

Versatility of Liposomes for Antisense Oligonucleotide Delivery: A Special Focus on Various Therapeutic Areas


 , , ,
, , , 
Abstract
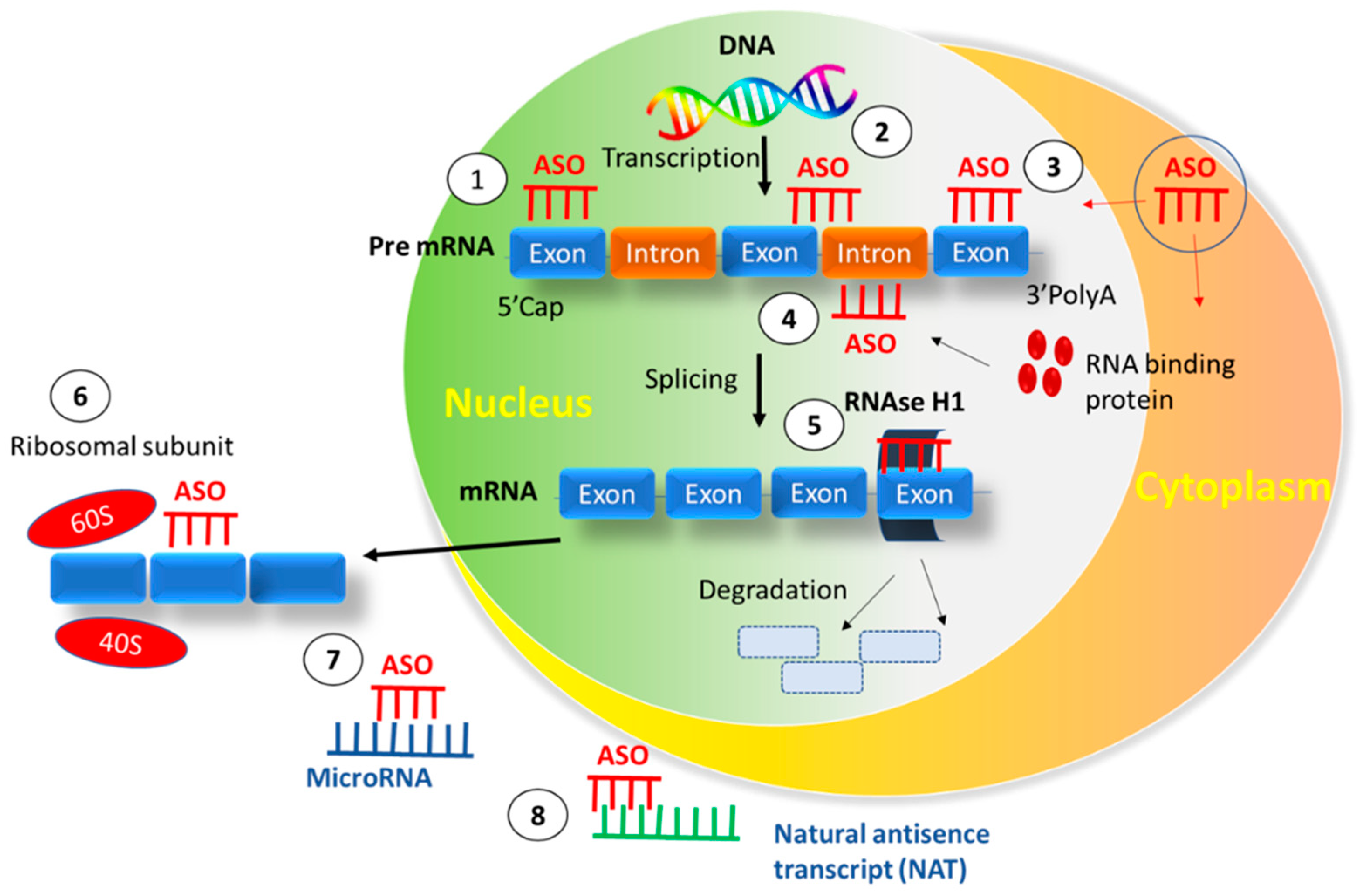
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
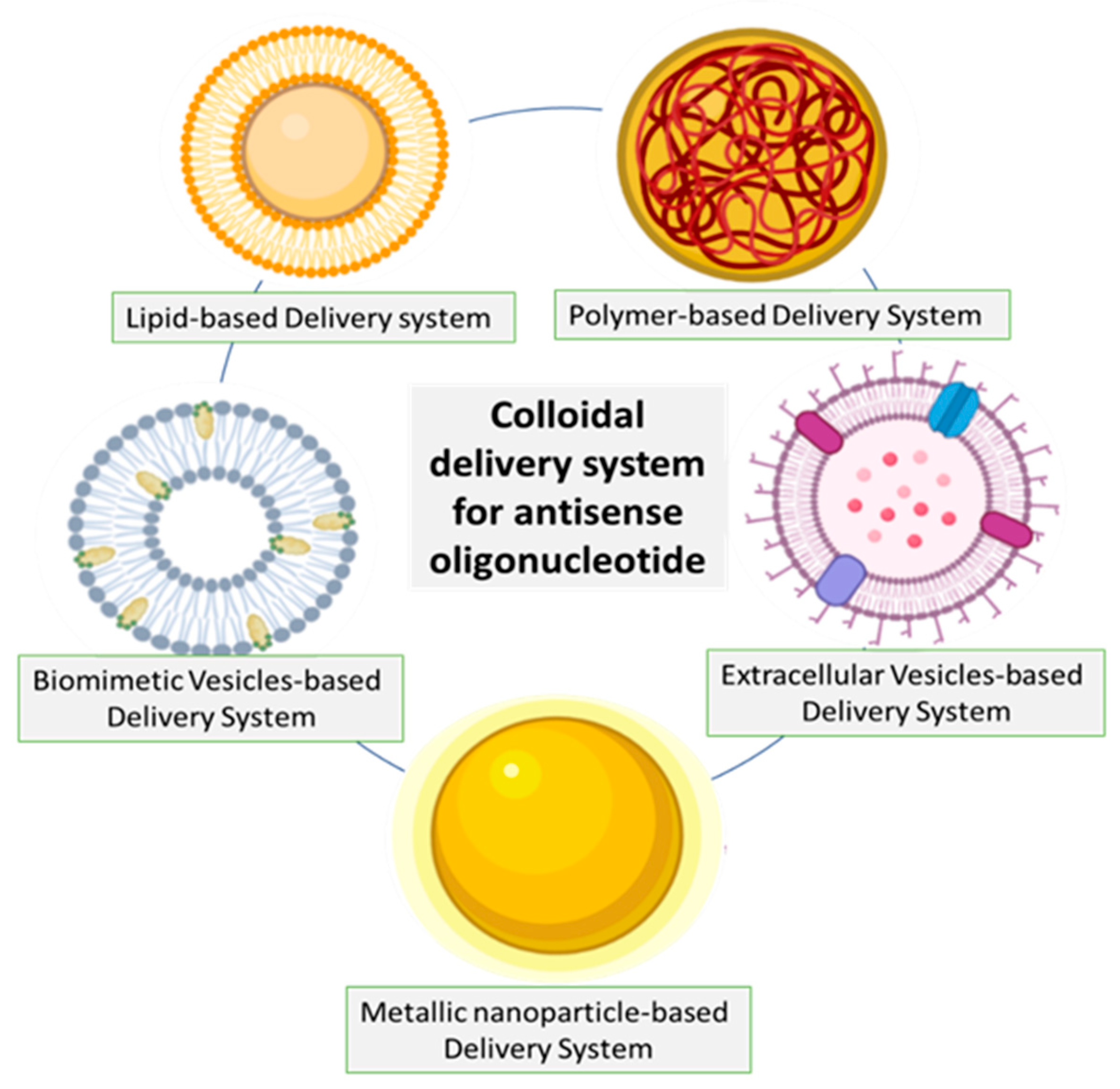{kind=link}
| Name of ASO/ Company | Chemical Class | Target Organ | Route | Mechanism of Action | Indication | Approval Year |
|---|---|---|---|---|---|---|
| Vitravene (Fomivirsen) Ionis Pharma, Novartis | Phosphorothioate oligonucleotide | Eye | Intravitreal | Inhibit the replication of human CMV | Cytomegalovirus Retinitis | 1998 |
| Kynamro (Mipomersen) Sanofi/Isis | 20-mer synthetic second-generation 2′-methoxyethyl antisense oligonucleotide | Liver | Subcutaneous | Hamper translation of ApoB-100 mRNA | Homozygous familial hypercholesterolemia | 2013 |
| Spinraza (Nusinersen) Biogen/Ionis | 2′-O-(2-methoxyethyl) phosphorothioate antisense oligonucleotide | Central Nervous System | Intrathecal | Inhibits the SMN2 pre-RNA splicing | Spinal muscular atrophy | 2016 |
