
Protective Efficacy of a Mucosal Influenza Vaccine Formulation Based on the Recombinant Nucleoprotein Co-Administered with a TLR2/6 Agonist BPPcysMPEG
 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- (i)
- (ii)
- (iii)
2. Material and Methods
2.1. Vaccine Design
2.2. Mice
2.3. Ethics Section
2.4. Influenza Challenge Studies
2.5. ELISA
2.6. ELISPOT
2.7. Proliferation Assay
2.8. Flow Cytometric Analysis of Multifunctional T Cells and Cytokine Profiling
2.9. Multiplex FlowCytomix (Cytometric Bead Array)
2.10. Statistical Analysis
3. Results
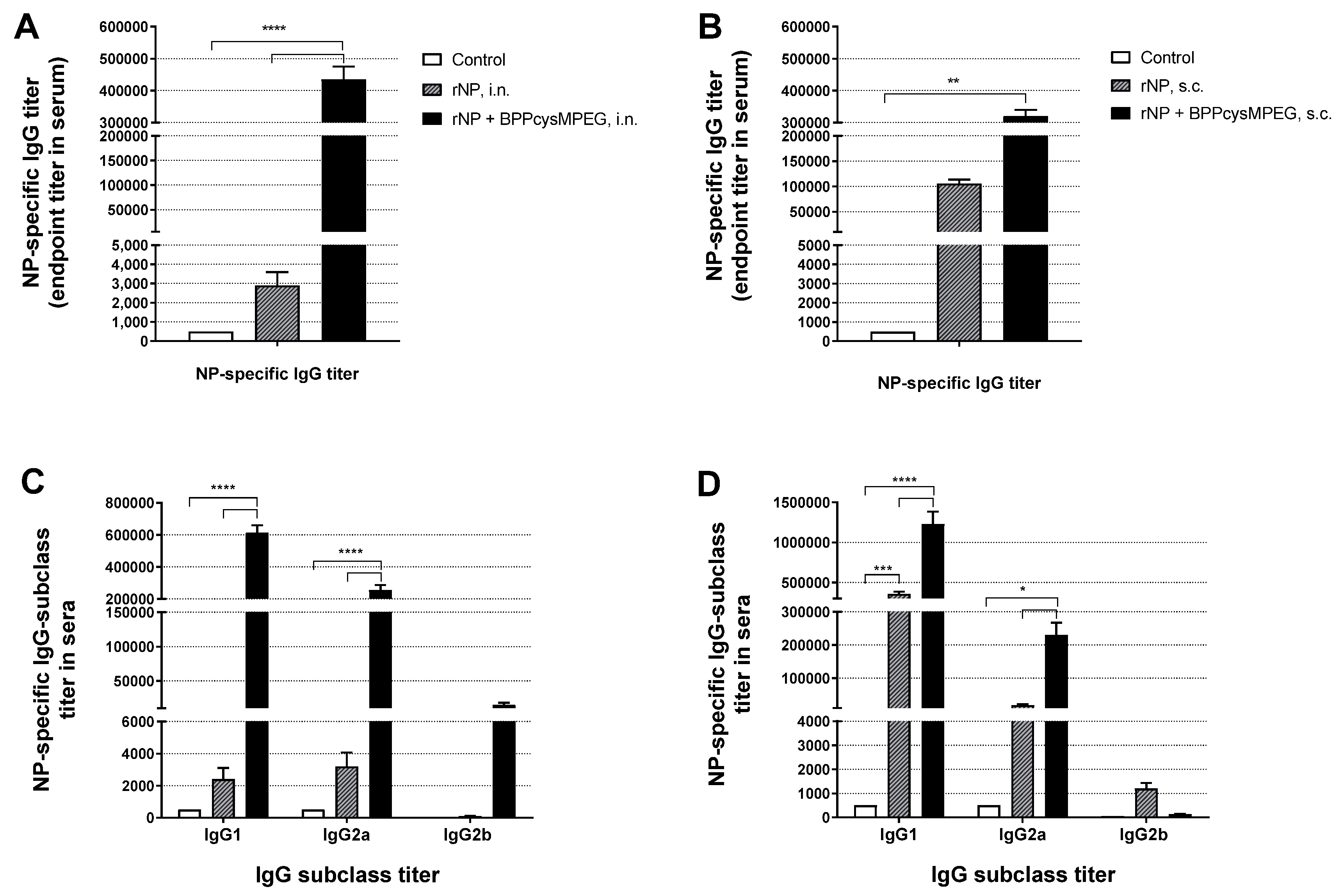
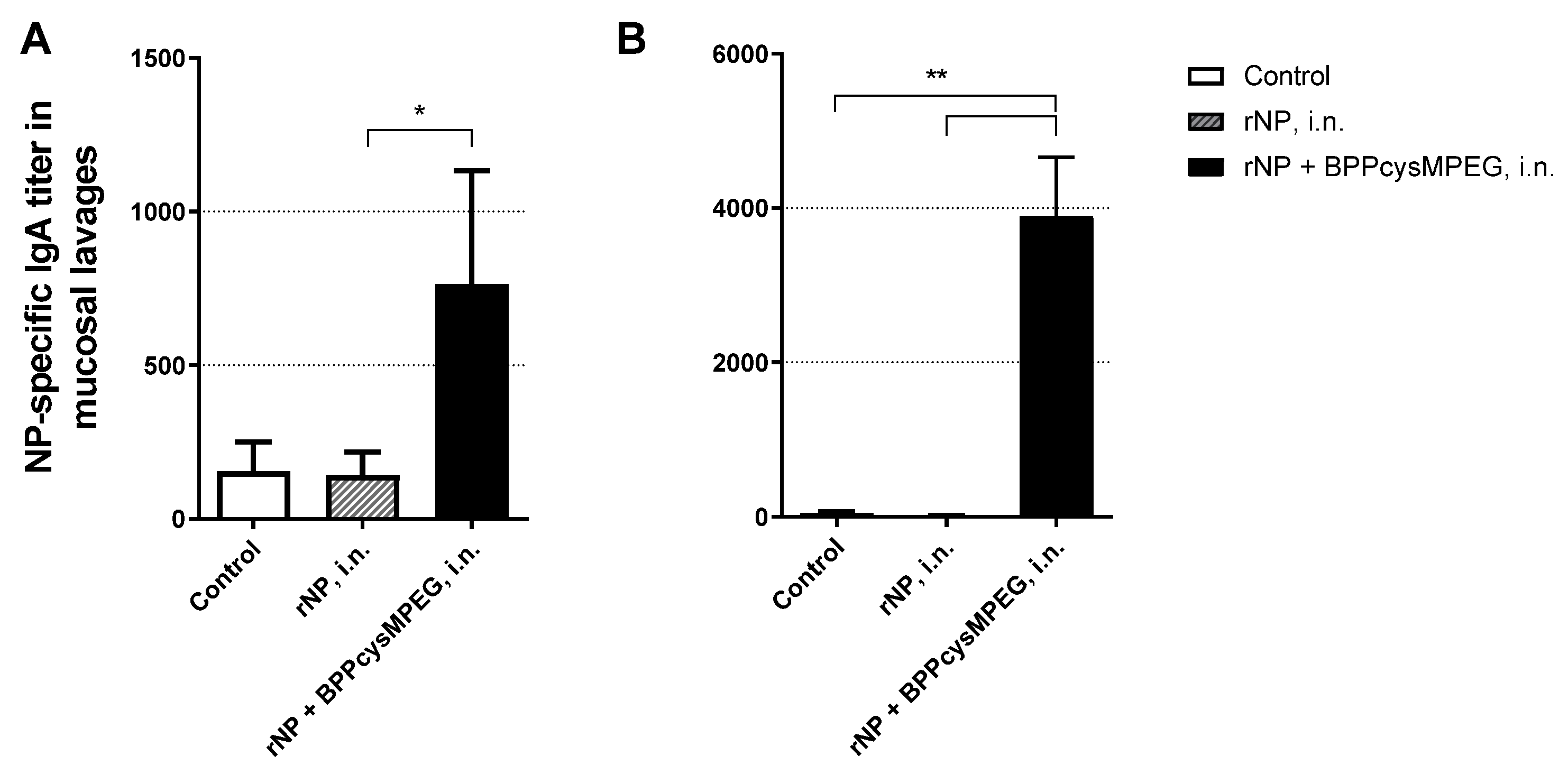
3.1. Mucosal Administration of rNP with BPPcysMPEG Stimulates Strong NP-Specific Systemic and Local Humoral Immune Responses
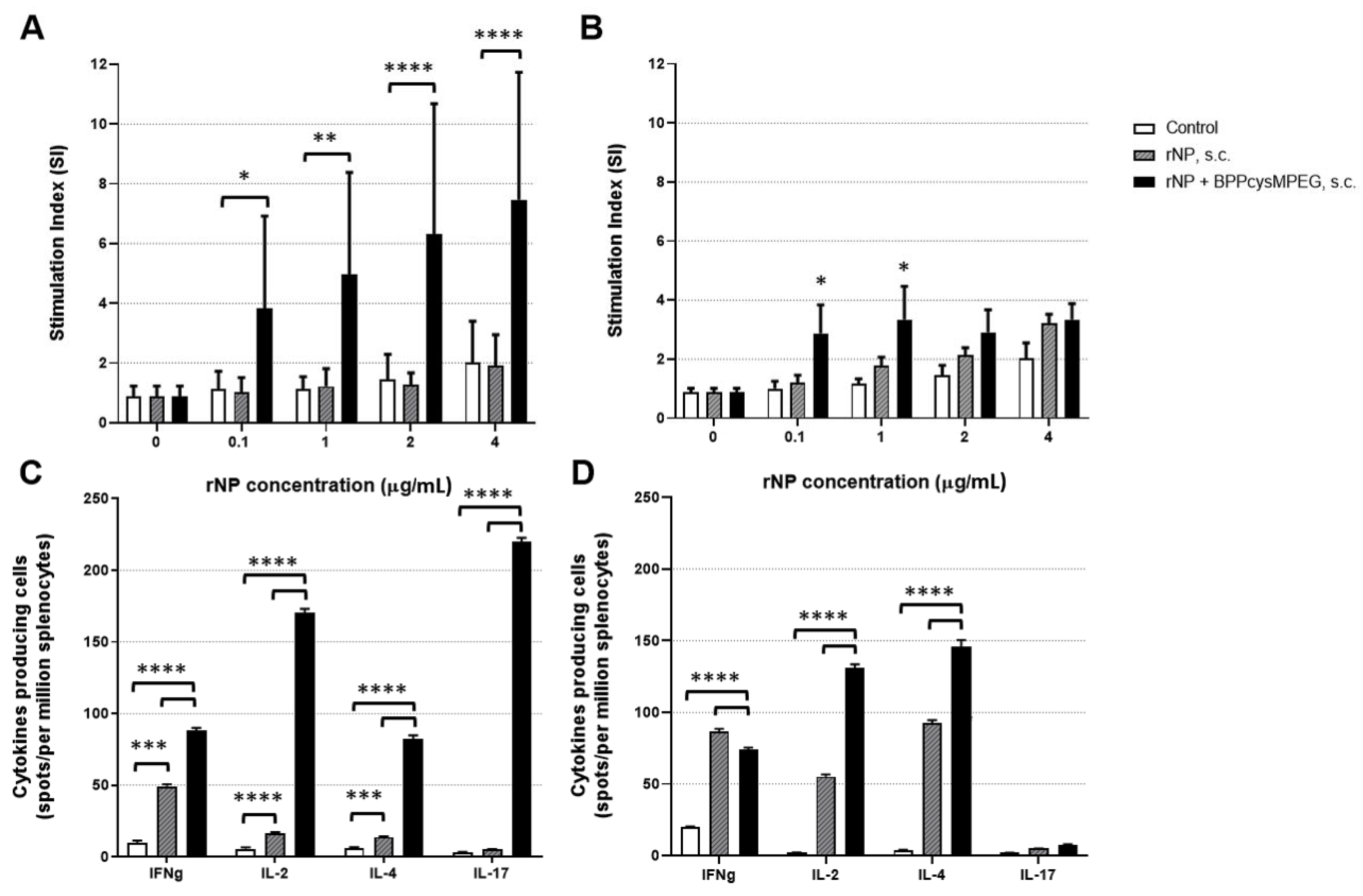
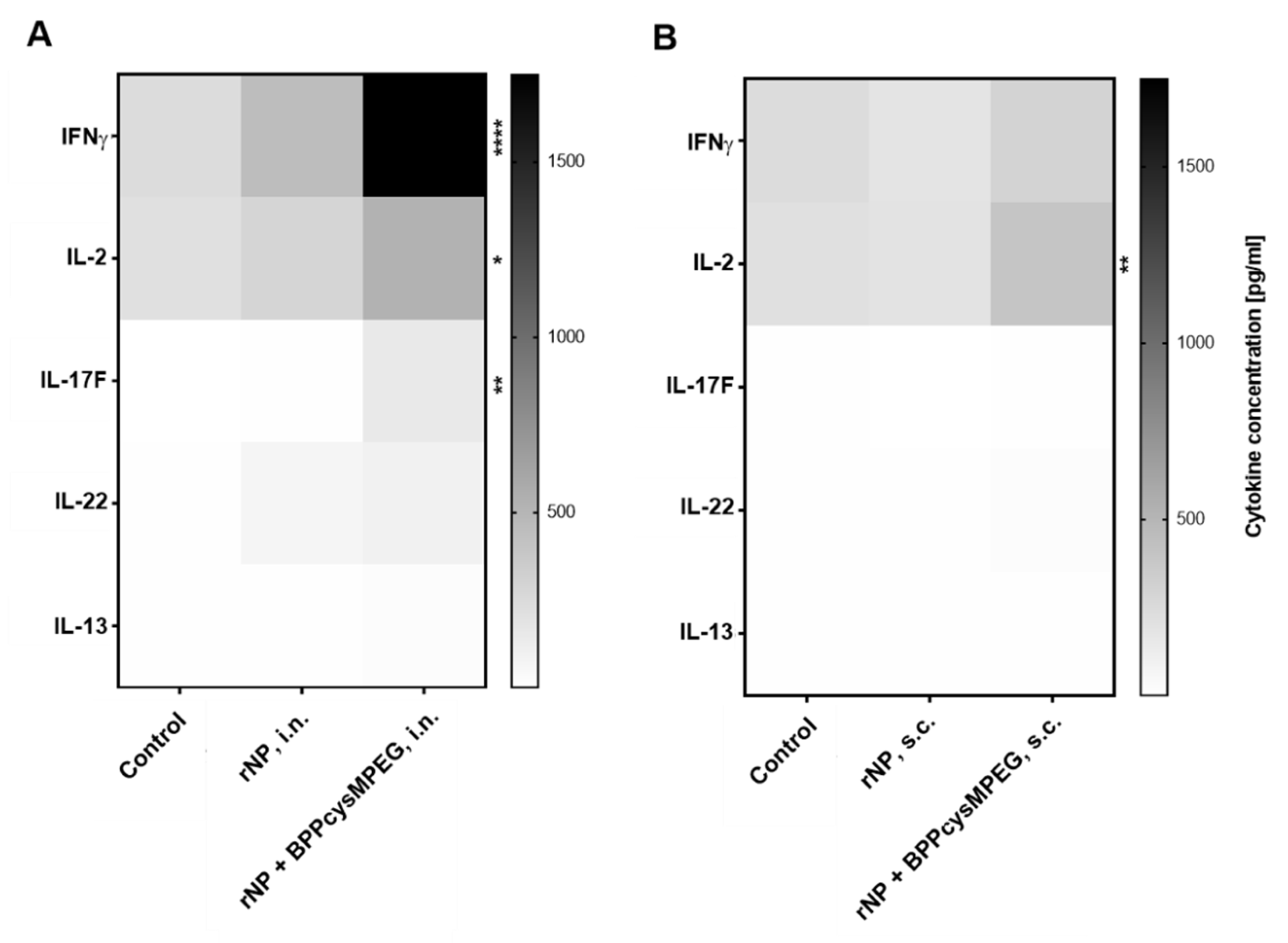
3.2. Mucosal Administration of rNP with BPPcysMPEG Promotes Strong Th1/Th2/Th17 Cellular Immune Responses
3.3. Mucosal Vaccination of Mice with rNP Co-Administered with BPPcysMPEG Enhances the Quality of Antigen-Specific Cellular Response by Stimulating Multifunctional CD4+ T Cells
