
Saponin Fraction CIL1 from Lysimachia ciliata L. Enhances the Effect of a Targeted Toxin on Cancer Cells
 , ,
, ,  , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. The CIL1 Saponin Fraction
2.2. Expression and Purification of DE
2.3. Determination of Enzymatic Activity
2.4. NIH-3T3 and HER14 Cell Culture
2.5. Experimental Procedure and Viability Assay
2.6. Proliferation Assay
2.7. Apoptosis
2.7.1. Flow Cytometry Analysis
2.7.2. Luminescence Detection of Caspase 3/7
2.8. Safety Study
2.8.1. Hepatotoxicity, Neurotoxicity and Cardiotoxicity
2.8.2. Mutagenic Potential
3. Results
3.1. CIL1 Increases DE-Mediated Inhibition of Proliferation and Cytotoxicity of HER14 and NIH3T3 Cells
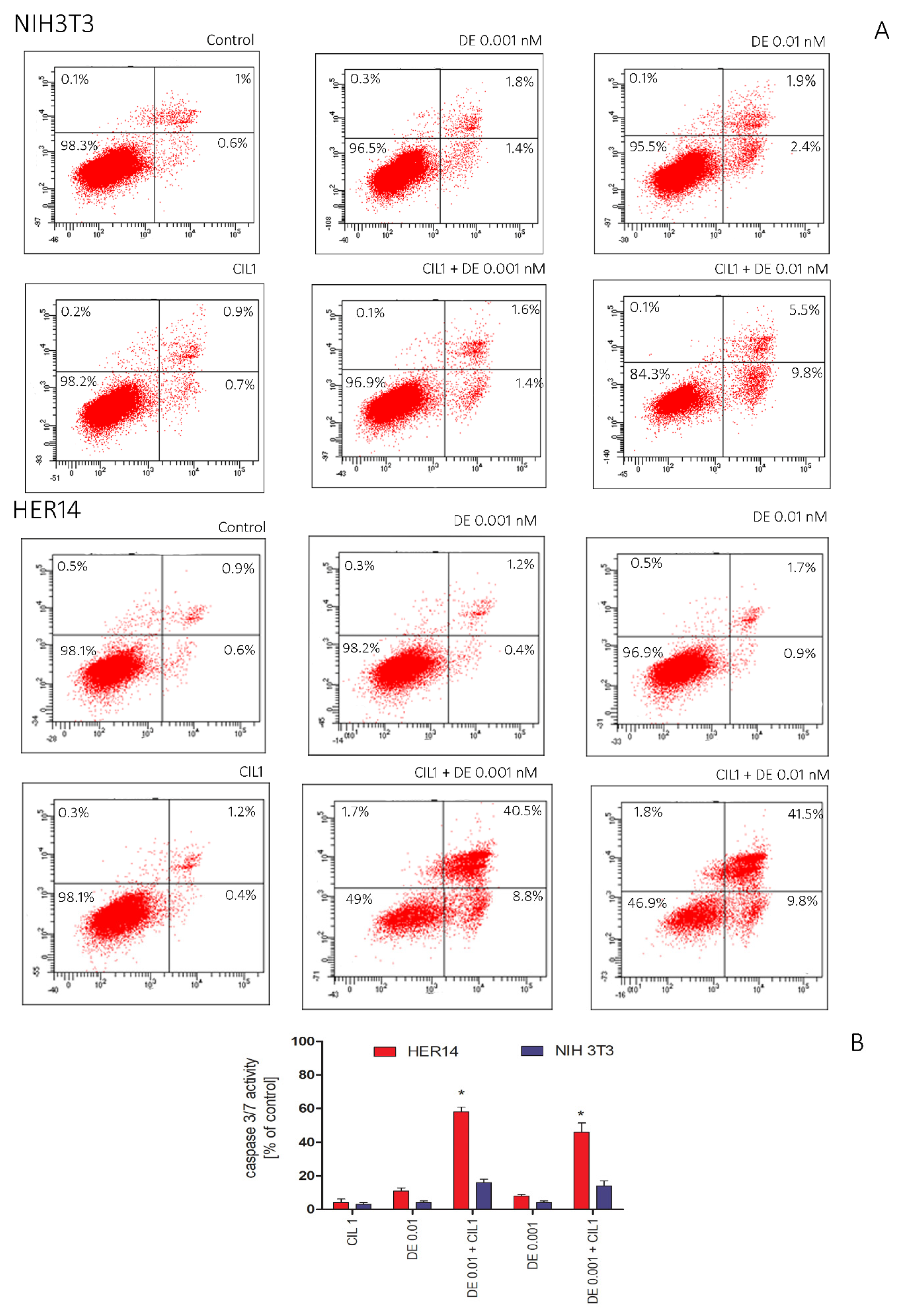
3.2. CIL1 Saponin Exhibits a Synergistic Effect with DE and Enhances Apoptosis of HER14 and NIH3T3 Cells
3.3. CIL1 Revealed a Satisfactory Safety Profile Confirmed by an In Vitro Cytotoxicity Analysis and a Mutagenicity Assay
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Magedans, Y.V.; Yendo, A.C.; Costa, F.; Gosmann, G.; Fett-Neto, A.G. Foamy matters: An update on Quillaja saponins and their use as immunoadjuvants. Future Med. Chem. 2019, 11, 1485–1499. [Google Scholar] [CrossRef] [PubMed]
- Wang, P. Natural and Synthetic Saponins as Vaccine Adjuvants. Vaccines 2021, 9, 222. [Google Scholar] [CrossRef] [PubMed]
- Koczurkiewicz, P.; Kowolik, E.; Podolak, I.; Wnuk, D.; Piska, K.; Łabędź-Masłowska, A.; Wójcik-Pszczoła, K.; Pękala, E.; Czyż, J.; Michalik, M. Synergistic Cytotoxic and Anti-invasive Effects of Mitoxantrone and Triterpene Saponins from Lysimachia ciliata on Human Prostate Cancer Cells. Planta Med. 2016, 82, 1546–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Min, W.; Lin, S.; Song, L.; Yang, P.; Ma, Q.; Guo, J. Saikosaponin-d increases radiation-induced apoptosis of hepatoma cells by promoting autophagy via inhibiting mTOR phosphorylation. Int. J. Med. Sci. 2021, 18, 1465–1473. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhan, Y.; Chen, B.; Lu, Y.; Yin, T.; Zhou, S.; Zhang, W.; Liu, X.; Du, B.; Wei, X.; et al. Tubeimoside I-induced lung cancer cell death and the underlying crosstalk between lysosomes and mitochondria. Cell Death Dis. 2020, 11, 708. [Google Scholar] [CrossRef]
