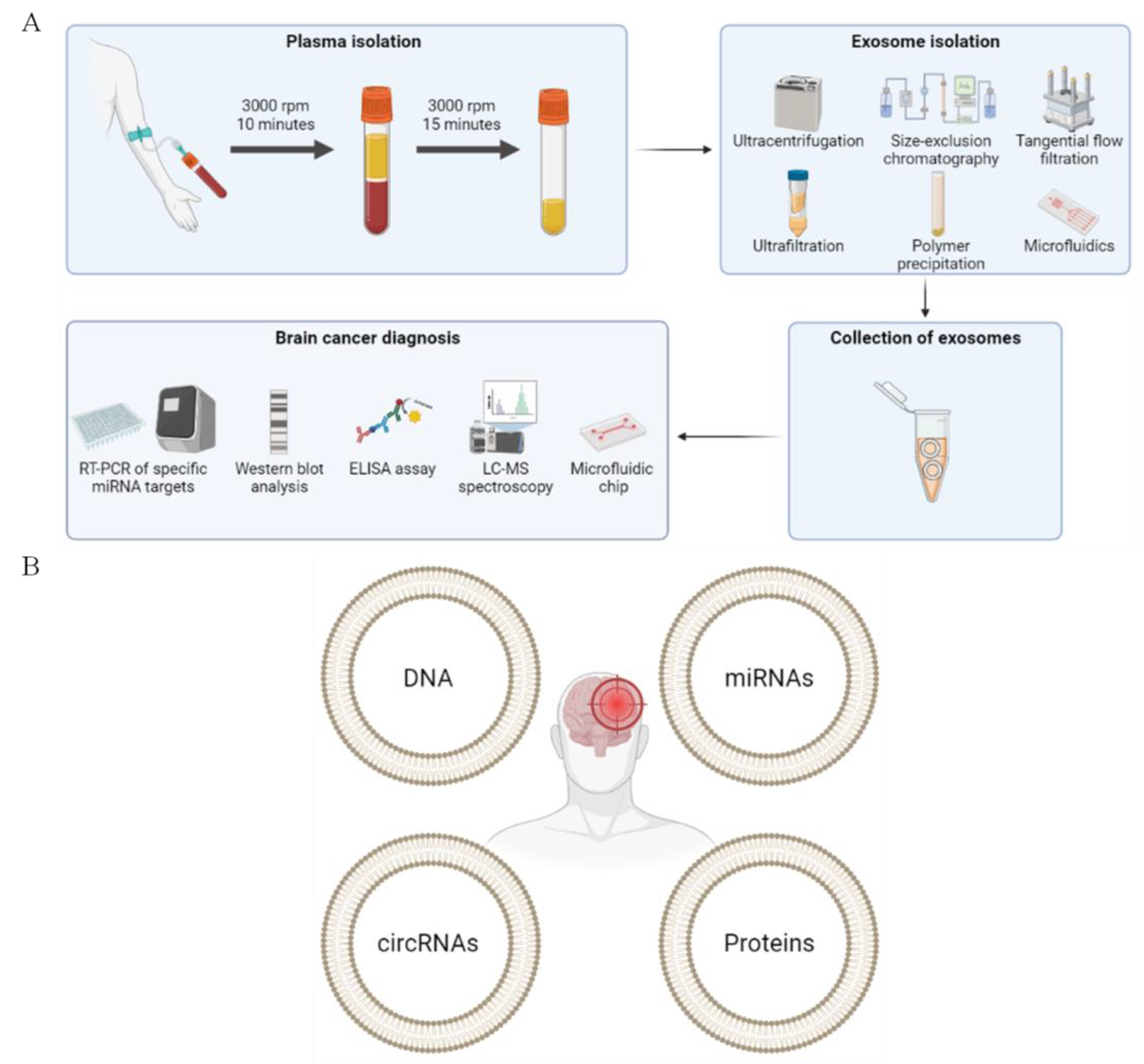
2.1. Exosome Isolation
2.2. Exosome Size
- Exosomes (30–150 nm);
- Microvesicles (100–1000 nm);
- Large oncosomes (1000–10,000 nm); and
2.3. Exosome Stability
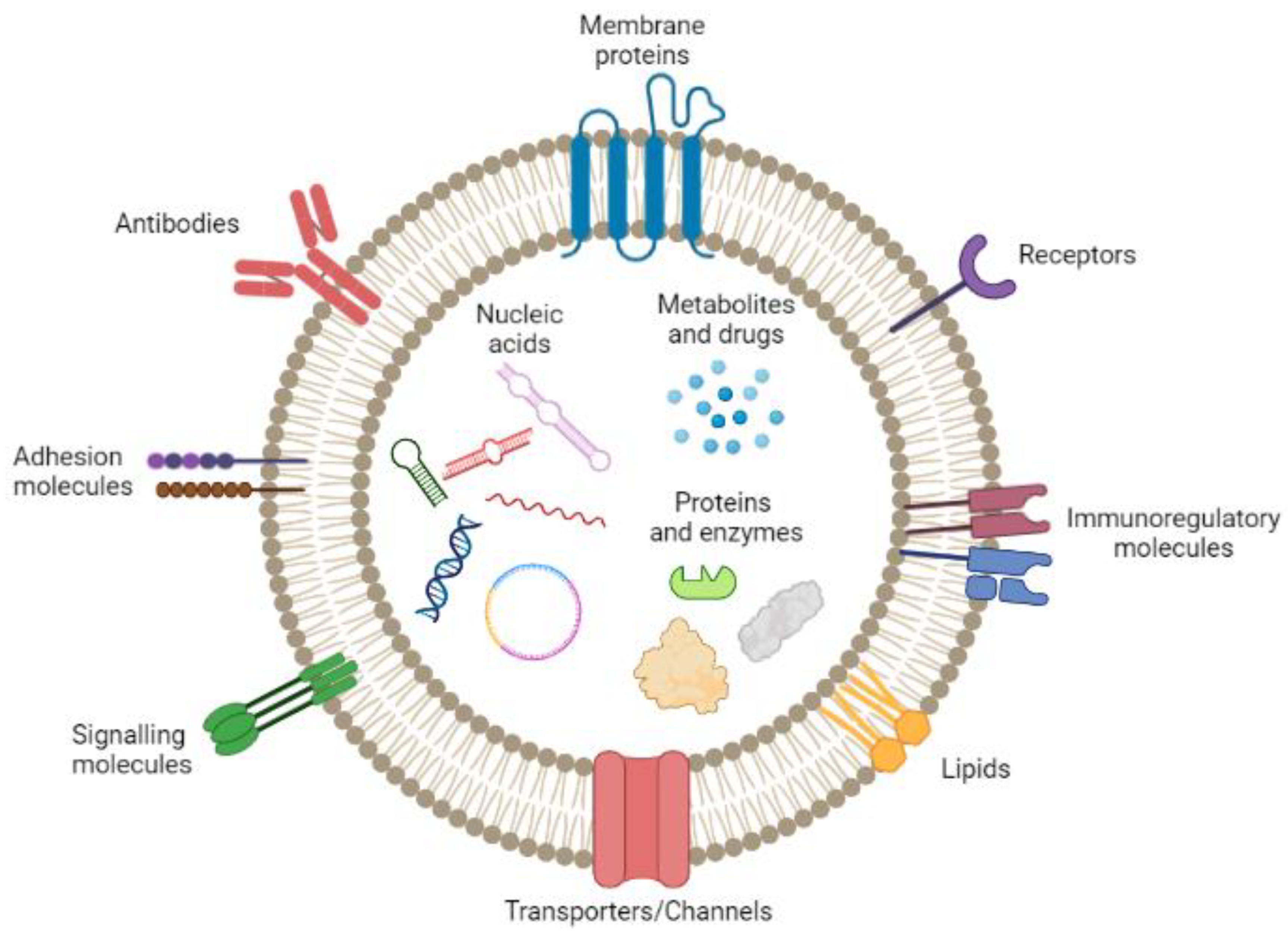
2.4. Exosome Biological Properties
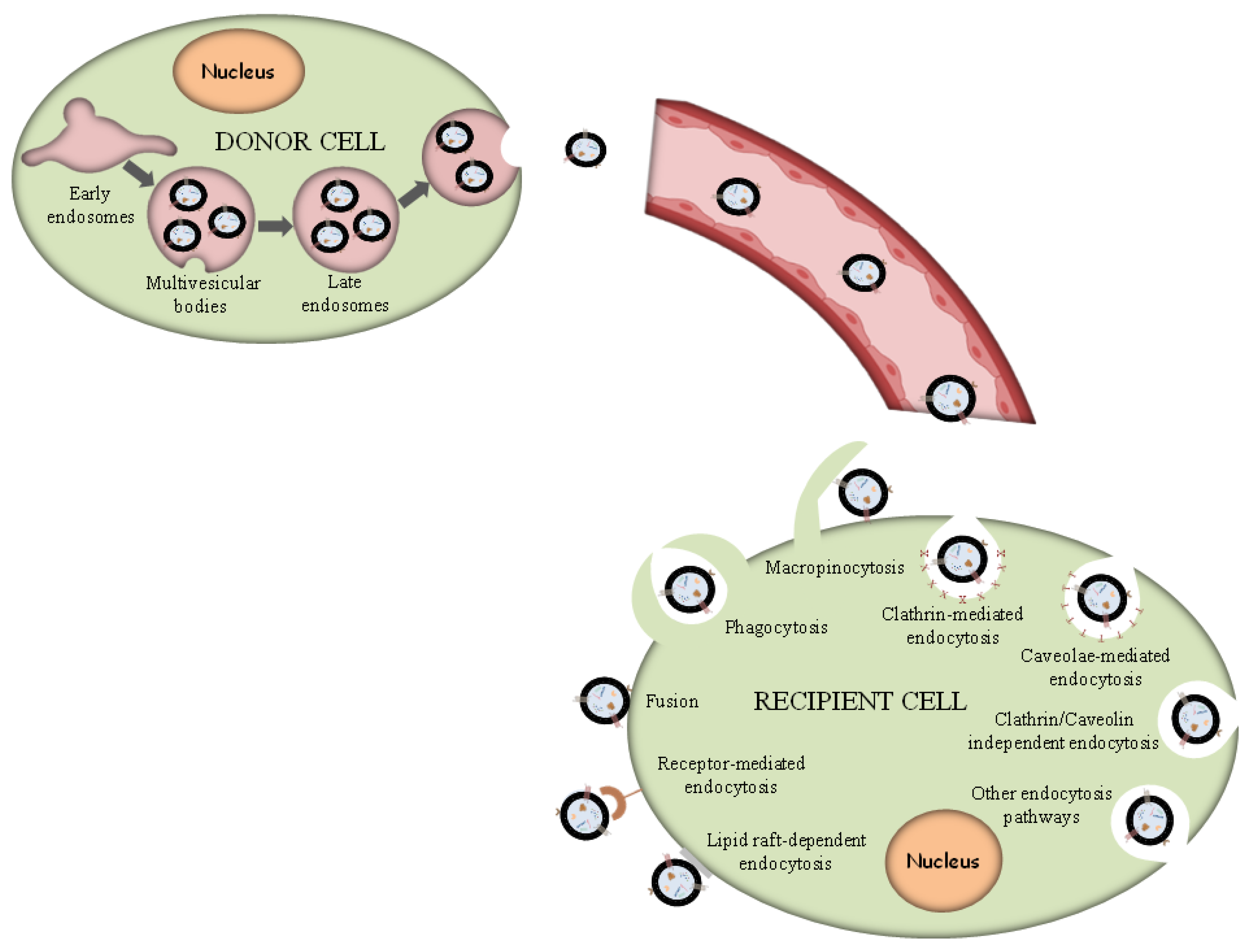
2.5. Exosome Biogenesis and Internalization
- (i)
- Phagocytosis. Phagocytosis is almost a similar internalization process to macropinocytosis where exosomes, along with some extracellular fluids, are phagocytosed. This process is common and adopted by both phagocytic cells, i.e., macrophages and dendritic cells (DCs) and non-phagocytic cells, i.e., T cells [91].
- (ii)
- Macropinocytosis. Macropinocytosis is a process in which the deformation of the plasma membrane leads to the formation of protrusions that surround the extracellular fluid and exosomes, therefrom uptaking exosomes. This complicated process is mediated by many factors, since it relies on Ras-related C3 botulinum toxun substrate 1 (Rac1), actin and cholesterol and it requires Na+/H+ exchange [92].
- (iii)
- Clathrin/caveolin-mediated endocytosis. The Clathrin protein shapes a basket knitted-like structure around the exosomes for their internalization. The plasma membrane of the target cells creates an internal folding, which is followed by squeezing off the clathrin-coated vesicle from the membrane. Subsequently, the exosome releases all its cargo in the targets cell’s endosomes to implement particular functions [93].
- (iv)
- Internalization through lipid raft. The endocytosis process may be also mediated by caveolin-1, similar to the clathrin-dependent way, whose clusters in plasma membrane are configured in rafts. The invagination of the plasma membrane named caveolae is abundant in caveolin 1, glycolipids, and cholesterol [94], and
- (v)
- Direct fusion with the plasma membrane. In this fusion process, the Lysosomal-Associated Membrane Protein 1 (LAMP-1) and glycoprotein type I transmembrane protein are implicated primarily residing across lysosomal membranes, and in certain cases, it can be expressed across the plasma membrane of the cell, and the integrins or tetraspanins [95,96].
