
Extracellular Vesicle-Based Drug Delivery Systems for Head and Neck Squamous Cell Carcinoma: A Systematic Review


Abstract
:1. Introduction
2. Materials and Methods
2.1. Search Strategy and Eligibility Criteria
2.2. Data Extraction
2.3. Assessment of Quality of Studies
3. Results
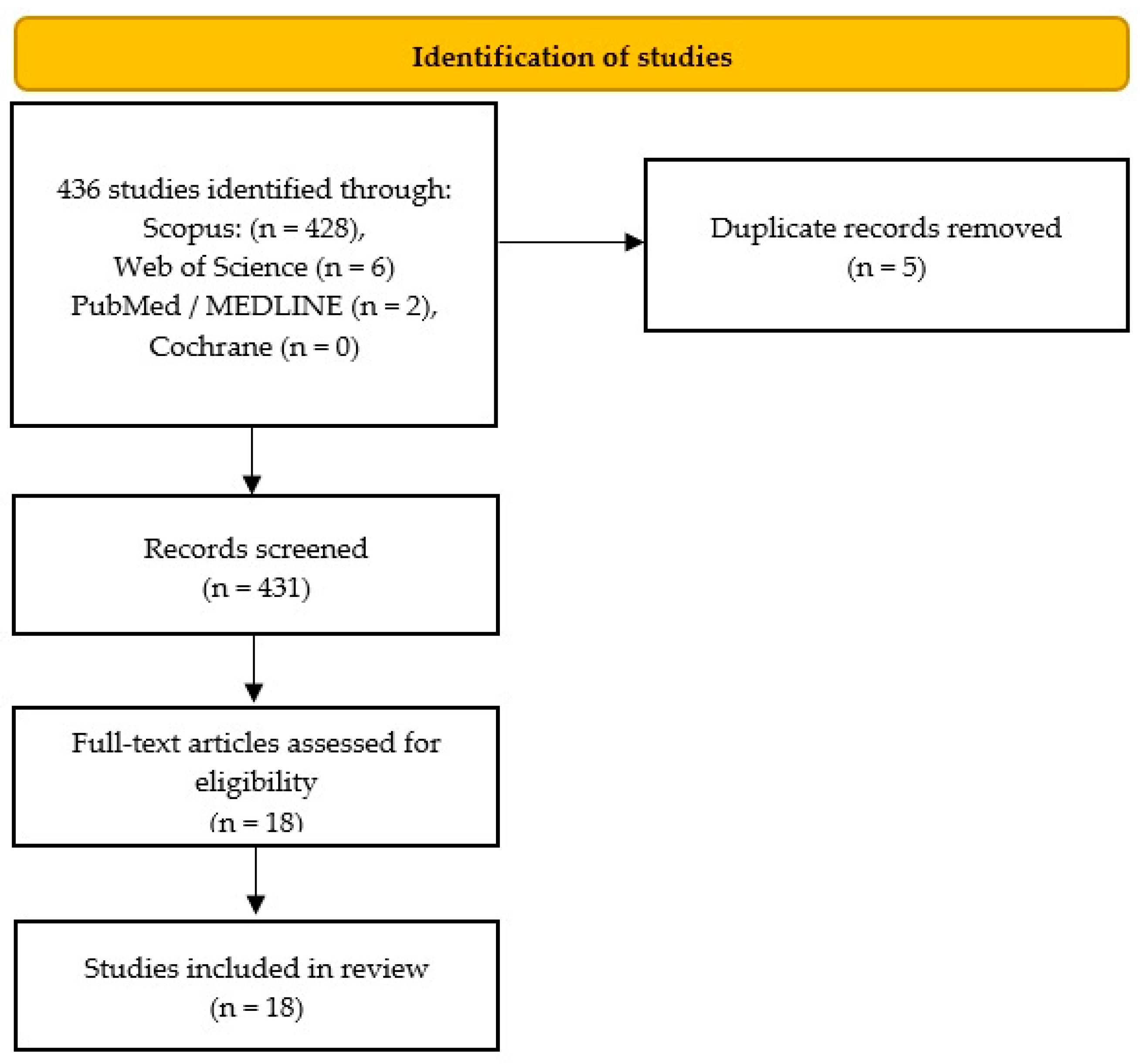
3.1. Search Results, Study Characteristics and Study Quality
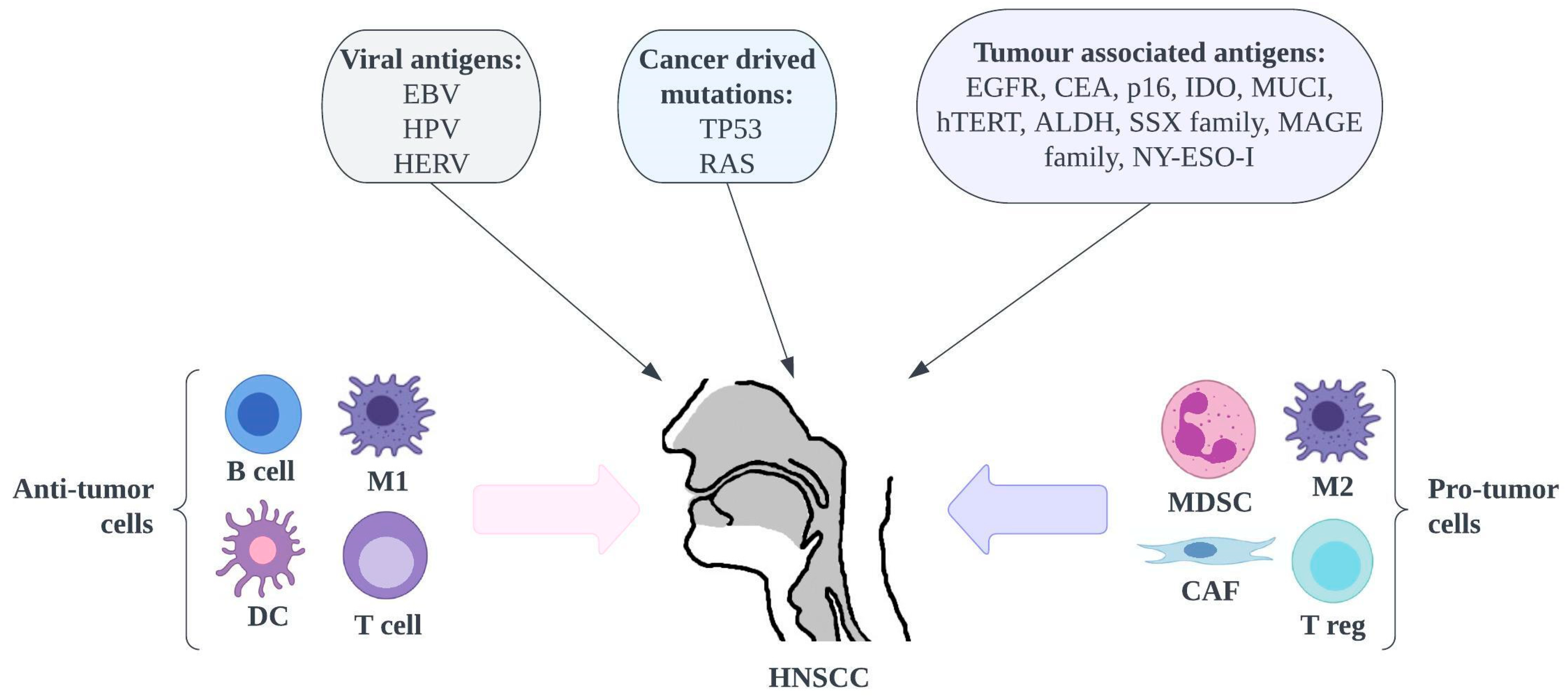
3.2. Uniqueness of HNSCC
- Unique risk factors—Cancers for Which a Vaccine Has Been Discovered.
- B.
- Specific Biomarkers for HNSCC and Second Primary Tumors.
- C.
- High Suicide Rate and Poor Quality of Life.
