
Evaluation of In Vitro and In Vivo Antiviral Activities of Vitamin D for SARS-CoV-2 and Variants

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell lines and Viruses
2.2. Preparation of Compound Libraries for Primary Screen
2.3. Primary Screen with Pre-Treatment Using ACE2-Targeted Inhibitors and 57-Compound Natural Product Libraries
2.4. Primary Screen with Post-Treatment Using FDA-Approved Drugs and Flavonoids Library
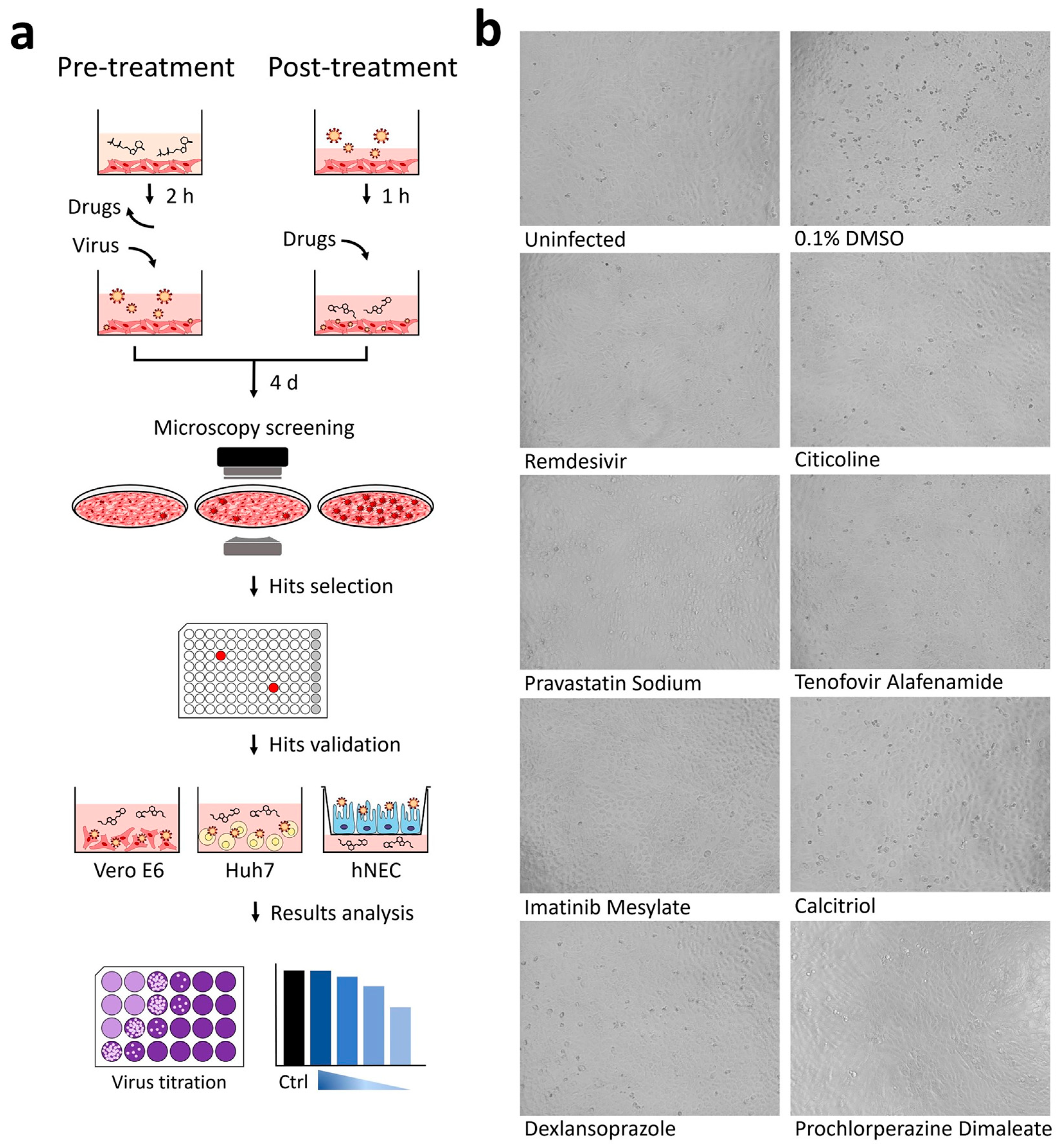
2.5. Selection of Hits via Analysis of Cytopathic Effects (CPE)
2.6. Validation of Hits
- Pre-infection treatment with citicoline, pravastatin sodium, and tenofovir alafenamide at working concentrations of 0.01, 0.1, 1, 10, 20 µM was carried out for 2 h at 37 °C. The inhibitors were removed and washed twice with PBS prior to SARS-CoV-2 infection at an MOI of 1. After infection, unbound viruses were removed by washing twice with PBS and replaced with DMEM media.
