
Urtica dioica Agglutinin Prevents Rabies Virus Infection in a Muscle Explant Model
 , ,
, ,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Virus, and Lectins
2.2. Antiviral Assay
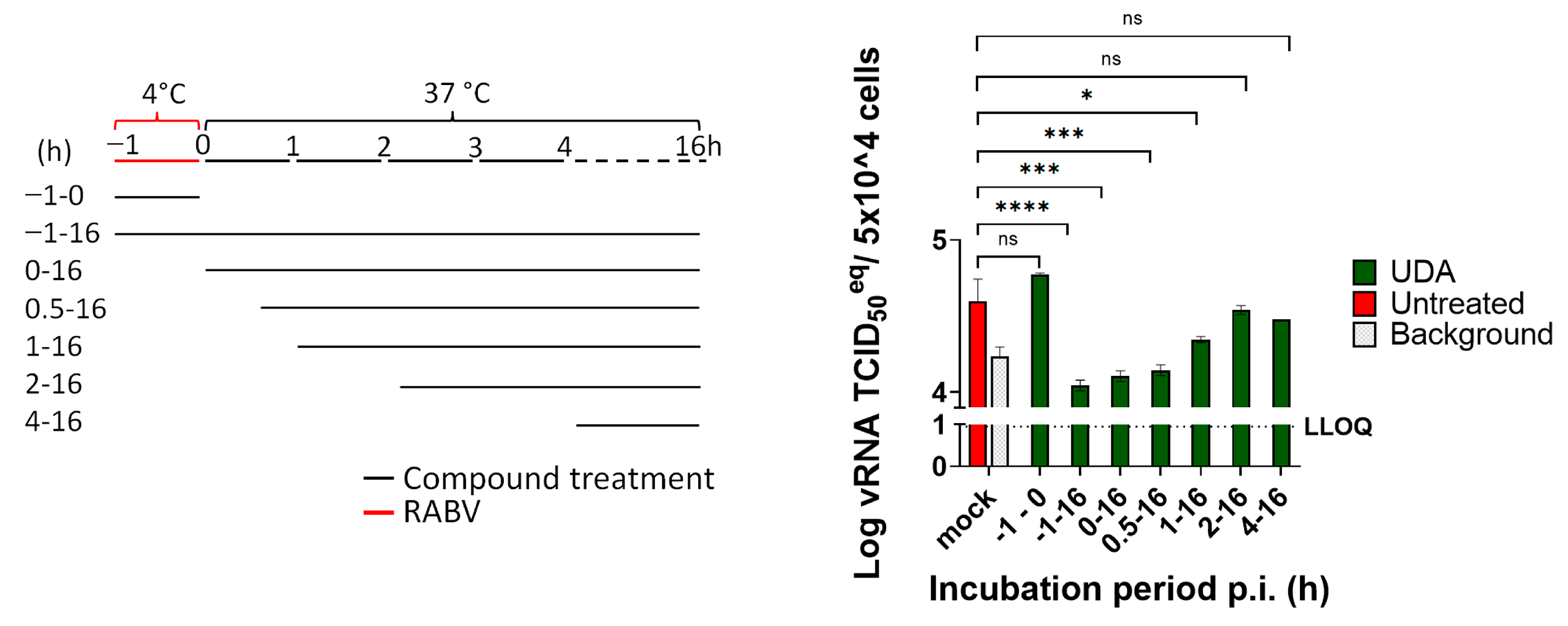
2.3. Time of Drug Addition Assay
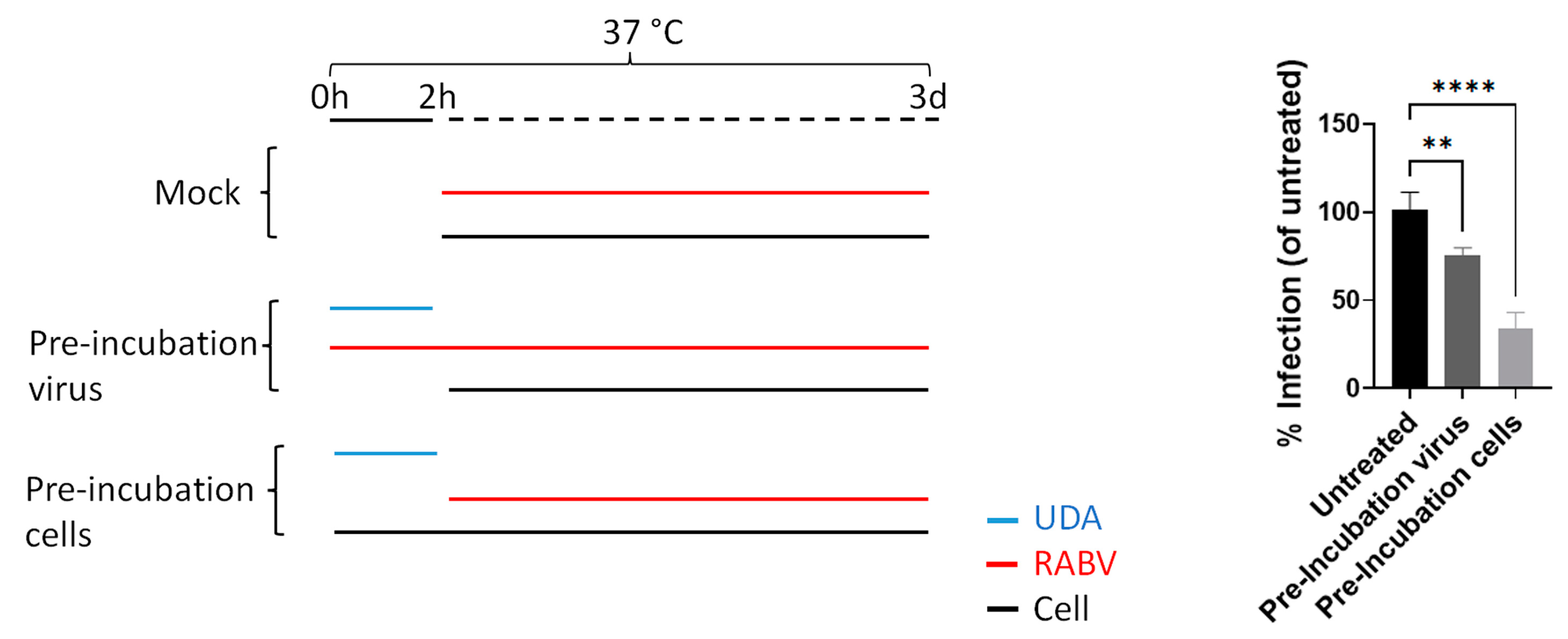
2.4. Pre-Incubation of RABV to Lectins Prior to Infection
2.5. Pre-Incubation of BHK to Lectins Prior to Infection
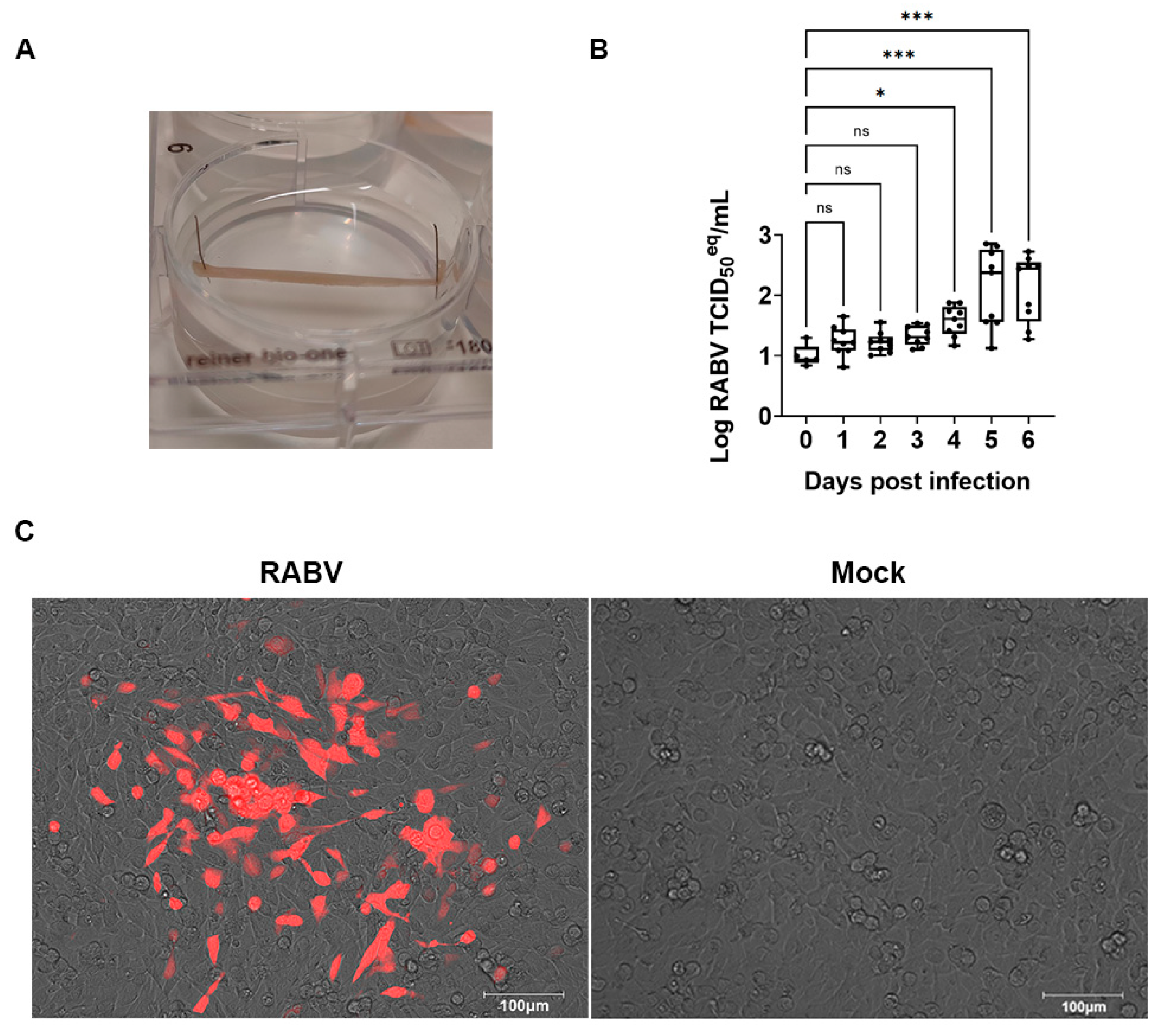