2.6. Cancer Cell-Derived Exosomes
3. Exosomes in the Diagnosis of Brain Cancer
3.1. Exosomal miRNA Biomarkers in Brain Cancer
3.2. Exosomal circRNA Biomarkers in Brain Cancer
3.3. Exosomal Gene Biomarkers in Brain Cancer
3.4. Exosomal Protein Biomarkers in Brain Cancer
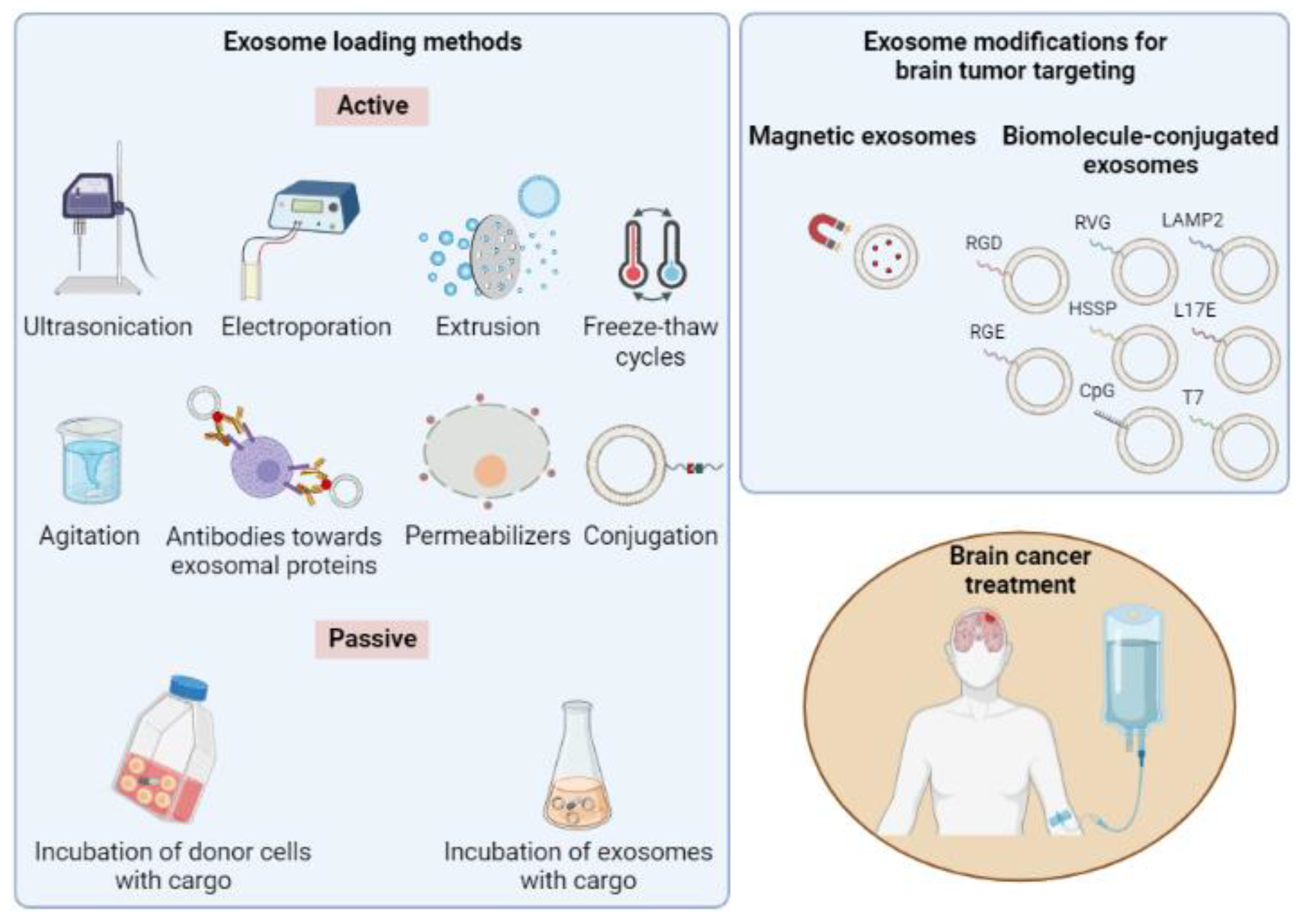
4. Exosomes in the Treatment of Brain Cancer
4.1. Mesenchymal Stem Cells-Derived Exosomes
4.2. Embryonic Stem Cells-Derived Exosomes
4.3. Neural Stem Cells-Derived Exosomes
4.4. Macrophage-Derived Exosomes
4.5. Glioma Cells-Derived Exosomes
4.6. Blood Exosomes
4.7. Natural Brain Endothelia Cell-Derived Exosomes
4.8. 293T Cell-Derived Exosomes
4.9. Milk Exosomes
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 5-FC | 5-Fluorocytosine |
| 5-FU | 5-Fluorouracil |
| 14-3-3E | 14-3-3 protein epsilon |
| AGAP2 | ArfGAP with GTPase Domain, Ankyrin Repeat And PH Domain 2 |
| AgNCs | Silver nanoparticles |
| Agos | Argonautes |
| AKT | Protein kinase B |
| ANXA1 | Annexin A1 |
| ANXA2 | Annexin A2 |
| APC | Adenomatous polyposis coli |
| APNG | Alkylpurine-DNA-N-glycosylase |
| ARF6 | ADP Ribosylation Factor 6 |
| ASOs | Antisense Oligonucleotides |
| ATG12 | Autophagy Related 12 |
| B7-H1 | B7-Homolog 1 |
| BAF-TiN | Biotinylated Antibody-Functionalized Titanium Nitride |
| BBB | Blood-brain-barrier |
| BECs | Brain Endothelial Cells |
| BG | Benzylguanine |
| BM | Brain metastases |
| BMSC | Bone mesenchymal stem cells |
| BMSCExo | Bone marrow mesenchymal stem cells-derived exosomes |
| BNCT | Boron Neutron Caption Therapy |
| BQR | Brequinar |
| BPA | Boron Phenylalanine |
| BTG2 | B-cell translocation gene 2 |
| CA9 | Carbonic Anhydrase 9 |
| CCND1 | Cyclin D1 |
| CD | Cluster of Differentiation |
| CD117 | KIT proto-oncogene receptor tyrosine kinase |
| CDA | Cytosine Deaminase |
| CDK6 | Cyclin-dependent kinase 6 |
| circRNA | circular RNA |
| COF1 | Cofillin |
| COP I | Coat protein I |
| CpG | 5′—C—phosphate—G—3′ |
| cPLA2 | cytosolic Phospholipase A2 |
| CSF | Cerebrospinal fluid |
| CT | Computed Tomography |
| Cur | Curcumin |
| CYP2J2 | Cytochrome P450 2J2 isoform |
| DCs | Dendritic Cells |
| DHODH | Dihydroorotate Dehydrogenase |
| DHT | Dihydrotanshinone |
| DIPG | Diffuse Intrinsic Pontine Glioma |
| DNA | Deoxyribonucleic acid |
| DNM3 | Dynamin 3 |
| DOX | Doxorubicin |
| dsDNA | Double-stranded DNA |
| ECM | Extracellular Matrix |
| EGFR | Epidermic Growth Factor Receptor |
| EGFRvIII | Epidermal Growth Factor Receptor variant III |
| ENOA | Alpha-enolase |
| ESCRT | Endosomal Sorting Complex Required for Transport |
| ESCs | Embryonic Stem Cells |
| EVs | Extracellular vehicles |
| EZH2 | Enhancer of Zeste 2 Polycomb Repressive Complex 2 Subunit |
| FasL | Fas Ligand |
| FBXO11 | F-Box Protein 11 |
| FGFR | Fibroblast Growth Factor Receptor |
| FLOT2 | Flotillin 2 |
| FOXA2 | Forkhead box protein A2 |
| FUS | Focused Ultrasound |
| G3P | Glyceraldehyde-3-phosphate dehydrogenase |
| GATA2 | GATA-binding factor 2 |
| GBM | Glioblastoma |
| GL | Glycyrrhizic acid |
| GLI1 | GLI family zinc finger 1 |
| GMP | Good manufacturing practice |
| GPX4 | Glutatheione peroxidase 4 |
| GSCs | Glioblastoma Stem Cells |
| hEnMSCs | Human Endometrial Mesenchymal Stem Cells |
| HCQ | Hydroxychloroquine |
| HGA | High-grade astrocytoma |
| HGG | High-Grade Glioma |
| HIF-1a | Hypoxia-inducible factor-1a |
| HIFs | Hypoxia-inducible factors |
| HLA-G | Human leukocyte antigen-G |
| HMOX1 | Heme oxygenase 1 |
| hMSCs | Human Mesenchymal Stromal Cells |
| HSP90B | Heat-shock protein 90B |
| HSPs | Heat-shock proteins |
| HSSP | Heme oxygenase-1 specific short peptide |
| hUC-MSCs | Human Umbilical Cord Mesenchymal Stem Cells |
| HUVEC | Human Umbilical Vein Endothelial Cells |
| ICAM-1 | Intercellular Adhesion Molecule-1 |
| ICG | Indocyanine green |
| ICOSL | Inducible costimulator ligand |
| ICP-MS | Inductively coupled plasma mass spectometry |
| IDH1 | Isocitrate dehydrogenase-1 |
| IFN-g | Interferon-gamma |
| IL-10 | Interleukin-10 |
| IL-12 | Interleukin-12 |
| IL-3 | Interleukin-3 |
| iNKT | Invariant Natural Killer T |
| ISEV | International Society for Extracellular Vesicles |
| JAG1 | Jagged Canonical Notch Ligand 1 |
| KRAS | Kirsten rat sarcoma virus |
| LAMP-1 | Lysosomal-Associated Membrane Protein-1 |
| LAMP-2b | Lysosomal-Associated Membrane Protein-2b |
| LGG | Low-grade glioma |
| LLGL2 | LLGL Scribble Cell Polarity Complex Component 2 |
| lncRNA | Long non-coding RNA |
| MAPK | Mitogen-activated protein kinase |
| MaSC | Marrow stromal cells |
| MGMT | O(6)-methylguanine DNA methyltransferase |
| MHC-I | Major histocompatibility complex-I |
| MHC-II | Major histocompatibility complex-II |
| miRNA | MicroRNA |
| MRI | Magnetic Resonance Imaging |
| mRNA | Messenger RNA |
| MSCs | Mesenchymal stem cells |
| mTOR | Mammalian target of Rapamycin |
| ΜVs | Microvesicles |
| MVBs | Multivesicular bodies |
| MVP | Major Vault Protein |
| NF-κB | Nuclear Factor κΒ |
| NHEJ | Non-homologous end joining |
| NIR | Near-infrared radiation |
| NPM | Nucleophosmin |
| NPs | Nanoparticles |
| NSCs | Neural Stem Cells |
| PCNSL | Primary Central Nervous Lymphoma |
| PCR | Polymerase chain reaction |
| PD-1 | Programmed Death-1 |
| PDA | Polydopamine |
| PDCD4 | Programmed cell death protein 4 |
| PD-L1 | Programmed Death-Ligand 1 |
| PI3K | Phosphoinositide 3-kinase |
| PKM | Pyruvate Kinase M1/2 |
| PPM1D | Protein Phosphatase 1D |
| PRDX1 | Peroxiredoxin-1 |
| PRKDC | Protein Kinase, DNA-Activated, Catalytic Subunit |
| PTEN | Phosphatase and tensin homolog |
| PTENP1 | Phosphatase and tensin homolog pseudogene 1 |
| PTX | Paclitaxel |
| Rab | Ras-associated binding |
| RAB13 | Ras-related protein 13 |
| Rac1 | Ras-related C3 botulinum toxin substrate 1 |
| RAP1B | Ras-associated protein 1B |
| Rb | Retinoblastoma protein |
| RECK | Reversion Inducing Cysteine Rich Protein With Kazal Motifs |
| R-EXO | Reassembly-Exosomes |
| RNA | Ribonucleic acid |
| RNU6-1 | RNA, U6 Small Nuclear 1 |
| ROS | Reactive Oxygen Species |
| rRNA | Ribosomal RNA |
| RVG-15 | Rabies viral glycoprotein |
| SCI | Spinal Cord Injury |
| SEC | Size-exclusion chromatography |
| siRNA | Small-interfering RNA |
| SLC31A1 | Solute Carrier Family 31 (Copper Transporter), Member 1 |
| sncRNA | Small non-coding RNA |
| SPIONs | Superparamagnetic Iron Oxide Nanoparticles |
| ST14 | ST14 Transmembrane Serine Protease Matriptase |
| STAT3 | Signal transducer and activator of transcription 3 |
| TACC | Transforming Acidic Coiled-Coil Containing Protein 3 |
| TanIIA | Tanshinone IIA |
| TdEX | Tumor-derived Exosomes |
| TERA | Transitional endoplasmic reticulum ATPase |
| TFF | Tangential Flow Filtration |
| TIMP3 | Tissue inhibitor of metalloproteinase 3 |
| TLR-9 | Toll-like receptor 9 |
| TMZ | Temozolomide |
| TPIS | Triose phosphate isomerase |
| TPM1 | Tropomyosin 1 |
| TPM4 | Tropomyosin 4 |
| tRNA | Transfer RNA |
| TSG101 | Tumor susceptibility gene 101 |
| UC | Ultracentifiguration |
| UTR | Untranslated region |
| VB | Verbascoside |
| VCP | Vasolin-containing protein |
| VEGF | Vascular Endothelial Growth Factor |
| VIME | Vimentin |
| WHO | World Health Organization |
| yCD::UPRT | Yeast cytosine deaminase::uracilphosphoribosyl transferase |
| ZEB1 | Zinc Finger E-Box Binding Homeobox 1 |
| ZnS | Zinc sulfide |
References
- Arrighetti, N.; Corbo, C.; Evangelopoulos, M.; Pastò, A.; Zuco, V.; Tasciotti, E. Exosome-like Nanovectors for Drug Delivery in Cancer. Curr. Med. Chem. 2018, 26, 6132–6148. [Google Scholar] [CrossRef] [PubMed]
- Romano, E.; Netti, P.A.; Torino, E. Exosomes in Gliomas: Biogenesis, Isolation, and Preliminary Applications in Nanomedicine. Pharmaceuticals 2020, 13, 319. [Google Scholar] [CrossRef]
- Benecke, L.; Coray, M.; Umbricht, S.; Chiang, D.; Figueiró, F.; Muller, L. Exosomes: Small Evs with Large Immunomodulatory Effect in Glioblastoma. Int. J. Mol. Sci. 2021, 22, 3600. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wang, X.; Wang, J.; Hao, Y.; Liu, F.; Wang, X.; Yang, L.; Lu, Z. The Roles of Exosomes as Future Therapeutic Agents and Diagnostic Tools for Glioma. Front. Oncol. 2021, 11, 733529. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Feng, Y.; Gao, S.; Mu, Q.; Liu, C. Nanotherapeutics Overcoming the Blood-Brain Barrier for Glioblastoma Treatment. Front. Pharmacol. 2021, 12, 786700. [Google Scholar] [CrossRef] [PubMed]
- Ghaemmaghami, A.B.; Mahjoubin-Tehran, M.; Movahedpour, A.; Morshedi, K.; Sheida, A.; Taghavi, S.P.; Mirzaei, H.; Hamblin, M.R. Role of Exosomes in Malignant Glioma: MicroRNAs and Proteins in Pathogenesis and Diagnosis. Cell Commun. Signal. 2020, 18, 120. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Keramat, S.; Izadirad, M.; Chen, Z.S.; Soukhtanloo, M. The Potential Role of Exosomes in the Treatment of Brain Tumors, Recent Updates and Advances. Front. Oncol. 2022, 12, 869929. [Google Scholar] [CrossRef]
- Oliveira, F.D.; Castanho, M.A.R.B.; Neves, V. Exosomes and Brain Metastases: A Review on Their Role and Potential Applications. Int. J. Mol. Sci. 2021, 22, 10899. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Li, M.; Weng, B.; Mao, H.; Zhao, J. Exosome-Based Delivery Nanoplatforms: Next-Generation Theranostic Platforms for Breast Cancer. Biomater. Sci. 2022, 10, 1607–1625. [Google Scholar] [CrossRef]
- Halevas, E.G.; Avgoulas, D.I.; Katsipis, G.; Pantazaki, A.A. Flavonoid-Liposomes Formulations: Physico-Chemical Characteristics, Biological Activities and Therapeutic Applications. Eur. J. Med. Chem. Rep. 2022, 5, 100059. [Google Scholar] [CrossRef]
- Batrakova, E.V.; Kim, M.S. Using Exosomes, Naturally-Equipped Nanocarriers, for Drug Delivery. J. Control. Release 2015, 219, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Bi, J.; Huang, J.; Tang, Y.; Du, S.; Li, P. Exosome: A Review of Its Classification, Isolation Techniques, Storage, Diagnostic and Targeted Therapy Applications. Int. J. Nanomed. 2020, 15, 6917–6934. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Chen, Q.; Lin, L.; Sha, C.; Li, T.; Liu, Y.; Yin, X.; Xu, Y.; Chen, L.; Gao, W.; et al. Regulation of Exosome Production and Cargo Sorting. Int. J. Biol. Sci. 2021, 17, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Nam, G.H.; Choi, Y.; Kim, G.B.; Kim, S.; Kim, S.A.; Kim, I.S. Emerging Prospects of Exosomes for Cancer Treatment: From Conventional Therapy to Immunotherapy. Adv. Mater. 2020, 32, 2002440. [Google Scholar] [CrossRef]
- Gurung, S.; Perocheau, D.; Touramanidou, L.; Baruteau, J. The Exosome Journey: From Biogenesis to Uptake and Intracellular Signalling. Cell Commun. Signal. 2021, 19, 47. [Google Scholar] [CrossRef]
- Jan, A.T.; Rahman, S.; Badierah, R.; Lee, E.J.; Mattar, E.H.; Redwan, E.M.; Choi, I. Expedition into Exosome Biology: A Perspective of Progress from Discovery to Therapeutic Development. Cancers 2021, 13, 1157. [Google Scholar] [CrossRef] [PubMed]
- Lyu, L.; Hu, M.; Fu, A.; Xing, B. Extracellular Vesicle Directed Exogenous Ion Channel Transport for Precise Manipulation of Biological Events. Bioconjug. Chem. 2018, 29, 2715–2722. [Google Scholar] [CrossRef]
- Burtenshaw, D.; Regan, B.; Owen, K.; Collins, D.; McEneaney, D.; Megson, I.L.; Redmond, E.M.; Cahill, P.A. Exosomal Composition, Biogenesis and Profiling Using Point-of-Care Diagnostics—Implications for Cardiovascular Disease. Front. Cell Dev. Biol. 2022, 10, 853451. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Ji, W.; Zhao, R.; Yang, J.; Lu, Z.; Li, Y.; Zhang, X. Exosome: A Significant Nano-Scale Drug Delivery Carrier. J. Mater. Chem. B 2020, 8, 7591–7608. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Xu, K.; Zheng, X.; Chen, T.; Wang, J.; Song, Y.; Shao, Y.; Zheng, S. Application of Exosomes as Liquid Biopsy in Clinical Diagnosis. Signal Transduct. Target. Ther. 2020, 5, 144. [Google Scholar] [CrossRef]