- Post-infection treatment was performed for imatinib mesylate, calcitriol, dexlansoprazole, and prochlorperazine dimaleate. To this end, Vero E6 and HuH7 cells were first infected with SARS-CoV-2 at an MOI of 1 for 1 h at 37 °C and then incubated with the compound-containing DMEM media at working concentrations of 0.01, 0.1, 1, 10, 20 µM.
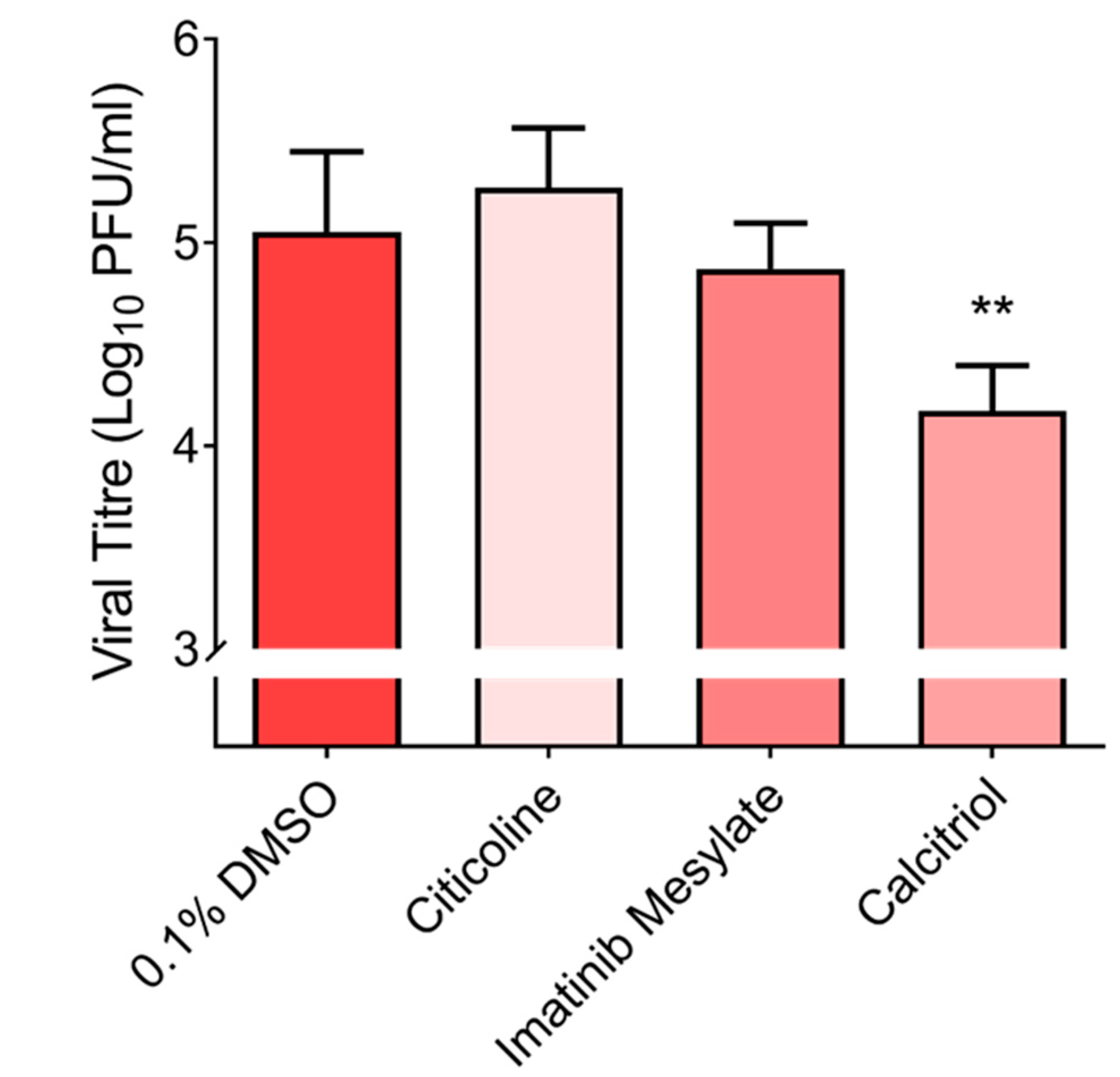
2.7. Validations of Imatinib Mesylate, Calcitrol and Citicoline with Primary Cell Line, hNEC at 10 µM
2.8. Plaque Assay
2.9. Gene Expression Levels of Vitamin D Receptor (VDR), 24-Hydroxylase (24(OH)ase) and Anti-Microbial Protein Cathelicidin (LL-37) by Real-Time Quantitative Polymerase Chain Reaction (RT qPCR)
2.10. Protein Expression and Analysis
2.10.1. Protein Extraction
2.10.2. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE) and Western Blot Analysis
2.11. Animals and Treatment Regimen
2.12. Statistical Analyses
3. Results
3.1. Cytopathic Effect (CPE) Based Screening of Compound Libraries
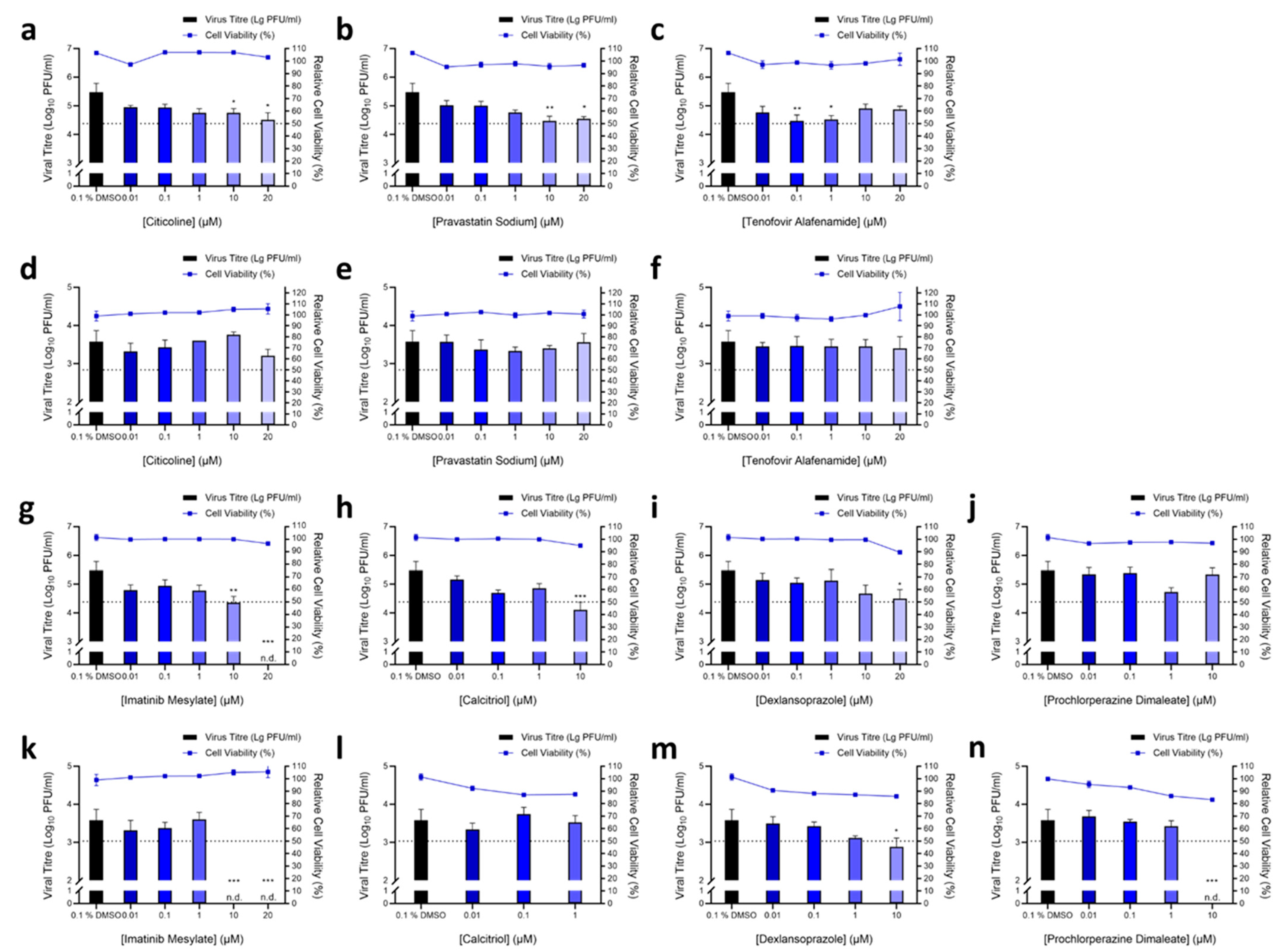
3.2. Validation of Hit Compounds via Dose-Dependent and Cell Viability Assays
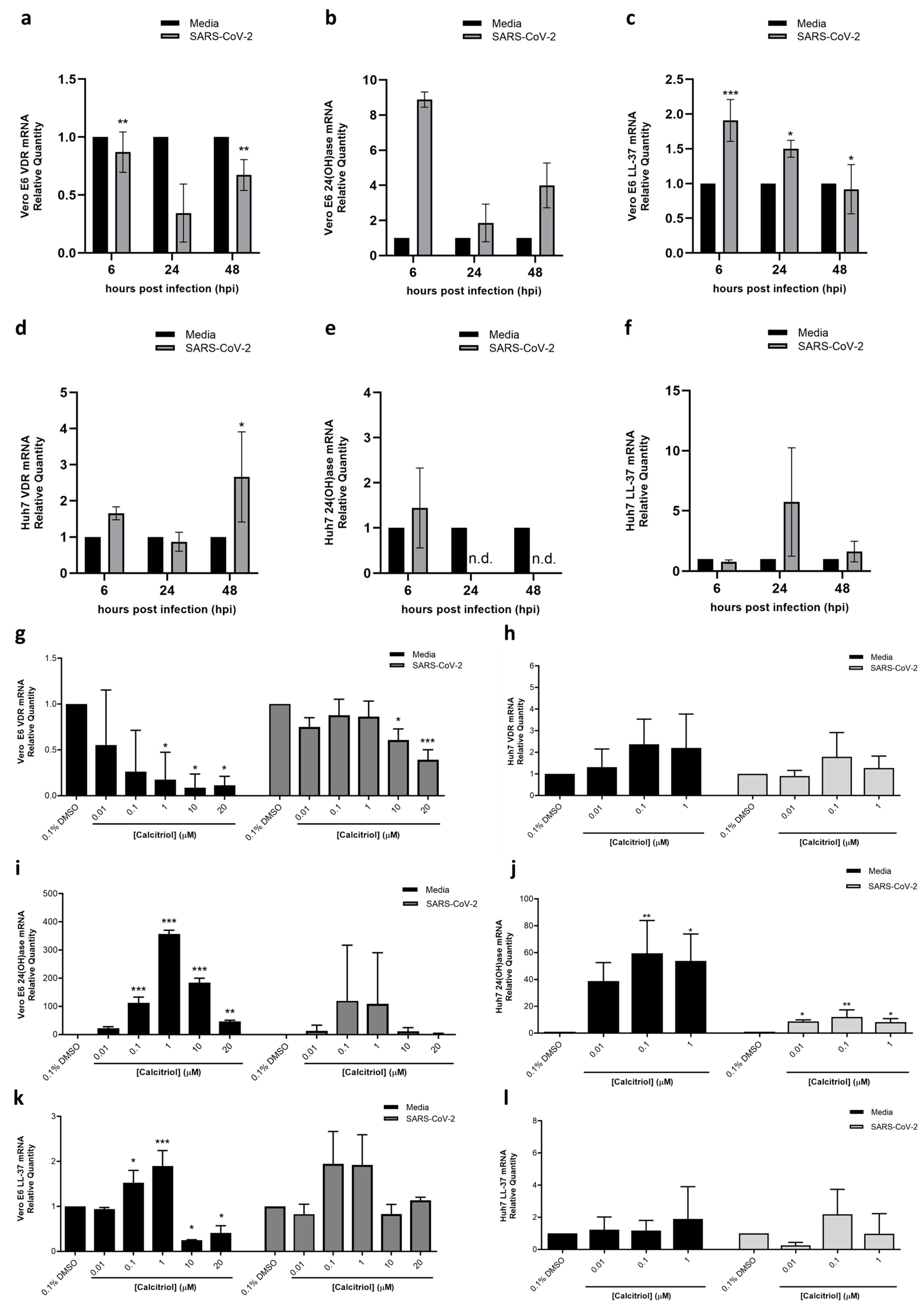
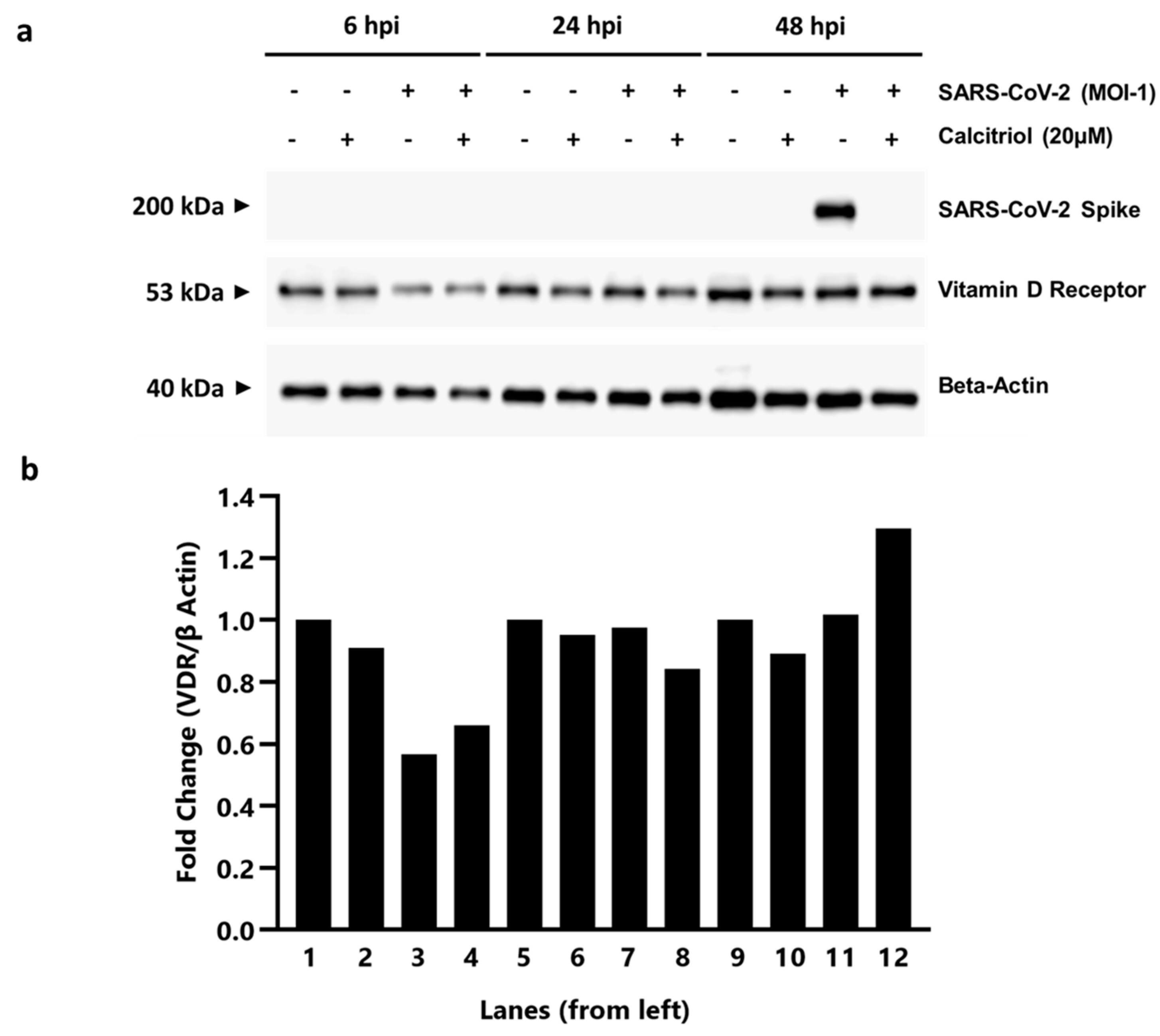
3.3. Elucidating the Antiviral Role of Calcitriol by RT qPCR
3.4. In Vivo Study of Calcitriol in K18-hACE2 Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gorbalenya, A.E.; Baker, S.C.; Baric, R.S.; de Groot, R.J.; Drosten, C.; Gulyaeva, A.A.; Haagmans, B.L.; Lauber, C.; Leontovich, A.M.; Neuman, B.W.; et al. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef] [Green Version]
- WHO. Rolling Updates on Coronavirus Disease (COVID-19). Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/events-as-they-happen (accessed on 8 June 2020).