2.6. Swine Skeletal Muscle Explant Culture and Antiviral Assay
2.7. Muscle Culture Supernatant Titration
2.8. Muscle Explant Viability Assay
2.9. Fluorescent Immunostaining and Microscopy of Muscle
2.10. Real-Time RT-qPCR
2.11. Statistics
3. Results
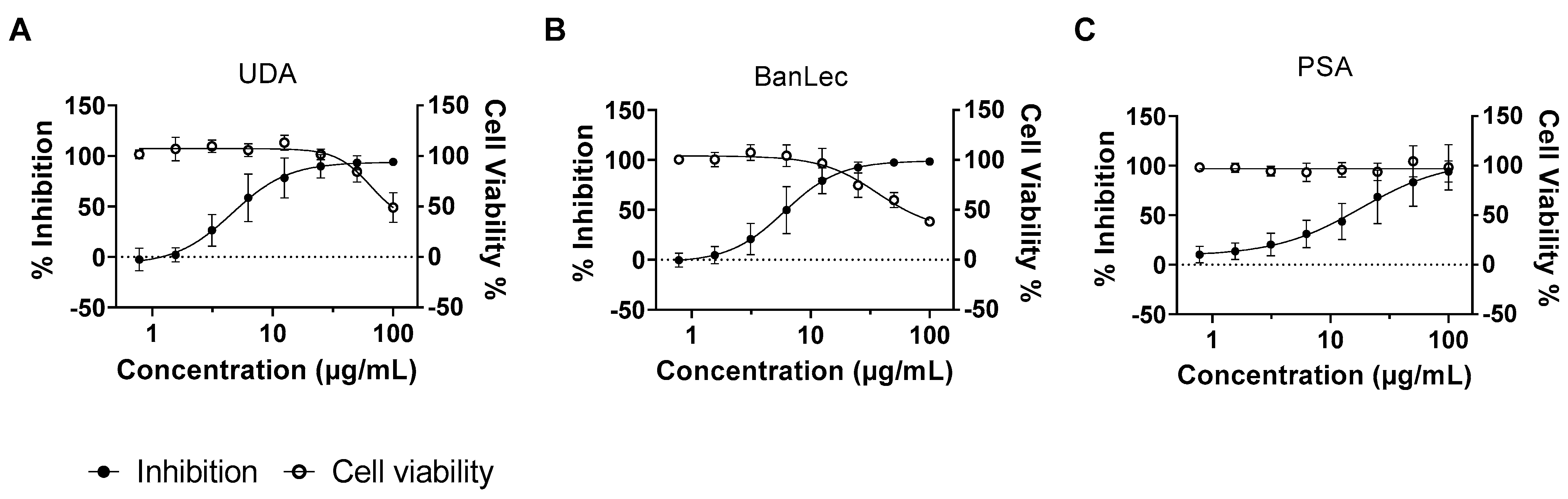
3.1. Several Lectins Inhibit RABV in Cell Culture
3.2. RABV Replicates in Swine Skeletal Muscle Explants
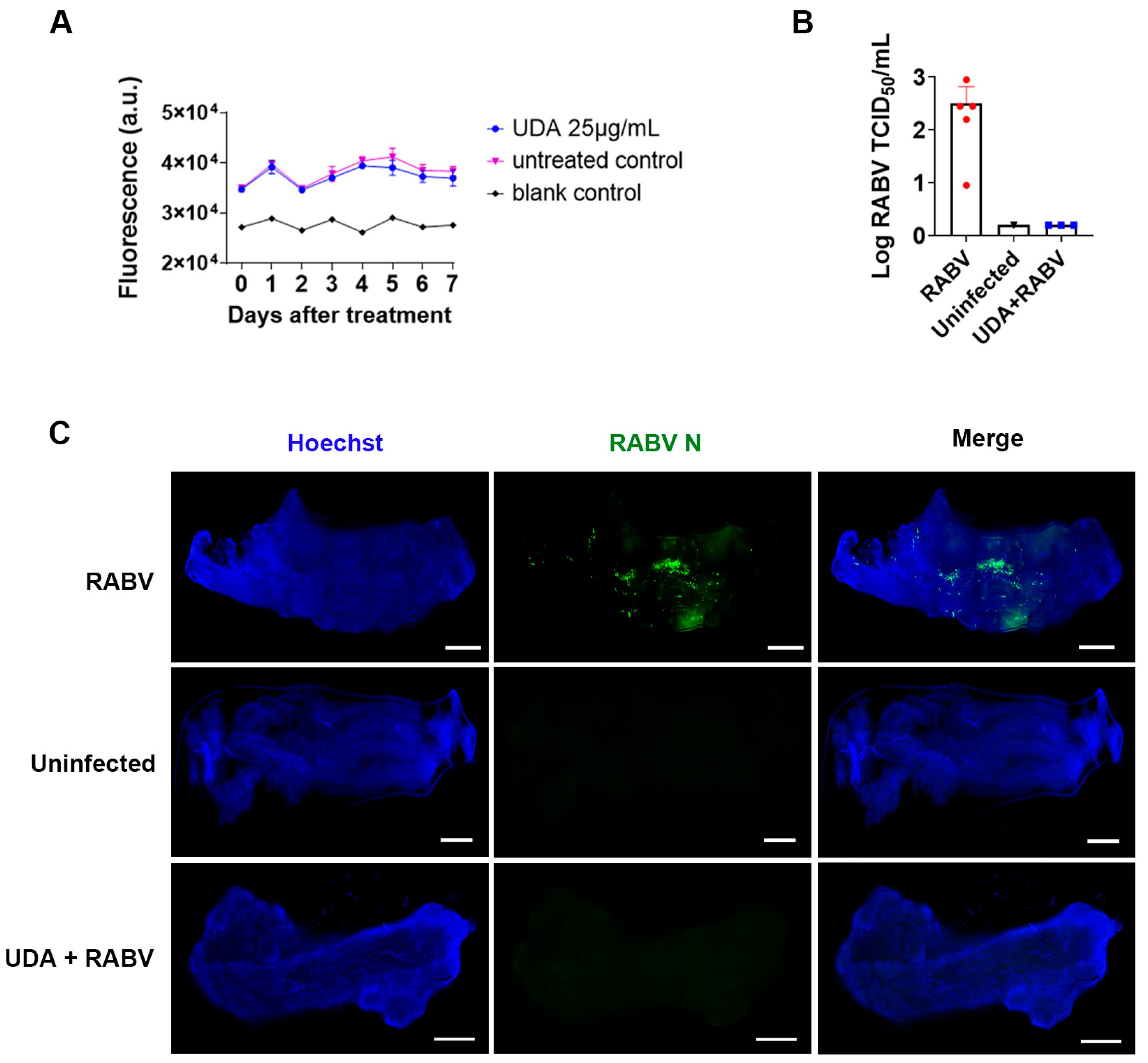
3.3. UDA Inhibits RABV Infection in Swine Skeletal Muscle