| Tegsedi (Inotersen) Ionis | 2′-O (2-methoxyethyl)-modified phosphorothioate antisense oligonucleotide (ASO) | Liver | Subcutaneous | Degradation of wild-type and mutant TTR mRNA. | Hereditary-transthyretin mediated amyloidosis | 2018 |
| Waylivra (Volanesorsen) Ionis | 2′-O-2-methoxyethyl (2′-MOE) antisense oligonucleotide (ASO) | Liver | Subcutaneous | Inhibits apo-lipoprotein C-III | Familial Chylomicronaemia | 2019 |
| Amondys 45 (Casimersen) Sarepta therapeutics | Phosphorodiamidate morpholino oligomer (PMO) | Muscle | Intravenous | Bind to exon 45 of the Duchenne muscular dystrophy (DMD) pre-mRNA and prevent translation | DMD | 2021 |
| Viltolarsen (Viltepso) NS Pharma | Phosphorodiamidate morpholino oligomer (PMO) | Muscle | Intravenous | Exclude the exon via binding to exon 53 of dystrophin pre-mRNA | DMD | 2020 |
| Exondys 51 (Eteplirsen) Sarepta therapeutics | Phosphorodiamidate morpholino oligomer (PMO) | Muscle | Intravenous | Alteration in exon splicing by binding to dystrophin | DMD | 2016 |
| Golodirsen (Vyondys 53) Sarepta therapeutics | Phosphorodiamidate morpholino oligomer (PMO) subclass | Muscle | Intravenous | Induce exon splicing | DMD | 2019 |
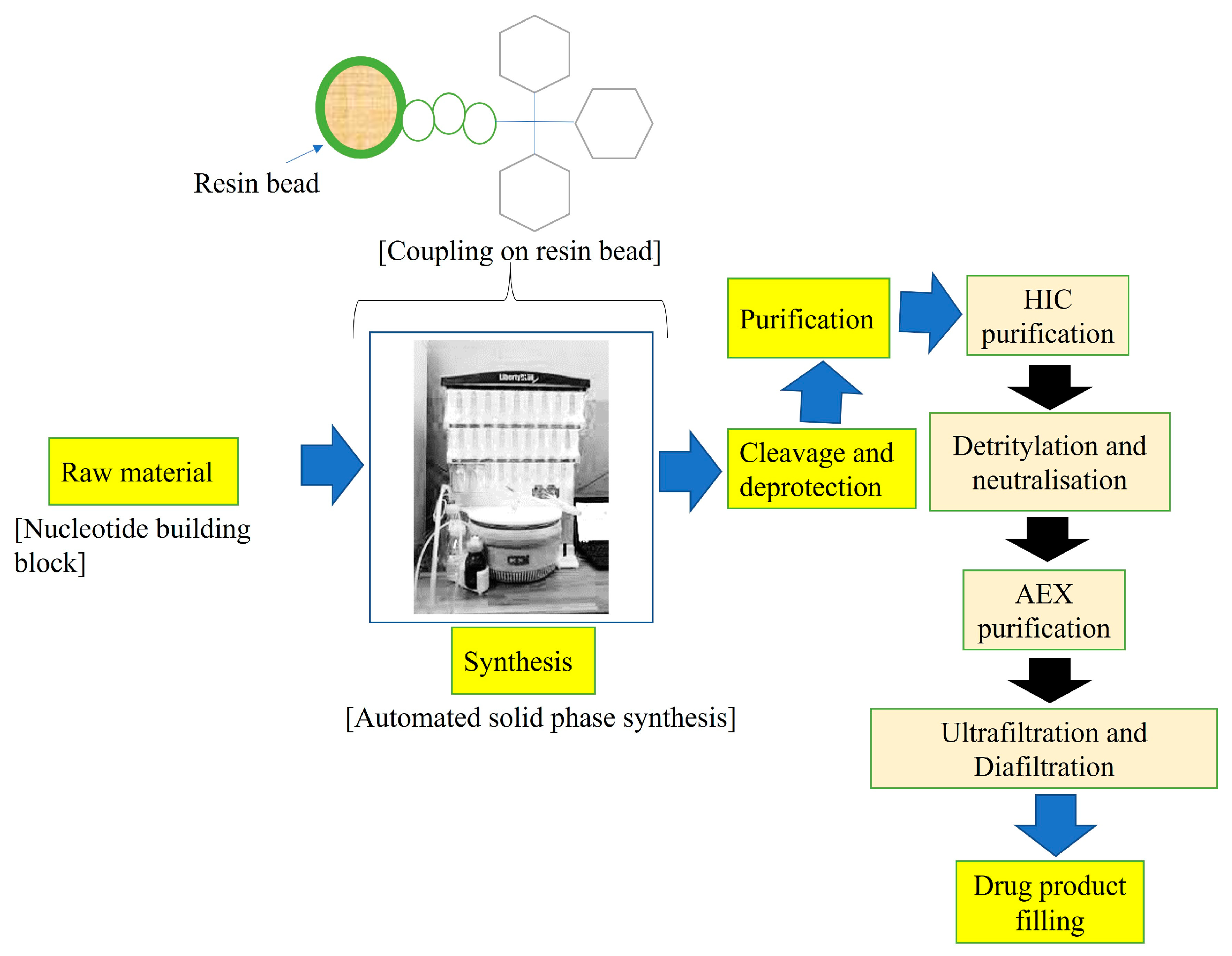