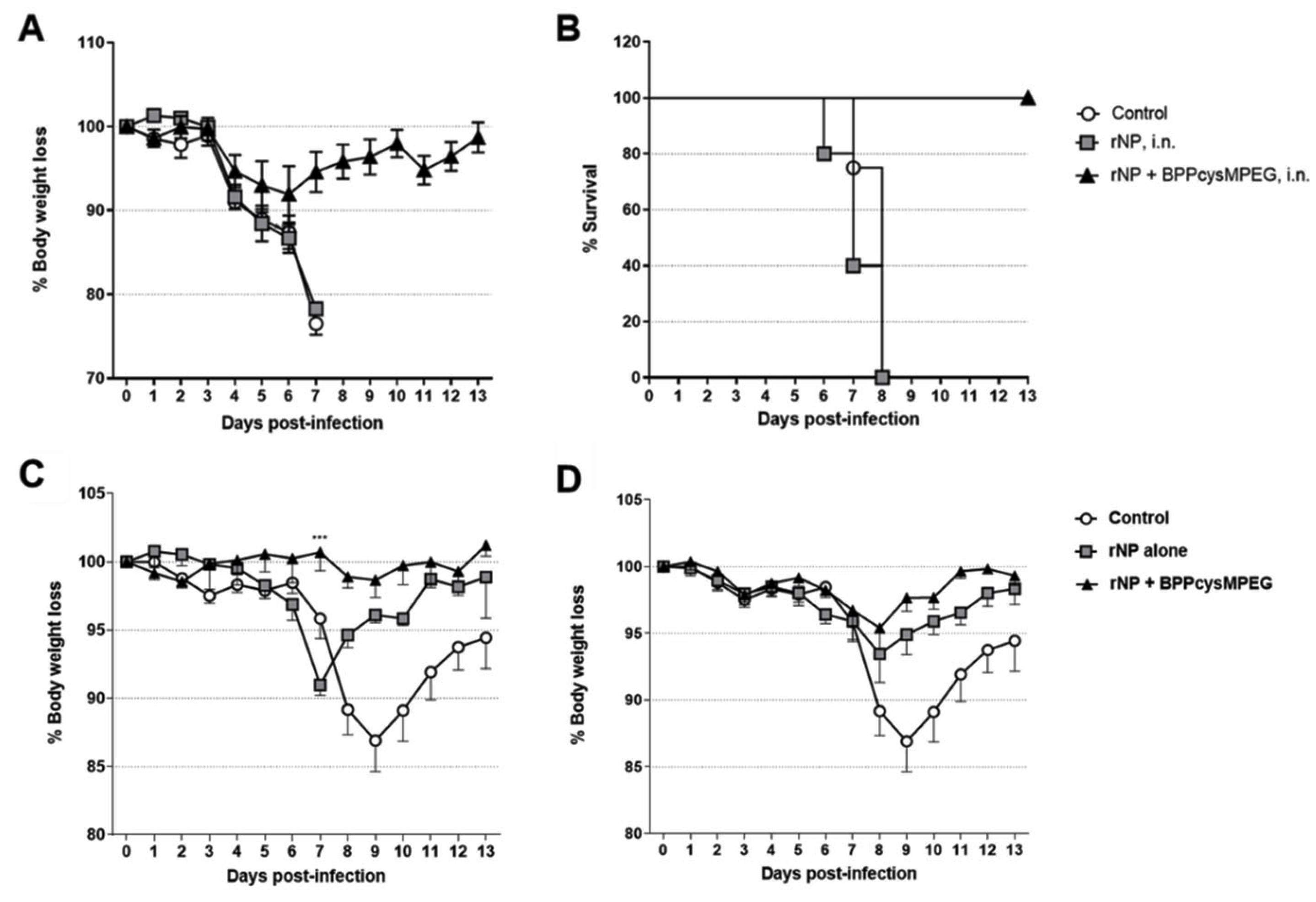
3.4. Mucosal Vaccination of Mice with rNP plus BPPcysMPEG Confers Protection against Influenza Infection
4. Discussion
5. Patent
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jang, Y.H.; Seong, B.L. Options and Obstacles for Designing a Universal Influenza Vaccine. Viruses 2014, 6, 3159–3180. [Google Scholar] [CrossRef]
- Lee, L.Y.-H.; Ha, D.L.A.; Simmons, C.; de Jong, M.D.; Chau, N.V.V.; Schumacher, R.; Peng, Y.C.; McMichael, A.J.; Farrar, J.J.; Smith, G.L.; et al. Memory T cells established by seasonal human influenza A infection cross-react with avian influenza A (H5N1) in healthy individuals. J. Clin. Investig. 2008, 118, 3478–3490. [Google Scholar] [CrossRef] [Green Version]
- Brokstad, K.A.; Eriksson, J.-C.H.; Cox, R.J.; Tynning, T.L.; Olofsson, J.; Jonsson, R.; Davidsson, Å. Parenteral Vaccination against Influenza Does Not Induce a Local Antigen-Specific Immune Response in the Nasal Mucosa. J. Infect. Dis. 2002, 185, 878–884. [Google Scholar] [CrossRef] [Green Version]