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fooks, A.R.; Cliquet, F.; Finke, S.; Freuling, C.; Hemachudha, T.; Mani, R.S.; Müller, T.; Nadin-Davis, S.; Picard-Meyer, E.; Wilde, H.; et al. Rabies. Nat. Rev. Dis. Prim. 2017, 3, 17091. [Google Scholar] [CrossRef] [PubMed]
- WHO. WHO Guide for Rabies Pre and Post-Exposure Prophylaxis in Humans; World Health Organization: Geneva, Switzerland, 2014; pp. 1–21.
- Lankester, F.; Hampson, K.; Lembo, T.; Palmer, G.; Taylor, L.; Cleaveland, S. Implementing Pasteur’s vision for rabies elimination. Science 2014, 345, 1562–1564. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasan, N.; Li, A.; Shiferaw, M.; Tran, C.H.; Wallace, R.; Blanton, J.; Knopf, L.; Abela-Ridder, B.; Hyde, T.; Siddiqi, U.R.; et al. Overview of rabies post-exposure prophylaxis access, procurement and distribution in selected countries in Asia and Africa, 2017–2018. Vaccine 2019, 37, A6–A13. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhao, W.; Zhang, S.F.; Miao, F.M.; Cao, Y.; Chen, C.; Li, Y.F.; Gao, J.; Lv, R.Y.; Zhang, S.X.; et al. Efficacy of ormutivimab, a novel recombinant human anti-rabies monoclonal antibody, in post-exposure prophylaxis animal models. Travel Med. Infect. Dis. 2022, 46, 102267. [Google Scholar] [CrossRef]
- WHO. Quality Assurance of Pharmaceuticals: A Compendium of Guidelines and Related Materials. Good Manufacturing Practices and Inspection, 2nd ed.; WHO: Geneva, Switzerland, 2007; Volume 2.