- Fajgenbaum, D.C.; Khor, J.S.; Gorzewski, A.; Tamakloe, M.A.; Powers, V.; Kakkis, J.J.; Repasky, M.; Taylor, A.; Beschloss, A.; Hernandez-Miyares, L.; et al. Treatments Administered to the First 9152 Reported Cases of COVID-19: A Systematic Review. Infect. Dis. Ther. 2020, 9, 435–449. [Google Scholar] [CrossRef] [PubMed]
- Ahidjo, B.A.; Loe, M.; Ng, Y.L.; Mok, C.K.; Chu, J.J.H. Current Perspective of Antiviral Strategies against COVID-19. ACS Infect. Dis. 2020, 6, 1624–1634. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Tan, K.S.; Li, C.; Tran, T.; Chao, S.S.; Sugrue, R.J.; Shi, L.; Chow, V.T.; Wang, D.Y. Human nasal epithelial cells derived from multiple subjects exhibit differential responses to H3N2 influenza virus infection in vitro. J. Allergy. Clin. Immunol. 2016, 138, 276–281.e215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Yu, F.; Li, C.; Li, Y.; Chao, S.S.; Loh, W.S.; Pan, X.; Shi, L.; Wang, D.Y. The use of nasal epithelial stem/progenitor cells to produce functioning ciliated cells in vitro. Am. J. Rhinol. Allergy. 2012, 26, 345–350. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic. Acids. Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Dyall, J.; Coleman, C.M.; Hart, B.J.; Venkataraman, T.; Holbrook, M.R.; Kindrachuk, J.; Johnson, R.F.; Olinger, G.G., Jr.; Jahrling, P.B.; Laidlaw, M.; et al. Repurposing of clinically developed drugs for treatment of Middle East respiratory syndrome coronavirus infection. Antimicrob. Agents. Chemother. 2014, 58, 4885–4893. [Google Scholar] [CrossRef] [Green Version]
- Yan, D.; Liu, X.Y.; Zhu, Y.N.; Huang, L.; Dan, B.T.; Zhang, G.J.; Gao, Y.H. Factors associated with prolonged viral shedding and impact of Lopinavir/Ritonavir treatment in hospitalised non-critically ill patients with SARS-CoV-2 infection. Eur. Respir. J. 2020, 56, 2000799. [Google Scholar] [CrossRef]