- Sridhar, S.; Begom, S.; Bermingham, A.; Hoschler, K.; Adamson, W.; Carman, W.; Bean, T.; Barclay, W.; Deeks, J.J.; Lalvani, A. Cellular immune correlates of protection against symptomatic pandemic influenza. Nat. Med. 2013, 19, 1305–1312. [Google Scholar] [CrossRef]
- Saletti, G.; Gerlach, T.; Rimmelzwaan, G.F. Influenza vaccines: ‘tailor-made’ or ‘one fits all’. Curr. Opin. Immunol. 2018, 53, 102–110. [Google Scholar] [CrossRef]
- Osterholm, M.T.; Kelley, N.S.; Sommer, A.; Belongia, E.A. Efficacy and effectiveness of influenza vaccines: A systematic review and meta-analysis. Lancet Infect. Dis. 2012, 12, 36–44. [Google Scholar] [CrossRef]
- O’Hagan, D.; Ott, G.; De Gregorio, E.; Seubert, A. The mechanism of action of MF59—An innately attractive adjuvant formulation. Vaccine 2012, 30, 4341–4348. [Google Scholar] [CrossRef]
- Scheifele, D.W.; Ward, B.J.; Dionne, M.; Vanderkooi, O.G.; Loeb, M.; Coleman, B.L.; Li, Y. Compatibility of ASO3-adjuvanted H1N1pdm09 and seasonal trivalent influenza vaccines in adults: Results of a randomized, controlled trial. Vaccine 2012, 30, 4728–4732. [Google Scholar] [CrossRef]