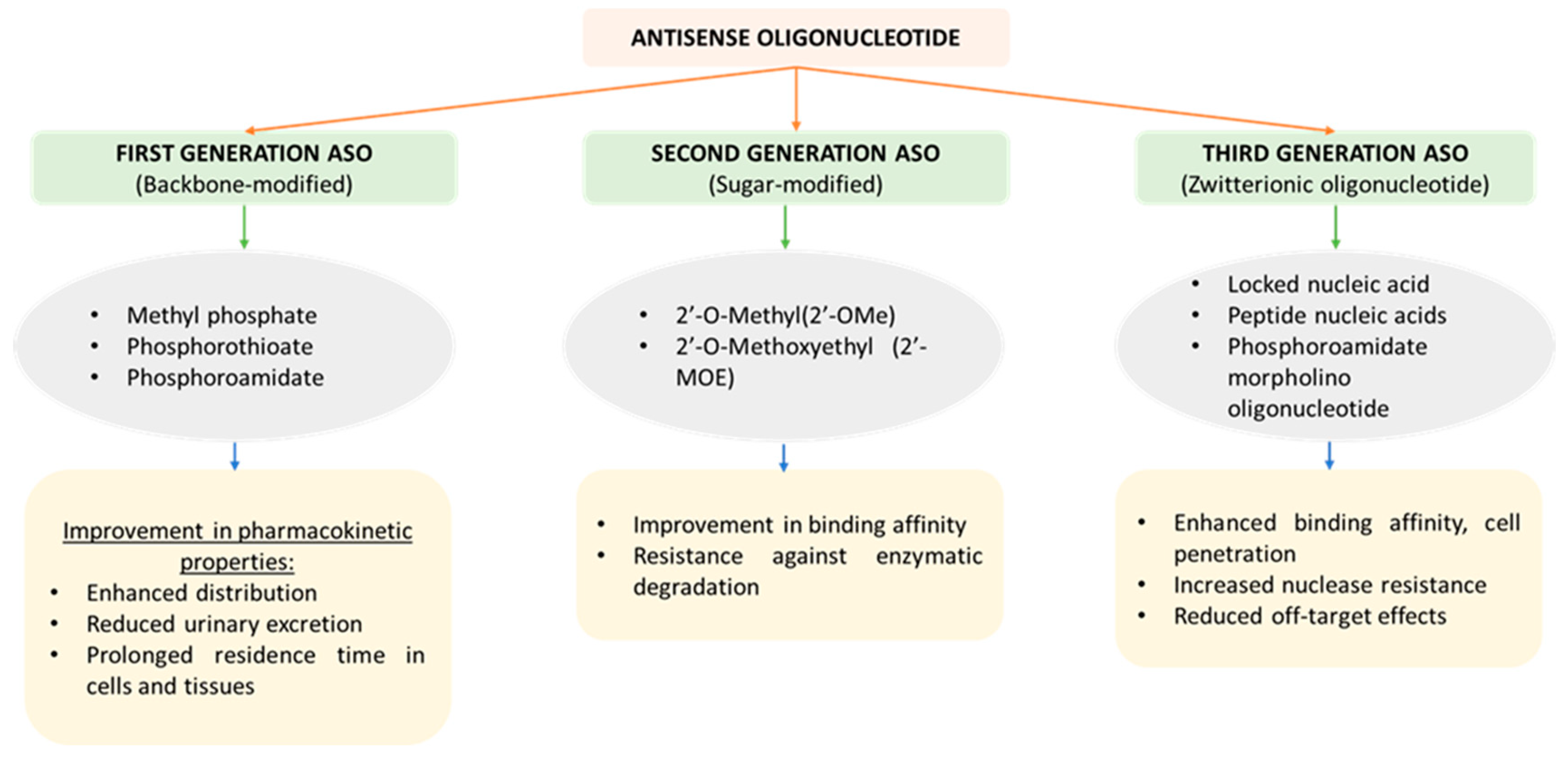
2. Synthesis and Generations of ASOs
2.1. First-Generation ASO
2.2. Second-Generation ASO
2.3. Third-Generation ASO
3. Interaction between ASO and Cell Membrane
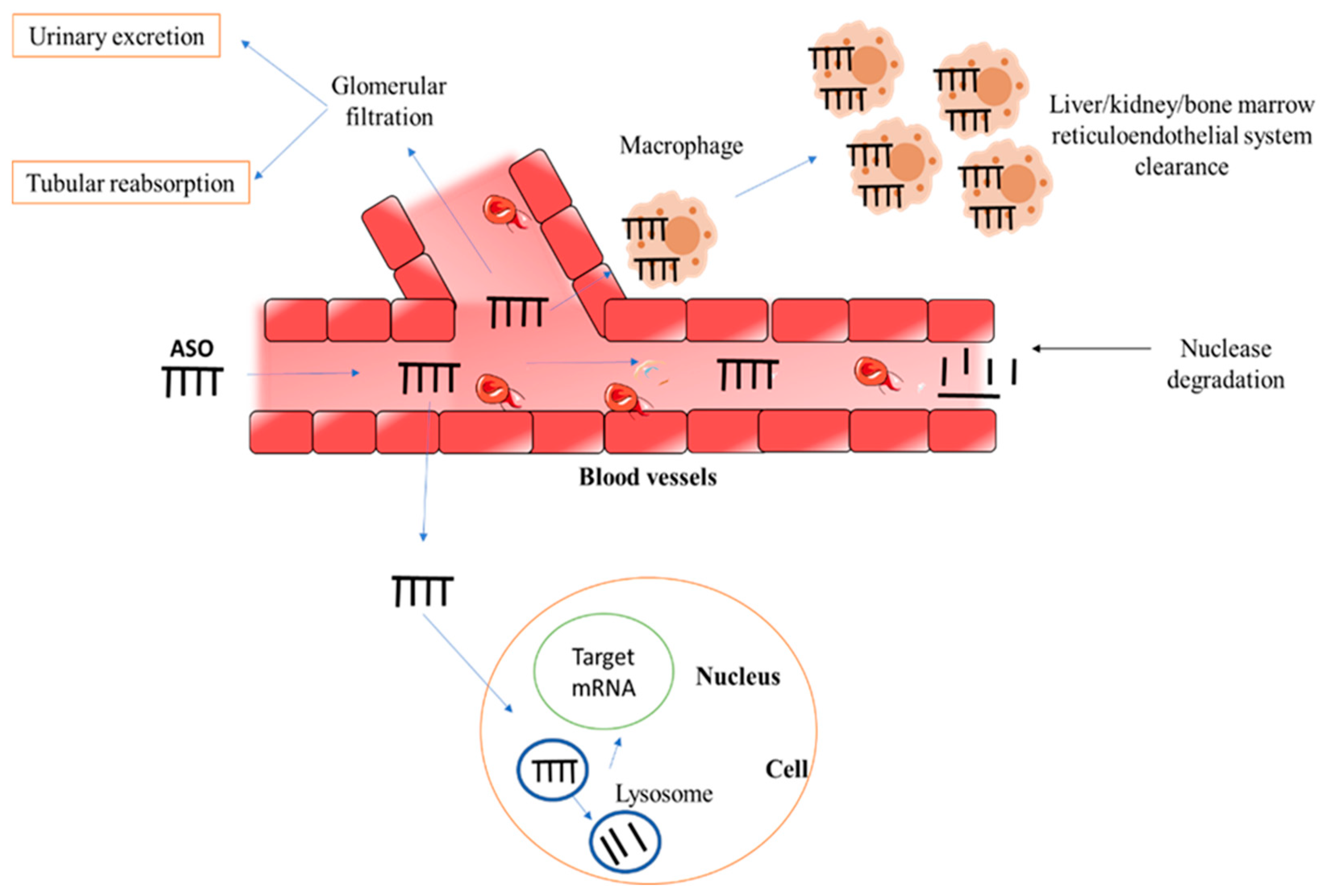
4. Stability of ASOs
5. Route of Administration
6. Approaches for ASO Delivery
7. Liposomes as a Promising Approach for ASO Delivery
8. Evaluation of Liposomal ASO Formulation
9. Applications of Liposomal ASO Formulations
9.1. Myotonic Dystrophy
9.2. Respiratory Diseases
9.3. Myocardium Dysfunction
9.4. Hepatic Disorders
9.5. Treatment of Colitis
9.6. Antiviral Therapy
9.7. Ophthalmic Therapy
9.8. Antibacterial Therapy
9.9. Anti-Parasite Therapy
9.10. Anticancer Therapy
10. Limitations of Liposomes for ASO Delivery
- (i)
- Binding of the liposome to the cell surface;
- (ii)
- Endocytosis-mediated entry of the liposome into the cells or direct traversing of the plasma membrane (e.g., via membrane fusion);
- (iii)
- Liposomal escape from the endosome;
- (iv)
- Dissociation of the liposome to release the nucleic acid payload;
- (v)
- Transport through the cytosol;
- (vi)
- Entry into the nucleus.