- Barnard, D.L.; Hubbard, V.D.; Burton, J.; Smee, D.F.; Morrey, J.D.; Otto, M.J.; Sidwell, R.W. Inhibition of severe acute respiratory syndrome-associated coronavirus (SARSCoV) by calpain inhibitors and beta-D-N4-hydroxycytidine. Antivir. Chem. Chemother. 2004, 15, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Nie, Y.; Wang, P.; Shi, X.; Wang, G.; Chen, J.; Zheng, A.; Wang, W.; Wang, Z.; Qu, X.; Luo, M.; et al. Highly infectious SARS-CoV pseudotyped virus reveals the cell tropism and its correlation with receptor expression. Biochem. Biophys. Res. Commun. 2004, 321, 994–1000. [Google Scholar] [CrossRef] [PubMed]
- Freymuth, F.; Vabret, A.; Rozenberg, F.; Dina, J.; Petitjean, J.; Gouarin, S.; Legrand, L.; Corbet, S.; Brouard, J.; Lebon, P. Replication of respiratory viruses, particularly influenza virus, rhinovirus, and coronavirus in HuH7 hepatocarcinoma cell line. J. Med. Virol. 2005, 77, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Sungnak, W.; Huang, N.; Becavin, C.; Berg, M.; Queen, R.; Litvinukova, M.; Talavera-Lopez, C.; Maatz, H.; Reichart, D.; Sampaziotis, F.; et al. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat. Med. 2020, 26, 681–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilie, P.C.; Stefanescu, S.; Smith, L. The role of vitamin D in the prevention of coronavirus disease 2019 infection and mortality. Aging Clin. Exp. Res. 2020, 32, 1195–1198. [Google Scholar] [CrossRef]
- Grant, W.B.; Lahore, H.; McDonnell, S.L.; Baggerly, C.A.; French, C.B.; Aliano, J.L.; Bhattoa, H.P. Evidence that Vitamin D Supplementation Could Reduce Risk of Influenza and COVID-19 Infections and Deaths. Nutrients 2020, 12, 988. [Google Scholar] [CrossRef] [Green Version]