3.3. Head and Neck Squamous Cell Carcinoma Model
3.4. Carriers for Head and Neck Squamous Cell Carcinoma Drug Delivery Systems
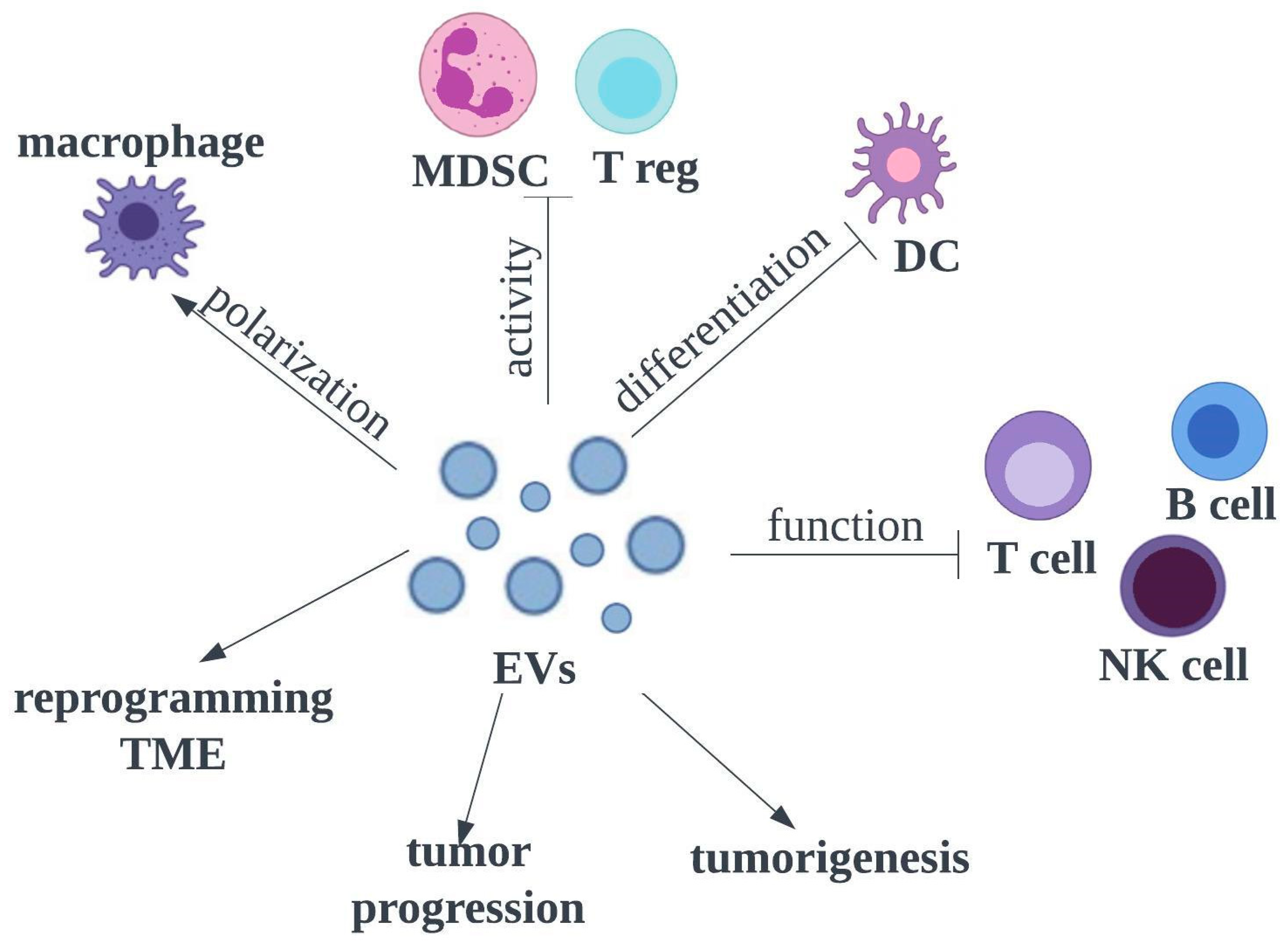
3.5. Membrane Vesicles for Drug Delivery Systems
3.5.1. Source of Extracellular Vesicles
γδT Cell-Derived Extracellular Vesicles (γδTDEVs)
Macrophage-Derived Extracellular Vesicles (M-EVs)
Mesenchymal Stem Cell-Derived Extracellular Vesicles
Carcinoma Cell and Normal Cell Lines—Derived Extracellular Vesicles
Plant-Derived Extracellular Vesicles
Bovine Milk-Derived Extracellular Vesicles
3.5.2. Extracellular Vesicles—Purification and Isolation
3.5.3. Extracellular Vesicles’ Characterization
3.5.4. Extracellular Vesicles’ Labeling and Tracking
3.5.5. Modified Extracellular Vesicles for Drug Delivery
Surface-Modified Extracellular Vesicles
Extracellular Vesicles Combined with pH-Sensitive Peptide for Drug Delivery
Extracellular Vesicle Capturing Used in Biomimetic Strategy
Engineering Extracellular Vesicles by Fusion with Liposome
Extracellular Vesicle-Coated Metal-Organic Framework Nanoparticles
ExomiR-Tracker—A Novel Drug Delivery System
Comparison of Methods for Drug Delivery Systems
3.5.6. Loading of Extracellular Vesicles
3.6. Therapeutic Effects of Membrane Vesicle-Based Drug Delivery System for Head and Neck Squamous Cell Carcinoma
3.6.1. Evaluation of Treatment Effectiveness
3.6.2. Anticancer Agents Derived from Extracellular Vesicles for Head and Neck Squamous Cell Carcinoma Treatment
3.6.3. RNA-Based Gene Therapy Delivered by EVs in the HNSCC Treatment
Extracellular Vesicle Derived miRs
- MiR-9
- 2
- MiR-18a and BART10-5p
- 3
- MiR-34a
- 4
- MiR-100-5p and miR-1246
- 5
- MiR-138
- 6
- MiR-144/451a
- 7
- MiR-155
- 8
- MiR-200c
Extracellular Vesicles-Derived Small Interfering RNA
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and neck squamous cell carcinoma. Nat. Rev. Dis. Prim. 2020, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, A.J.; Vokes, E.E. Optimizing Treatment De-Escalation in Head and Neck Cancer: Current and Future Perspectives. Oncologist 2021, 26, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Forastiere, A.A.; Zhang, Q.; Weber, R.S.; Maor, M.H.; Goepfert, H.; Pajak, T.F.; Morrison, W.; Glisson, B.; Trotti, A.; Ridge, J.A.; et al. Long-term results of RTOG 91-11: A comparison of three nonsurgical treatment strategies to preserve the larynx in patients with locally advanced larynx cancer. J. Clin. Oncol. 2013, 31, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Elsharkasy, O.M.; Nordin, J.Z.; Hagey, D.W.; de Jong, O.G.; Schiffelers, R.M.; Andaloussi, S.E.; Vader, P. Extracellular vesicles as drug delivery systems: Why and how? Adv. Drug Deliv. Rev. 2020, 159, 332–343. [Google Scholar] [CrossRef] [PubMed]
- Fischbach, C.; Chen, R.; Matsumoto, T.; Schmelzle, T.; Brugge, J.S.; Polverini, P.J.; Mooney, D.J. Engineering tumors with 3D scaffolds. Nat. Methods 2007, 4, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, I.K.; Wood, M.J.A.; Fuhrmann, G. Extracellular vesicles as a next-generation drug delivery platform. Nat. Nanotechnol. 2021, 16, 748–759. [Google Scholar] [CrossRef]
- Meng, W.; He, C.; Hao, Y.; Wang, L.; Li, L.; Zhu, G. Prospects and challenges of extracellular vesicle-based drug delivery system: Considering cell source. Drug Deliv. 2020, 27, 585–598. [Google Scholar] [CrossRef]
- Sharma, S.; Masud, M.K.; Kaneti, Y.V.; Rewatkar, P.; Koradia, A.; Hossain, M.S.A.; Yamauchi, Y.; Popat, A.; Salomon, C. Extracellular Vesicle Nanoarchitectonics for Novel Drug Delivery Applications. Small 2021, 17, e2102220. [Google Scholar] [CrossRef]
- Boucherit, N.; Gorvel, L.; Olive, D. 3D Tumor Models and Their Use for the Testing of Immunotherapies. Front. Immunol. 2020, 11, 603640. [Google Scholar] [CrossRef]
- Page, M.J.; Moher, D.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. PRISMA 2020 explanation and elaboration: Updated guidance and exemplars for reporting systematic reviews. BMJ 2021, 372, n160. [Google Scholar] [CrossRef]
- Program, N.T. Handbook for Conducting a Literature-Based Health Assessment Using OHAT Approach for Systematic Review and Evidence Integration; Office of Health Assessment and Translation (OHAT), Division of the National Toxicology Program, National Institute of Environmental Health Sciences: Durham, NC, USA, 2015; Available online: https://ntpniehsnihgov/ntp/ohat/pubs/handbookjan2015_508pdf (accessed on 31 January 2023).