- Zahaf, N.I.; Schmidt, G. Bacterial Toxins for Cancer Therapy. Toxins 2017, 9, 236. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.K.; Brock, D.J.; Kondow-McConaghy, H.M.; Pellois, J.P. Efficient Delivery of Macromolecules into Human Cells by Improving the Endosomal Escape Activity of Cell-Penetrating Peptides: Lessons Learned from dfTAT and its Analogs. Biomolecules 2018, 8, 50. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, H.; Niesler, N.; Trautner, A.; Sama, S.; Jerz, G.; Panjideh, H.; Weng, A. Glycosylated Triterpenoids as Endosomal Escape Enhancers in Targeted Tumor Therapies. Biomedicines 2017, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, H.; Bachran, C. Targeted tumor therapies at a glance. Curr. Drug Targets 2009, 10, 89–93. [Google Scholar] [CrossRef]
- Bachran, C.; Sutherland, M.; Heisler, I.; Hebestreit, P.; Melzig, M.F.; Fuchs, H. The saponin-mediated enhanced uptake of targeted saporin-based drugs is strongly dependent on the saponin structure. Exp. Biol. Med. 2006, 231, 412–420. [Google Scholar] [CrossRef]
- Podolak, I.; Koczurkiewicz, P.; Galanty, A.; Michalik, M. Cytotoxic triterpene saponins from the underground parts of six Lysimachia L. species. Biochem. Syst. Ecol. 2013, 47, 116–120. [Google Scholar] [CrossRef]
- Koczurkiewicz, P.; Podolak, I.; Skrzeczyńska-Moncznik, J.; Sarna, M.; Wójcik, K.A.; Ryszawy, D.; Galanty, A.; Lasota, S.; Madeja, Z.; Czyż, J.; et al. Triterpene saponosides from Lysimachia ciliata differentially attenuate invasive potential of prostate cancer cells. Chem. Biol. Interact. 2013, 206, 6–17. [Google Scholar] [CrossRef]
- Fuchs, H. Dianthin and Its Potential in Targeted Tumor Therapies. Toxins 2019, 11, 592. [Google Scholar] [CrossRef] [Green Version]
- Weng, A.; Thakur, M.; Beceren-Braun, F.; Bachran, D.; Bachran, C.; Riese, S.B.; Jenett-Siems, K.; Gilabert-Oriol, R.; Melzig, M.F.; Fuchs, H. The toxin component of targeted anti-tumor toxins determines their efficacy increase by saponins. Mol. Oncol. 2012, 6, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Koczurkiewicz-Adamczyk, P.; Klaś, K.; Gunia-Krzyżak, A.; Piska, K.; Andrysiak, K.; Stępniewski, J.; Lasota, S.; Wójcik-Pszczoła, K.; Dulak, J.; Madeja, Z.; et al. Cinnamic Acid Derivatives as Cardioprotective Agents against Oxidative and Structural Damage Induced by Doxorubicin. Int. J. Mol. Sci. 2021, 22, 6217. [Google Scholar] [CrossRef]
- Gong, G.; Kam, H.; Tse, Y.; Lee, S.M. Cardiotoxicity of forchlorfenuron (CPPU) in zebrafish (Danio rerio) and H9c2 cardiomyocytes. Chemosphere 2019, 235, 153–162. [Google Scholar] [CrossRef]