- Hemachudha, T.; Ugolini, G.; Wacharapluesadee, S.; Sungkarat, W.; Shuangshoti, S.; Laothamatas, J. Human rabies: Neuropathogenesis, diagnosis, and management. Lancet Neurol. 2013, 12, 498–513. [Google Scholar] [CrossRef]
- Hanham, C.A.; Zhao, F.; Tignor, G.H. Evidence from the anti-idiotypic network that the acetylcholine receptor is a rabies virus receptor. J. Virol. 1993, 67, 530–542. [Google Scholar] [CrossRef]
- Lentz, T.L. Rabies virus binding to an acetylcholine receptor alpha-subunit peptide. J. Mol. Recognit. 1990, 3, 82–88. [Google Scholar] [CrossRef]
- Lentz, T.L.; Burrage, T.G.; Smith, A.L.; Crick, J.; Tignor, G.H. Is the acetylcholine receptor a rabies virus receptor? Science 1982, 215, 182–184. [Google Scholar] [CrossRef]
- Lentz, T.L.; Burrage, T.G.; Smith, A.L.; Tignor, G.H. The acetylcholine receptor as a cellular receptor for rabies virus. Yale J. Biol. Med. 1983, 56, 315–322. [Google Scholar]
- Lafon, M. Rabies virus receptors. J. Neurovirol. 2005, 11, 82–87. [Google Scholar] [CrossRef]
- Sharon, N.; Lis, H. Lectins as cell recognition molecules. Science 1989, 246, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Lis, H.; Sharon, N. Lectins: Carbohydrate-Specific Proteins That Mediate Cellular Recognition. Chem. Rev. 1998, 98, 637–674. [Google Scholar] [CrossRef] [PubMed]
- Balzarini, J. Inhibition of HIV entry by carbohydrate-binding proteins. Antivir. Res. 2006, 71, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.K.; Ng, T.B. Lectins: Production and practical applications. Appl. Microbiol. Biotechnol. 2011, 89, 45–55. [Google Scholar] [CrossRef]
- Akkouh, O.; Ng, T.B.; Singh, S.S.; Yin, C.; Dan, X.; Chan, Y.S.; Pan, W.; Cheung, R.C.F. Lectins with Anti-HIV Activity: A Review. Molecules 2015, 20, 648–668. [Google Scholar] [CrossRef]
- Keyaerts, E.; Vijgen, L.; Pannecouque, C.; Van Damme, E.; Peumans, W.; Egberink, H.; Balzarini, J.; Van Ranst, M. Plant lectins are potent inhibitors of coronaviruses by interfering with two targets in the viral replication cycle. Antivir. Res. 2007, 75, 179–187. [Google Scholar] [CrossRef]
- Gordts, S.C.; Renders, M.; Férir, G.; Huskens, D.; Van Damme, E.J.M.; Peumans, W.; Balzarini, J.; Schols, D. NICTABA and UDA, two GlcNAc-binding lectins with unique antiviral activity profiles. J. Antimicrob. Chemother. 2014, 70, 1674–1685. [Google Scholar] [CrossRef]
- Vanhulle, E.; D’huys, T.; Provinciael, B.; Stroobants, J.; Camps, A.; Noppen, S.; Schols, D.; Van Damme, E.J.M.; Maes, P.; Stevaert, A.; et al. Carbohydrate-binding protein from stinging nettle as fusion inhibitor for SARS-CoV-2 variants of concern. Front. Cell. Infect. Microbiol. 2022, 12, 1267. [Google Scholar] [CrossRef]
- Vanderlinden, E.; Van Winkel, N.; Naesens, L.; Van Damme, E.J.M.; Persoons, L.; Schols, D. In Vitro Characterization of the Carbohydrate-Binding Agents HHA, GNA, and UDA as Inhibitors of Influenza A and B Virus Replication. Antimicrob. Agents Chemother. 2021, 65, e01732-20. [Google Scholar] [CrossRef]