- Bikle, D.D. Vitamin D: Production, metabolism and mechanisms of action. In Endotext; Feingold, K.R., Anawalt, B., Blackman, M.R., Boyce, A., Chrousos, G., Corpas, E., de Herder, W.W., Dhatariya, K., Hofland, J., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Andrault, P.M.; Samsonov, S.A.; Weber, G.; Coquet, L.; Nazmi, K.; Bolscher, J.G.; Lalmanach, A.C.; Jouenne, T.; Bromme, D.; Pisabarro, M.T.; et al. Antimicrobial Peptide LL-37 Is Both a Substrate of Cathepsins S and K and a Selective Inhibitor of Cathepsin L. Biochemistry 2015, 54, 2785–2798. [Google Scholar] [CrossRef]
- Simmons, G.; Gosalia, D.N.; Rennekamp, A.J.; Reeves, J.D.; Diamond, S.L.; Bates, P. Inhibitors of cathepsin L prevent severe acute respiratory syndrome coronavirus entry. Proc. Natl. Acad. Sci. USA 2005, 102, 11876–11881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, I.C.; Bosch, B.J.; Li, F.; Li, W.; Lee, K.H.; Ghiran, S.; Vasilieva, N.; Dermody, T.S.; Harrison, S.C.; Dormitzer, P.R.; et al. SARS coronavirus, but not human coronavirus NL63, utilizes cathepsin L to infect ACE2-expressing cells. J. Biol. Chem. 2006, 281, 3198–3203. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.; Prosser, D.E.; Kaufmann, M. Cytochrome P450-mediated metabolism of vitamin D. J. Lipid. Res. 2014, 55, 13–31. [Google Scholar] [CrossRef] [Green Version]
- Healy, K.D.; Zella, J.B.; Prahl, J.M.; DeLuca, H.F. Regulation of the murine renal vitamin D receptor by 1,25-dihydroxyvitamin D3 and calcium. Proc. Natl. Acad. Sci. USA 2003, 100, 9733–9737. [Google Scholar] [CrossRef] [Green Version]