- Kizhedath, A.; Wilkinson, S.; Glassey, J. Assessment of hepatotoxicity and dermal toxicity of butyl paraben and methyl paraben using HepG2 and HDFn in vitro models. Toxicol. Vitr. 2019, 55, 108–115. [Google Scholar] [CrossRef]
- Thimmappa, R.; Wang, S.; Zheng, M.; Misra, R.C.; Huang, A.C.; Saalbach, G.; Chang, Y.; Zhou, Z.; Hinman, V.; Bao, Z.; et al. Biosynthesis of saponin defensive compounds in sea cucumbers. Nat. Chem. Biol. 2022, 18, 774–781. [Google Scholar] [CrossRef]
- Podolak, I.; Grabowska, K.; Sobolewska, D.; Wróbel-Biedrawa, D.; Makowska-Wąs, J.; Galanty, A. Saponins as cytotoxic agents: An update (2010–2021). Part II—Triterpene saponins. Phytochem. Rev. 2022, 22, 113–167. [Google Scholar] [CrossRef]
- Sharma, P.; Tyagi, A.; Bhansali, P.; Pareek, S.; Singh, V.; Ilyas, A.; Mishra, R.; Poddar, N.K. Saponins: Extraction, bio-medicinal properties and way forward to anti-viral representatives. Food Chem. Toxicol. 2021, 150, 112075. [Google Scholar] [CrossRef]
- Zhu, H.; He, Y.S.; Ma, J.; Zhou, J.; Kong, M.; Wu, C.Y.; Mao, Q.; Lin, G.; Li, S.L. The dual roles of ginsenosides in improving the anti-tumor efficiency of cyclophosphamide in mammary carcinoma mice. J. Ethnopharmacol. 2021, 265, 113271. [Google Scholar] [CrossRef] [PubMed]
- Koczurkiewicz, P.; Klaś, K.; Grabowska, K.; Piska, K.; Rogowska, K.; Wójcik-Pszczoła, K.; Podolak, I.; Galanty, A.; Michalik, M.; Pękala, E. Saponins as chemosensitizing substances that improve effectiveness and selectivity of anticancer drug-Minireview of in vitro studies. Phytother. Res. 2019, 33, 2141–2151. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Basu, S.; Yang, Z.; Deb, A.; Hu, M. Bioavailability challenges associated with development of saponins as therapeutic and chemopreventive agents. Curr. Drug Targets 2012, 13, 1885–1899. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.L.; Gao, M.Z.; Gao, D.M.; Guo, Y.H.; Gao, Z.; Gao, X.J.; Wang, J.Q.; Qiao, M.Q. Tubeimoside-1: A review of its antitumor effects, pharmacokinetics, toxicity, and targeting preparations. Front. Pharm. 2022, 13, 941270. [Google Scholar] [CrossRef]
- Wolf, P. Targeted Toxins for the Treatment of Prostate Cancer. Biomedicines 2021, 9, 986. [Google Scholar] [CrossRef]
- Darvishi, B.; Farahmand, L.; Jalili, N.; Majidzadeh, A.K. Probable Mechanisms Involved in Immunotoxin Mediated Capillary Leak Syndrome (CLS) and Recently Developed Countering Strategies. Curr. Mol. Med. 2018, 18, 335–342. [Google Scholar] [CrossRef]