- Cohen, O.; Betzer, O.; Elmaliach-Pnini, N.; Motiei, M.; Sadan, T.; Cohen-Berkman, M.; Dagan, O.; Popovtzer, A.; Yosepovich, A.; Barhom, H.; et al. ‘Golden’ exosomes as delivery vehicles to target tumors and overcome intratumoral barriers: In vivo tracking in a model for head and neck cancer. Biomater. Sci. 2021, 9, 2103–2114. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Wang, H.; Zhang, X.; Sun, X.; Zhang, J.; Ma, J. Exosomal miR-200c suppresses chemoresistance of docetaxel in tongue squamous cell carcinoma by suppressing TUBB3 and PPP2R1B. Aging 2020, 12, 6756–6773. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Meng, Y.; Wang, B.; Wang, C.X.; Hou, C.X.; Zhu, Q.H.; Tang, Y.T.; Ye, J.H. In vitro experimental study on the formation of microRNA-34a loaded exosomes and their inhibitory effect in oral squamous cell carcinoma. Cell Cycle 2022, 21, 1775–1783. [Google Scholar] [CrossRef] [PubMed]
- Kase, Y.; Uzawa, K.; Wagai, S.; Yoshimura, S.; Yamamoto, J.I.; Toeda, Y.; Okubo, M.; Eizuka, K.; Ando, T.; Nobuchi, T.; et al. Engineered exosomes delivering specific tumor-suppressive RNAi attenuate oral cancer progression. Sci. Rep. 2021, 11, 5897. [Google Scholar] [CrossRef]
- Li, K.; Qiu, Y.; Liu, X.; Huang, F. Biomimetic Nanosystems for the Synergistic Delivery of miR-144/451a for Oral Squamous Cell Carcinoma. Balk. Med. J. 2022, 39, 178–186. [Google Scholar] [CrossRef]
- Li, L.; Lu, S.; Liang, X.; Cao, B.; Wang, S.; Jiang, J.; Luo, H.; He, S.; Lang, J.; Zhu, G. γδTDEs: An Efficient Delivery System for miR-138 with Anti-tumoral and Immunostimulatory Roles on Oral Squamous Cell Carcinoma. Mol. Ther. Nucleic Acids 2019, 14, 101–113. [Google Scholar] [CrossRef]
- Liu, P.; Zhang, Q.; Mi, J.; Wang, S.; Xu, Q.; Zhuang, D.; Chen, W.; Liu, C.; Zhang, L.; Guo, J.; et al. Exosomes derived from stem cells of human deciduous exfoliated teeth inhibit angiogenesis in vivo and in vitro via the transfer of miR-100-5p and miR-1246. Stem Cell Res. Ther. 2022, 13, 89. [Google Scholar] [CrossRef]
- Liu, Y.; Li, H.; Qiu, Y.L.; Li, K.S.; Lu, Y.; Wang, W. Nanoscale biomimetic nano system for the co-delivery of sns032 and tumor necrosis factor related apoptosis inducing ligand to enhance therapeutic efficacy in oral squamous cell carcinoma cell line scc25. Mater. Express 2021, 11, 1321–1330. [Google Scholar] [CrossRef]
- Qiu, Y.L.; Sun, J.M.; Qiu, J.P.; Chen, G.L.; Wang, X.; Mu, Y.X.; Li, K.S.; Wang, W.J. Antitumor Activity of Cabazitaxel and MSC-TRAIL Derived Extracellular Vesicles in Drug-Resistant Oral Squamous Cell Carcinoma. Cancer Manag. Res. 2020, 12, 10809–10820. [Google Scholar] [CrossRef]
- Sayyed, A.A.; Gondaliya, P.; Mali, M.; Pawar, A.; Bhat, P.; Khairnar, A.; Arya, N.; Kalia, K. MiR-155 Inhibitor-Laden Exosomes Reverse Resistance to Cisplatin in a 3D Tumor Spheroid and Xenograft Model of Oral Cancer. Mol. Pharm. 2021, 18, 3010–3025. [Google Scholar] [CrossRef]
- Tong, F.; Mao, X.; Zhang, S.; Xie, H.; Yan, B.; Wang, B.; Sun, J.; Wei, L. HPV + HNSCC-derived exosomal miR-9 induces macrophage M1 polarization and increases tumor radiosensitivity. Cancer Lett. 2020, 478, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, L.E.I.; Huang, Y.; Li, K.U.N.; Jinye, A.; Fan, T.; Zhao, R.E.N.; Xia, X.; Shen, B.; Du, J.; et al. Exosome-delivered TRPP2 siRNA inhibits the epithelial-mesenchymal transition of FaDu cells. Oncol. Lett. 2019, 17, 1953–1961. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jiang, Q.; Faleti, O.D.; Tsang, C.M.; Zhao, M.; Wu, G.; Tsao, S.W.; Fu, M.; Chen, Y.; Ding, T.; et al. Exosomal Delivery of AntagomiRs Targeting Viral and Cellular MicroRNAs Synergistically Inhibits Cancer Angiogenesis. Mol. Ther. Nucleic Acids 2020, 22, 153–165. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Y.; Mu, X.; Tu, C.R.; Chung, Y.; Tsao, S.W.; Chan, G.C.F.; Leung, W.H.; Lau, Y.L.; Liu, Y.; et al. Exosomes derived from 3δ -T cells synergize with radiotherapy and preserve antitumor activities against nasopharyngeal carcinoma in immunosuppressive microenvironment. J. ImmunoTher. Cancer 2022, 10, e003832. [Google Scholar] [CrossRef]
- Yakavets, I.; Francois, A.; Lamy, L.; Piffoux, M.; Gazeau, F.; Wilhelm, C.; Zorin, V.; Silva, A.K.A.; Bezdetnaya, L. Effect of stroma on the behavior of temoporfin-loaded lipid nanovesicles inside the stroma-rich head and neck carcinoma spheroids. J. Nanobiotechnology 2021, 19, 3. [Google Scholar] [CrossRef] [PubMed]
- Yamayoshi, A.; Oyama, S.; Kishimoto, Y.; Konishi, R.; Yamamoto, T.; Kobori, A.; Harada, H.; Ashihara, E.; Sugiyama, H.; Murakami, A. Development of antibody–oligonucleotide complexes for targeting exosomal microrna. Pharmaceutics 2020, 12, 545. [Google Scholar] [CrossRef]
- Yang, M.; Luo, Q.; Chen, X.; Chen, F. Bitter melon derived extracellular vesicles enhance the therapeutic effects and reduce the drug resistance of 5-fluorouracil on oral squamous cell carcinoma. J. Nanobiotechnol. 2021, 19, 259. [Google Scholar] [CrossRef]
- Zhang, Q.; Xiao, Q.; Yin, H.; Xia, C.; Pu, Y.; He, Z.; Hu, Q.; Wang, J.; Wang, Y. Milk-exosome based pH/light sensitive drug system to enhance anticancer activity against oral squamous cell carcinoma. RSC Adv. 2020, 10, 28314–28323. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Trivedi, S.; Sun, L.; Aggarwal, C. Immunotherapy for Head and Neck Cancer. Hematol. Oncol. Clin. N. Am. 2021, 35, 1021–1037. [Google Scholar] [CrossRef]
- Jurkiewicz, D.; Dżaman, K.; Rapiejko, P. Laryngeal cancer risk factors. Pol. Merkur. Lek. 2006, 21, 94–98. [Google Scholar]
- Gillison, M.L.; Chaturvedi, A.K.; Lowy, D.R. HPV prophylactic vaccines and the potential prevention of noncervical cancers in both men and women. Cancer 2008, 113, 3036–3046. [Google Scholar] [CrossRef] [PubMed]
- Tsur, N.; Frig, O.; Steinberg-Shemer, O.; Tamary, H.; Kurman, N.; Mizrachi, A.; Popovtzer, A. Characterization of Fanconi Anemia Patients with Head and Neck Squamous Cell Carcinoma: Israel Fanconi Registry. Isr. Med. Assoc. J. 2022, 24, 491–496. [Google Scholar] [PubMed]