- Yu, D.; Li, Y.; Wang, M.; Gu, J.; Xu, W.; Cai, H.; Fang, X.; Zhang, X. Exosomes as a New Frontier of Cancer Liquid Biopsy. Mol. Cancer 2022, 21, 56. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Pang, B.; Li, J.; Gao, N.; Fan, T.; Li, Y. Emerging Role of Exosomes in Liquid Biopsy for Monitoring Prostate Cancer Invasion and Metastasis. Front. Cell Dev. Biol. 2021, 9, 679527. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Li, M.; Wang, T.; Liang, Y.; Zhong, Z.; Wang, X.; Zhou, Q.; Chen, L.; Lang, Q.; He, Z.; et al. Lactation-Related MicroRNA Expression Profiles of Porcine Breast Milk Exosomes. PLoS ONE 2012, 7, e43691. [Google Scholar] [CrossRef] [PubMed]
- Rehman, F.U.; Liu, Y.; Zheng, M.; Shi, B. Exosomes Based Strategies for Brain Drug Delivery. Biomaterials 2023, 293, 121949. [Google Scholar] [CrossRef]
- Karami Fath, M.; Azami, J.; Masoudi, A.; Mosaddeghi Heris, R.; Rahmani, E.; Alavi, F.; Alagheband Bahrami, A.; Payandeh, Z.; Khalesi, B.; Dadkhah, M.; et al. Exosome-Based Strategies for Diagnosis and Therapy of Glioma Cancer. Cancer Cell Int. 2022, 22, 262. [Google Scholar] [CrossRef] [PubMed]
- Katakowski, M.; Chopp, M. Exosomes as Tools to Suppress Primary Brain Tumor. Cell. Mol. Neurobiol. 2016, 36, 343–352. [Google Scholar] [CrossRef]
- Zottel, A.; Videtič Paska, A.; Jovčevska, I. Nanotechnology Meets Oncology: Nanomaterials in Brain Cancer Research, Diagnosis and Therapy. Materials 2019, 12, 1588. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhang, Y.; Yao, B.; Sun, P.; Hao, Y.; Piao, H.; Zhao, X. Role of Exosomes in the Progression, Diagnosis, and Treatment of Gliomas. Med. Sci. Monit. 2020, 26, e924023. [Google Scholar] [CrossRef] [PubMed]
- Davatgaran-Taghipour, Y.; Masoomzadeh, S.; Farzaei, M.H.; Bahramsoltani, R.; Karimi-Soureh, Z.; Rahimi, R.; Abdollahi, M. Polyphenol Nanoformulations for Cancer Therapy: Experimental Evidence and Clinical Perspective. Int. J. Nanomed. 2017, 12, 2689–2702. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.G.; Venkatesan, J.; Shim, M.S. Selective Anticancer Therapy Using Pro-Oxidant Drug-Loaded Chitosan–Fucoidan Nanoparticles. Int. J. Mol. Sci. 2019, 20, 3220. [Google Scholar] [CrossRef]
- Abdelaziz, H.M.; Gaber, M.; Abd-Elwakil, M.M.; Mabrouk, M.T.; Elgohary, M.M.; Kamel, N.M.; Kabary, D.M.; Freag, M.S.; Samaha, M.W.; Mortada, S.M.; et al. Inhalable Particulate Drug Delivery Systems for Lung Cancer Therapy: Nanoparticles, Microparticles, Nanocomposites and Nanoaggregates. J. Control. Release 2018, 269, 374–392. [Google Scholar] [CrossRef] [PubMed]
- Curcio, M.; Diaz-Gomez, L.; Cirillo, G.; Nicoletta, F.P.; Leggio, A.; Iemma, F. Dual-Targeted Hyaluronic Acid/Albumin Micelle-Like Nanoparticles for the Vectorization of Doxorubicin. Pharmaceutics 2021, 13, 304. [Google Scholar] [CrossRef] [PubMed]
- de la Torre, P.; Pérez-Lorenzo, M.J.; Alcázar-Garrido, Á.; Flores, A.I. Cell-Based Nanoparticles Delivery Systems for Targeted Cancer Therapy: Lessons from Anti-Angiogenesis Treatments. Molecules 2020, 25, 715. [Google Scholar] [CrossRef] [PubMed]
- AlMalki, F.A.; Hassan, A.M.; Klaab, Z.M.; Abdulla, S.; Pizzi, A. Tannin Nanoparticles (NP99) Enhances the Anticancer Effect of Tamoxifen on ER+ Breast Cancer Cells. J. Renew. Mater. 2021, 9, 2077–2092. [Google Scholar] [CrossRef]
- Hua, S.; de Matos, M.B.C.; Metselaar, J.M.; Storm, G. Current Trends and Challenges in the Clinical Translation of Nanoparticulate Nanomedicines: Pathways for Translational Development and Commercialization. Front. Pharmacol. 2018, 9, 790. [Google Scholar] [CrossRef] [PubMed]
- Ion, D.; Niculescu, A.G.; Păduraru, D.N.; Andronic, O.; Mușat, F.; Grumezescu, A.M.; Bolocan, A. An Up-to-Date Review of Natural Nanoparticles for Cancer Management. Pharmaceutics 2022, 14, 18. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular Vesicles: Exosomes, Microvesicles, and Friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef]
- Hood, J.L. Post Isolation Modification of Exosomes for Nanomedicine Applications. Nanomedicine 2016, 11, 1745–1756. [Google Scholar] [CrossRef]
- Hood, J.L.; Wickline, S.A. A Systematic Approach to Exosome-Based Translational Nanomedicine. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2012, 4, 458–467. [Google Scholar] [CrossRef]
- You, B.; Xu, W.; Zhang, B. Engineering Exosomes: A New Direction for Anticancer Treatment. Am. J. Cancer Res. 2018, 8, 1332–1342. [Google Scholar]
- Samanta, S.; Rajasingh, S.; Drosos, N.; Zhou, Z.; Dawn, B.; Rajasingh, J. Exosomes: New Molecular Targets of Diseases. Acta Pharmacol. Sin. 2018, 39, 501–513. [Google Scholar] [CrossRef]
- Jia, S.; Zocco, D.; Samuels, M.L.; Chou, M.F.; Chammas, R.; Skog, J.; Zarovni, N.; Momen-Heravi, F.; Kuo, W.P. Emerging Technologies in Extracellular Vesicle-Based Molecular Diagnostics. Expert Rev. Mol. Diagn. 2014, 14, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Yuana, Y.; Levels, J.; Grootemaat, A.; Sturk, A.; Nieuwland, R. Co-Isolation of Extracellular Vesicles and High-Density Lipoproteins Using Density Gradient Ultracentrifugation. J. Extracell. Vesicles 2014, 3, 23262. [Google Scholar] [CrossRef]
- Cheruvanky, A.; Zhou, H.; Pisitkun, T.; Kopp, J.B.; Knepper, M.A.; Yuen, P.S.T.; Star, R.A. Rapid Isolation of Urinary Exosomal Biomarkers Using a Nanomembrane Ultrafiltration Concentrator. Am. J. Physiol. Physiol. 2007, 292, F1657–F1661. [Google Scholar] [CrossRef]
- Böing, A.N.; van der Pol, E.; Grootemaat, A.E.; Coumans, F.A.W.; Sturk, A.; Nieuwland, R. Single-Step Isolation of Extracellular Vesicles by Size-Exclusion Chromatography. J. Extracell. Vesicles 2014, 3, 23430. [Google Scholar] [CrossRef] [PubMed]
- Deregibus, M.C.; Figliolini, F.; D’antico, S.; Manzini, P.M.; Pasquino, C.; De Lena, M.; Tetta, C.; Brizzi, M.F.; Camussi, G. Charge-Based Precipitation of Extracellular Vesicles. Int. J. Mol. Med. 2016, 38, 1359–1366. [Google Scholar] [CrossRef]
- Niu, Z.; Pang, R.T.K.; Liu, W.; Li, Q.; Cheng, R.; Yeung, W.S.B. Polymer-Based Precipitation Preserves Biological Activities of Extracellular Vesicles from an Endometrial Cell Line. PLoS ONE 2017, 12, e0186534. [Google Scholar] [CrossRef]
- Lobb, R.J.; Becker, M.; Wen Wen, S.; Wong, C.S.F.; Wiegmans, A.P.; Leimgruber, A.; Möller, A. Optimized Exosome Isolation Protocol for Cell Culture Supernatant and Human Plasma. J. Extracell. Vesicles 2015, 4, 27031. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Skog, J.; Hsu, C.-H.; Lessard, R.T.; Balaj, L.; Wurdinger, T.; Carter, B.S.; Breakefield, X.O.; Toner, M.; Irimia, D. Microfluidic Isolation and Transcriptome Analysis of Serum Microvesicles. Lab Chip 2010, 10, 505–511. [Google Scholar] [CrossRef]
- Li, P.; Kaslan, M.; Lee, S.H.; Yao, J.; Gao, Z. Progress in Exosome Isolation Techniques. Theranostics 2017, 7, 789–804. [Google Scholar] [CrossRef]
- Lee, K.; Shao, H.; Weissleder, R.; Lee, H. Acoustic Purification of Extracellular Microvesicles. ACS Nano 2015, 9, 2321–2327. [Google Scholar] [CrossRef]
- Busatto, S.; Vilanilam, G.; Ticer, T.; Lin, W.-L.; Dickson, D.; Shapiro, S.; Bergese, P.; Wolfram, J. Tangential Flow Filtration for Highly Efficient Concentration of Extracellular Vesicles from Large Volumes of Fluid. Cells 2018, 7, 273. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, C.; Vizio, D.D.; Sahoo, S.; Théry, C.; Witwer, K.W.; Wauben, M.; Hill, A.F. Techniques Used for the Isolation and Characterization of Extracellular Vesicles: Results of a Worldwide Survey. J. Extracell. Vesicles 2016, 5, 32945. [Google Scholar] [CrossRef]
- Tauro, B.J.; Greening, D.W.; Mathias, R.A.; Ji, H.; Mathivanan, S.; Scott, A.M.; Simpson, R.J. Comparison of Ultracentrifugation, Density Gradient Separation, and Immunoaffinity Capture Methods for Isolating Human Colon Cancer Cell Line LIM1863-Derived Exosomes. Methods 2012, 56, 293–304. [Google Scholar] [CrossRef]
- Morishita, M.; Takahashi, Y.; Nishikawa, M.; Sano, K.; Kato, K.; Yamashita, T.; Imai, T.; Saji, H.; Takakura, Y. Quantitative Analysis of Tissue Distribution of the B16BL6-Derived Exosomes Using a Streptavidin-Lactadherin Fusion Protein and Iodine-125-Labeled Biotin Derivative After Intravenous Injection in Mice. J. Pharm. Sci. 2015, 104, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Smyth, T.; Kullberg, M.; Malik, N.; Smith-Jones, P.; Graner, M.W.; Anchordoquy, T.J. Biodistribution and Delivery Efficiency of Unmodified Tumor-Derived Exosomes. J. Control. Release 2015, 199, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Haney, M.J.; Klyachko, N.L.; Zhao, Y.; Gupta, R.; Plotnikova, E.G.; He, Z.; Patel, T.; Piroyan, A.; Sokolsky, M.; Kabanov, A.V.; et al. Exosomes as Drug Delivery Vehicles for Parkinson’s Disease Therapy. J. Control. Release 2015, 207, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Didiot, M.-C.; Hall, L.M.; Coles, A.H.; Haraszti, R.A.; Godinho, B.M.; Chase, K.; Sapp, E.; Ly, S.; Alterman, J.F.; Hassler, M.R.; et al. Exosome-Mediated Delivery of Hydrophobically Modified SiRNA for Huntingtin MRNA Silencing. Mol. Ther. 2016, 24, 1836–1847. [Google Scholar] [CrossRef]
- Faruqu, F.N.; Wang, J.T.-W.; Xu, L.; McNickle, L.; Chong, E.M.-Y.; Walters, A.; Gurney, M.; Clayton, A.; Smyth, L.A.; Hider, R.; et al. Membrane Radiolabelling of Exosomes for Comparative Biodistribution Analysis in Immunocompetent and Immunodeficient Mice—A Novel and Universal Approach. Theranostics 2019, 9, 1666–1682. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, J.; Gao, W.; Xie, N. Exosomes as Anticancer Drug Delivery Vehicles: Prospects and Challenges. Front. Biosci. Landmark 2022, 27, 293. [Google Scholar] [CrossRef]
- Rezaie, J.; Feghhi, M.; Etemadi, T. A Review on Exosomes Application in Clinical Trials: Perspective, Questions, and Challenges. Cell Commun. Signal. 2022, 20, 145. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Cocucci, E.; Racchetti, G.; Meldolesi, J. Shedding Microvesicles: Artefacts No More. Trends Cell Biol. 2009, 19, 43–51. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA Delivery by Extracellular Vesicles in Mammalian Cells and Its Applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef]
- Patil, A.A.; Rhee, W.J. Exosomes: Biogenesis, Composition, Functions, and Their Role in Pre-Metastatic Niche Formation. Biotechnol. Bioprocess Eng. 2019, 24, 689–701. [Google Scholar] [CrossRef]
- Teng, F.; Fussenegger, M. Shedding Light on Extracellular Vesicle Biogenesis and Bioengineering. Adv. Sci. 2021, 8, 2003505. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, C.-J.; Hou, W.-H.; Jiang, D.-G.; Tian, H.-J.; Wang, L.-N.; Jia, F.; Zhou, C.-H.; Zhu, J.-J. Circular RNAs in Early Brain Development and Their Influence and Clinical Significance in Neuropsychiatric Disorders. Neural Regen. Res. 2020, 15, 817. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, J.P.K.; Stevens, M.M. Strategic Design of Extracellular Vesicle Drug Delivery Systems. Adv. Drug Deliv. Rev. 2018, 130, 12–16. [Google Scholar] [CrossRef]