- Duvic, M.; Talpur, R. Optimizing denileukin diftitox (Ontak) therapy. Future Oncol. 2008, 4, 457–469. [Google Scholar] [CrossRef] [Green Version]
- Abou Dalle, I.; Ravandi, F. Moxetumomab pasudotox for the treatment of relapsed and/or refractory hairy cell leukemia. Expert. Rev. Hematol. 2019, 12, 707–714. [Google Scholar] [CrossRef]
- Pemmaraju, N.; Konopleva, M. Approval of tagraxofusp-erzs for blastic plasmacytoid dendritic cell neoplasm. Blood Adv. 2020, 4, 4020–4027. [Google Scholar] [CrossRef]
- Bachran, C.; Sutherland, M.; Bachran, D.; Fuchs, H. Patents on immunotoxins and chimeric toxins for the treatment of cancer. Recent Pat. Drug Deliv. 2007, 1, 105–115. [Google Scholar] [CrossRef]
- Bachran, D.; Schneider, S.; Bachran, C.; Urban, R.; Weng, A.; Melzig, M.F.; Hoffmann, C.; Kaufmann, A.M.; Fuchs, H. Epidermal growth factor receptor expression affects the efficacy of the combined application of saponin and a targeted toxin on human cervical carcinoma cells. Int. J. Cancer 2010, 127, 1453–1461. [Google Scholar] [CrossRef]
- Bhargava, C.; Dürkop, H.; Zhao, X.; Weng, A.; Melzig, M.F.; Fuchs, H. Targeted dianthin is a powerful toxin to treat pancreatic carcinoma when applied in combination with the glycosylated triterpene SO1861. Mol. Oncol. 2017, 11, 1527–1543. [Google Scholar] [CrossRef] [Green Version]
- Panjideh, H.; Niesler, N.; Weng, A.; Fuchs, H. Improved Therapy of B-Cell Non-Hodgkin Lymphoma by Obinutuzumab-Dianthin Conjugates in Combination with the Endosomal Escape Enhancer SO1861. Toxins 2022, 14, 478. [Google Scholar] [CrossRef]
- von Mallinckrodt, B.; Thakur, M.; Weng, A.; Gilabert-Oriol, R.; Dürkop, H.; Brenner, W.; Lukas, M.; Beindorff, N.; Melzig, M.F.; Fuchs, H. Dianthin-EGF is an effective tumor targeted toxin in combination with saponins in a xenograft model for colon carcinoma. Future Oncol. 2014, 10, 2161–2175. [Google Scholar] [CrossRef]
- Koczurkiewicz, P.; Podolak, I.; Wójcik, K.A.; Galanty, A.; Madeja, Z.; Michalik, M.; Czyż, J. Lclet 4 enhances pro-apoptotic and anti-invasive effects of mitoxantrone on human prostate cancer cells—In vitro study. Acta Biochim. Pol. 2013, 60, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Gilabert-Oriol, R.; Weng, A.; Trautner, A.; Weise, C.; Schmid, D.; Bhargava, C.; Niesler, N.; Wookey, P.J.; Fuchs, H.; Thakur, M. Combinatorial approach to increase efficacy of Cetuximab, Panitumumab and Trastuzumab by dianthin conjugation and co-application of SO1861. Biochem. Pharm. 2015, 97, 247–255. [Google Scholar] [CrossRef]