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Anti-GD2 | Anti-disialoganglioside |
| ASGPR | Asialoglycoprotein receptor |
| ASO | Antisense oligonucleotides |
| CCLs | Coated cationic liposomes |
| CHEMS | Cholesterylhemisuccinate |
| CLCN1 | Chloride channel 1 gene |
| CLL | Chronic lymphocytic leukemia |
| CMV | Cytomegalovirus |
| DMD | Duchenne mus-cular dystrophy |
| DMPG | Dimiristoyl phosphatidylglycerol |
| D-oligos | Antisense deoxyoligonucleotides |
| DOPE | Dioleoyl-phosphatidylethanolamine |
| DOTAP | Dioleoyl-3-trimethylammonium propane |
| DPPC | Dipalmitoyl phosphatidylcholine |
| DSC | Differential scanning calorimetry |
| DSPE | 1,2-distearoyl-sn-glycero-3-phosphoethanolamine |
| EPC | Egg phosphatidylcholine |
| HPLC | High-performance liquid chromatography |
| L-cL | Liposome-coated lipoplexes |
| MP-oligos | Methyl phosphonate |
| mRNA | Messenger RNA |
| PCS | Photon correlation spectroscopy |
| PEG | Polyethylene glycol |
| PMO | Phosphorodiamidate morpholino oligomer |
| PMO | Phosphorodiamidate morpholino oligonucleotide |
| TGF-β2 | Transforming growth factor beta-2 |
| Tm | Glass transition temperature |
| VEGF | Vascular endothelial growth factor |
References
- Wang, Y.; Miao, L.; Satterlee, A.; Huang, L. Delivery of Oligonucleotides with Lipid Nanoparticles. Adv. Drug Deliv. Rev. 2015, 87, 68. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, J.A.; Witzigmann, D.; Thomson, S.B.; Chen, S.; Leavitt, B.R.; Cullis, P.R.; van der Meel, R. The current landscape of nucleic acid therapeutics. Nat. Nanotechnol. 2021, 16, 630–643. [Google Scholar] [CrossRef] [PubMed]
- Kerlin, R.L.; Li, X. Pathology in Non-Clinical Drug Safety Assessment. In Haschek and Rousseaux’s Handbook of Toxicologic Pathology; Elseiver: Amsterdam, The Netherlands, 2013; pp. 725–750. [Google Scholar]
- Roberts, T.C.; Langer, R.; Wood, M.J.A. Advances in oligonucleotide drug delivery. Nat. Rev. Drug Discov. 2020, 19, 673–694. [Google Scholar] [CrossRef] [PubMed]
- Jacob, D. How Do Antisense Oligonucleotides Work? Available online: https://www.rxlist.com/neurologics_antisense_oligonucleotides/drug-class.htm (accessed on 13 March 2023).
- Di Fusco, D.; Dinallo, V.; Marafini, I.; Figliuzzi, M.M.; Romano, B.; Monteleone, G. Antisense oligonucleotide: Basic concepts and therapeutic application in inflammatory bowel disease. Front. Pharmacol. 2019, 10, 305. [Google Scholar] [CrossRef]
- Drug Approval Package: Vitravene (Fomivirsen Sodium Intravitreal Injectable) NDA# 20-961. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/nda/98/20961_vitravene.cfm (accessed on 13 March 2023).
- European Medicines Agency (EMA); Committee for Medicinal Products for Human Use (CHMP). Assessment Report—Kynamro (Mipomersen). 2013; Volume 44, pp. 1–114. Available online: https://www.ema.europa.eu/en/documents/assessment-report/kynamro-epar-public-assessment-report_en.pdf (accessed on 25 March 2023).
- SPINRAZA (Nusinersen) Injection, for Intrathecal Use. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2016/209531lbl.pdf (accessed on 13 March 2023).
- Tegsedi (Inotersen). Available online: Ema.europa.eu/en/documents/overview/tegsedi-epar-summary-public_en.pdf (accessed on 13 March 2023).
- Gales, L. Tegsedi (Inotersen): An Antisense Oligonucleotide Approved for the Treatment of Adult Patients with Hereditary Transthyretin Amyloidosis. Pharmaceuticals 2019, 12, 78. [Google Scholar] [CrossRef]
- Waylivra. Available online: https://www.ema.europa.eu/en/medicines/human/EPAR/waylivra (accessed on 13 March 2023).
- AMONDYS 45 (Casimersen) Injection, for Intravenous Use. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2021/213026lbl.pdf (accessed on 13 March 2023).
- LABEL: VILTEPSO-Viltolarsen Injection, Solution. Available online: https://dailymed.nlm.nih.gov/dailymed/drugInfo.cfm?setid=1ffff9a8-6d6a-4dcb-8493-1b6cc3a5d123 (accessed on 13 March 2023).
- EXONDYS 51 (Eteplirsen) Injection, for Intravenous Use. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2016/206488lbl.pdf (accessed on 13 March 2023).
- VYONDYS 53 (Golodirsen) Injection, for Intravenous Use. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2019/211970s000lbl.pdf (accessed on 13 March 2023).
- Nakamura, A.; Ali, S.A.; Kapoor, M. Antisense oligonucleotide-based therapies for the treatment of osteoarthritis: Opportunities and roadblocks. Bone 2020, 138, 115461. [Google Scholar] [CrossRef]
- Urban, E.; Noe, C.R. Structural modifications of antisense oligonucleotides. Farmaco 2003, 58, 243–258. [Google Scholar] [CrossRef]
- Sharma, V.K.; Sharma, R.K.; Singh, S.K. Antisense oligonucleotides: Modifications and clinical trials. MedChemComm 2014, 5, 1454–1471. [Google Scholar] [CrossRef]
- Bege, M.; Borbás, A. Rise and fall of fomivirsen, the first approved gene silencing medicine—A historical review. Acta Pharm. Hung. 2022, 92, 38–44. [Google Scholar] [CrossRef]
- Dirin, M.; Winkler, J. Influence of diverse chemical modifications on the ADME characteristics and toxicology of antisense oligonucleotides. Expert Opin. Biol. Ther. 2013, 13, 875–888. [Google Scholar] [CrossRef]
- Yu, R.Z.; Grundy, J.S.; Geary, R.S. Clinical pharmacokinetics of second generation antisense oligonucleotides. Expert Opin. Drug Metab. Toxicol. 2013, 9, 169–182. [Google Scholar] [CrossRef]
- Altmann, K.H.; Martin, P.; Dean, N.M.; Mania, B.P. Second Generation Antisense Oligonucleotides—Inhibition of PKC-a and c-raf Kinase Expression by Chimeric Oligonucleotides Incorporating 6”-Substituted Carbocyclic Nucleosides and 2″-O-Ethylene Glycol Substituted Ribonucleosides. Nucl. Nucl. 2006, 16, 917–926. [Google Scholar] [CrossRef]
- Griffey, R.H.; Monia, B.P.; Cummins, L.L.; Freier, S.; Greig, M.J.; Guinosso, C.J.; Lesnik, E.; Manalili, S.M.; Mohan, V.; Owens, S.; et al. 2′-O-aminopropyl ribonucleotides: A zwitterionic modification that enhances the exonuclease resistance and biological activity of antisense oligonucleotides. J. Med. Chem. 1996, 39, 5100–5109. [Google Scholar] [CrossRef]
- Gagliardi, M.; Ashizawa, A.T. The Challenges and Strategies of Antisense Oligonucleotide Drug Delivery. Biomedicines 2021, 9, 433. [Google Scholar] [CrossRef]
- Quemener, A.M.; Centomo, M.L.; Sax, S.L.; Panella, R. Small Drugs, Huge Impact: The Extraordinary Impact of Antisense Oligonucleotides in Research and Drug Development. Molecules 2022, 27, 536. [Google Scholar] [CrossRef]