- Kibria, G.; Ramos, E.K.; Wan, Y.; Gius, D.R.; Liu, H. Exosomes as a Drug Delivery System in Cancer Therapy: Potential and Challenges. Mol. Pharm. 2018, 15, 3625–3633. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The Biology, Function, and Biomedical Applications of Exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Vahabi, A.; Rezaie, J.; Hassanpour, M.; Panahi, Y.; Nemati, M.; Rasmi, Y.; Nemati, M. Tumor Cells-Derived Exosomal CircRNAs: Novel Cancer Drivers, Molecular Mechanisms, and Clinical Opportunities. Biochem. Pharmacol. 2022, 200, 115038. [Google Scholar] [CrossRef] [PubMed]
- Monopoli, M.P.; Walczyk, D.; Campbell, A.; Elia, G.; Lynch, I.; Baldelli Bombelli, F.; Dawson, K.A. Physical–Chemical Aspects of Protein Corona: Relevance to in Vitro and in Vivo Biological Impacts of Nanoparticles. J. Am. Chem. Soc. 2011, 133, 2525–2534. [Google Scholar] [CrossRef] [PubMed]
- Lunova, M.; Smolková, B.; Lynnyk, A.; Uzhytchak, M.; Jirsa, M.; Kubinová, Š.; Dejneka, A.; Lunov, O. Targeting the MTOR Signaling Pathway Utilizing Nanoparticles: A Critical Overview. Cancers 2019, 11, 82. [Google Scholar] [CrossRef]
- Jeppesen, D.K.; Fenix, A.M.; Franklin, J.L.; Higginbotham, J.N.; Zhang, Q.; Zimmerman, L.J.; Liebler, D.C.; Ping, J.; Liu, Q.; Evans, R.; et al. Reassessment of Exosome Composition. Cell 2019, 177, 428–445.e18. [Google Scholar] [CrossRef] [PubMed]
- Lo Cicero, A.; Stahl, P.D.; Raposo, G. Extracellular Vesicles Shuffling Intercellular Messages: For Good or for Bad. Curr. Opin. Cell Biol. 2015, 35, 69–77. [Google Scholar] [CrossRef]
- Qu, Y.; Franchi, L.; Nunez, G.; Dubyak, G.R. Nonclassical IL-1β Secretion Stimulated by P2X7 Receptors Is Dependent on Inflammasome Activation and Correlated with Exosome Release in Murine Macrophages. J. Immunol. 2007, 179, 1913–1925. [Google Scholar] [CrossRef]
- Phoonsawat, W.; Aoki-Yoshida, A.; Tsuruta, T.; Sonoyama, K. Adiponectin Is Partially Associated with Exosomes in Mouse Serum. Biochem. Biophys. Res. Commun. 2014, 448, 261–266. [Google Scholar] [CrossRef]
- Chen, L.; Chen, R.; Kemper, S.; Charrier, A.; Brigstock, D.R. Suppression of Fibrogenic Signaling in Hepatic Stellate Cells by Twist1-Dependent MicroRNA-214 Expression: Role of Exosomes in Horizontal Transfer of Twist1. Am. J. Physiol. Liver Physiol. 2015, 309, G491–G499. [Google Scholar] [CrossRef] [PubMed]
- Mathivanan, S.; Fahner, C.J.; Reid, G.E.; Simpson, R.J. ExoCarta 2012: Database of Exosomal Proteins, RNA and Lipids. Nucleic Acids Res. 2012, 40, D1241–D1244. [Google Scholar] [CrossRef]
- Simpson, R.J.; Kalra, H.; Mathivanan, S. ExoCarta as a Resource for Exosomal Research. J. Extracell. Vesicles 2012, 1, 18374. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-K.; Kang, B.; Kim, O.Y.; Choi, D.; Lee, J.; Kim, S.R.; Go, G.; Yoon, Y.J.; Kim, J.H.; Jang, S.C.; et al. EVpedia: An Integrated Database of High-Throughput Data for Systemic Analyses of Extracellular Vesicles. J. Extracell. Vesicles 2013, 2, 20384. [Google Scholar] [CrossRef] [PubMed]
- Subra, C.; Laulagnier, K.; Perret, B.; Record, M. Exosome Lipidomics Unravels Lipid Sorting at the Level of Multivesicular Bodies. Biochimie 2007, 89, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Subra, C.; Grand, D.; Laulagnier, K.; Stella, A.; Lambeau, G.; Paillasse, M.; De Medina, P.; Monsarrat, B.; Perret, B.; Silvente-Poirot, S.; et al. Exosomes Account for Vesicle-Mediated Transcellular Transport of Activatable Phospholipases and Prostaglandins. J. Lipid Res. 2010, 51, 2105–2120. [Google Scholar] [CrossRef]
- Théry, C.; Zitvogel, L.; Amigorena, S. Exosomes: Composition, Biogenesis and Function. Nat. Rev. Immunol. 2002, 2, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-Mediated Transfer of MRNAs and MicroRNAs Is a Novel Mechanism of Genetic Exchange between Cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Skog, J.; Würdinger, T.; van Rijn, S.; Meijer, D.H.; Gainche, L.; Curry, W.T.; Carter, B.S.; Krichevsky, A.M.; Breakefield, X.O. Glioblastoma Microvesicles Transport RNA and Proteins That Promote Tumour Growth and Provide Diagnostic Biomarkers. Nat. Cell Biol. 2008, 10, 1470–1476. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Zaro, J.; Shen, Y. Advances in Exosome-Based Drug Delivery and Tumor Targeting: From Tissue Distribution to Intracellular Fate. Int. J. Nanomed. 2020, 15, 9355–9371. [Google Scholar] [CrossRef] [PubMed]
- EL Andaloussi, S.; Mäger, I.; Breakefield, X.O.; Wood, M.J.A. Extracellular Vesicles: Biology and Emerging Therapeutic Opportunities. Nat. Rev. Drug Discov. 2013, 12, 347–357. [Google Scholar] [CrossRef]
- Shaimardanova, A.; Solovyeva, V.; Chulpanova, D.; James, V.; Kitaeva, K.; Rizvanov, A. Extracellular Vesicles in the Diagnosis and Treatment of Central Nervous System Diseases. Neural Regen. Res. 2020, 15, 586. [Google Scholar] [CrossRef]
- Heusermann, W.; Hean, J.; Trojer, D.; Steib, E.; von Bueren, S.; Graff-Meyer, A.; Genoud, C.; Martin, K.; Pizzato, N.; Voshol, J.; et al. Exosomes Surf on Filopodia to Enter Cells at Endocytic Hot Spots, Traffic within Endosomes, and Are Targeted to the ER. J. Cell Biol. 2016, 213, 173–184. [Google Scholar] [CrossRef]
- Wu, Y.; Wu, W.; Wong, W.M.; Ward, E.; Thrasher, A.J.; Goldblatt, D.; Osman, M.; Digard, P.; Canaday, D.H.; Gustafsson, K. Human Γδ T Cells: A Lymphoid Lineage Cell Capable of Professional Phagocytosis. J. Immunol. 2009, 183, 5622–5629. [Google Scholar] [CrossRef] [PubMed]
- Gonda, A.; Kabagwira, J.; Senthil, G.N.; Wall, N.R. Internalization of Exosomes through Receptor-Mediated Endocytosis. Mol. Cancer Res. 2019, 17, 337–347. [Google Scholar] [CrossRef]
- Tian, T.; Zhu, Y.-L.; Zhou, Y.-Y.; Liang, G.-F.; Wang, Y.-Y.; Hu, F.-H.; Xiao, Z.-D. Exosome Uptake through Clathrin-Mediated Endocytosis and Macropinocytosis and Mediating MiR-21 Delivery. J. Biol. Chem. 2014, 289, 22258–22267. [Google Scholar] [CrossRef] [PubMed]
- Nabi, I.R.; Le, P.U. Caveolae/Raft-Dependent Endocytosis. J. Cell Biol. 2003, 161, 673–677. [Google Scholar] [CrossRef] [PubMed]
- Joshi, B.S.; de Beer, M.A.; Giepmans, B.N.G.; Zuhorn, I.S. Endocytosis of Extracellular Vesicles and Release of Their Cargo from Endosomes. ACS Nano 2020, 14, 4444–4455. [Google Scholar] [CrossRef] [PubMed]
- Andreu, Z.; Yáñez-MÃ3, M. Tetraspanins in Extracellular Vesicle Formation and Function. Front. Immunol. 2014, 5, 442. [Google Scholar] [CrossRef]
- D’Asti, E.; Garnier, D.; Lee, T.H.; Montermini, L.; Meehan, B.; Rak, J. Oncogenic Extracellular Vesicles in Brain Tumor Progression. Front. Physiol. 2012, 3, 294. [Google Scholar] [CrossRef]
- Whiteside, T.L. Immune Modulation of T-Cell and NK (Natural Killer) Cell Activities by TEXs (Tumour-Derived Exosomes). Biochem. Soc. Trans. 2013, 41, 245–251. [Google Scholar] [CrossRef]
- Peinado, H.; Alečković, M.; Lavotshkin, S.; Matei, I.; Costa-Silva, B.; Moreno-Bueno, G.; Hergueta-Redondo, M.; Williams, C.; García-Santos, G.; Ghajar, C.M.; et al. Melanoma Exosomes Educate Bone Marrow Progenitor Cells toward a Pro-Metastatic Phenotype through MET. Nat. Med. 2012, 18, 883–891. [Google Scholar] [CrossRef]
- Smyth, T.J.; Redzic, J.S.; Graner, M.W.; Anchordoquy, T.J. Examination of the Specificity of Tumor Cell Derived Exosomes with Tumor Cells in Vitro. Biochim. Biophys. Acta—Biomembr. 2014, 1838, 2954–2965. [Google Scholar] [CrossRef]
- Rajagopal, C.; Harikumar, K.B. The Origin and Functions of Exosomes in Cancer. Front. Oncol. 2018, 8, 66. [Google Scholar] [CrossRef] [PubMed]
- Santangelo, A.; Tamanini, A.; Cabrini, G.; Dechecchi, M.C. Circulating MicroRNAs as Emerging Non-Invasive Biomarkers for Gliomas. Ann. Transl. Med. 2017, 5, 1–8. [Google Scholar] [CrossRef]
- Vaidya, M.; Sugaya, K. DNA Associated with Circulating Exosomes as a Biomarker for Glioma. Genes 2020, 11, 1276. [Google Scholar] [CrossRef]
- Jafari, D.; Shajari, S.; Jafari, R.; Mardi, N.; Gomari, H.; Ganji, F.; Forouzandeh Moghadam, M.; Samadikuchaksaraei, A. Designer Exosomes: A New Platform for Biotechnology Therapeutics. BioDrugs 2020, 34, 567–586. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A Summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef]
- McLendon, R.E.; Halperin, E.C. Is the Long-Term Survival of Patients with Intracranial Glioblastoma Multiforme Overstated? Cancer 2003, 98, 1745–1748. [Google Scholar] [CrossRef]
- Pan, T.; Jia, P.; Chen, N.; Fang, Y.; Liang, Y.; Guo, M.; Ding, X. Delayed Remote Ischemic Preconditioning ConfersRenoprotection against Septic Acute Kidney Injury via Exosomal MiR-21. Theranostics 2019, 9, 405–423. [Google Scholar] [CrossRef] [PubMed]
- Jansen, F.; Li, Q. Exosomes as Diagnostic Biomarkers in Cardiovascular Diseases. In Exosomes in Cardiovascular Diseases. Advances in Experimental Medicine and Biology; Xiao, J., Cretoiu, S., Eds.; Springer: Singapore, 2017; pp. 61–70. [Google Scholar]
- Raj, D.A.A.; Fiume, I.; Capasso, G.; Pocsfalvi, G. A Multiplex Quantitative Proteomics Strategy for Protein Biomarker Studies in Urinary Exosomes. Kidney Int. 2012, 81, 1263–1272. [Google Scholar] [CrossRef]
- Baker, S.G.; Kramer, B.S.; McIntosh, M.; Patterson, B.H.; Shyr, Y.; Skates, S. Evaluating Markers for the Early Detection of Cancer: Overview of Study Designs and Methods. Clin. Trials 2006, 3, 43–56. [Google Scholar] [CrossRef]
- Properzi, F.; Logozzi, M.; Fais, S. Exosomes: The Future of Biomarkers in Medicine. Biomark. Med. 2013, 7, 769–778. [Google Scholar] [CrossRef]
- Sun, W.; Luo, J.; Jiang, H.; Duan, D.D. Tumor Exosomes: A Double-Edged Sword in Cancer Therapy. Acta Pharmacol. Sin. 2018, 39, 534–541. [Google Scholar] [CrossRef]
- Maisano, D.; Mimmi, S.; Russo, R.; Fioravanti, A.; Fiume, G.; Vecchio, E.; Nisticò, N.; Quinto, I.; Iaccino, E. Uncovering the Exosomes Diversity: A Window of Opportunity for Tumor Progression Monitoring. Pharmaceuticals 2020, 13, 180. [Google Scholar] [CrossRef] [PubMed]
- García-Romero, N.; Carrión-Navarro, J.; Esteban-Rubio, S.; Lázaro-Ibáñez, E.; Peris-Celda, M.; Alonso, M.M.; Guzmán-De-Villoria, J.; Fernández-Carballal, C.; de Mendivil, A.O.; García-Duque, S.; et al. DNA Sequences within Glioma-Derived Extracellular Vesicles Can Cross the Intact Blood-Brain Barrier and Be Detected in Peripheral Blood of Patients. Oncotarget 2017, 8, 1416–1428. [Google Scholar] [CrossRef] [PubMed]
- Ha, D.; Yang, N.; Nadithe, V. Exosomes as Therapeutic Drug Carriers and Delivery Vehicles across Biological Membranes: Current Perspectives and Future Challenges. Acta Pharm. Sin. B 2016, 6, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Touat, M.; Duran-Peña, A.; Alentorn, A.; Lacroix, L.; Massard, C.; Idbaih, A. Emerging Circulating Biomarkers in Glioblastoma: Promises and Challenges. Expert Rev. Mol. Diagn. 2015, 15, 1311–1323. [Google Scholar] [CrossRef]
- Schwarzenbach, H.; Nishida, N.; Calin, G.A.; Pantel, K. Clinical Relevance of Circulating Cell-Free MicroRNAs in Cancer. Nat. Rev. Clin. Oncol. 2014, 11, 145–156. [Google Scholar] [CrossRef]
- Takahashi, R.U.; Miyazaki, H.; Ochiya, T. The Roles of MicroRNAs in Breast Cancer. Cancers 2015, 7, 598–616. [Google Scholar] [CrossRef]
- Ahir, B.K.; Ozer, H.; Engelhard, H.H.; Lakka, S.S. MicroRNAs in Glioblastoma Pathogenesis and Therapy: A Comprehensive Review. Crit. Rev. Oncol. Hematol. 2017, 120, 22–33. [Google Scholar] [CrossRef]
- Ciafrè, S.A.; Galardi, S.; Mangiola, A.; Ferracin, M.; Liu, C.-G.; Sabatino, G.; Negrini, M.; Maira, G.; Croce, C.M.; Farace, M.G. Extensive Modulation of a Set of MicroRNAs in Primary Glioblastoma. Biochem. Biophys. Res. Commun. 2005, 334, 1351–1358. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, J.; Guo, C.; Sai, K.; Wang, J.; Chen, F.; Yang, Q.; Chen, Y.; Wang, J.; To, T.S.; et al. MiR-181b Sensitizes Glioma Cells to Teniposide by Targeting MDM2. BMC Cancer 2014, 14, 611. [Google Scholar] [CrossRef]