- Heisler, I.; Sutherland, M.; Bachran, C.; Hebestreit, P.; Schnitger, A.; Melzig, M.F.; Fuchs, H. Combined application of saponin and chimeric toxins drastically enhances the targeted cytotoxicity on tumor cells. J. Control. Release 2005, 106, 123–137. [Google Scholar] [CrossRef]
- Zhou, P.; Shi, W.; He, X.Y.; Du, Q.Y.; Wang, F.; Guo, J. Saikosaponin D: Review on the antitumour effects, toxicity and pharmacokinetics. Pharm. Biol. 2021, 59, 1480–1489. [Google Scholar] [CrossRef]
- Lin, B.; Qi, X.; Fang, L.; Zhao, L.; Zhang, R.; Jing, J.; Zhang, S.; Yang, X.; Hou, Z.; Xue, P. In vivo acute toxicity and mutagenic analysis of crude saponins from Chenopodium quinoa Willd husks. RSC Adv. 2021, 11, 4829–4841. [Google Scholar] [CrossRef]
- Vo, N.N.Q.; Fukushima, E.O.; Muranaka, T. Structure and hemolytic activity relationships of triterpenoid saponins and sapogenins. J. Nat. Med. 2017, 71, 50–58. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 (nM) | EF | |
|---|---|---|
| DE_HER14 | 0.54 | - |
| DE_NIH3T3 | 55.2 | - |
| DE_HER14_CIL1 | 0.000245 | 2200 |
| DE_NIH3T3_CIL1 | 8.01 | 6.9 |
| IC50 (nM) | EF | |
|---|---|---|
| DE_HER14 | 0.34 | - |
| DE_NIH3T3 | 39.4 | - |
| DE_HER14_CIL1 | 0.000156 | 2179 |
| DE_NIH3T3_CIL1 | 7.21 | 5.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koczurkiewicz-Adamczyk, P.; Grabowska, K.; Karnas, E.; Piska, K.; Wnuk, D.; Klaś, K.; Galanty, A.; Wójcik-Pszczoła, K.; Michalik, M.; Pękala, E.; et al. Saponin Fraction CIL1 from Lysimachia ciliata L. Enhances the Effect of a Targeted Toxin on Cancer Cells. Pharmaceutics 2023, 15, 1350. https://doi.org/10.3390/pharmaceutics15051350
Koczurkiewicz-Adamczyk P, Grabowska K, Karnas E, Piska K, Wnuk D, Klaś K, Galanty A, Wójcik-Pszczoła K, Michalik M, Pękala E, et al. Saponin Fraction CIL1 from Lysimachia ciliata L. Enhances the Effect of a Targeted Toxin on Cancer Cells. Pharmaceutics. 2023; 15(5):1350. https://doi.org/10.3390/pharmaceutics15051350
Chicago/Turabian StyleKoczurkiewicz-Adamczyk, Paulina, Karolina Grabowska, Elżbieta Karnas, Kamil Piska, Dawid Wnuk, Katarzyna Klaś, Agnieszka Galanty, Katarzyna Wójcik-Pszczoła, Marta Michalik, Elżbieta Pękala, and et al. 2023. "Saponin Fraction CIL1 from Lysimachia ciliata L. Enhances the Effect of a Targeted Toxin on Cancer Cells" Pharmaceutics 15, no. 5: 1350. https://doi.org/10.3390/pharmaceutics15051350