- Ainai, A.; Suzuki, T.; Tamura, S.-I.; Hasegawa, H. Intranasal Administration of Whole Inactivated Influenza Virus Vaccine as a Promising Influenza Vaccine Candidate. Viral Immunol. 2017, 30, 451–462. [Google Scholar] [CrossRef]
- Tamura, S.-I.; Ainai, A.; Suzuki, T.; Kurata, T.; Hasegawa, H. Intranasal Inactivated Influenza Vaccines: A Reasonable Approach to Improve the Efficacy of Influenza Vaccine? Jpn. J. Infect. Dis. 2016, 69, 165–179. [Google Scholar] [CrossRef] [Green Version]
- Kaur, A.; Piplani, S.; Kaushik, D.; Fung, J.; Sakala, I.G.; Honda-Okubo, Y.; Mehta, S.K.; Petrovsky, N.; Salunke, D.B. Stereoisomeric Pam2CS based TLR2 agonists: Synthesis, structural modelling and activity as vaccine adjuvants. RSC Med. Chem. 2022, 13, 622–637. [Google Scholar] [CrossRef]
- Kaur, A.; Kanwar, R.; Kaushik, D.; Sakala, I.G.; Honda-Okubo, Y.; Petrovsky, N.; Salunke, D.B.; Mehta, S.K. Combined delivery of TLR2 and TLR7 agonists by Nanostructured lipid carriers induces potent vaccine adjuvant activity in mice. Int. J. Pharm. 2022, 613, 121378. [Google Scholar] [CrossRef]