- Fang, M.; Huang, W.; Mo, D.; Zhao, W.; Huang, R. Association of Five Snps in Cytotoxic T-Lymphocyte Antigen 4 and Cancer Susceptibility: Evidence from 67 Studies. Cell Physiol. Biochem. 2018, 47, 414–427. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, H.; Duan, G.; Wang, H. The association of the CYP1A1 Ile462Val polymorphism with head and neck cancer risk: Evidence based on a cumulative meta-analysis. OncoTargets Ther. 2016, 9, 2927–2934. [Google Scholar] [CrossRef] [PubMed]
- Wong, I.C.; Ng, Y.K.; Lui, V.W. Cancers of the lung, head and neck on the rise: Perspectives on the genotoxicity of air pollution. Chin. J. Cancer 2014, 33, 476–480. [Google Scholar] [CrossRef]
- Yu, S.S.; Cirillo, N. The molecular markers of cancer stem cells in head and neck tumors. J. Cell Physiol. 2020, 235, 65–73. [Google Scholar] [CrossRef]
- Zhang, Q.; Shi, S.; Yen, Y.; Brown, J.; Ta, J.Q.; Le, A.D. A subpopulation of CD133(+) cancer stem-like cells characterized in human oral squamous cell carcinoma confer resistance to chemotherapy. Cancer Lett. 2010, 289, 151–160. [Google Scholar] [CrossRef]
- Antra; Parashar, P.; Hungyo, H.; Jain, A.; Ahmad, S.; Tandon, V. Unraveling molecular mechanisms of head and neck cancer. Crit. Rev. Oncol. Hematol. 2022, 178, 103778. [Google Scholar] [CrossRef]
- Leon, X.; Garcia, J.; Lopez, M.; Rodriguez, C.; Gutierrez, A.; Quer, M. Risk of onset of second neoplasms and successive neoplasms in patients with a head and neck index tumour. Acta Otorrinolaringol. Esp. 2020, 71, 9–15. [Google Scholar] [CrossRef]
- Osazuwa-Peters, N.; Simpson, M.C.; Zhao, L.; Boakye, E.A.; Olomukoro, S.I.; Deshields, T.; Loux, T.M.; Varvares, M.A.; Schootman, M. Suicide risk among cancer survivors: Head and neck versus other cancers. Cancer 2018, 124, 4072–4079. [Google Scholar] [CrossRef]
- Nayak, S.G.; Pai, M.S.; George, L.S. Quality of life of patients with head and neck cancer: A mixed method study. J. Cancer Res. Ther. 2019, 15, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Bonomo, P.; Paderno, A.; Mattavelli, D.; Zenda, S.; Cavalieri, S.; Bossi, P. Quality Assessment in Supportive Care in Head and Neck Cancer. Front. Oncol. 2019, 9, 926. [Google Scholar] [CrossRef] [PubMed]
- Hutcheson, K.A.; Nurgalieva, Z.; Zhao, H.; Gunn, G.B.; Giordano, S.H.; Bhayani, M.K.; Lewin, J.S.; Lewis, C.M. Two-year prevalence of dysphagia and related outcomes in head and neck cancer survivors: An updated SEER-Medicare analysis. Head Neck 2019, 41, 479–487. [Google Scholar] [CrossRef]
- Fais, S. NK cell-released exosomes: Natural nanobullets against tumors. Oncoimmunology 2013, 2, e22337. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.M.; Andre, F.; Amigorena, S.; Soria, J.C.; Eggermont, A.; Kroemer, G.; Zitvogel, L. Dendritic cell-derived exosomes for cancer therapy. J. Clin. Investig. 2016, 126, 1224–1232. [Google Scholar] [CrossRef] [PubMed]
- Ventimiglia, L.N.; Alonso, M.A. Biogenesis and Function of T Cell-Derived Exosomes. Front. Cell Dev. Biol. 2016, 4, 84. [Google Scholar] [CrossRef]
- Yoshikawa, T.; Takahara, M.; Tomiyama, M.; Nieda, M.; Maekawa, R.; Nakatsura, T. Large-scale expansion of gammadelta T cells and peptide-specific cytotoxic T cells using zoledronate for adoptive immunotherapy. Int. J. Oncol. 2014, 45, 1847–1856. [Google Scholar] [CrossRef]
- Wang, X.; Xiang, Z.; Liu, Y.; Huang, C.; Pei, Y.; Wang, X.; Zhi, H.; Wong, W.H.; Wei, H.; Ng, I.O.; et al. Exosomes derived from Vdelta2-T cells control Epstein-Barr virus-associated tumors and induce T cell antitumor immunity. Sci. Transl. Med. 2020, 12, eaaz3426. [Google Scholar] [CrossRef]
- Gordon, S.; Martinez, F.O. Alternative activation of macrophages: Mechanism and functions. Immunity 2010, 32, 593–604. [Google Scholar] [CrossRef]
- Liu, J.; Wu, F.; Zhou, H. Macrophage-derived exosomes in cancers: Biogenesis, functions and therapeutic applications. Immunol. Lett. 2020, 227, 102–108. [Google Scholar] [CrossRef]
- Lai, C.P.; Kim, E.Y.; Badr, C.E.; Weissleder, R.; Mempel, T.R.; Tannous, B.A.; Breakefield, X.O. Visualization and tracking of tumour extracellular vesicle delivery and RNA translation using multiplexed reporters. Nat. Commun. 2015, 6, 7029. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Shen, B.; Jiang, F.; Xia, L.; Fan, T.; Qin, M.; Yang, L.; Guo, J.; Li, Y.; Zhu, M.; et al. TRPP2 Enhances Metastasis by Regulating Epithelial-Mesenchymal Transition in Laryngeal Squamous Cell Carcinoma. Cell Physiol. Biochem. 2016, 39, 2203–2215. [Google Scholar] [CrossRef] [PubMed]
- Martinez Saez, D.; Sasaki, R.T.; Neves, A.D.; da Silva, M.C. Stem Cells from Human Exfoliated Deciduous Teeth: A Growing Literature. Cells Tissues Organs 2016, 202, 269–280. [Google Scholar] [CrossRef]
- Zhou, J.; Yang, Y.; Wang, W.; Zhang, Y.; Chen, Z.; Hao, C.; Zhang, J. Melanoma-released exosomes directly activate the mitochondrial apoptotic pathway of CD4(+) T cells through their microRNA cargo. Exp. Cell Res. 2018, 371, 364–371. [Google Scholar] [CrossRef]
- Raimondo, S.; Naselli, F.; Fontana, S.; Monteleone, F.; Lo Dico, A.; Saieva, L.; Zito, G.; Flugy, A.; Manno, M.; Di Bella, M.A.; et al. Citrus limon-derived nanovesicles inhibit cancer cell proliferation and suppress CML xenograft growth by inducing TRAIL-mediated cell death. Oncotarget 2015, 6, 19514–19527. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Yoo, H.J.; Jung, J.H.; Lee, R.; Hyun, J.K.; Park, J.H.; Na, D.; Yeon, J.H. Cytotoxic Effects of Plant Sap-Derived Extracellular Vesicles on Various Tumor Cell Types. J. Funct. Biomater. 2020, 11, 22. [Google Scholar] [CrossRef]
- Kameli, N.; Dragojlovic-Kerkache, A.; Savelkoul, P.; Stassen, F.R. Plant-Derived Extracellular Vesicles: Current Findings, Challenges, and Future Applications. Membranes 2021, 11, 411. [Google Scholar] [CrossRef]
- Hata, T.; Murakami, K.; Nakatani, H.; Yamamoto, Y.; Matsuda, T.; Aoki, N. Isolation of bovine milk-derived microvesicles carrying mRNAs and microRNAs. Biochem. Biophys. Res. Commun. 2010, 396, 528–533. [Google Scholar] [CrossRef]