- Uhmann, A.; Niemann, H.; Lammering, B.; Henkel, C.; Hess, I.; Rosenberger, A.; Dullin, C.; Schraepler, A.; Schulz-Schaeffer, W.; Hahn, H. Calcitriol inhibits hedgehog signaling and induces vitamin d receptor signaling and differentiation in the patched mouse model of embryonal rhabdomyosarcoma. Sarcoma 2012, 2012, 357040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, I.; Han, G.; Seshadri, M.; Gillard, B.M.; Yu, W.D.; Foster, B.A.; Trump, D.L.; Johnson, C.S. Role of vitamin D receptor in the antiproliferative effects of calcitriol in tumor-derived endothelial cells and tumor angiogenesis in vivo. Cancer Res. 2009, 69, 967–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muindi, J.R.; Modzelewski, R.A.; Peng, Y.; Trump, D.L.; Johnson, C.S. Pharmacokinetics of 1α,25-Dihydroxyvitamin D<sub>3</sub> in Normal Mice after Systemic Exposure to Effective and Safe Antitumor Doses. Oncology 2004, 66, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Tukaj, S.; Bieber, K.; Witte, M.; Ghorbanalipoor, S.; Schmidt, E.; Zillikens, D.; Ludwig, R.J.; Kasperkiewicz, M. Calcitriol Treatment Ameliorates Inflammation and Blistering in Mouse Models of Epidermolysis Bullosa Acquisita. J. Investig. Dermatol. 2018, 138, 301–309. [Google Scholar] [CrossRef] [Green Version]
- Gui, B.; Chen, Q.; Hu, C.; Zhu, C.; He, G. Effects of calcitriol (1, 25-dihydroxy-vitamin D3) on the inflammatory response induced by H9N2 influenza virus infection in human lung A549 epithelial cells and in mice. Virol. J. 2017, 14, 10. [Google Scholar] [CrossRef] [Green Version]
- ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/results/browse?cond=COVID-19&term=&cntry=&state=&city=&dist=&Search=Search&brwse=cond_alpha_all&type=Intr (accessed on 21 April 2020).