- Quemener, A.M.; Bachelot, L.; Forestier, A.; Donnou-Fournet, E.; Gilot, D.; Galibert, M.D. The powerful world of antisense oligonucleotides: From bench to bedside. Wiley Interdiscip. Rev. RNA 2020, 11, e1594. [Google Scholar] [CrossRef]
- Loke, S.L.; Stein, C.A.; Zhang, X.H.; Mori, K.; Nakanishi, M.; Subasinghe, C.; Cohen, J.S.; Neckers, L.M. Characterization of oligonucleotide transport into living cells. Proc. Natl. Acad. Sci. USA 1989, 86, 3474–3478. [Google Scholar] [CrossRef]
- Stein, C.A.; Mori, K.; Loke, S.L.; Subasinghe, C.; Shinozuka, K.; Cohen, J.S.; Neckers, L.M. Phosphorothioate and normal oligodeoxyribonucleotides with 5′-linked acridine: Characterization and preliminary kinetics of cellular uptake. Gene 1988, 72, 333–341. [Google Scholar] [CrossRef]
- Miller, P.S.; McParland, K.B.; Jayaraman, K.; Ts’o, P.O.P. Biochemical and Biological Effects of Nonionic Nucleic Acid Methylphosphonatest. Biochemistry 1981, 20, 1874–1880. [Google Scholar] [CrossRef]
- Akhtar, S.; Basu, S.; Wickstrom, E.; Juliano, R.L. Interactions of antisense DNA oligonucleotide analogs with phospholipid membranes (liposomes). Nucleic Acids Res. 1991, 19, 5551. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, C.; Wood, M.J.A. Antisense oligonucleotides: The next frontier for treatment of neurological disorders. Nat. Rev. Neurol. 2017, 14, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Eder, P.S.; Devine, R.J.; Dagle, J.M.; Walder, J.A. Substrate Specificity and Kinetics of Degradation of Antisense Oligonucleotides by a 3′ Exonuclease in Plasma. Antisense Res. Dev. 2009, 1, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Karaki, S.; Paris, C.; Rocchi, P. Antisense Oligonucleotides, A Novel Developing Targeting Therapy. In Antisense Therapy; Intechopen: London, UK, 2019. [Google Scholar]
- White, P.J.; Anastasopoulos, F.; Pouton, C.W.; Boyd, B.J. Overcoming biological barriers to in vivo efficacy of antisense oligonucleotides. Expert Rev. Mol. Med. 2009, 11, e10. [Google Scholar] [CrossRef] [PubMed]
- Muslehiddinoglu, J.; Simler, R.; Hill, M.L.; Mueller, C.; Amery, J.H.A.; Dixon, L.; Watson, A.; Storch, K.; Gazziola, C.; Gielen, F.; et al. Technical Considerations for Use of Oligonucleotide Solution API. Nucleic Acid Ther. 2020, 30, 189–197. [Google Scholar] [CrossRef]
- Tari, A.M.; Lopez-Berestein, G. Oligonucleotide Therapy for Hematological Malignancies. J. Liposome Res. 2008, 7, 19–30. [Google Scholar] [CrossRef]
- Yadava, P.; Gibbs, M.; Castro, C.; Hughes, J.A. Effect of lyophilization and freeze-thawing on the stability of siRNA-liposome complexes. AAPS PharmSciTech 2008, 9, 335–341. [Google Scholar] [CrossRef]
- Meissner, J.M.; Toporkiewicz, M.; Czogalla, A.; Matusewicz, L.; Kuliczkowski, K.; Sikorski, A.F. Novel antisense therapeutics delivery systems: In vitro and in vivo studies of liposomes targeted with anti-CD20 antibody. J. Control. Release 2015, 220, 515–528. [Google Scholar] [CrossRef]
- Zhong, J.; Yao, X.; Li, D.L.; Li, L.Q.; Zhou, L.F.; Huang, H.L.; Min, L.S.; Li, J.; Fu, F.F.; Dai, L.C. Large scale preparation of midkine antisense oligonucleotides nanoliposomes by a cross-flow injection technique combined with ultrafiltration and high-pressure extrusion procedures. Int. J. Pharm. 2013, 441, 712–720. [Google Scholar] [CrossRef]
- Bochot, A.; Couvreur, P.; Fattal, E. Intravitreal administration of antisense oligonucleotides: Potential of liposomal delivery. Prog. Retin. Eye Res. 2000, 19, 131–147. [Google Scholar] [CrossRef]
- Juliano, R.L.; Akhtar, S. Liposomes as a Drug Delivery System for Antisense Oligonucleotides. Antisense Res. Dev. 1992, 2, 165–176. [Google Scholar] [CrossRef]
- Liu, M.; Li, M.; Sun, S.; Li, B.; Du, D.; Sun, J.; Cao, F.; Li, H.; Jia, F.; Wang, T.; et al. The use of antibody modified liposomes loaded with AMO-1 to deliver oligonucleotides to ischemic myocardium for arrhythmia therapy. Biomaterials 2014, 35, 3697–3707. [Google Scholar] [CrossRef]
- Peddada, L.Y.; Garbuzenko, O.B.; Devore, D.I.; Minko, T.; Roth, C.M. Delivery of antisense oligonucleotides using poly(alkylene oxide)–poly(propylacrylic acid) graft copolymers in conjunction with cationic liposomes. J. Control. Release 2014, 194, 103–112. [Google Scholar] [CrossRef]
- Brignole, C.; Pastorino, F.; Marimpietri, D.; Pagnan, G.; Pistorio, A.; Allen, T.M.; Pistioa, V.; Ponzoni, M. Immune cell-mediated antitumor activities of GD2- targeted liposomal c-myb antisense oligonucleotides containing CpG motifs. J. Natl. Cancer Inst. 2004, 96, 1171–1180. [Google Scholar] [CrossRef]
- Wyrozumska, P.; Meissner, J.; Toporkiewicz, M.; Szarawarska, M.; Kuliczkowski, K.; Ugorski, M.; Walasek, M.A.; Sikorski, A.F. Liposome-coated lipoplex–based carrier for antisense oligonucleotides. Cancer Biol. 2015, 16, 66–76. [Google Scholar] [CrossRef]
- Yao, Y.; Feng, L.; Wang, Z.; Chen, H.; Tan, N. Programmed delivery of cyclopeptide RA-V and antisense oligonucleotides for combination therapy on hypoxic tumors and for therapeutic self-monitoring. Biomater. Sci. 2020, 8, 256–265. [Google Scholar] [CrossRef]
- Rudin, C.M.; Marshall, J.L.; Huang, C.H.; Kindler, H.L.; Zhang, C.; Kumar, D.; Gokhale, P.C.; Steinberg, J.; Wanaski, S.; Kasid, U.N.; et al. Delivery of a Liposomal c-raf-1 Antisense Oligonucleotide by Weekly Bolus Dosing in Patients with Advanced Solid TumorsA Phase I Study. Clin. Cancer Res. 2004, 10, 7244–7251. [Google Scholar] [CrossRef]
- Dritschilo, A.; Huang, C.H.; Rudin, C.M.; Marshall, J.; Collins, B.; Dul, J.L.; Zhang, C.; Kumar, D.; Gokhale, P.C.; Ahmad, A.; et al. Phase I study of liposome-encapsulated c-raf antisense oligodeoxyribonucleotide infusion in combination with radiation therapy in patients with advanced malignancies. Clin. Cancer Res. 2006, 12, 1251–1259. [Google Scholar] [CrossRef]
- Garbuzenko, O.B.; Saad, M.; Betigeri, S.; Zhang, M.; Vetcher, A.A.; Soldatenkov, V.A.; Reimer, D.C.; Pozharov, V.P.; Minko, T. Intratracheal versus intravenous liposomal delivery of siRNA, antisense oligonucleotides and anticancer drug. Pharm. Res. 2009, 26, 382–394. [Google Scholar] [CrossRef]
- Kilicay, E.; Karahaliloglu, Z.; Alpaslan, P.; Hazer, B.; Denkbas, E.B. In vitro evaluation of antisense oligonucleotide functionalized core-shell nanoparticles loaded with α-tocopherol succinate. J. Biomater. Sci. Polym. Ed. 2017, 28, 1762–1785. [Google Scholar] [CrossRef]