- Samuel, M.; Fonseka, P.; Sanwlani, R.; Gangoda, L.; Chee, S.H.; Keerthikumar, S.; Spurling, A.; Chitti, S.V.; Zanker, D.; Ang, C.S.; et al. Oral administration of bovine milk-derived extracellular vesicles induces senescence in the primary tumor but accelerates cancer metastasis. Nat. Commun. 2021, 12, 3950. [Google Scholar] [CrossRef]
- Vader, P.; Mol, E.A.; Pasterkamp, G.; Schiffelers, R.M. Extracellular vesicles for drug delivery. Adv. Drug Deliv. Rev. 2016, 106, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Bahl, A.; Wilson, W.; Ball, J.; Renninson, E.; Dubey, S.; Bravo, A.; Foulstone, E.; Spensley, S.; Bowen, R.; Mansi, J.; et al. Concept: A randomised multicentre trial of first line chemotherapy comparing three weekly cabazitaxel versus weekly paclitaxel in HER2 negative metastatic breast cancer. Breast 2022, 66, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Momen-Heravi, F.; Balaj, L.; Alian, S.; Mantel, P.Y.; Halleck, A.E.; Trachtenberg, A.J.; Soria, C.E.; Oquin, S.; Bonebreak, C.M.; Saracoglu, E.; et al. Current methods for the isolation of extracellular vesicles. Biol. Chem. 2013, 394, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Buschmann, D.; Kirchner, B.; Hermann, S.; Marte, M.; Wurmser, C.; Brandes, F.; Kotschote, S.; Bonin, M.; Steinlein, O.K.; Pfaffl, M.W.; et al. Evaluation of serum extracellular vesicle isolation methods for profiling miRNAs by next-generation sequencing. J. Extracell. Vesicles 2018, 7, 1481321. [Google Scholar] [CrossRef]
- Gurunathan, S.; Kang, M.H.; Jeyaraj, M.; Qasim, M.; Kim, J.H. Review of the Isolation, Characterization, Biological Function, and Multifarious Therapeutic Approaches of Exosomes. Cells 2019, 8, 307. [Google Scholar] [CrossRef]
- Noble, J.M.; Roberts, L.M.; Vidavsky, N.; Chiou, A.E.; Fischbach, C.; Paszek, M.J.; Estroff, L.A.; Kourkoutis, L.F. Direct comparison of optical and electron microscopy methods for structural characterization of extracellular vesicles. J. Struct. Biol. 2020, 210, 107474. [Google Scholar] [CrossRef]
- Betzer, O.; Barnoy, E.; Sadan, T.; Elbaz, I.; Braverman, C.; Liu, Z.; Popovtzer, R. Advances in imaging strategies for in vivo tracking of exosomes. WIREs Nanomed. Nanobiotechnol. 2020, 12, e1594. [Google Scholar] [CrossRef]
- Qiao, L.; Hu, S.; Huang, K.; Su, T.; Li, Z.; Vandergriff, A.; Cores, J.; Dinh, P.U.; Allen, T.; Shen, D.; et al. Tumor cell-derived exosomes home to their cells of origin and can be used as Trojan horses to deliver cancer drugs. Theranostics 2020, 10, 3474–3487. [Google Scholar] [CrossRef]
- Smyth, T.; Kullberg, M.; Malik, N.; Smith-Jones, P.; Graner, M.W.; Anchordoquy, T.J. Biodistribution and delivery efficiency of unmodified tumor-derived exosomes. J. Control. Release 2015, 199, 145–155. [Google Scholar] [CrossRef]
- Choi, H.; Lee, D.S. Illuminating the physiology of extracellular vesicles. Stem Cell Res. Ther. 2016, 7, 55. [Google Scholar] [CrossRef]
- Tian, T.; Zhu, Y.L.; Hu, F.H.; Wang, Y.Y.; Huang, N.P.; Xiao, Z.D. Dynamics of exosome internalization and trafficking. J. Cell Physiol. 2013, 228, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.P.; Mardini, O.; Ericsson, M.; Prabhakar, S.; Maguire, C.; Chen, J.W.; Tannous, B.A.; Breakefield, X.O. Dynamic biodistribution of extracellular vesicles in vivo using a multimodal imaging reporter. ACS Nano 2014, 8, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Perets, N.; Betzer, O.; Shapira, R.; Brenstein, S.; Angel, A.; Sadan, T.; Ashery, U.; Popovtzer, R.; Offen, D. Golden Exosomes Selectively Target Brain Pathologies in Neurodegenerative and Neurodevelopmental Disorders. Nano Lett. 2019, 19, 3422–3431. [Google Scholar] [CrossRef] [PubMed]
- Counihan, J.L.; Grossman, E.A.; Nomura, D.K. Cancer Metabolism: Current Understanding and Therapies. Chem. Rev. 2018, 118, 6893–6923. [Google Scholar] [CrossRef] [PubMed]
- Duan, L.; Xu, L.; Xu, X.; Qin, Z.; Zhou, X.; Xiao, Y.; Liang, Y.; Xia, J. Exosome-mediated delivery of gene vectors for gene therapy. Nanoscale 2021, 13, 1387–1397. [Google Scholar] [CrossRef]
- Kang, Y.; Xu, C.; Meng, L.; Dong, X.; Qi, M.; Jiang, D. Exosome-functionalized magnesium-organic framework-based scaffolds with osteogenic, angiogenic and anti-inflammatory properties for accelerated bone regeneration. Bioact. Mater. 2022, 18, 26–41. [Google Scholar] [CrossRef]
- Miroshnichenko, S.K.; Patutina, O.A.; Burakova, E.A.; Chelobanov, B.P.; Fokina, A.A.; Vlassov, V.V.; Altman, S.; Zenkova, M.A.; Stetsenko, D.A. Mesyl phosphoramidate antisense oligonucleotides as an alternative to phosphorothioates with improved biochemical and biological properties. Proc. Natl. Acad. Sci. USA 2019, 116, 1229–1234. [Google Scholar] [CrossRef]
- Li, L.; Li, C.; Wang, S.; Wang, Z.; Jiang, J.; Wang, W.; Li, X.; Chen, J.; Liu, K.; Li, C.; et al. Exosomes Derived from Hypoxic Oral Squamous Cell Carcinoma Cells Deliver miR-21 to Normoxic Cells to Elicit a Prometastatic Phenotype. Cancer Res. 2016, 76, 1770–1780. [Google Scholar] [CrossRef]
- Yanez-Mo, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borras, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef]
- Zheng, H.C. The molecular mechanisms of chemoresistance in cancers. Oncotarget 2017, 8, 59950–59964. [Google Scholar] [CrossRef]
- Corn, P.G.; Heath, E.I.; Zurita, A.; Ramesh, N.; Xiao, L.; Sei, E.; Li-Ning-Tapia, E.; Tu, S.M.; Subudhi, S.K.; Wang, J.; et al. Cabazitaxel plus carboplatin for the treatment of men with metastatic castration-resistant prostate cancers: A randomised, open-label, phase 1–2 trial. Lancet Oncol. 2019, 20, 1432–1443. [Google Scholar] [CrossRef] [PubMed]
- Huo, R.; Wang, L.; Liu, P.; Zhao, Y.; Zhang, C.; Bai, B.; Liu, X.; Shi, C.; Wei, S.; Zhang, H. Cabazitaxel-induced autophagy via the PI3K/Akt/mTOR pathway contributes to A549 cell death. Mol. Med. Rep. 2016, 14, 3013–3020. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Xuan, Z.; Chen, J.; Song, W.; Zhang, S.; Jin, C.; Zhou, M.; Zheng, S.; Song, P. Inhibiting the NF-kappaB pathway enhances the antitumor effect of cabazitaxel by downregulating Bcl-2 in pancreatic cancer. Int. J. Oncol. 2020, 57, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Vikas; Mehata, A.K.; Suseela, M.N.L.; Behera, C.; Kumari, P.; Mahto, S.K.; Muthu, M.S. Chitosan-alginate nanoparticles of cabazitaxel: Design, dual-receptor targeting and efficacy in lung cancer model. Int. J. Biol. Macromol. 2022, 221, 874–890. [Google Scholar] [CrossRef]