- Chen, W.W.; Balaj, L.; Liau, L.M.; Samuels, M.L.; Kotsopoulos, S.K.; Maguire, C.A.; LoGuidice, L.; Soto, H.; Garrett, M.; Zhu, L.D.; et al. BEAMing and Droplet Digital PCR Analysis of Mutant IDH1 MRNA in Glioma Patient Serum and Cerebrospinal Fluid Extracellular Vesicles. Mol. Ther.—Nucleic Acids 2013, 2, e109. [Google Scholar] [CrossRef]
- Siegal, T.; Charbit, H.; Paldor, I.; Zelikovitch, B.; Canello, T.; Benis, A.; Wong, M.L.; Morokoff, A.P.; Kaye, A.H.; Lavon, I. Dynamics of Circulating Hypoxia-Mediated miRNAs and Tumor Response in Patients with High-Grade Glioma Treated with Bevacizumab. J. Neurosurg. 2016, 125, 1008–1015. [Google Scholar] [CrossRef]
- Bronisz, A.; Wang, Y.; Nowicki, M.O.; Peruzzi, P.; Ansari, K.I.; Ogawa, D.; Balaj, L.; De Rienzo, G.; Mineo, M.; Nakano, I.; et al. Extracellular Vesicles Modulate the Glioblastoma Microenvironment via a Tumor Suppression Signaling Network Directed by MiR-1. Cancer Res. 2014, 74, 738–750. [Google Scholar] [CrossRef] [PubMed]
- Shi, J. Considering Exosomal MiR-21 as a Biomarker for Cancer. J. Clin. Med. 2016, 5, 42. [Google Scholar] [CrossRef] [PubMed]
- Lai, N.; Wu, D.; Fang, X.; Lin, Y.; Chen, S.; Li, Z.; Xu, S. Serum MicroRNA-210 as a Potential Noninvasive Biomarker for the Diagnosis and Prognosis of Glioma. Br. J. Cancer 2015, 112, 1241–1246. [Google Scholar] [CrossRef]
- Chevillet, J.; Lee, I.; Briggs, H.; He, Y.; Wang, K. Issues and Prospects of MicroRNA-Based Biomarkers in Blood and Other Body Fluids. Molecules 2014, 19, 6080–6105. [Google Scholar] [CrossRef]
- Catelan, S.; Olioso, D.; Santangelo, A.; Scapoli, C.; Tamanini, A.; Pinna, G.; Sala, F.; Lippi, G.; Nicolato, A.; Cabrini, G.; et al. miRNAs in Serum Exosomes for Differential Diagnosis of Brain Metastases. Cancers 2022, 14, 3493. [Google Scholar] [CrossRef] [PubMed]
- Manterola, L.; Guruceaga, E.; Pérez-Larraya, J.G.; González-Huarriz, M.; Jauregui, P.; Tejada, S.; Diez-Valle, R.; Segura, V.; Samprón, N.; Barrena, C.; et al. A Small Noncoding RNA Signature Found in Exosomes of GBM Patient Serum as a Diagnostic Tool. Neuro. Oncol. 2014, 16, 520–527. [Google Scholar] [CrossRef]
- White, R.J.; Trouche, D.; Martin, K.; Jackson, S.P.; Kouzarides, T. Repression of RNA Polymerase III Transcription by the Retinoblastoma Protein. Nature 1996, 382, 88–90. [Google Scholar] [CrossRef]
- Cabarcas Inhibition of U6 SnRNA Transcription by PTEN. Online J. Biol. Sci. 2010, 10, 114–125. [CrossRef]
- Marshall, L.; White, R.J. Non-Coding RNA Production by RNA Polymerase III Is Implicated in Cancer. Nat. Rev. Cancer 2008, 8, 911–914. [Google Scholar] [CrossRef]
- Su, Y.; Ni, Z.; Wang, G.; Cui, J.; Wei, C.; Wang, J.; Yang, Q.; Xu, Y.; Li, F. Aberrant Expression of MicroRNAs in Gastric Cancer and Biological Significance of MiR-574-3p. Int. Immunopharmacol. 2012, 13, 468–475. [Google Scholar] [CrossRef]
- Tatarano, S.; Chiyomaru, T.; Kawakami, K.; Enokida, H.; Yoshino, H.; Hidaka, H.; Nohata, N.; Yamasaki, T.; Gotanda, T.; Tachiwada, T.; et al. Novel Oncogenic Function of Mesoderm Development Candidate 1 and Its Regulation by MiR-574-3p in Bladder Cancer Cell Lines. Int. J. Oncol. 2012, 40, 951–959. [Google Scholar] [CrossRef]
- Ebrahimkhani, S.; Vafaee, F.; Hallal, S.; Wei, H.; Lee, M.Y.T.; Young, P.E.; Satgunaseelan, L.; Beadnall, H.; Barnett, M.H.; Shivalingam, B.; et al. Deep Sequencing of Circulating Exosomal MicroRNA Allows Non-Invasive Glioblastoma Diagnosis. npj Precis. Oncol. 2018, 2, 28. [Google Scholar] [CrossRef] [PubMed]
- Fm, K.; La, H.; Wl, D.; Es, D.; Hua, Y.; Hao, L.; Cy, P.; Tj, M.; Ma, Q.; Ritner, C. Differentiation Programs in Glioblastoma. Cold Spring Harb. Lab. Press 2015, 2, 25838542. [Google Scholar]
- Jiang, L.; Mao, P.; Song, L.; Wu, J.; Huang, J.; Lin, C.; Yuan, J.; Qu, L.; Cheng, S.Y.; Li, J. MiR-182 as a Prognostic Marker for Glioma Progression and Patient Survival. Am. J. Pathol. 2010, 177, 29–38. [Google Scholar] [CrossRef]
- Song, L.; Lin, C.; Gong, H.; Wang, C.; Liu, L.; Wu, J.; Tao, S.; Hu, B.; Cheng, S.-Y.; Li, M.; et al. MiR-486 Sustains NF-ΚB Activity by Disrupting Multiple NF-ΚB-Negative Feedback Loops. Cell Res. 2013, 23, 274–289. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Zheng, Z.; Zheng, Y.; Lu, X.; Xu, L.; Lin, L. MicroRNA-328 Is a Favorable Prognostic Marker in Human Glioma via Suppressing Invasive and Proliferative Phenotypes of Malignant Cells. Int. J. Neurosci. 2016, 126, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Qiu, S.; Ge, R.; He, L.; Li, M.; Li, Y.; Peng, Y. MiR-340 Suppresses Glioblastoma Multiforme. Oncotarget 2015, 6, 9257–9270. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Yu, J.; Wang, Z.; Zhu, Q.; Wang, W.; Lan, Q. MiR-543 Functions as a Tumor Suppressor in Glioma in Vitro and in Vivo. Oncol. Rep. 2017, 38, 725–734. [Google Scholar] [CrossRef]
- Ueda, R.; Kohanbash, G.; Sasaki, K.; Fujita, M.; Zhu, X.; Kastenhuber, E.R.; McDonald, H.A.; Potter, D.M.; Hamilton, R.L.; Lotze, M.T.; et al. Dicer-Regulated MicroRNAs 222 and 339 Promote Resistance of Cancer Cells to Cytotoxic T-Lymphocytes by down-Regulation of ICAM-1. Proc. Natl. Acad. Sci. USA 2009, 106, 10746–10751. [Google Scholar] [CrossRef] [PubMed]
- Lan, F.; Yue, X.; Xia, T. Exosomal MicroRNA-210 Is a Potentially Non-invasive Biomarker for the Diagnosis and Prognosis of Glioma. Oncol. Lett. 2020, 19, 1967–1974. [Google Scholar] [CrossRef] [PubMed]
- Majmundar, A.J.; Wong, W.J.; Simon, M.C. Hypoxia-Inducible Factors and the Response to Hypoxic Stress. Mol. Cell 2010, 40, 294–309. [Google Scholar] [CrossRef] [PubMed]
- Colwell, N.; Larion, M.; Giles, A.J.; Seldomridge, A.N.; Sizdahkhani, S.; Gilbert, M.R.; Park, D.M. Hypoxia in the Glioblastoma Microenvironment: Shaping the Phenotype of Cancer Stem-like Cells. Neuro. Oncol. 2017, 19, 887–896. [Google Scholar] [CrossRef]
- Dang, K.; Myers, K.A. The Role of Hypoxia-Induced MiR-210 in Cancer Progression. Int. J. Mol. Sci. 2015, 16, 6353–6372. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Ma, J.; Zhou, W.; Zhou, X.; Cao, B.; Fan, D.; Hong, L. Biological Implications and Clinical Value of Mir-210 in Gastrointestinal Cancer. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 539–548. [Google Scholar] [CrossRef]
- Sun, J.; Sun, Z.; Gareev, I.; Yan, T.; Chen, X.; Ahmad, A.; Zhang, D.; Zhao, B.; Beylerli, O.; Yang, G.; et al. Exosomal MiR-2276-5p in Plasma Is a Potential Diagnostic and Prognostic Biomarker in Glioma. Front. Cell Dev. Biol. 2021, 9, 671202. [Google Scholar] [CrossRef]
- Hutagalung, A.H.; Novick, P.J. Role of Rab GTPases in Membrane Traffic and Cell Physiology. Physiol. Rev. 2011, 91, 119–149. [Google Scholar] [CrossRef]
- D’Souza-Schorey, C.; Clancy, J.W. Tumor-Derived Microvesicles: Shedding Light on Novel Microenvironment Modulators and Prospective Cancer Biomarkers. Genes Dev. 2012, 26, 1287–1299. [Google Scholar] [CrossRef]
- Hoshino, D.; Kirkbride, K.C.; Costello, K.; Clark, E.S.; Sinha, S.; Grega-Larson, N.; Tyska, M.J.; Weaver, A.M. Exosome Secretion Is Enhanced by Invadopodia and Drives Invasive Behavior. Cell Rep. 2013, 5, 1159–1168. [Google Scholar] [CrossRef]
- Shao, N.; Xue, L.; Wang, R.; Luo, K.; Zhi, F.; Lan, Q. MiR-454-3p Is an Exosomal Biomarker and Functions as a Tumor Suppressor in Glioma. Mol. Cancer Ther. 2019, 18, 459–469. [Google Scholar] [CrossRef]
- Vo, J.N.; Cieslik, M.; Zhang, Y.; Shukla, S.; Xiao, L.; Zhang, Y.; Wu, Y.-M.; Dhanasekaran, S.M.; Engelke, C.G.; Cao, X.; et al. The Landscape of Circular RNA in Cancer. Cell 2019, 176, 869–881.e13. [Google Scholar] [CrossRef]
- Rybak-Wolf, A.; Stottmeister, C.; Glažar, P.; Jens, M.; Pino, N.; Giusti, S.; Hanan, M.; Behm, M.; Bartok, O.; Ashwal-Fluss, R.; et al. Circular RNAs in the Mammalian Brain Are Highly Abundant, Conserved, and Dynamically Expressed. Mol. Cell 2015, 58, 870–885. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Zhou, H.; Feng, Z.; Xu, Z.; Tang, Y.; Li, P.; Wu, M. CircRNA: Functions and Properties of a Novel Potential Biomarker for Cancer. Mol. Cancer 2017, 16, 94. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zheng, Q.; Bao, C.; Li, S.; Guo, W.; Zhao, J.; Chen, D.; Gu, J.; He, X.; Huang, S. Circular RNA Is Enriched and Stable in Exosomes: A Promising Biomarker for Cancer Diagnosis. Cell Res. 2015, 25, 981–984. [Google Scholar] [CrossRef]
- Kaiser, J. ‘Liquid Biopsy’ for Cancer Promises Earlydetection. Science 2018, 359, 259. [Google Scholar] [CrossRef]
- Dong, W.-W.; Li, H.-M.; Qing, X.-R.; Huang, D.-H.; Li, H.-G. Identification and Characterization of Human Testis Derived Circular RNAs and Their Existence in Seminal Plasma. Sci. Rep. 2016, 6, 39080. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, Y.; Qi, L.; Ding, L.; Jiang, H.; Yu, H. NFIX Circular RNA Promotes Glioma Progression by Regulating MiR-34a-5p via Notch Signaling Pathway. Front. Mol. Neurosci. 2018, 11, 225. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhao, K.; Xu, X.; Yang, Y.; Yan, S.; Wei, P.; Liu, H.; Xu, J.; Xiao, F.; Zhou, H.; et al. A Peptide Encoded by Circular Form of LINC-PINT Suppresses Oncogenic Transcriptional Elongation in Glioblastoma. Nat. Commun. 2018, 9, 4475. [Google Scholar] [CrossRef]
- Yin, K.; Liu, X. CircMMP1promotes the Progression of Glioma through MiR-433 HMGB3 Axis in Vitro and in Vivo. IUBMB Life 2020, 72, 2508–2524. [Google Scholar] [CrossRef]
- Li, P.; Xu, Z.; Liu, T.; Liu, Q.; Zhou, H.; Meng, S.; Feng, Z.; Tang, Y.; Liu, C.; Feng, J.; et al. Circular RNA Sequencing Reveals Serum Exosome Circular RNA Panel for High-Grade Astrocytoma Diagnosis. Clin. Chem. 2022, 68, 332–343. [Google Scholar] [CrossRef]
- Thakur, B.K.; Zhang, H.; Becker, A.; Matei, I.; Huang, Y.; Costa-Silva, B.; Zheng, Y.; Hoshino, A.; Brazier, H.; Xiang, J.; et al. Double-Stranded DNA in Exosomes: A Novel Biomarker in Cancer Detection. Cell Res. 2014, 24, 766–769. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.K.; Song, J.; Huo, H.R.; Zhao, Y.L.; Zhang, G.Y.; Zhao, Z.M.; Sun, G.Z.; Jiao, B.H. DNM3, P65 and P53 from Exosomes Represent Potential Clinical Diagnosis Markers for Glioblastoma Multiforme. Ther. Adv. Med. Oncol. 2017, 9, 741–754. [Google Scholar] [CrossRef] [PubMed]
- Manda, S.V.; Kataria, Y.; Tatireddy, B.R.; Ramakrishnan, B.; Ratnam, B.G.; Lath, R.; Ranjan, A.; Ray, A. Exosomes as a Biomarker Platform for Detecting Epidermal Growth Factor Receptor-Positive High-Grade Gliomas. J. Neurosurg. 2018, 128, 1091–1101. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Huang, A.C.; Zhang, W.; Zhang, G.; Wu, M.; Xu, W.; Yu, Z.; Yang, J.; Wang, B.; Sun, H.; et al. Exosomal PD-L1 Contributes to Immunosuppression and Is Associated with Anti-PD-1 Response. Nature 2018, 560, 382–386. [Google Scholar] [CrossRef]
- Francisco, L.M.; Salinas, V.H.; Brown, K.E.; Vanguri, V.K.; Freeman, G.J.; Kuchroo, V.K.; Sharpe, A.H. PD-L1 Regulates the Development, Maintenance, and Function of Induced Regulatory T Cells. J. Exp. Med. 2009, 206, 3015–3029. [Google Scholar] [CrossRef] [PubMed]
- Litak, J.; Mazurek, M.; Grochowski, C.; Kamieniak, P.; Roliński, J. PD-L1/PD-1 Axis in Glioblastoma Multiforme. Int. J. Mol. Sci. 2019, 20, 5347. [Google Scholar] [CrossRef]
- Shao, H.; Chung, J.; Lee, K.; Balaj, L.; Min, C.; Carter, B.S.; Hochberg, F.H.; Breakefield, X.O.; Lee, H.; Weissleder, R. Chip-Based Analysis of Exosomal MRNA Mediating Drug Resistance in Glioblastoma. Nat. Commun. 2015, 6, 6999. [Google Scholar] [CrossRef]