- Gong, N.; Teng, X.; Li, J.; Liang, X.J. Antisense Oligonucleotide-Conjugated Nanostructure-Targeting lncRNA MALAT1 Inhibits Cancer Metastasis. ACS Appl. Mater. Interfaces 2019, 11, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Luo, S.; Zhang, J.; Yu, T.; Fu, Z.; Zheng, Y.; Xu, X.; Liu, C.; Fan, M.; Zhang, Z. Exosome-mediated delivery of antisense oligonucleotides targeting α-synuclein ameliorates the pathology in a mouse model of Parkinson’s disease. Neurobiol. Dis. 2021, 148, 105218. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Pang, J.; Wang, Q.; Yan, L.; Wang, L.; Xing, Z.; Wang, C.; Zhang, J.; Dong, L.; Wang, Y.; et al. Delivering Antisense Oligonucleotides across the Blood-Brain Barrier by Tumor Cell-Derived Small Apoptotic Bodies. Adv. Sci. 2021, 8, 2004929. [Google Scholar] [CrossRef] [PubMed]
- Hoyer, D.; Zorrilla, E.P.; Cottone, P.; Parylak, S.; Morelli, M.; Simola, N.; Simola, N.; Morelli, M.; Lane, J.D.; Morgan, M.M.; et al. Chemical Modifications of Antisense Oligonucleotides. In Encyclopedia of Psychopharmacology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 277–278. [Google Scholar]
- Geary, R.S.; Norris, D.; Yu, R.; Bennett, C.F. Pharmacokinetics, biodistribution and cell uptake of antisense oligonucleotides. Adv. Drug Deliv. Rev. 2015, 87, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Brad Wan, W.; Seth, P.P. The Medicinal Chemistry of Therapeutic Oligonucleotides. J. Med. Chem. 2016, 59, 9645–9667. [Google Scholar]
- Bijsterbosch, M.K.; Manoharan, M.; Rump, E.T.; De Vrueh, R.L.A.; Van Veghel, R.; Tivel, K.L.; Biessen, E.A.L.; Bennett, C.F.; Dan Cook, P.; Van Berkel, T.J.C. In vivo fate of phosphorothioate antisense oligodeoxynucleotides: Predominant uptake by scavenger receptors on endothelial liver cells. Nucleic Acids Res. 1997, 25, 3290–3296. [Google Scholar] [CrossRef]
- Juliano, R.L. The delivery of therapeutic oligonucleotides. Nucleic Acids Res. 2016, 44, 6518–6548. [Google Scholar] [CrossRef]
- Rana, D.; Salave, S.; Jain, S.; Shah, R.; Benival, D. Systematic Development and Optimization of Teriparatide-Loaded Nanoliposomes Employing Quality by Design Approach for Osteoporosis. J. Pharm. Innov. 2022, 1–15. [Google Scholar] [CrossRef]
- Salave, S.; Rana, D.; Benival, D. Peptide Functionalised Nanocarriers for Bone Specific Delivery of PTH (1-34) in Osteoporosis. Curr. Nanomed. 2021, 11, 142–148. [Google Scholar] [CrossRef]
- Salave, S.; Rana, D.; Pardhe, R.; Bule, P.; Benival, D. Unravelling Micro and Nano vesicular System in Intranasal Drug Delivery for Epilepsy. Pharm. Nanotechnol. 2022, 10, 182–193. [Google Scholar]
- Salave, S.; Rana, D.; Benival, D. Encapsulation of Anabolic Peptide in Lipid Nano Vesicles for Osteoporosis. Curr. Protein Pept. Sci. 2022, 23, 495–503. [Google Scholar]
- Rawat, G.; Kolhe, S.; Rana, D.; Salave, S.; Benival, D. Exploring the Therapeutic Potential of Cyclosporin for Ophthalmic Indications by virtue of Novel Carrier Systems. Crit. Rev. Trade Ther. Drug Carr. Syst. 2022, 1–45. [Google Scholar] [CrossRef]
- Salave, S.; Rana, D.; Benival, D. Dual Targeting Anti-Osteoporotic Therapy through Potential Nanotherapeutic Approaches. Pharm. Nanotechnol. 2022, 10, 384–392. [Google Scholar]
- Rana, D.; Salave, S.; Longare, S.; Agarwal, R.; Kalia, K.; Benival, D. Nanotherapeutics in Tumour Microenvironment for Cancer Therapy. Nanosci. Nanotechnol.-Asia 2021, 12, e080921196283. [Google Scholar] [CrossRef]
- Lakkaraju, A.; Dubinsky, J.M.; Low, W.C.; Rahman, Y.E. Neurons Are Protected from Excitotoxic Death by p53 Antisense Oligonucleotides Delivered in Anionic Liposomes. J. Biol. Chem. 2001, 276, 32000–32007. [Google Scholar] [CrossRef]
- Noguchi, A.; Furuno, T.; Kawaura, C.; Nakanishi, M. Membrane fusion plays an important role in gene transfection mediated by cationic liposomes. FEBS Lett. 1998, 433, 169–173. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Rusckowski, M.; Liu, N.; Liu, C.; Hnatowich, D.J. Cationic liposomes enhance cellular/nuclear localization of 99mTc-antisense oligonucleotides in target tumor cells. Cancer Biother. Radiopharm. 2001, 16, 411–419. [Google Scholar] [CrossRef]
- Sicard, G.; Giacometti, S.; Rodallec, A.; Ciccolini, J. Enhanced Antisense Oligonucleotide Delivery Using Cationic Liposomes Grafted with Trastuzumab: A Proof-of-Concept Study in Prostate Cancer. Pharmaceutics 2020, 12, 1166. [Google Scholar] [CrossRef]
- Coutinho, G.; Omri, A.; Jerzykiewicz, J.; Czogalla, A. Polyethyleneimine-Based Lipopolyplexes as Carriers in Anticancer Gene Therapies. Materials 2021, 15, 179. [Google Scholar]
- Rezaee, M.; Oskuee, R.K.; Nassirli, H.; Malaekeh-Nikouei, B. Progress in the development of lipopolyplexes as efficient non-viral gene delivery systems. J. Control. Release 2016, 236, 1–14. [Google Scholar] [CrossRef]
- Zhang, Q.Y.; Ho, P.Y.; Tu, M.J.; Jilek, J.L.; Chen, Q.X.; Zeng, S.; Yu, A.M. Lipidation of polyethylenimine-based polyplex increases serum stability of bioengineered RNAi agents and offers more consistent tumoral gene knockdown in vivo. Int. J. Pharm. 2018, 547, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Koh, C.G.; Yu, B.; Liu, S.; Piao, L.; Marcucci, G.; Lee, R.J.; Lee, L.J. Transferrin Receptor Targeted Lipopolyplexes for Delivery of Antisense Oligonucleotide G3139 in a Murine K562 Xenograft Model. Pharm. Res. 2009, 26, 1516. [Google Scholar] [CrossRef] [PubMed]
- Fattal, E.; Couvreur, P.; Dubernet, C. “Smart” delivery of antisense oligonucleotides by anionic pH-sensitive liposomes. Adv. Drug Deliv. Rev. 2004, 56, 931–946. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, C.; Fattal, E.; Couvreur, P.; Lesieur, P. pH-sensitive liposomes as a carrier for oligonucleotides: A physico-chemical study of the interaction between DOPE and a 15-mer oligonucleotide in quasi-anhydrous samples. Biochim. Biophys. Acta 1998, 1372, 301–310. [Google Scholar] [CrossRef]
- Juliano, R.L.; Stamp, D. The effect of particle size and charge on the clearance rates of liposomes and liposome encapsulated drugs. Biochem. Biophys. Res. Commun. 1975, 63, 651–658. [Google Scholar] [CrossRef]
- Ulrich, A.S. Biophysical aspects of using liposomes as delivery vehicles. Biosci. Rep. 2002, 22, 129–150. [Google Scholar] [CrossRef]