- Di Leva, G.; Garofalo, M.; Croce, C.M. MicroRNAs in cancer. Annu. Rev. Pathol. 2014, 9, 287–314. [Google Scholar] [CrossRef]
- Garzon, R.; Calin, G.A.; Croce, C.M. MicroRNAs in Cancer. Annu. Rev. Med. 2009, 60, 167–179. [Google Scholar] [CrossRef]
- Shi, X.; Shiao, S.L. The role of macrophage phenotype in regulating the response to radiation therapy. Transl. Res. 2018, 191, 64–80. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, S.; Tong, F.; Wang, Y.; Wei, L. HPV(+) HNSCC-derived exosomal miR-9-5p inhibits TGF-beta signaling-mediated fibroblast phenotypic transformation through NOX4. Cancer Sci. 2022, 113, 1475–1487. [Google Scholar] [CrossRef]
- Cai, L.; Ye, Y.; Jiang, Q.; Chen, Y.; Lyu, X.; Li, J.; Wang, S.; Liu, T.; Cai, H.; Yao, K.; et al. Epstein-Barr virus-encoded microRNA BART1 induces tumour metastasis by regulating PTEN-dependent pathways in nasopharyngeal carcinoma. Nat. Commun. 2015, 6, 7353. [Google Scholar] [CrossRef]
- Ge, X.; Gao, J.; Sun, Q.W.; Wang, C.X.; Deng, W.; Mao, G.Y.; Li, H.Q.; Guo, S.S.; Cheng, J.; Wu, Y.N.; et al. MiR-34a inhibits the proliferation, migration, and invasion of oral squamous cell carcinoma by directly targeting SATB2. J. Cell Physiol. 2020, 235, 4856–4864. [Google Scholar] [CrossRef]
- Roy, S.K.; Shrivastava, A.; Srivastav, S.; Shankar, S.; Srivastava, R.K. SATB2 is a novel biomarker and therapeutic target for cancer. J. Cell Mol. Med. 2020, 24, 11064–11069. [Google Scholar] [CrossRef]
- Bai, Y.; Wang, W.; Zhang, Y.; Zhang, F.; Zhang, H. lncRNA MIAT suppression alleviates corneal angiogenesis through regulating miR-1246/ACE. Cell Cycle 2019, 18, 661–669. [Google Scholar] [CrossRef]
- Pakravan, K.; Babashah, S.; Sadeghizadeh, M.; Mowla, S.J.; Mossahebi-Mohammadi, M.; Ataei, F.; Dana, N.; Javan, M. MicroRNA-100 shuttled by mesenchymal stem cell-derived exosomes suppresses in vitro angiogenesis through modulating the mTOR/HIF-1alpha/VEGF signaling axis in breast cancer cells. Cell Oncol. 2017, 40, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, Z.; Yu, J.; Xia, J.; Zhou, X. MicroRNA profiling and head and neck cancer. Comp. Funct. Genom. 2009, 2009, 837514. [Google Scholar] [CrossRef] [PubMed]
- Manikandan, M.; Rao, A.K.D.M.; Rajkumar, K.S.; Rajaraman, R.; Munirajan, A.K. Altered levels of miR-21, miR-125b-2*, miR-138, miR-155, miR-184, and miR-205 in oral squamous cell carcinoma and association with clinicopathological characteristics. J. Oral Pathol. Med. 2015, 44, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Wong, T.S.; Liu, X.B.; Wong, B.Y.; Ng, R.W.; Yuen, A.P.; Wei, W.I. Mature miR-184 as Potential Oncogenic microRNA of Squamous Cell Carcinoma of Tongue. Clin. Cancer Res. 2008, 14, 2588–2592. [Google Scholar] [CrossRef]
- Jin, Y.; Chen, D.; Cabay, R.J.; Wang, A.; Crowe, D.L.; Zhou, X. Role of microRNA-138 as a potential tumor suppressor in head and neck squamous cell carcinoma. Int. Rev. Cell Mol. Biol. 2013, 303, 357–385. [Google Scholar] [CrossRef]
- Wang, H.M.; Lu, Y.J.; He, L.; Gu, N.J.; Wang, S.Y.; Qiu, X.S.; Wang, E.H.; Wu, G.P. HPV16 E6/E7 promote the translocation and glucose uptake of GLUT1 by PI3K/AKT pathway via relieving miR-451 inhibitory effect on CAB39 in lung cancer cells. Ther. Adv. Chronic Dis. 2020, 11, 2040622320957143. [Google Scholar] [CrossRef]
- Zeng, Q.; Tao, X.; Huang, F.; Wu, T.; Wang, J.; Jiang, X.; Kuang, Z.; Cheng, B. Overexpression of miR-155 promotes the proliferation and invasion of oral squamous carcinoma cells by regulating BCL6/cyclin D2. Int. J. Mol. Med. 2016, 37, 1274–1280. [Google Scholar] [CrossRef]
- Ahn, H.; Kim, H.; Abdul, R.; Kim, Y.; Sim, J.; Choi, D.; Paik, S.S.; Shin, S.J.; Kim, D.H.; Jang, K. Overexpression of Forkhead Box O3a and Its Association with Aggressive Phenotypes and Poor Prognosis in Human Hepatocellular Carcinoma. Am. J. Clin. Pathol. 2018, 149, 117–127. [Google Scholar] [CrossRef]
- Tian, Y.; Qi, P.; Hu, X. Downregulated FOXO3a Associates with Poor Prognosis and Promotes Cell Invasion and Migration via WNT/beta-catenin Signaling in Cervical Carcinoma. Front. Oncol. 2020, 10, 903. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, R.D.; Ghuwalewala, S.; Das, P.; Mandloi, S.; Alam, S.K.; Chakraborty, J.; Sarkar, S.; Chakrabarti, S.; Panda, C.K.; Roychoudhury, S. MicroRNA profiling of cisplatin-resistant oral squamous cell carcinoma cell lines enriched with cancer-stem-cell-like and epithelial-mesenchymal transition-type features. Sci. Rep. 2016, 6, 23932. [Google Scholar] [CrossRef] [PubMed]
- Zheng, R.; Liu, Y.; Zhang, X.; Zhao, P.; Deng, Q. miRNA-200c enhances radiosensitivity of esophageal cancer by cell cycle arrest and targeting P21. Biomed Pharmacother. 2017, 90, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Zhang, C.; Hu, Y.; Qin, H.; Gu, A.; Li, Y.; Zhang, L.; Li, Z.; Wang, Y. miRNA-200c mediates mono-butyl phthalate-disrupted steroidogenesis by targeting vimentin in Leydig tumor cells and murine adrenocortical tumor cells. Toxicol. Lett. 2016, 241, 95–102. [Google Scholar] [CrossRef]
- Luo, Z.; Wen, G.; Wang, G.; Pu, X.; Ye, S.; Xu, Q.; Wang, W.; Xiao, Q. MicroRNA-200C and -150 play an important role in endothelial cell differentiation and vasculogenesis by targeting transcription repressor ZEB1. Stem Cells 2013, 31, 1749–1762. [Google Scholar] [CrossRef]
- Ma, C.; Ding, Y.C.; Yu, W.; Wang, Q.; Meng, B.; Huang, T. MicroRNA-200c overexpression plays an inhibitory role in human pancreatic cancer stem cells by regulating epithelial-mesenchymal transition. Minerva Med. 2015, 106, 193–202. [Google Scholar]
- Chen, Y.; Sun, Y.; Chen, L.; Xu, X.; Zhang, X.; Wang, B.; Min, L.; Liu, W. miRNA-200c increases the sensitivity of breast cancer cells to doxorubicin through the suppression of E-cadherin-mediated PTEN/Akt signaling. Mol. Med. Rep. 2013, 7, 1579–1584. [Google Scholar] [CrossRef]