- Chun, S.; Ahn, S.; Yeom, C.H.; Park, S. Exosome Proteome of U-87MG Glioblastoma Cells. Biology 2016, 5, 50. [Google Scholar] [CrossRef]
- Burgeson, R.E. Type VII Collagen. In Structure and Function of Collagen Types; Elsevier: Amsterdam, The Netherlands, 1987; pp. 145–172. [Google Scholar]
- Stenbeck, G.; Harter, C.; Brecht, A.; Herrmann, D.; Lottspeich, F.; Orci, L.; Wieland, F.T. Beta′-COP, a Novel Subunit of Coatomer. EMBO J. 1993, 12, 2841–2845. [Google Scholar] [CrossRef]
- Markoff, A.; Bogdanova, N.; Knop, M.; Rüffer, C.; Kenis, H.; Lux, P.; Reutelingsperger, C.; Todorov, V.; Dworniczak, B.; Horst, J.; et al. Annexin A5 Interacts with Polycystin-1 and Interferes with the Polycystin-1 Stimulated Recruitment of E-Cadherin into Adherens Junctions. J. Mol. Biol. 2007, 369, 954–966. [Google Scholar] [CrossRef] [PubMed]
- Naryzhny, S.; Volnitskiy, A.; Kopylov, A.; Zorina, E.; Kamyshinsky, R.; Bairamukov, V.; Garaeva, L.; Shlikht, A.; Shtam, T. Proteome of Glioblastoma-Derived Exosomes as a Source of Biomarkers. Biomedicines 2020, 8, 216. [Google Scholar] [CrossRef]
- Daido, S.; Yamamoto, A.; Fujiwara, K.; Sawaya, R.; Kondo, S.; Kondo, Y. Inhibition of the DNA-Dependent Protein Kinase Catalytic Subunit Radiosensitizes Malignant Glioma Cells by Inducing Autophagy. Cancer Res. 2005, 65, 4368–4375. [Google Scholar] [CrossRef]
- Jiang, N.; Shen, Y.; Fei, X.; Sheng, K.; Sun, P.; Qiu, Y.; Larner, J.; Cao, L.; Kong, X.; Mi, J. Valosin-Containing Protein Regulates the Proteasome-Mediated Degradation of DNA-PKcs in Glioma Cells. Cell Death Dis. 2013, 4, e647. [Google Scholar] [CrossRef] [PubMed]
- Lara, P.C.; Pruschy, M.; Zimmermann, M.; Henríquez-Hernández, L.A. MVP and Vaults: A Role in the Radiation Response. Radiat. Oncol. 2011, 6, 148. [Google Scholar] [CrossRef]
- Stavrovskaya, A.A.; Shushanov, S.S.; Rybalkina, E.Y. Problems of Glioblastoma Multiforme Drug Resistance. Biochem. 2016, 81, 91–100. [Google Scholar] [CrossRef]
- Cui, D.; Xu, Q.; Wang, K.; Che, X. Gli1 Is a Potential Target for Alleviating Multidrug Resistance of Gliomas. J. Neurol. Sci. 2010, 288, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Volnitskiy, A.; Shtam, T.; Burdakov, V.; Kovalev, R.; Konev, A.; Filatov, M. Abnormal Activity of Transcription Factors Gli in High-Grade Gliomas. PLoS ONE 2019, 14, e0211980. [Google Scholar] [CrossRef]
- Cumba Garcia, L.M.; Peterson, T.E.; Cepeda, M.A.; Johnson, A.J.; Parney, I.F. Isolation and Analysis of Plasma-Derived Exosomes in Patients With Glioma. Front. Oncol. 2019, 9, 651. [Google Scholar] [CrossRef]
- Qiu, G.; Thakur, A.; Xu, C.; Ng, S.P.; Lee, Y.; Wu, C.M.L. Detection of Glioma-Derived Exosomes with the Biotinylated Antibody-Functionalized Titanium Nitride Plasmonic Biosensor. Adv. Funct. Mater. 2019, 29, 1806761. [Google Scholar] [CrossRef]
- Salomon, C.; Ryan, J.; Sobrevia, L.; Kobayashi, M.; Ashman, K.; Mitchell, M.; Rice, G.E. Exosomal Signaling during Hypoxia Mediates Microvascular Endothelial Cell Migration and Vasculogenesis. PLoS ONE 2013, 8, e68451. [Google Scholar] [CrossRef]
- Pucci, C.; Martinelli, C.; Ciofani, G. Innovative Approaches for Cancer Treatment: Current Perspectives and New Challenges. Ecancermedicalscience 2019, 13, 961. [Google Scholar] [CrossRef] [PubMed]
- Zugazagoitia, J.; Guedes, C.; Ponce, S.; Ferrer, I.; Molina-Pinelo, S.; Paz-Ares, L. Current Challenges in Cancer Treatment. Clin. Ther. 2016, 38, 1551–1566. [Google Scholar] [CrossRef]
- Li, C.; Li, Y.; Li, G.; Wu, S. Functional Nanoparticles for Enhanced Cancer Therapy. Pharmaceutics 2022, 14, 1682. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Jang, H.; Cho, H.; Choi, J.; Hwang, K.Y.; Choi, Y.; Kim, S.H.; Yang, Y. Recent Advances in Exosome-based Drug Delivery for Cancer Therapy. Cancers 2021, 13, 4435. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, L.; Zhu, L.; Xu, Z.; Liu, Y.; Li, Z.; Zhou, J.; Luo, F. Exosomes as Drug Carriers in Anti-Cancer Therapy. Front. Cell Dev. Biol. 2022, 10, 728616. [Google Scholar] [CrossRef]
- Walker, S.; Busatto, S.; Pham, A.; Tian, M.; Suh, A.; Carson, K.; Quintero, A.; Lafrence, M.; Malik, H.; Santana, M.X.; et al. Extracellular Vesicle-Based Drug Delivery Systems for Cancer Treatment. Theranostics 2019, 9, 8001–8017. [Google Scholar] [CrossRef]
- Piffoux, M.; Silva, A.K.A.; Wilhelm, C.; Gazeau, F.; Tareste, D. Modification of Extracellular Vesicles by Fusion with Liposomes for the Design of Personalized Biogenic Drug Delivery Systems. ACS Nano 2018, 12, 6830–6842. [Google Scholar] [CrossRef]
- Zou, Y.; Liu, Y.; Yang, Z.; Zhang, D.; Lu, Y.; Zheng, M.; Xue, X.; Geng, J.; Chung, R.; Shi, B. Effective and Targeted Human Orthotopic Glioblastoma Xenograft Therapy via a Multifunctional Biomimetic Nanomedicine. Adv. Mater. 2018, 30, 1803717. [Google Scholar] [CrossRef]
- Vader, P.; Mol, E.A.; Pasterkamp, G.; Schiffelers, R.M. Extracellular Vesicles for Drug Delivery. Adv. Drug Deliv. Rev. 2016, 106, 148–156. [Google Scholar] [CrossRef]
- Akuma, P.; Okagu, O.D.; Udenigwe, C.C. Naturally Occurring Exosome Vesicles as Potential Delivery Vehicle for Bioactive Compounds. Front. Sustain. Food Syst. 2019, 3, 23. [Google Scholar] [CrossRef]
- Luan, X.; Sansanaphongpricha, K.; Myers, I.; Chen, H.; Yuan, H.; Sun, D. Engineering Exosomes as Refined Biological Nanoplatforms for Drug Delivery. Acta Pharmacol. Sin. 2017, 38, 754–763. [Google Scholar] [CrossRef] [PubMed]
- Butreddy, A.; Kommineni, N.; Dudhipala, N. Exosomes as Naturally Occurring Vehicles for Delivery of Biopharmaceuticals: Insights from Drug Delivery to Clinical Perspectives. Nanomaterials 2021, 11, 1481. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, J.-H.; Chakraborty, K.; Hwang, J.; Lee, Y.-K. Exosome-Based Drug Delivery Systems and Their Therapeutic Applications. RSC Adv. 2022, 12, 18475–18492. [Google Scholar] [CrossRef]
- Liu, Q.; Li, S.; Dupuy, A.; Le Mai, H.; Sailliet, N.; Logé, C.; Robert, J.M.H.; Brouard, S. Exosomes as New Biomarkers and Drug Delivery Tools for the Prevention and Treatment of Various Diseases: Current Perspectives. Int. J. Mol. Sci. 2021, 22, 7763. [Google Scholar] [CrossRef]
- Ghasempour, E.; Hesami, S.; Movahed, E.; keshel, S.H.; Doroudian, M. Mesenchymal Stem Cell-Derived Exosomes as a New Therapeutic Strategy in the Brain Tumors. Stem Cell Res. Ther. 2022, 13, 1–12. [Google Scholar] [CrossRef]
- Tibensky, M.; Jakubechova, J.; Altanerova, U.; Pastorakova, A.; Rychly, B.; Baciak, L.; Mravec, B.; Altaner, C. Gene-Directed Enzyme/Prodrug Therapy of Rat Brain Tumor Mediated by Human Mesenchymal Stem Cell Suicide Gene Extracellular Vesicles In Vitro and In Vivo. Cancers 2022, 14, 735. [Google Scholar] [CrossRef]
- Lee, H.K.; Finniss, S.; Cazacu, S.; Bucris, E.; Ziv-Av, A.; Xiang, C.; Bobbitt, K.; Rempel, S.A.; Hasselbach, L.; Mikkelsen, T.; et al. Mesenchymal Stem Cells Deliver Synthetic MicroRNA Mimics to Glioma Cells and Glioma Stem Cells and Inhibit Their Cell Migration and Self-Renewal. Oncotarget 2013, 4, 346–361. [Google Scholar] [CrossRef]
- Munoz, J.L.; Bliss, S.A.; Greco, S.J.; Ramkissoon, S.H.; Ligon, K.L.; Rameshwar, P. Delivery of Functional Anti-MiR-9 by Mesenchymal Stem Cell-Derived Exosomes to Glioblastoma Multiforme Cells Conferred Chemosensitivity. Mol. Ther.—Nucleic Acids 2013, 2, e126. [Google Scholar] [CrossRef]
- Yan, T.; Wu, M.; Lv, S.; Hu, Q.; Xu, W.; Zeng, A.; Huang, K.; Zhu, X. Exosomes Derived from MicroRNA-512-5p-Transfected Bone Mesenchymal Stem Cells Inhibit Glioblastoma Progression by Targeting JAG1. Aging 2021, 13, 9911–9926. [Google Scholar] [CrossRef]
- Yu, L.; Gui, S.; Liu, Y.; Qiu, X.; Zhang, G.; Zhang, X.; Pan, J.; Fan, J.; Qi, S.; Qiu, B. Exosomes Derived from MicroRNA-199a-Overexpressing Mesenchymal Stem Cells Inhibit Glioma Progression by down-Regulating AGAP2. Aging 2019, 11, 5300–5318. [Google Scholar] [CrossRef]
- Zhang, Z.; Guo, X.; Guo, X.; Yu, R.; Qian, M.; Wang, S.; Gao, X.; Qiu, W.; Guo, Q.; Xu, J.; et al. MicroRNA-29a-3p Delivery via Exosomes Derived from Engineered Human Mesenchymal Stem Cells Exerts Tumour Suppressive Effects by Inhibiting Migration and Vasculogenic Mimicry in Glioma. Aging 2021, 13, 5055–5068. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhao, G.; Zhang, Y.; Jiang, H.; Wang, W.; Zhao, D.; Hong, J.; Yu, H.; Qi, L. Mesenchymal Stem Cell-Derived Exosomal MicroRNA-133b Suppresses Glioma Progression via Wnt/β-Catenin Signaling Pathway by Targeting EZH2. Stem Cell Res. Ther. 2019, 10, 381. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.; Lee, S.; Lee, J.; Kim, M.; Kim, W.J.; Lee, H.W.; Lee, M.Y.; Kim, J.; Chang, W. Exosomes Derived from MicroRNA-584 Transfected Mesenchymal Stem Cells: Novel Alternative Therapeutic Vehicles for Cancer Therapy. BMB Rep. 2018, 51, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Katakowski, M.; Buller, B.; Zheng, X.; Lu, Y.; Rogers, T.; Osobamiro, O.; Shu, W.; Jiang, F.; Chopp, M. Exosomes from Marrow Stromal Cells Expressing MiR-146b Inhibit Glioma Growth. Cancer Lett. 2013, 335, 201–204. [Google Scholar] [CrossRef]
- Parsaei, H.; Moosavifar, M.J.; Eftekharzadeh, M.; Ramezani, R.; Barati, M.; Mirzaei, S.; Nobakht, M. Exosomes to Control Glioblastoma Multiforme: Investigating the Effects of Mesenchymal Stem Cell-Derived Exosomes on C6 Cells in Vitro. Cell Biol. Int. 2022, 46, 2028–2040. [Google Scholar] [CrossRef]
- Rehman, F.U.; Liu, Y.; Yang, Q.; Yang, H.; Liu, R.; Zhang, D.; Muhammad, P.; Liu, Y.; Hanif, S.; Ismail, M.; et al. Heme Oxygenase-1 Targeting Exosomes for Temozolomide Resistant Glioblastoma Synergistic Therapy. J. Control. Release 2022, 345, 696–708. [Google Scholar] [CrossRef]
- Lang, F.M.; Hossain, A.; Gumin, J.; Momin, E.N.; Shimizu, Y.; Ledbetter, D.; Shahar, T.; Yamashita, S.; Parker Kerrigan, B.; Fueyo, J.; et al. Mesenchymal Stem Cells as Natural Biofactories for Exosomes Carrying MiR-124a in the Treatment of Gliomas. Neuro. Oncol. 2018, 20, 380–390. [Google Scholar] [CrossRef]
- Valipour, E.; Ranjbar, F.E.; Mousavi, M.; Ai, J.; Malekshahi, Z.V.; Mokhberian, N.; Taghdiri-Nooshabadi, Z.; Khanmohammadi, M.; Nooshabadi, V.T. The Anti-Angiogenic Effect of Atorvastatin Loaded Exosomes on Glioblastoma Tumor Cells: An in Vitro 3D Culture Model. Microvasc. Res. 2022, 143, 104385. [Google Scholar] [CrossRef]
- Nooshabadi, V.T.; Khanmohammadi, M.; Shafei, S.; Banafshe, H.R.; Malekshahi, Z.V.; Ebrahimi-Barough, S.; Ai, J. Impact of Atorvastatin Loaded Exosome as an Anti-Glioblastoma Carrier to Induce Apoptosis of U87 Cancer Cells in 3D Culture Model. Biochem. Biophys. Rep. 2020, 23, 100792. [Google Scholar] [CrossRef]
- Li, B.; Chen, X.; Qiu, W.; Zhao, R.; Duan, J.; Zhang, S.; Pan, Z.; Zhao, S.; Guo, Q.; Qi, Y.; et al. Synchronous Disintegration of Ferroptosis Defense Axis via Engineered Exosome-Conjugated Magnetic Nanoparticles for Glioblastoma Therapy. Adv. Sci. 2022, 9, 2105451. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.Z.; Lai, M.F.; Li, Y.P.; Xu, C.H.; Zhang, H.R.; Kuang, J.G. Human Marrow Stromal Cells Secrete MicroRNA-375-Containing Exosomes to Regulate Glioma Progression. Cancer Gene Ther. 2020, 27, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Hao, S.C.; Ma, H.; Niu, Z.F.; Sun, S.Y.; Zou, Y.R.; Xia, H.C. HUC-MSCs Secreted Exosomes Inhibit the Glioma Cell Progression through PTENP1/MIR-10a-5p/PTEN Pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 10013–10023. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhu, X.; Li, C.; Cai, L.; Pei, W.; Ni, M.; He, J.; Jiang, H.; Chen, J. Efficient Exosome Extraction through the Conjugation of Superparamagnetic Iron Oxide Nanoparticles for the Targeted Delivery in Rat Brain. Mater. Today Chem. 2022, 23, 100637. [Google Scholar] [CrossRef]