- Girkin, J.L.; Maltby, S.; Bartlett, N.W. Toll-like receptor-agonist-based therapies for respiratory viral diseases: Thinking outside the cell. Eur. Respir. Rev. 2022, 31, 210274. [Google Scholar] [CrossRef]
- Hossain, M.J.; Bourgeois, M.; Quan, F.-S.; Lipatov, A.S.; Song, J.-M.; Chen, L.-M.; Compans, R.W.; York, I.; Kang, S.-M.; Donis, R.O. Virus-Like Particle Vaccine Containing Hemagglutinin Confers Protection against 2009 H1N1 Pandemic Influenza. Clin. Vaccine Immunol. 2011, 18, 2010–2017. [Google Scholar] [CrossRef] [Green Version]
- Impagliazzo, A.; Milder, F.; Kuipers, H.; Wagner, M.V.; Zhu, X.; Hoffman, R.M.; Van Meersbergen, R.; Huizingh, J.; Wanningen, P.; Verspuij, J.; et al. A stable trimeric influenza hemagglutinin stem as a broadly protective immunogen. Science 2015, 349, 1301–1306. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, J.A.; Buonocore, L.; Suguitan, A.; Hunter, M.; Marx, P.A.; Subbarao, K.; Rose, J.K. Vesicular Stomatitis Virus-Based H5N1 Avian Influenza Vaccines Induce Potent Cross-Clade Neutralizing Antibodies in Rhesus Macaques. J. Virol. 2011, 85, 4602–4605. [Google Scholar] [CrossRef] [Green Version]