- Choi, Y.J.; Lee, K.; Yoon, J.H.; Cho, S.G.; Kim, Y.G.; Jeong, M.; Hwang, H.H.; Lee, S.Y.; Jung, S.E.; Ko, S.G. SH003 and Docetaxel Show Synergistic Anticancer Effects by Inhibiting EGFR Activation in Triple-Negative Breast Cancer. Biomed Res. Int. 2022, 2022, 3647900. [Google Scholar] [CrossRef]
- Feng, B.; Wang, R.; Song, H.Z.; Chen, L.B. MicroRNA-200b reverses chemoresistance of docetaxel-resistant human lung adenocarcinoma cells by targeting E2F3. Cancer 2012, 118, 3365–3376. [Google Scholar] [CrossRef]
- Han, G.; Xu, C.; Yu, D. Mechanisms correlated with chemotherapy resistance in tongue cancers. J. Cancer Res. Ther. 2018, 14, 1–5. [Google Scholar] [CrossRef]
- Jiang, Y.; Huang, W.; Sun, X.; Yang, X.; Wu, Y.; Shi, J.; Zheng, J.; Fan, S.; Liu, J.; Wang, J.; et al. DTX-P7, a peptide-drug conjugate, is highly effective for non-small cell lung cancer. J. Hematol. Oncol. 2022, 15, 73. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Lee, Y.J. Synergistic anticancer activity of resveratrol in combination with docetaxel in prostate carcinoma cells. Nutr. Res. Pract. 2021, 15, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Wang, H.; Zhang, L.; Mei, X.; Hu, J.; Huang, K. Receptor for advanced glycation end product blockade enhances the chemotherapeutic effect of cisplatin in tongue squamous cell carcinoma by reducing autophagy and modulating the Wnt pathway. Anti-Cancer Drugs 2017, 28, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, J.; Chandrashekar, C.; Radhakrishnan, R. Critical biomarkers of epithelial-mesenchymal transition in the head and neck cancers. J. Cancer Res. Ther. 2014, 10, 512–518. [Google Scholar] [PubMed]
- Koide, N.; Kasamatsu, A.; Endo-Sakamoto, Y.; Ishida, S.; Shimizu, T.; Kimura, Y.; Miyamoto, I.; Yoshimura, S.; Shiiba, M.; Tanzawa, H.; et al. Evidence for Critical Role of Lymphocyte Cytosolic Protein 1 in Oral Cancer. Sci. Rep. 2017, 7, 43379. [Google Scholar] [CrossRef]
- Czerwaty, K.; Dzaman, K.; Miechowski, W. Application of Extracellular Vesicles in Allergic Rhinitis: A Systematic Review. Int. J. Mol. Sci. 2022, 24, 367. [Google Scholar] [CrossRef]
- Dzaman, K.; Czerwaty, K. Roles of Exosomes in Chronic Rhinosinusitis: A Systematic Review. Int. J. Mol. Sci. 2022, 23, 11284. [Google Scholar] [CrossRef]
- Hofmann, L.; Ludwig, S.; Vahl, J.M.; Brunner, C.; Hoffmann, T.K.; Theodoraki, M.N. The Emerging Role of Exosomes in Diagnosis, Prognosis, and Therapy in Head and Neck Cancer. Int. J. Mol. Sci. 2020, 21, 4072. [Google Scholar] [CrossRef]
- Mathew, M.; Zade, M.; Mezghani, N.; Patel, R.; Wang, Y.; Momen-Heravi, F. Extracellular Vesicles as Biomarkers in Cancer Immunotherapy. Cancers 2020, 12, 2825. [Google Scholar] [CrossRef]
- Xiao, C.; Song, F.; Zheng, Y.L.; Lv, J.; Wang, Q.F.; Xu, N. Exosomes in Head and Neck Squamous Cell Carcinoma. Front. Oncol. 2019, 9, 894. [Google Scholar] [CrossRef]
- Li, T.; Li, J.; Wang, H.; Zhao, J.; Yan, M.; He, H.; Yu, S. Exosomes: Potential Biomarkers and Functions in Head and Neck Squamous Cell Carcinoma. Front. Mol. Biosci. 2022, 9, 881794. [Google Scholar] [CrossRef] [PubMed]
- Perez-Herrero, E.; Fernandez-Medarde, A. Advanced targeted therapies in cancer: Drug nanocarriers, the future of chemotherapy. Eur. J. Pharm. Biopharm. 2015, 93, 52–79. [Google Scholar] [CrossRef] [PubMed]
- Riley, R.S.; June, C.H.; Langer, R.; Mitchell, M.J. Delivery technologies for cancer immunotherapy. Nat. Rev. Drug Discov. 2019, 18, 175–196. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.J.; Sun, X.Y.; Huang, K.M.; Zhang, L.; Yang, Z.S.; Zou, D.D.; Wang, B.; Warnock, G.L.; Dai, L.J.; Luo, J. Therapeutic potential of CAR-T cell-derived exosomes: A cell-free modality for targeted cancer therapy. Oncotarget 2015, 6, 44179–44190. [Google Scholar] [CrossRef]
- Singh, K.; Nalabotala, R.; Koo, K.M.; Bose, S.; Nayak, R.; Shiddiky, M.J.A. Separation of distinct exosome subpopulations: Isolation and characterization approaches and their associated challenges. Analyst 2021, 146, 3731–3749. [Google Scholar] [CrossRef]
- Maisano, D.; Mimmi, S.; Dattilo, V.; Marino, F.; Gentile, M.; Vecchio, E.; Fiume, G.; Nistico, N.; Aloisio, A.; de Santo, M.P.; et al. A novel phage display based platform for exosome diversity characterization. Nanoscale 2022, 14, 2998–3003. [Google Scholar] [CrossRef]
- Janiak-Kiszka, J.; Nowaczewska, M.; Kazmierczak, W. Oral squamous cell carcinoma—Clinical characteristics, treatment, and outcomes in a single institution retrospective cohort study. Otolaryngol. Pol. 2022, 76, 12–17. [Google Scholar] [CrossRef]
- Przybylski, K.; Majchrzak, E.; Weselik, L.; Golusinski, W. Immunotherapy of head and neck squamous cell carcinoma (HNSCC). Immune checkpoint blockade. Otolaryngol. Pol. 2018, 72, 10–16. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Database | Number of Results | Search Strategy |
|---|---|---|
| Scopus | 428 | TITLE-ABS-KEY ((extracellular vesicle *) OR (multivesicular bodies) OR (exosome *)) AND ((squamous cell carcinoma *) AND ((head and neck) OR (larynx) OR (laryngeal) OR (nasal cavity) OR (oral) OR (pharyngeal) OR (hypopharyngeal) OR (nasopharyngeal) OR (oropharyngeal) OR (mouth))) AND ((delivery system *) AND (drug *)) Limited to: Article |
| Web of Science | 6 | AB = (((extracellular vesicle *) OR (multivesicular bodies) OR (exosome *)) AND ((squamous cell carcinoma *) AND ((head and neck) OR (larynx) OR (laryngeal) OR (nasal cavity) OR (oral) OR (pharyngeal) OR (hypopharyngeal) OR (nasopharyngeal) OR (oropharyngeal) OR (mouth))) AND ((delivery system *) AND (drug *))) |
| Pubmed | 2 | ((“Extracellular Vesicles”[Mesh]) OR (“Multivesicular Bodies”[Mesh]) OR (“Exosomes”[Mesh)) AND (“Squamous Cell Carcinoma of Head and Neck”[Mesh]) AND (“Drug Delivery Systems”[Mesh]) |
| Cochrane | 0 | #1 MeSH descriptor: [Extracellular Vesicles] explode all trees #2 MeSH descriptor: [Multivesicular Bodies] explode all trees #3 MeSH descriptor: [Exosomes] explode all trees #4 MeSH descriptor: [Drug Delivery Systems] explode all trees #5 MeSH descriptor: [Squamous Cell Carcinoma of Head and Neck] explode all trees #6 (#1 OR #2 OR #3) AND #4 AND #5 |
| Type of Study | Type of EVs | EVs Cargo | Main Results | |