- Liu, W.; Wei, L.; Li, M.; Mo, J. Zinc Sulfide-Based Hybrid Exosome-Coated Nanoplatform for Targeted Treatment of Glioblastoma in an Orthotopic Mouse Glioblastoma Model. Mater. Today Adv. 2023, 17, 100327. [Google Scholar] [CrossRef]
- Gečys, D.; Kazlauskas, A.; Gečytė, E.; Paužienė, N.; Kulakauskienė, D.; Lukminaitė, I.; Jekabsone, A. Internalisation of RGD-Engineered Extracellular Vesicles by Glioblastoma Cells. Biology 2022, 11, 1483. [Google Scholar] [CrossRef]
- Zhu, Q.; Ling, X.; Yang, Y.; Zhang, J.; Li, Q.; Niu, X.; Hu, G.; Chen, B.; Li, H.; Wang, Y.; et al. Embryonic Stem Cells-Derived Exosomes Endowed with Targeting Properties as Chemotherapeutics Delivery Vehicles for Glioblastoma Therapy. Adv. Sci. 2019, 6, 1801899. [Google Scholar] [CrossRef]
- Ludwig, B.S.; Kessler, H.; Kossatz, S.; Reuning, U. Rgd-Binding Integrins Revisited: How Recently Discovered Functions and Novel Synthetic Ligands (Re-)Shape an Ever-Evolving Field. Cancers 2021, 13, 1711. [Google Scholar] [CrossRef]
- Alvarez-Erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; Wood, M.J.A. Delivery of SiRNA to the Mouse Brain by Systemic Injection of Targeted Exosomes. Nat. Biotechnol. 2011, 29, 341–345. [Google Scholar] [CrossRef]
- Hung, M.E.; Leonard, J.N. Stabilization of Exosome-Targeting Peptides via Engineered Glycosylation. J. Biol. Chem. 2015, 290, 8166–8172. [Google Scholar] [CrossRef]
- Rahmani, R.; Kiani, J.; Tong, W.Y.; Soleimani, M.; Voelcker, N.H.; Arefian, E. Engineered Anti-EGFRvIII Targeted Exosomes Induce Apoptosis in Glioblastoma Multiforme. J. Drug Target. 2023, 31, 310–319. [Google Scholar] [CrossRef]
- Epidermal Growth Factor Receptor Variant III (EGFRvIII); Prominin 1 (PROM1; CD133). Sci.-Bus. Exch. 2014, 7, 172. [CrossRef]
- Parker Kerrigan, B.C.; Ledbetter, D.; Kronowitz, M.; Phillips, L.; Gumin, J.; Hossain, A.; Yang, J.; Mendt, M.; Singh, S.; Cogdell, D.; et al. RNAi Technology Targeting the FGFR3-TACC3 Fusion Breakpoint: An Opportunity for Precision Medicine. Neuro-Oncol. Adv. 2020, 2, vdaa132. [Google Scholar] [CrossRef]
- Chen, B.; Sun, Y.; Zhang, J.; Zhu, Q.; Yang, Y.; Niu, X.; Deng, Z.; Li, Q.; Wang, Y. Human Embryonic Stem Cell-Derived Exosomes Promote Pressure Ulcer Healing in Aged Mice by Rejuvenating Senescent Endothelial Cells. Stem Cell Res. Ther. 2019, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Shi, H.Y.; Ma, Y.M.; Liu, J.W. Neural Stem Cell Therapy for Brain Disease. World J. Stem Cells 2021, 13, 1278–1292. [Google Scholar] [CrossRef]
- Adamus, T.; Hung, C.Y.; Yu, C.; Kang, E.; Hammad, M.; Flores, L.; Nechaev, S.; Zhang, Q.; Gonzaga, J.M.; Muthaiyah, K.; et al. Glioma-Targeted Delivery of Exosome-Encapsulated Antisense Oligonucleotides Using Neural Stem Cells. Mol. Ther.—Nucleic Acids 2022, 27, 611–620. [Google Scholar] [CrossRef]
- Qian, C.; Wang, Y.; Ji, Y.; Chen, D.; Wang, C.; Zhang, G.; Wang, Y. Neural Stem Cell-Derived Exosomes Transfer MiR-124-3p into Cells to Inhibit Glioma Growth by Targeting FLOT2. Int. J. Oncol. 2022, 61, 115. [Google Scholar] [CrossRef]
- Mahla, R.S.; Kumar, A.; Tutill, H.J.; Krishnaji, S.T.; Sathyamoorthy, B.; Noursadeghi, M.; Breuer, J.; Pandey, A.K.; Kumar, H. NIX-Mediated Mitophagy Regulate Metabolic Reprogramming in Phagocytic Cells during Mycobacterial Infection. Tuberculosis 2021, 126, 102046. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, P.; Dong, Y.; Xie, H.; Wang, Y.; Soto, F.; Ma, P.; Feng, X.; Du, W.; Liu, B.F. Designer Exosomes Enabling Tumor Targeted Efficient Chemo/Gene/Photothermal Therapy. Biomaterials 2021, 276, 121056. [Google Scholar] [CrossRef]
- Jia, G.; Han, Y.; An, Y.; Ding, Y.; He, C.; Wang, X.; Tang, Q. NRP-1 Targeted and Cargo-Loaded Exosomes Facilitate Simultaneous Imaging and Therapy of Glioma in Vitro and in Vivo. Biomaterials 2018, 178, 302–316. [Google Scholar] [CrossRef]
- Wu, T.; Liu, Y.; Cao, Y.; Liu, Z. Engineering Macrophage Exosome Disguised Biodegradable Nanoplatform for Enhanced Sonodynamic Therapy of Glioblastoma. Adv. Mater. 2022, 34, 2110364. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Kong, J.; Ma, S.; Li, J.; Mao, M.; Chen, K.; Chen, Z.; Zhang, J.; Chang, Y.; Yuan, H.; et al. Exosome-Coated 10B Carbon Dots for Precise Boron Neutron Capture Therapy in a Mouse Model of Glioma In Situ. Adv. Funct. Mater. 2021, 31, 2100969. [Google Scholar] [CrossRef]
- Liang, S.; Xu, H.; Ye, B.C. Membrane-Decorated Exosomes for Combination Drug Delivery and Improved Glioma Therapy. Langmuir 2022, 38, 299–308. [Google Scholar] [CrossRef]
- Shan, S.; Chen, J.; Sun, Y.; Wang, Y.; Xia, B.; Tan, H.; Pan, C.; Gu, G.; Zhong, J.; Qing, G.; et al. Functionalized Macrophage Exosomes with Panobinostat and PPM1D-SiRNA for Diffuse Intrinsic Pontine Gliomas Therapy. Adv. Sci. 2022, 9, 2200353. [Google Scholar] [CrossRef]
- Shi, L.; Cao, Y.; Yuan, W.; Guo, J.; Sun, G. Exosomal CircRNA BTG2 Derived from RBP-J Overexpressed-Macrophages Inhibits Glioma Progression via MiR-25-3p/PTEN. Cell Death Dis. 2022, 13, 506. [Google Scholar] [CrossRef]
- Yao, J.; Wang, Z.; Cheng, Y.; Ma, C.; Zhong, Y.; Xiao, Y.; Gao, X.; Li, Z. M2 Macrophage-Derived Exosomal MicroRNAs Inhibit Cell Migration and Invasion in Gliomas through PI3K/AKT/MTOR Signaling Pathway. J. Transl. Med. 2021, 19, 99. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Liu, Y.; Guo, K.; Zhang, K.; Liu, Q.; Wang, P.; Wang, X. Ultrasound Facilitates Naturally Equipped Exosomes Derived from Macrophages and Blood Serum for Orthotopic Glioma Treatment. ACS Appl. Mater. Interfaces 2019, 11, 14576–14587. [Google Scholar] [CrossRef] [PubMed]
- Fan, B.; Yang, S.; Wang, Y.-Y.; Zhang, C.; Yang, J.-P.; Wang, L.-Q.; Lv, Z.-Q.; Shi, X.-F.; Fan, Z.-Z.; Yang, J.-K. Indocyanine Green-Loaded Exosomes for Image-Guided Glioma Nano-Therapy. J. Exp. Nanosci. 2022, 17, 187–196. [Google Scholar] [CrossRef]
- Wang, R.; Liang, Q.; Zhang, X.; Di, Z.; Wang, X.; Di, L. Tumor-Derived Exosomes Reversing TMZ Resistance by Synergistic Drug Delivery for Glioma-Targeting Treatment. Colloids Surf. B Biointerfaces 2022, 215, 112505. [Google Scholar] [CrossRef]
- Zeng, A.; Wei, Z.; Yan, W.; Yin, J.; Huang, X.; Zhou, X.; Li, R.; Shen, F.; Wu, W.; Wang, X.; et al. Exosomal Transfer of MiR-151a Enhances Chemosensitivity to Temozolomide in Drug-Resistant Glioblastoma. Cancer Lett. 2018, 436, 10–21. [Google Scholar] [CrossRef]
- Lee, H.; Bae, K.; Baek, A.R.; Kwon, E.-B.; Kim, Y.H.; Nam, S.W.; Lee, G.H.; Chang, Y. Glioblastoma-Derived Exosomes as Nanopharmaceutics for Improved Glioma Treatment. Pharmaceutics 2022, 14, 1002. [Google Scholar] [CrossRef] [PubMed]
- Salarpour, S.; Pardakhty, A.; Ahmadi-Zeidabadi, M.; Pournamdari, M.; Forootanfar, H.; Esmaeeli, M.; Afsharipour, S. Exosome-Loaded Paclitaxel: Preparation and Toxicity Evaluation on Two Glioblastoma Cell Lines. Nanomed. Res. J. 2019, 4, 239–246. [Google Scholar] [CrossRef]
- Wang, H.; Feng, J.; Ao, F.; Tang, Y.; Xu, P.; Wang, M.; Huang, M. Tumor-Derived Exosomal MicroRNA-7-5p Enhanced by Verbascoside Inhibits Biological Behaviors of Glioblastoma in Vitro and in Vivo. Mol. Ther.—Oncolytics 2021, 20, 569–582. [Google Scholar] [CrossRef]
- Thakur, A.; Sidu, R.K.; Zou, H.; Alam, M.K.; Yang, M.; Lee, Y. Inhibition of Glioma Cells’ Proliferation by Doxorubicin-Loaded Exosomes via Microfluidics. Int. J. Nanomed. 2020, 15, 8331–8343. [Google Scholar] [CrossRef] [PubMed]
- Tanziela, T.; Shaikh, S.; ur Rehman, F.; Semcheddine, F.; Jiang, H.; Lu, Z.; Wang, X. Cancer-Exocytosed Exosomes Loaded with Bio-Assembled AgNCs as Smart Drug Carriers for Targeted Chemotherapy. Chem. Eng. J. 2022, 440, 135980. [Google Scholar] [CrossRef]
- Liu, H.; Chen, L.; Liu, J.; Meng, H.; Zhang, R.; Ma, L.; Wu, L.; Yu, S.; Shi, F.; Li, Y.; et al. Co-Delivery of Tumor-Derived Exosomes with Alpha-Galactosylceramide on Dendritic Cell-Based Immunotherapy for Glioblastoma. Cancer Lett. 2017, 411, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Shi, J.; Xie, J.; Wang, Y.; Sun, J.; Liu, T.; Zhao, Y.; Zhao, X.; Wang, X.; Ma, Y.; et al. Large-Scale Generation of Functional MRNA-Encapsulating Exosomes via Cellular Nanoporation. Nat. Biomed. Eng. 2020, 4, 69–83. [Google Scholar] [CrossRef]
- Zhan, Q.; Yi, K.; Qi, H.; Li, S.; Li, X.; Wang, Q.; Wang, Y.; Liu, C.; Qiu, M.; Yuan, X.; et al. Engineering Blood Exosomes for Tumor-Targeting Efficient Gene/Chemo Combination Therapy. Theranostics 2020, 10, 7889–7905. [Google Scholar] [CrossRef]
- Cui, J.; Wang, X.; Li, J.; Zhu, A.; Du, Y.; Zeng, W.; Guo, Y.; Di, L.; Wang, R. Immune Exosomes Loading Self-Assembled Nanomicelles Traverse the Blood-Brain Barrier for Chemo-Immunotherapy against Glioblastoma. ACS Nano 2023, 17, 1464–1484. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Q.; Yi, K.; Cui, X.; Li, X.; Yang, S.; Wang, Q.; Fang, C.; Tan, Y.; Li, L.; Xu, C.; et al. Blood Exosomes-Based Targeted Delivery of CPLA2 SiRNA and Metformin to Modulate Glioblastoma Energy Metabolism for Tailoring Personalized Therapy. Neuro. Oncol. 2022, 24, 1871–1883. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xia, T.; Fang, Y.; Zuo, H.; Dong, X.; Xu, P.; Ouyang, J. Overcoming the Blood–Brain Barrier by Using a Multistage Exosome Delivery System to Inhibit Central Nervous System Lymphoma. Nanomed. Nanotechnol. Biol. Med. 2022, 41, 102523. [Google Scholar] [CrossRef]
- Zhuang, X.; Xiang, X.; Grizzle, W.; Sun, D.; Zhang, S.; Axtell, R.C.; Ju, S.; Mu, J.; Zhang, L.; Steinman, L.; et al. Treatment of Brain Inflammatory Diseases by Delivering Exosome Encapsulated Anti-Inflammatory Drugs from the Nasal Region to the Brain. Mol. Ther. 2011, 19, 1769–1779. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Martin, P.; Fogarty, B.; Brown, A.; Schurman, K.; Phipps, R.; Yin, V.P.; Lockman, P.; Bai, S. Exosome Delivered Anticancer Drugs across the Blood-Brain Barrier for Brain Cancer Therapy in Danio rerio. Pharm. Res. 2015, 32, 2003–2014. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.B.; Yang, A.C.; Yousef, H.; Lee, D.; Chen, W.; Schaum, N.; Lehallier, B.; Quake, S.R.; Wyss-Coray, T. Brain Endothelial Cells Are Exquisite Sensors of Age-Related Circulatory Cues. Cell Rep. 2020, 30, 4418–4432.e4. [Google Scholar] [CrossRef]
- Yang, T.; Fogarty, B.; LaForge, B.; Aziz, S.; Pham, T.; Lai, L.; Bai, S. Delivery of Small Interfering RNA to Inhibit Vascular Endothelial Growth Factor in Zebrafish Using Natural Brain Endothelia Cell-Secreted Exosome Nanovesicles for the Treatment of Brain Cancer. AAPS J. 2017, 19, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Kim, M.; Lee, Y.; Byun, J.W.; Hwang, D.W.; Lee, M. Systemic Delivery of MicroRNA-21 Antisense Oligonucleotides to the Brain Using T7-Peptide Decorated Exosomes. J. Control. Release 2020, 317, 273–281. [Google Scholar] [CrossRef]
- Monfared, H.; Jahangard, Y.; Nikkhah, M.; Mirnajafi-Zadeh, J.; Mowla, S.J. Potential Therapeutic Effects of Exosomes Packed with a MiR-21-Sponge Construct in a Rat Model of Glioblastoma. Front. Oncol. 2019, 9, 782. [Google Scholar] [CrossRef]
- Aqil, F.; Munagala, R.; Jeyabalan, J.; Agrawal, A.K.; Kyakulaga, A.H.; Wilcher, S.A.; Gupta, R.C. Milk Exosomes—Natural Nanoparticles for SiRNA Delivery. Cancer Lett. 2019, 449, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Tepeoglu, M.; Borcek, P.; Ozen, O.; Altinors, M.N. Microsatellite Instability in Glioblastoma: Is It Really Relevant in Tumor Prognosis? Turk. Neurosurg. 2019, 29. [Google Scholar] [CrossRef]