- Tu, Y.F.; Chien, C.S.; Yarmishyn, A.A.; Lin, Y.Y.; Luo, Y.H.; Lin, Y.T.; Lai, W.Y.; Yang, D.M.; Chou, S.J.; Yang, Y.P.; et al. A Review of SARS-CoV-2 and the Ongoing Clinical Trials. Int. J. Mol. Sci. 2020, 21, 2657. [Google Scholar] [CrossRef] [Green Version]
- WHO. WHO Consolidated Guidelines on Tuberculosis: Module 1: Prevention: Tuberculosis Preventive Treatment. 2020. Available online: http://www.who.int/publications/i/item/9789240001503 (accessed on 7 February 2022).
- Balicer, R.D.; Huerta, M.; Grotto, I. Tackling the next influenza pandemic. BMJ 2004, 328, 1391–1392. [Google Scholar] [CrossRef] [PubMed]
- van Genugten, M.L.; Heijnen, M.L.; Jager, J.C. Pandemic influenza and healthcare demand in the Netherlands: Scenario analysis. Emerg. Infect. Dis. 2003, 9, 531–538. [Google Scholar] [CrossRef]
- Gombart, A.F. The vitamin D-antimicrobial peptide pathway and its role in protection against infection. Future Microbiol. 2009, 4, 1151–1165. [Google Scholar] [CrossRef] [Green Version]
- Pislar, A.; Mitrovic, A.; Sabotic, J.; Pecar Fonovic, U.; Perisic Nanut, M.; Jakos, T.; Senjor, E.; Kos, J. The role of cysteine peptidases in coronavirus cell entry and replication: The therapeutic potential of cathepsin inhibitors. PLoS Pathog. 2020, 16, e1009013. [Google Scholar] [CrossRef]
- Zhao, M.M.; Yang, W.L.; Yang, F.Y.; Zhang, L.; Huang, W.J.; Hou, W.; Fan, C.F.; Jin, R.H.; Feng, Y.M.; Wang, Y.C.; et al. Cathepsin L plays a key role in SARS-CoV-2 infection in humans and humanized mice and is a promising target for new drug development. Signal Transduct. Target. Ther. 2021, 6, 134. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, S.; Li, D.; Chen, P.; Han, S.; Zhao, G.; Chen, Y.; Zhao, J.; Xiong, J.; Qiu, J.; et al. Human Cathelicidin Inhibits SARS-CoV-2 Infection: Killing Two Birds with One Stone. ACS Infect. Dis. 2021, 7, 1545–1554. [Google Scholar] [CrossRef] [PubMed]
- Gombart, A.F.; Borregaard, N.; Koeffler, H.P. Human cathelicidin antimicrobial peptide (CAMP) gene is a direct target of the vitamin D receptor and is strongly up-regulated in myeloid cells by 1,25-dihydroxyvitamin D3. FASEB J. 2005, 19, 1067–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pharmaceutical Class | No. of Compounds with Activity against SARS-CoV-2 | Total No. of Compounds for Each Class | ||
|---|---|---|---|---|
| FDA-Approved Drugs | ACE2 Inhibitors | Flavonoids | ||
| Antiviral agents | 3 | 2 | 5 | |
| Antibacterial agents | 9 | 4 | 13 | |
| Antifungal agents | 3 | 3 | ||
| Antiparasitic agents | 2 | 2 | ||
| Anticancer agents | 3 | 3 | 6 | |
| Antihistamines | 5 | 5 | ||
| Antihypertensives | 1 | 1 | 2 | |
| Antiarrhythmic | 1 | 1 | ||
| Anticoagulants | 1 | 1 | ||
| Antispasmodic | 1 | 1 | ||
| Proton pump inhibitors | 2 | 2 | ||
| Ion channel blockers | 3 | 3 | ||
| Non-steroidal anti-inflammatory drugs | 3 | 3 | ||
| Anti-inflammatory agents | 2 | 4 | 6 | |
| Phosphodiesterase inhibitors | 1 | 1 | ||
| Lipid, sterol metabolism inhibitors | 1 | 1 | 2 | |
| Signalling kinase inhibitors | 2 | 2 | 4 | |
| Neurotransmitter inhibitors | 3 | 3 | ||
| Nucleic acid synthesis inhibitors | 2 | 2 | ||
| Histone deacetylase inhibitors | 1 | 1 | ||
| Flavonoids | 1 | 34 | 35 | |