- Jones, M.N. The surface properties of phospholipid liposome systems and their characterisation. Adv. Colloid Interface Sci. 1995, 54, 93–128. [Google Scholar] [CrossRef]
- Guan, J.; Guo, H.; Tang, T.; Wang, Y.; Wei, Y.; Seth, P.; Li, Y.; Dehm, S.M.; Ruoslahti, E.; Pang, H.B. iRGD-Liposomes Enhance Tumor Delivery and Therapeutic Efficacy of Antisense Oligonucleotide Drugs against Primary Prostate Cancer and Bone Metastasis. Adv. Funct. Mater. 2021, 31, 2100478. [Google Scholar] [CrossRef]
- Fan, Y.; Marioli, M.; Zhang, K. Analytical characterization of liposomes and other lipid nanoparticles for drug delivery. J. Pharm. Biomed. Anal. 2021, 192, 113642. [Google Scholar] [CrossRef]
- Lo, Y.L.; Liu, Y. Reversing multidrug resistance in Caco-2 by silencing MDR1, MRP1, MRP2, and BCL-2/BCL-xL using liposomal antisense oligonucleotides. PLoS ONE 2014, 9, e90180. [Google Scholar] [CrossRef]
- Galderisi, U.; Cipollaro, M.; Melone, M.A.B.; Iacomino, G.; Di Bernardo, G.; Galano, G.; Contrufo, R.; Zappia, V.; Cascino, A. Myotonic dystrophy: Antisense oligonucleotide inhibition of DMPK gene expression in vitro. Biochem. Biophys. Res. Commun. 1996, 221, 750–754. [Google Scholar] [CrossRef]
- Alamoudi, K.; Martins, P.; Croissant, J.G.; Patil, S.; Omar, H.; Khashab, N.M. Thermoresponsive pegylated bubble liposome nanovectors for efficient siRNA delivery via endosomal escape. Nanomedicine 2017, 12, 1421–1433. [Google Scholar] [CrossRef]
- Koebis, M.; Kiyatake, T.; Yamaura, H.; Nagano, K.; Higashihara, M.; Sonoo, M.; Hayashi, Y.; Negishi, Y.; Endo-Takahashi, Y.; Yanagihara, D.; et al. Ultrasound-enhanced delivery of Morpholino with Bubble liposomes ameliorates the myotonia of myotonic dystrophy model mice. Sci. Rep. 2013, 3, 2242. [Google Scholar] [CrossRef]
- Ulanova, M.; Schreiber, A.D.; Befus, A.D. The future of antisense oligonucleotides in the treatment of respiratory diseases. BioDrugs 2006, 20, 1–11. [Google Scholar] [CrossRef]
- Sachetelli, S.; Khalil, H.; Chen, T.; Beaulac, C.; Sénéchal, S.; Lagacé, J. Demonstration of a fusion mechanism between a fluid bactericidal liposomal formulation and bacterial cells. Biochim. et Biophys. Acta (BBA)—Biomembr. 2000, 1463, 254–266. [Google Scholar] [CrossRef]
- Fillion, P.; Desjardins, A.; Sayasith, K.; Lagacé, J. Encapsulation of DNA in negatively charged liposomes and inhibition of bacterial gene expression with fluid liposome-encapsulated antisense oligonucleotides. Biochim. Biophys. Acta (BBA)-Biomembr. 2001, 1515, 44–54. [Google Scholar] [CrossRef]
- Morishita, R.; Gibbons, G.H.; Ellison, K.E.; Nakajima, M.; Zhang, L.; Kaneda, Y.; Ogihara, T.; Dzau, V.J. Single intraluminal delivery of antisense cdc2 kinase and proliferating-cell nuclear antigen oligonucleotides results in chronic inhibition of neointimal hyperplasia. Proc. Natl. Acad. Sci. USA 1993, 90, 8474–8478. [Google Scholar] [CrossRef]
- Simons, M.; Edelman, E.R.; DeKeyser, J.L.; Langer, R.; Rosenberg, R.D. Antisense c-myb oligonucleotides inhibit intimal arterial smooth muscle cell accumulation in vivo. Nature 1992, 359, 67–70. [Google Scholar] [CrossRef]
- Aoki, M.; Morishita, R.; Higaki, J.; Moriguchi, A.; Kida, I.; Hayashi, S.I.; Matsushita, H.; Kaneda, Y.; Ogihara, T. In vivo transfer efficiency of antisense oligonucleotides into the myocardium using HVJ–liposome method. Biochem. Biophys. Res. Commun. 1997, 231, 540–545. [Google Scholar] [CrossRef]
- Zhang, Y.; Rong Qi, X.; Gao, Y.; Wei, L.; Maitani, Y.; Nagai, T. Mechanisms of co-modified liver-targeting liposomes as gene delivery carriers based on cellular uptake and antigens inhibition effect. J. Control. Release 2007, 117, 281–290. [Google Scholar] [CrossRef]
- Arranz, A.; Reinsch, C.; Papadakis, K.A.; Dieckmann, A.; Rauchhaus, U.; Androulidaki, A.; Zacharioudaki, V.; Margioris, A.N.; Tsatsanis, C.; Panzner, S. Treatment of experimental murine colitis with CD40 antisense oligonucleotides delivered in amphoteric liposomes. J. Control. Release 2013, 165, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Bochot, A.; Fattal, E.; Boutet, V.; Deverre, J.R.; Jeanny, J.C.; Chacun, H.; Couvreur, P. Intravitreal Delivery of Oligonucleotides by Sterically Stabilized Liposomes. Investig. Ophthalmol. Vis. Sci. 2002, 43, 253–259. [Google Scholar]
- Hélène, C.; Toulmé, J.J. Specific regulation of gene expression by antisense, sense and antigene nucleic acids. Biochim. Biophys. Acta 1990, 1049, 99–125. [Google Scholar] [CrossRef]
- Cantin, E.M.; Woolf, T.M. Antisense oligonucleotides as antiviral agents: Prospects and problems. Trends Microbiol. 1993, 1, 270–276. [Google Scholar] [CrossRef]
- Ropert, C.; Malvy, C.; Couvreur, P. Inhibition of the Friend Retrovirus by Antisense Oligonucleotides Encapsulated in Liposomes: Mechanism of Action. Pharm. Res. Off. J. Am. Assoc. Pharm. Sci. 1993, 10, 1427–1433. [Google Scholar]
- DENG, X.; CAI, X.; KE, Z. Preliminary studies of prevention of interleukin-6 antisense oligonucleotide liposomes on after cataract in rabbit. J. Med. Postgrad. 2003, 12, wpr-585555. [Google Scholar]
- Xue, X.Y.; Mao, X.G.; Zhou, Y.; Chen, Z.; Hu, Y.; Hou, Z.; Li, M.K.; Meng, J.R.; Luo, X.X. Advances in the delivery of antisense oligonucleotides for combating bacterial infectious diseases. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 745–758. [Google Scholar] [CrossRef]
- Das, A.R.; Dattagupta, N.; Sridhar, C.N.; Wu, W.K. A novel thiocationic liposomal formulation of antisense oligonucleotides with activity against mycobacterium tuberculosis. Scand. J. Infect. Dis. 2003, 35, 168–174. [Google Scholar] [CrossRef]
- Sartorius, C.; Franklin, R.M. The use of antisense oligonucleotides as chemotherapeutic agents for parasites. Parasitol. Today 1991, 7, 90–93. [Google Scholar] [CrossRef]
- Siwak, D.R.; Tari, A.M.; Lopez-Berestein, G. Liposomal antisense oligonucleotides for cancer therapy. Methods Enzymol. 2004, 387, 241–253. [Google Scholar]
- Rodríguez, M.; Coma, S.; Noé, V.; Ciudad, C.J. Development and effects of immunoliposomes carrying an antisense oligonucleotide against DHFR RNA and directed toward human breast cancer cells overexpressing HER2. Antisense Nucleic Acid Drug Dev. 2002, 12, 311–325. [Google Scholar] [CrossRef]
- Pakunlu, R.I.; Wang, Y.; Saad, M.; Khandare, J.J.; Starovoytov, V.; Minko, T. In vitro and in vivo intracellular liposomal delivery of antisense oligonucleotides and anticancer drug. J. Control. Release 2006, 114, 153–162. [Google Scholar] [CrossRef]