- Ou, A.; Yung, W.K.A.; Majd, N. Molecular Mechanisms of Treatment Resistance in Glioblastoma. Int. J. Mol. Sci. 2020, 22, 351. [Google Scholar] [CrossRef]
- Delgado-Martín, B.; Medina, M.Á. Advances in the Knowledge of the Molecular Biology of Glioblastoma and Its Impact in Patient Diagnosis, Stratification, and Treatment. Adv. Sci. 2020, 7, 1902971. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.S.; Lin, E.Y.; Chiou, T.W.; Harn, H.J. Exosomes in Clinical Trial and Their Production in Compliance with Good Manufacturing Practice. Tzu Chi Med. J. 2020, 32, 113–120. [Google Scholar] [CrossRef]
- Charoenviriyakul, C.; Takahashi, Y.; Morishita, M.; Matsumoto, A.; Nishikawa, M.; Takakura, Y. Cell Type-Specific and Common Characteristics of Exosomes Derived from Mouse Cell Lines: Yield, Physicochemical Properties, and Pharmacokinetics. Eur. J. Pharm. Sci. 2017, 96, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Takahashi, Y.; Nishikawa, M.; Takakura, Y. Effect of Exosome Isolation Methods on Physicochemical Properties of Exosomes and Clearance of Exosomes from the Blood Circulation. Eur. J. Pharm. Biopharm. 2016, 98, 1–8. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug | Chemical Structure | Free | Exosome Encapsulated | Reference |
|---|---|---|---|---|
| 5-FC |  | Non-active | Tumor growth suppression of C6 GBM cells | [199] |
| TMZ |  | Non-active | Inhibition of the TMZ-resistant GBM growth in cell lines | [209,235] |
| Atorvastatin |  | Reduced tumor-targeting efficiency | Tumor growth inhibition with reduced cell propagation, migration and VEGF secretion | [211,212] |
| BQR |  | Limited BBB penetration | Improved ferroptosis, BBB penetration and tumor-targeting ability | [213] |
| DOX |  | Limited BBB penetration | High apoptotic effect and tumor growth suppression | [216,231,239,247,250,253] |
| PTX |  | Limited BBB penetration | High cytotoxicity and improvement of therapeutic effects through optimized targeting | [219,244] |
| Cur |  | Limited BBB penetration and tumor-targeting ability as well as poor stability in liquid environment | Increased imaging and therapeutic effects with reduced side effects and augmented survival in mice models | [232,254] |
| ICG |  | Limited BBB penetration, tumor-targeting ability and poor stability in aqueous solution | Relief of the intracellular hypoxia microenvironment, BBB penetration, tumor accumulation and potent anticancer effect | [233,240] |
| Panobinostat |  | Limited BBB penetration and poor water solubility | Inhibition of DIPG cell proliferation | [236] |
| HCQ |  | Limited BBB penetration and non-selective distribution of the drug in vivo | Strong antitumor activity | [217] |
| TMZ & DHT |  | Non-active/TMZ-resistance inversion and apoptosis induction | Good antitumor efficacy and targeting of the tumor site of TMZ-resistant cells and mice models | [241] |
| Selumetinib |  | Limited BBB penetration | Strong antitumor effect with zero toxicity in vivo | [243] |
| VB |  | Strong anticancer effect in vitro and in vivo | Not studied | [245] |
| DOX & PTX |  | Limited BBB penetration | Autologous exosomes represent the most potent weapon for glioma drug targeting with increased cytotoxicity and BBB penetration | [246,255] |
| TanIIA and GL |  | Poor solubility in water and quick metabolism/Self-assembly with fat-soluble ingredients for nanomicelle formation | Great BBB penetration, apoptosis inducement and when co-administered with TMZ an enhanced synergistic effect as well as an inhibition of postoperative recurrence was observed | [251] |
| Metformin |  | Limited BBB penetration, poor stability an inefficient cell uptake | Increased cell uptake, suppression of tumor growth and significant extension of mice survival time | [252] |
| Cell Source | Encapsulated Cargo | Outcome In Vitro | Outcome In Vivo | Reference | |
|---|---|---|---|---|---|
| Macromolecule | Drug | ||||
| Mesenchymal stem cell-derived exosomes | yCD::UPRT | 5-FC | Tumor growth inhibition of C6 GBM cells | [199] | |
| miR-124 and miR-145 | n.p. | Reduced migration rate of tumor cells and diminished the regeneration capacity of GSCs | U87-derived xenograft penetration and downregulation of miR-124 target genes, such as CDK6 | [200] | |
| Anti-miR-9 | n.p. | Chemosensitivity, increased cell death as well as elevated caspase activity | n.s. | [201] | |
| miR-512-5p | n.p. | Cell propagation inhibition and cell cycle arrest induction | n.s. | [202] | |
| miR-199a | n.p. | Suppression in proliferation, invasion and migration as well as TMZ chemosensitivity induction | Repression of tumor growth | [203] | |
| miR-29a-3p | n.p. | Migration and vasculogenic mimicry formation attenuation | [204] | ||
| miR-133b | n.p. | Suppression in proliferation, invasion and migration | n.s. | [205] | |
| miR-584 | n.p. | Decreased proliferation and invasion rate of glioma cells | Attenuation of malignant gliomas development | [206] | |
| miR-146b | n.p. | n.s. | Glioma xenograft growth inhibition | [207] | |
| n.p. | n.p. | Apoptosis induction in C6 cells | n.s. | [208] | |
| STAT3-siRNA | TMZ | Inhibition of TMZ-resistant GBM growth | [209] | ||
| miR-124a | n.p. | Significant reduction in viability and clonogenicity of GSCs | Tumor growth retention and increased survival | [210] | |
| n.p. | Atorvastatin | Reduced cell propagation, migration and VEGF secretion as well as increased apoptotic activity | n.s. | [211] | |
| n.p. | Atorvastatin | Tumor growth suppression in a 3D GBM model | n.s. | [212] | |
| GPX4-siRNA | BQR | Enhanced ferroptosis in GBM cell lines | BBB penetration and tumor-targeting ability | [213] | |
| miR-375 | n.p. | Inhibition in cell propagation, migration and infiltration as well as apoptosis induction | Repression of xenograft tumor in nude mice | [214] | |
| lncRNA PTENP1 | n.p. | Suppression of malignant phenotype of U87 cells | n.s. | [215] | |
| n.p. | DOX | Increased cytotoxicity against cancer cells | n.s. | [216] | |
| Cytosine deaminase and miR-34a | n.p. | Increased apoptosis | n.s. | [223] | |
| FGFR3-TACC3 siRNA | n.p. | Decreased cell viability | Improved survival in glioma-bearing mice without toxicity to adjacent normal tissues and affecting the wild-type FGFR3 or TACC3 | [225] | |
| Embryonic stem cells-derived exosomes | n.p. | PTX | Improvement in the therapeutic effects of PTX through optimized targeting | n.s. | [219] |
| Neural stem cells-derived exosomes | STAT3 ASOs | n.p. | Stimulation of the immunogenic activity of human dendritic cells or mouse macrophages | Inhibit growth of GL261 tumors | [228] |
| miR-124-3p | n.p. | Glioma cell proliferation, migration, and invasion as well as FLOT2 expression suppression | Tumor growth inhibition in a mouse tumor xenograft model | [229] | |
| Macrophage-derived exosomes | miR-21 | DOX | High apoptotic effect in glioma cells | n.s. | [231] |
| n.p. | Curcumin | Targeted imaging and curative effects with reduced side effects | [232] | ||
| Catalase | ICG | Relief of the intracellular hypoxia microenvironment | BBB penetration, brain accumulation ability and strong anticancer effect | [233] | |
| n.p. | n.p. | n.s. | 100% Mice survival ratio | [234] | |
| O6-benzylguanine (BG) | TMZ | Proliferation inhibition of U87MG and GSCs | n.s. | [235] | |
| PPM1D-siRNA | Panobinostat | DIPG cell proliferation inhibition | n.s. | [236] | |
| circBTG2 | n.p. | Inhibition in cell proliferation and invasion | n.s. | [237] | |
| miR-15a and miR-92a | n.p. | Inhibition of malignant behavior of glioma cells | n.s. | [238] | |
| n.p. | DOX | Tumor growth suppression | [239] | ||
| Glioma cells-derived exosomes | n.p. | HCQ | High antitumor potency with visible light illumination | [217] | |
| n.p. | ICG | n.s. | Abolishment of tumor growth | [240] | |
| n.p. | TMZ and DHT | Good antitumor efficacy and targeting of the tumor site of TMZ-resistant cells and mice models | [241] | ||
| miR-151a | n.p. | chemosensitivity recovery to TMZ | [242] | ||
| n.p. | Selumetinib | Strong antitumor effect | Strong antitumor effect with zero toxicity | [243] | |
| n.p. | PTX | High cytotoxicity to U87 and T98G cell lines | n.s. | [244] | |
| miR-7-5p | VB | Inhibition of cell proliferation, invasion, and migration as well as vessel-like tube formation | Diminish tumor formation and metastasis | [245] | |
| n.p. | DOX and PTX | Autologous exosomes represent the most potent weapon for glioma drug targeting | n.s. | [246] | |
| [Ag (GSH)]+ | DOX | High glioma cells uptake and constant DOX release in cancerous cells | n.s. | [247] | |
| alpha-Galactosylceramide | n.p. | n.s. | Stimulation of tumor-specific cytotoxic T lymphocytes proliferation, synergistically breaking the immune tolerance and improving the immunosuppressive environment | [248] | |
| Blood exosomes | n.p. | DOX | Neoplastic growth suppression | [239] | |
| PTEN mRNA | n.p. | n.s. | Restoration of tumor suppression, enhancement of tumor growth inhibition, and improvement of survival in an orthotopic PTEN-deficient glioma mouse | [249] | |
| n.p. | DOX and miR-21i | Acceleration of the endocytic uptake and enhanced tumor accumulation ability | Improved and synergistic antitumor effect | [250] | |
| n.p. | TanIIA and GL | Great BBB penetration and apoptosis inducement in GL261 cells | Extension of mice median survival and when co-administered with TMZ, a synergistic therapeutic effect was recorded as well as inhibition of postoperative recurrence | [251] | |
| cPLA2-siRNA | Metformin | Increased cellular uptake and strong antitumor effect | Inhibition of tumor growth and significant extension of mice survival time | [252] | |
| Natural brain endothelia cell-derived exosomes | VEGF-siRNA | n.p. | BBB penetration efficiency and increased cellular uptake | Strong BBB penetration and aggregation abortion of xenotransplanted cancer cells | [257] |
| n.p. | DOX & PTX | High cytotoxicity and intracellular uptake to U87 cell line | Efficient BBB penetration | [255] | |
| 293T cell-derived exosomes | AMO-21 | n.p. | Increased delivery efficiency to C6 GBM cells | Expression induction of PDCD4 and PTEN as well as tumor shrinkage | [258] |
| miR-21-sponge construct | n.p. | Cell proliferation reduction and increased apoptosis | Tumor growth retention and tumor retardation inducement as well as reduction in tumors volume | [259] | |
| n.p. | DOX | Increased apoptosis | Life expectancy extension and antitumor activity enhancement | [253] | |
| n.p. | Cur/ STAT3-inhibitor | n.s. | Abortion in tumor growth and mice survival increase | [254] | |
| Milk exosomes | siRNA against a plethora of oncogenes | n.p. | Knockdown of oncogenes and reduced cell proliferation | n.s. | [260] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avgoulas, D.I.; Tasioulis, K.S.; Papi, R.M.; Pantazaki, A.A. Therapeutic and Diagnostic Potential of Exosomes as Drug Delivery Systems in Brain Cancer. Pharmaceutics 2023, 15, 1439. https://doi.org/10.3390/pharmaceutics15051439
Avgoulas DI, Tasioulis KS, Papi RM, Pantazaki AA. Therapeutic and Diagnostic Potential of Exosomes as Drug Delivery Systems in Brain Cancer. Pharmaceutics. 2023; 15(5):1439. https://doi.org/10.3390/pharmaceutics15051439
Chicago/Turabian StyleAvgoulas, Dimitrios I., Konstantinos S. Tasioulis, Rigini M. Papi, and Anastasia A. Pantazaki. 2023. "Therapeutic and Diagnostic Potential of Exosomes as Drug Delivery Systems in Brain Cancer" Pharmaceutics 15, no. 5: 1439. https://doi.org/10.3390/pharmaceutics15051439