| Others | 10 | 10 | 20 | |
| Total | 56 | 31 | 34 | 121 |
| Compound | Chemical Structure | Pharmaceutical Class | Known Mechanism | Indication | FDA-Approved |
|---|---|---|---|---|---|
| Citicoline |  | Others | Intermediate in the synthesis of Phospatidylcholine | Proposed for use in traumatic brain injuries, stroke and vascular dementia due to potential neuroprotective effects | No |
| Pravastatin Sodium |  | Lipid, sterol metabolism Inhibitor | Inhibits HMG-CoA reductase, thus inhibiting the synthesis of cholesterol | Prevention of cardiovascular disease and treatment of hypercholesterolemia | Yes |
| Tenofovir Alafenamide |  | Antiviral | Nucleoside inhibitor of viral reverse transcriptase | Prodrug form of tenofovir, used for treatment of HIV and chronic Hepatitis B infections | Yes |
| Imatinib Mesylate |  | Signalling kinase inhibitor | Inhibits Bcr-Abl tyrosine kinase | Treatment of chronic myelogenous leukemia, gastrointestinal stromal tumors and various other cancers | Yes |
| Calcitriol |  | Others | Active metabolite of vitamin D3 | Treatment of secondary hyperparathyroidism and metabolic bone disease, hypocalcemia, osteoporosis | Yes |
| Dexlansoprazole |  | Proton pump Inhibitor | Inhibits H+/K+ ATPase, resulting in decreased secretion of HCl into the gastric lumen | Treatment and management of gastroesophageal reflux, erosive esophagitis | Yes |
| Prochlorperazine Dimaleate |  | Neurotransmitter inhibitor | Blocks D2 dopamine receptors in the brain | Treatment of severe nausea and vomiting, as well as short-term management of anxiety and schizophrenia | Yes |
| Compound | CC50 (μM) | IC50 (μM) | Selectivity Index |
|---|---|---|---|
| Vero E6 Cells | |||
| Citicoline | >20 | - | - |
| Pravastatin Sodium | >20 | <0.01 | >2000 |
| Tenofovir Alafenamide | >20 | - | - |
| Imatinib Mesylate | >20 | <0.01 | >2000 |
| Calcitriol | >20 | <0.01 | >2000 |
| Dexlansoprazole | >20 | <0.01 | >2000 |
| Prochlorperazine Dimaleate | >20 | - | - |
| HuH7 Cells | |||
| Citicoline | >20 | - | - |
| Pravastatin Sodium | >20 | - | - |
| Tenofovir Alafenamide | >20 | - | - |
| Imatinib Mesylate | >20 | - | - |
| Calcitriol | 4.692 | - | - |
| Dexlansoprazole | >20 | 2.479 | >8.0678 |
| Prochlorperazine Dimaleate | 15.73 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mok, C.-K.; Ng, Y.L.; Ahidjo, B.A.; Aw, Z.Q.; Chen, H.; Wong, Y.H.; Lee, R.C.H.; Loe, M.W.C.; Liu, J.; Tan, K.S.; et al. Evaluation of In Vitro and In Vivo Antiviral Activities of Vitamin D for SARS-CoV-2 and Variants. Pharmaceutics 2023, 15, 925. https://doi.org/10.3390/pharmaceutics15030925
Mok C-K, Ng YL, Ahidjo BA, Aw ZQ, Chen H, Wong YH, Lee RCH, Loe MWC, Liu J, Tan KS, et al. Evaluation of In Vitro and In Vivo Antiviral Activities of Vitamin D for SARS-CoV-2 and Variants. Pharmaceutics. 2023; 15(3):925. https://doi.org/10.3390/pharmaceutics15030925
Chicago/Turabian StyleMok, Chee-Keng, Yan Ling Ng, Bintou Ahmadou Ahidjo, Zhen Qin Aw, Huixin Chen, Yi Hao Wong, Regina Ching Hua Lee, Marcus Wing Choy Loe, Jing Liu, Kai Sen Tan, and et al. 2023. "Evaluation of In Vitro and In Vivo Antiviral Activities of Vitamin D for SARS-CoV-2 and Variants" Pharmaceutics 15, no. 3: 925. https://doi.org/10.3390/pharmaceutics15030925