- Van Damme, E.J.M.; Peumans, W.J.; Pusztai, A.; Bardocz, S. Handbook of Plant Lectins: Properties and Biomedical Applications; John Wiley & Sons: Hoboken, NJ, USA, 1998; ISBN 047196445X. [Google Scholar]
- Jochmans, D.; Leyssen, P.; Neyts, J. A novel method for high-throughput screening to quantify antiviral activity against viruses that induce limited CPE. J. Virol. Methods 2012, 183, 176–179. [Google Scholar] [CrossRef]
- WHO. Rabies vaccines: WHO position paper—April 2018. Wkly. Epidemiol. Rec. 2018, 93, 201–219. [Google Scholar]
- De Melo, G.D.; Hellert, J.; Gupta, R.; Corti, D.; Bourhy, H. Monoclonal antibodies against rabies: Current uses in prophylaxis and in therapy. Curr. Opin. Virol. 2022, 53, 101204. [Google Scholar] [CrossRef] [PubMed]
- Conti, C.; Tsiang, H. Effect of Concanavalin A on the Early Events of Rabies Virus Infection of CER Cells. Intervirology 1985, 24, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.C.M.; Malveira, E.A.; Mendonça, L.P.; Maia, M.E.S.; Silva, R.R.S.; Roma, R.R.; Aguiar, T.K.B.; Grangeiro, Y.A.; Souza, P.F.N. Plant Lectins: A Review on their Biotechnological Potential Toward Human Pathogens. Curr. Protein Pept. Sci. 2022, 23, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Meiers, J.; Siebs, E.; Zahorska, E.; Titz, A. Lectin antagonists in infection, immunity, and inflammation. Curr. Opin. Chem. Biol. 2019, 53, 51–67. [Google Scholar] [CrossRef]
- Hansen, J.E.; Nielsen, C.M.; Nielsen, C.; Heegaard, P.; Mathiesen, L.R.; Nielsen, J.O. Correlation between carbohydrate structures on the envelope glycoprotein gp120 of HIV-1 and HIV-2 and syncytium inhibition with lectins. AIDS 1989, 3, 635–641. [Google Scholar] [CrossRef]
- Seganti, L.; Superti, F.; Sinibaldi, L.; Marchetti, M.; Bianchi, S.; Orsi, N. Rabies virus infection in Aedes pseudoscutellaris cells: A study on receptorial structures. Comp. Immunol. Microbiol. Infect. Dis. 1991, 14, 265–275. [Google Scholar] [CrossRef]
- Saul, F.A.; Rovira, P.; Boulot, G.; Van Damme, E.J.; Peumans, W.J.; Truffa-Bachi, P.; Bentley, G.A. Crystal structure of Urtica dioica agglutinin, a superantigen presented by MHC molecules of class I and class II. Structure 2000, 8, 593–603. [Google Scholar] [CrossRef]
- Grauso, L.; de Falco, B.; Lanzotti, V.; Motti, R. Stinging nettle, Urtica dioica L.: Botanical, phytochemical and pharmacological overview. Phytochem. Rev. 2020, 19, 1341–1377. [Google Scholar] [CrossRef]
- Badirzadeh, A.; Heidari-Kharaji, M.; Fallah-Omrani, V.; Dabiri, H.; Araghi, A.; Chirani, A.S. Antileishmanial activity of Urtica dioica extract against zoonotic cutaneous leishmaniasis. PLoS Negl. Trop. Dis. 2020, 14, e0007843. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lectin | Species | 1EC50 (μg/mL) | CC50 (μg/mL) | SI |
|---|---|---|---|---|
| Gal/GalNAc-specific agglutinins | ||||