- Yu, B.; Mao, Y.; Bai, L.Y.; Herman, S.E.M.; Wang, X.; Ramanunni, A.; Jin, Y.; Mo, X.; Cheney, C.; Chan, K.K.; et al. Targeted nanoparticle delivery overcomes off-target immunostimulatory effects of oligonucleotides and improves therapeutic efficacy in chronic lymphocytic leukemia. Blood 2013, 121, 136–147. [Google Scholar] [CrossRef]
- Matthay, K.K.; Maris, J.M.; Schleiermacher, G.; Nakagawara, A.; Mackall, C.L.; Diller, L.; Weiss, W.A. Neuroblastoma. Nat. Rev. Dis. Prim. 2016, 2, 1–21. [Google Scholar] [CrossRef]
- Brignole, C.; Pagnan, G.; Marimpietri, D.; Cosimo, E.; Allen, T.M.; Ponzoni, M.; Pastorino, F. Targeted delivery system for antisense oligonucleotides: A novel experimental strategy for neuroblastoma treatment. Cancer Lett. 2003, 197, 231–235. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, L.; Liu, Y.H.; Wilkinson, J.E.; Krainer, A.R. Antisense oligonucleotide therapy for H3.3K27M diffuse midline glioma. Sci. Transl. Med. 2023, 15, eadd8280. [Google Scholar] [CrossRef]
- Schlingensiepen, K.H.; Schlingensiepen, R.; Steinbrecher, A.; Hau, P.; Bogdahn, U.; Fischer-Blass, B.; Jachimczak, P. Targeted tumor therapy with the TGF-β2 antisense compound AP 12009. Cytokine Growth Factor Rev. 2006, 17, 129–139. [Google Scholar] [CrossRef]
- Min, H.S.; Kim, H.J.; Naito, M.; Ogura, S.; Toh, K.; Hayashi, K.; Kim, B.S.; Fukushima, S.; Anraku, Y.; Miyata, K.; et al. Systemic Brain Delivery of Antisense Oligonucleotides across the Blood–Brain Barrier with a Glucose-Coated Polymeric Nanocarrier. Angew. Chem. Int. Ed. 2020, 59, 8173–8180. [Google Scholar] [CrossRef]
- Bochot, A.; Mashhour, B.; Puisieux, F.; Couvreur, P.; Fattal, E. Comparison of the ocular distribution of a model oligonucleotide after topical instillation in rabbits of conventional and new dosage forms. J. Drug Target. 1998, 6, 309–313. [Google Scholar] [CrossRef]
- Saffari, M.; Moghimi, H.R.; Dass, C.R. Barriers to Liposomal Gene Delivery: From Application Site to the Target. Iran. J. Pharm. Res. IJPR 2016, 15, 3. [Google Scholar]
- Pastorino, F.; Stuart, D.; Ponzoni, M.; Allen, T.M. Targeted delivery of antisense oligonucleotides in cancer. J. Control. Release 2001, 74, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Lysik, M.A.; Wu-Pong, S. Innovations in Oligonucleotide Drug Delivery. J. Pharm. Sci. 2003, 92, 1559–1573. [Google Scholar] [CrossRef] [PubMed]
- Ponnappa, B.C.; Israel, Y.; Aini, M.; Zhou, F.; Russ, R.; Cao, Q.N.; Hu, Y.; Rubin, R. Inhibition of tumor necrosis factor alpha secretion and prevention of liver injury in ethanol-fed rats by antisense oligonucleotides. Biochem. Pharmacol. 2005, 69, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Inglut, C.T.; Sorrin, A.J.; Kuruppu, T.; Vig, S.; Cicalo, J.; Ahmad, H.; Huang, H.C. Immunological and Toxicological Considerations for the Design of Liposomes. Nanomaterials 2020, 10, 190. [Google Scholar] [CrossRef]
- Kedmi, R.; Ben-Arie, N.; Peer, D. The systemic toxicity of positively charged lipid nanoparticles and the role of Toll-like receptor 4 in immune activation. Biomaterials 2010, 31, 6867–6875. [Google Scholar] [CrossRef]
- Berdugo, M.; Valamanesh, F.; Andrieu, C.; Klein, C.; Benezra, D.; Courtois, Y.; Behar-Cohen, F. Delivery of antisense oligonucleotide to the cornea by iontophoresis. Antisense Nucleic Acid Drug Dev. 2003, 13, 107–114. [Google Scholar] [CrossRef]





| Sr. No. | Disease | ASO | Lipid Composition | Particle Size | Route of Administration | Reference |
|---|---|---|---|---|---|---|
| 1. | Chronic lymphocytic leuakemia | Phosphorothioate-modified oligos G3139 (5′-TCT CCC AGC GTG CGC CAT-3′), G3622 (5′-TAC CGC GTG CGA CCC TCT-3′), and a fluorescein-modified ODN (5′-(6)-FAM-TAC CGC GTG CGA CCC TCT-3′) | 3β-N-(N′, N′-dimethyl amino ethane)-carbamoyl cholesterol/EPC/methoxy polyethylene glycol–distearoyl phosphatidylethanolamine (28/70/2) | 56.3 ± 7.5 nm | Intraperitoneal | [105] |
| 2. | Myotonic dystrophy | Phosphorodiamidate morpholino oligonucleotide (PMO) | DPPC:1,2-distearoyl-sn-glycero-3-phosphatidyl-ethanolamine-polyethyleneglycol (DSPE-PEG2000-OMe) (94:6) | <200 nm | Intramuscular | [85] |
| 3. | Bacterial Infections | Anti-β-galactosidase antisense oligonucleotide (5′- GGT CAT AGC TGT TTC-3′) | DPPC: DMPG (10:1) | 316.2–562.3 nm | - | [88] |
| 4. | Neointimal hyperplasia | Antisense cdc2 kinase [5′-GTCTTCCATAGTTACTCA-3′] | Phosphatidylserine:Phosphatidylcholine:Cholesterol (1:4:8:2) | - | - | [89] |
| 5. | Cancer | 5′-CAG CGT GCG CCA TCC TTC CC-3′ and 5′-TTC AAG ATC CAT CCC GAC CTC GCG-3′ ASO | EPC/DPPC/Cholesterol (7:3:10) | 100–200 nm | Subcutaneous | [104] |
| 6. | Viral infections | 5′-TGAACACGCCATGTC-3′ ASO | DOPE/Oleic acid/Cholesterol (10:5:2) | 170 nm | - | [97] |
| 7. | Ocular diseases | 16-mer oligothymidylate (pdT16) | PC:Cholesterol:PEG-DSPE (64:30:6) | 150 nm | Intravitreal | [111] |
| 8. | Inflammatory Bowel Disease | Phosphorothioate and 2′methoxyethyl (MOE) modified ASO | POPC:DOPE:CHEMS:MoChol (15:45:20:20) | 162 nm | Intravenous | [93] |
| 9. | Cardiac diseases | FITC-labelled phosphorothioate ODN | Phosphatidylserine:Phosphatidylcholine:Cholesterol (1:4.8:2) | - | - | [91] |
| 10. | Cardiac arrythmia | Anti-miR-1 ASOs (AMO-1) | EPC: CHO:DSPE-PEG2000 (49:50:1) | 105 ± 0.5 | Intravenous | [43] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gupta, R.; Salave, S.; Rana, D.; Karunakaran, B.; Butreddy, A.; Benival, D.; Kommineni, N. Versatility of Liposomes for Antisense Oligonucleotide Delivery: A Special Focus on Various Therapeutic Areas. Pharmaceutics 2023, 15, 1435. https://doi.org/10.3390/pharmaceutics15051435
Gupta R, Salave S, Rana D, Karunakaran B, Butreddy A, Benival D, Kommineni N. Versatility of Liposomes for Antisense Oligonucleotide Delivery: A Special Focus on Various Therapeutic Areas. Pharmaceutics. 2023; 15(5):1435. https://doi.org/10.3390/pharmaceutics15051435
Chicago/Turabian StyleGupta, Raghav, Sagar Salave, Dhwani Rana, Bharathi Karunakaran, Arun Butreddy, Derajram Benival, and Nagavendra Kommineni. 2023. "Versatility of Liposomes for Antisense Oligonucleotide Delivery: A Special Focus on Various Therapeutic Areas" Pharmaceutics 15, no. 5: 1435. https://doi.org/10.3390/pharmaceutics15051435