|---|---|---|---|---|
| Cohen [12] | in vitro–A431 cell line in vivo–mice model (xenograft HNSCC) | Human MSC-EVs, A431-EVs | GNPs as labeling | 1. The EVs origin has a direct impact on the tumor targeting and penetration ability. 2. MSC-EVs had superior tumor accumulation when compared to A431-EVs. 3. The use of biocompatible GNP-labeled EVs, combined with CT imaging, has the potential for application in cancer therapy. |
| Cui [13] | in vitro–HSC-3 cell line in vivo–mice model (xenograft TSCC) | NTECs-EVs | miR-200c | 1. HSC-3DR cells showed downregulation of miR-200c. 2. The level of miR-200c was lower in TSCC lines relative to NTECs. 3. MiR-200c delivered by EVs to carcinoma cells decreased DTX resistance by targeting TUBB3 and PPP2R1B both in vitro and in vivo. |
| Deng [14] | in vitro–HN6 cells | HEK293T c-EVs | miR-34a | MiR-34a-EVs led to significant inhibition of HN6 cell proliferation, migration, and invasion by down-regulating SATB2 expression. |
| Kase [15] | in vitro–cell lines (OSCC-derived) in vivo–mice model (xenograft OSCC) | Oct-EVs | siLCP1 | The silencing of LCP1 by siRNA-suppressed OSCC tumor. |
| Li [17] | in vitro–CAL27 in vivo–mice model (xenograft OSCC) | γδ Tcell-EVs | miR-138 | 1. MiR-138—γδ Tcell-EVs increased expansion and cytotoxicity of γδ Tcells resulting in inhibition of OSCC both in vitro and in vivo. 2. MiR-138—γδ Tcell-EVs were more effective than liposome transfected miR-138 and scramble-cargo γδ Tcell- EVs. |
| Li [16] | in vitro–UM-SCC083A, UPCI-SCC029B | M-EVs | CA-miR-144/451a | M-EVs/CA-miR-144/451a biomimetic system effectively reduced the migration, invasion, and viability of OSCC cells and was more effective than free miR-144/451a. |
| Liu [18] | in vitro–HUVEC in vivo–mice model (xenograft OSCC) | SHED-EVs | miR-100-5p miR-1246 | 1. SHED-EVs inhibited cell proliferation and migration and induced apoptosis in HUVECs. 2. SHED-EVs downregulated several angiogenesis-related factors, (VEGFA, MMP-9, and ANGPT) and inhibited micro-vascular formation in OSCC tumor. |
| Liu [19] | in vitro–SCC25 | MSCT-EVs | SNS032 | SNS032/TRAIL delivered by gelatin biomimetic system can effectively induce apoptosis of tumor cells, reduces the dosage of free drugs, and shows a high inhibitory effect on OSCC. |
| Qiu [20] | in vitro–SCC25 in vivo–mice model (xenograft OSCC) | MSCT-EVs | CTX/TRAIL | 1. The antitumor effect of MSCT-EVs/CTX was confirmed in vitro and in vivo. 2. MSCT-EVs/CTX inhibited cell proliferation and migration and induced apoptosis in SCC cell lines and mice models. |
| Sayyed [21] | ex vivo–3D tumor spheroids model of UPCI-SCC-131 in vivo–mice model (xenograft OSCC) | UPCI-SCC-131-EVs | miR-155 inhibitor | 1. Cisplatin-resistant OSCC tumors showed a more malignant phenotype and elevated miR-155 level. 2. Treatment with miR-155 inhibitor-loaded EVs caused chemosensitization toward cisplatin via upregulation of FOXO3a and reducing EMT. |
| Tong [22] | human HNSCC cell lines: SCC90, SCC47, SCC104, SAS, CAL33 HPV- cell line CAL27 | HPV + HNSCC-EVs | miR-9-5p | 1. HPV + HNSCC-EVs were rich in miR-9-5p. 2. HPV + HNSCC-EVs miR-9 increased the radiosensitivity of HNSCC cells by polarizing macrophages into the M1 phenotypes which produce iNOS. |
| Wang [23] | in vitro–FaDu cells | HEK293T cells-EVs | TRPP2 siRNA | EV/TRPP2 siRNA complexes reduced EMT in FaDu cells and inhibited FaDu cell migration and invasion. |
| Wang [24] | in vitro–EBV-negative and positive NPC cell lines in vivo–NPC mice model | HUVEC–EVs | antagomiR-BART10-5p; antagomiR-18a | 1. IRGD-EVs-antagomiRs that contained both antagomiR-BART10-5p and antagomiR-18a attenuated the angiogenesis and growth of NPC with greater efficiency than a single treatment. 2. The findings established a synergistic role for virus and host miRs in the regulation of virus-associated tumor angiogenesis. |
| Wang [25] | in vitro–EBV-negative and positive NPC cell lines, radioresistant CSCs in vivo–NPC mice model | γδ TD-EVs | - | 1. γδ-T-EVs can effectively interact with and kill both EBV-positive and negative NPC cells. 2. The combination therapy of radiotherapy and γδ-T-EVs was more efficient because γδ-T-EVs improved radiosensitivity by eradicating the NPC CSCs. |
| Yakavets [26] | ex vivo–3D tumor spheroids model of PSCC (FaDu cells) | HUVEC–EVs | mTHPC | MTHPC—EVs most effectively improved the drug delivery to PSCC due to extremely high loading capacity. |
| Yamayoshi [27] | in vitro–CAL27 in vivo–hind model | CAL27-EVs | ExomiR-Tracker | ExomiR-Tracker successfully inhibited the function of miR-21 and the Cal27cells growth but also inhibited tumorigenesis in vivo. |
| Yang [28] | in vitro – CAL 27, WSU-HN6 ex vivo–3D spheroid-model of OSCC in vivo–mice model | BMEVs | 24 miRs identified in BMEVs | 1. BMEVs had a synergistic therapeutic effect of 5-FU against OSCC both in vitro and in vivo. 2. BMEVs significantly downregulated NLRP3 expression. 3. BMEVs suppressed OSCC proliferation and induced apoptosis by the generation of reactive oxygen species (ROS). |
| Zhang [29] | in vitro–HSC-3, SCC-9, CAL-27), HCM cell line in vivo–mice model | Milk- EVs | EV@Dox loaded with EPT1 and Ce6 | 1. EV@Dox–EPT1 caused significantly more cytotoxicity in cancer cells under 808 nm laser irradiation than free Dox. 2. NPs produced synergistic effects of photochemistry triggered by acid TME and NIR. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dżaman, K.; Czerwaty, K. Extracellular Vesicle-Based Drug Delivery Systems for Head and Neck Squamous Cell Carcinoma: A Systematic Review. Pharmaceutics 2023, 15, 1327. https://doi.org/10.3390/pharmaceutics15051327
Dżaman K, Czerwaty K. Extracellular Vesicle-Based Drug Delivery Systems for Head and Neck Squamous Cell Carcinoma: A Systematic Review. Pharmaceutics. 2023; 15(5):1327. https://doi.org/10.3390/pharmaceutics15051327
Chicago/Turabian StyleDżaman, Karolina, and Katarzyna Czerwaty. 2023. "Extracellular Vesicle-Based Drug Delivery Systems for Head and Neck Squamous Cell Carcinoma: A Systematic Review" Pharmaceutics 15, no. 5: 1327. https://doi.org/10.3390/pharmaceutics15051327