| PHA-L4 | Phaseolus vulgaris | 47 ± 13 | >100 | >2.1 |
| RPA | Robinia pseudoacacia | 5.4 ± 2.5 | 11 ± 1.4 | 2.1 |
| RSA | Rhizoctonia solani | >100 | >100 | Na |
| SJA | Styphnolobium japonicum | 32 ± 19 | >100 | >3.2 |
| GalNAc-specific agglutinins | ||||
| BPA | Bauhinia purpurea | 3.2 ± 1.0 | 9.0 ± 2.1 | 2.8 |
| CAA | Caragana arborescens | 95 ± 5 | >100 | >1.0 |
| DBA | Dolichos biflorus | >100 | >100 | Na |
| SBA | Soybean | >100 | >100 | Na |
| WFA | Wisteria floribunda | >100 | >100 | Na |
| Gal-specific agglutinins | ||||
| Jacalin | Jackfruit | >100 | >100 | Na |
| Morniga G | Morus nigra | >100 | 24 ± 8.6 | <0.2 |
| PHA-E | Phaseolus vulgaris | 47 ± 1.5 | 86 ± 18 | 1.8 |
| PNA | Arachis hypogaea | >100 | >100 | Na |
| GlcNAc-specific agglutinins | ||||
| DSL | Datura stramonium | 57 ± 2.2 | 69 ± 12.4 | 1.2 |
| GS II | Griffonia simplicifolia | >100 | >100 | Na |
| Nictaba | Nicotiana tabacum | 29.0 ± 3.9 | >100 | >3.4 |
| UDA | Urtica dioica | 8.1 ± 1.3 | 54 ± 6.8 | 6.7 |
| UEA II | Ulex europaeus | >100 | >100 | Na |
| WGA | Wheat germ | 21 ± 7.0 | 17 ± 6.3 | 0.8 |
| Man/GalNAc-specific agglutinins | ||||
| AMA | Arum maculatum | 7.1 ± 2.9 | 9.1 ± 2.2 | 1.3 |
| TLC I | Tulipa hybrid | 32 ± 5.7 | >100 | >3.2 |
| Mannose-specific agglutinins | ||||
| APA | Allium porrum L. | 36 ± 11 | >100 | >2.9 |
| Banana lectin | Musa acuminata | 6.2 ± 2.6 | 40.7 ± 11.5 | 6.5 |
| Calsepa | Calystegia sepium | >100 | >100 | Na |
| Con A | Canavalia ensiformis | 13 ± 3.7 | 22 ± 4 | 1.7 |
| Conarv A | Convolvulus arvensis | 90.7 ± 9.3 | >100 | >1.1 |
| GNA | Galanthus nivalis | >100 | >100 | Na |
| HHA | Hippeastrum hybrid | 29 ± 2.3 | >100 | >3.4 |
| Morniga M | Morus nigra | >100 | 27.4 ± 14.1 | <0.3 |
| NPA | Narcissus pseudonarcissus | 16 ± 5.4 | 42 ± 5.8 | 2.6 |
| PSA | Pisum sativum | 13 ± 7.5 | >100 | >7.7 |
| Sialic acid-specific agglutinin | ||||
| ACA | Amaranthus caudatus | >100 | >100 | Na |
| MAA | Maackia amurensis | >100 | >100 | Na |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Terrie, L.; Wu, G.; Van Damme, E.J.M.; Thorrez, L.; Fooks, A.R.; Banyard, A.C.; Jochmans, D.; Neyts, J. Urtica dioica Agglutinin Prevents Rabies Virus Infection in a Muscle Explant Model. Pharmaceutics 2023, 15, 1353. https://doi.org/10.3390/pharmaceutics15051353
Wang X, Terrie L, Wu G, Van Damme EJM, Thorrez L, Fooks AR, Banyard AC, Jochmans D, Neyts J. Urtica dioica Agglutinin Prevents Rabies Virus Infection in a Muscle Explant Model. Pharmaceutics. 2023; 15(5):1353. https://doi.org/10.3390/pharmaceutics15051353
Chicago/Turabian StyleWang, Xinyu, Lisanne Terrie, Guanghui Wu, Els J. M. Van Damme, Lieven Thorrez, Anthony R. Fooks, Ashley C. Banyard, Dirk Jochmans, and Johan Neyts. 2023. "Urtica dioica Agglutinin Prevents Rabies Virus Infection in a Muscle Explant Model" Pharmaceutics 15, no. 5: 1353. https://doi.org/10.3390/pharmaceutics15051353