- Hessel, A.; Schwendinger, M.; Holzer, G.W.; Orlinger, K.K.; Coulibaly, S.; Savidis-Dacho, H.; Zips, M.-L.; Crowe, B.A.; Kreil, T.R.; Ehrlich, H.J.; et al. Vectors Based on Modified Vaccinia Ankara Expressing Influenza H5N1 Hemagglutinin Induce Substantial Cross-Clade Protective Immunity. PLoS ONE 2011, 6, e16247. [Google Scholar] [CrossRef]
- Steitz, J.; Barlow, P.G.; Hossain, J.; Kim, E.; Okada, K.; Kenniston, T.; Rea, S.; Donis, R.O.; Gambotto, A. A Candidate H1N1 Pandemic Influenza Vaccine Elicits Protective Immunity in Mice. PLoS ONE 2010, 5, e10492. [Google Scholar] [CrossRef] [Green Version]
- Haveri, A.; Ikonen, N.; Savolainen-Kopra, C.; Julkunen, I. Long-lasting heterologous antibody responses after sequential vaccination with A/Indonesia/5/2005 and A/Vietnam/1203/2004 pre-pandemic influenza A(H5N1) virus vaccines. Vaccine 2021, 39, 402–411. [Google Scholar] [CrossRef]
- Van Reeth, K.; Gracia, J.C.M.; Trus, I.; Sys, L.; Claes, G.; Versnaeyen, H.; Cox, E.; Krammer, F.; Qiu, Y. Heterologous prime-boost vaccination with H3N2 influenza viruses of swine favors cross-clade antibody responses and protection. NPJ Vaccines 2017, 2, 11. [Google Scholar] [CrossRef] [Green Version]
- Pica, N.; Palese, P. Toward a Universal Influenza Virus Vaccine: Prospects and Challenges. Annu. Rev. Med. 2013, 64, 189–202. [Google Scholar] [CrossRef] [Green Version]
- Jansen, J.M.; Gerlach, T.; Elbahesh, H.; Rimmelzwaan, G.F.; Saletti, G. Influenza virus-specific CD4+ and CD8+ T cell-mediated immunity induced by infection and vaccination. J. Clin. Virol. 2019, 119, 44–52. [Google Scholar] [CrossRef]
- Li, S.; Qiao, Y.; Xu, Y.; Li, P.; Nie, J.; Zhao, Q.; Chai, W.; Shi, Y.; Kong, W.; Shan, Y. Identification of Linear Peptide Immunogens with Verified Broad-spectrum Immunogenicity from the Conserved Regions within the Hemagglutinin Stem Domain of H1N1 Influenza Virus. Immunol. Investig. 2020, 51, 411–424. [Google Scholar] [CrossRef]
- McMahon, M.; Arunkumar, G.A.; Liu, W.-C.; Stadlbauer, D.; Albrecht, R.; Pavot, V.; Aramouni, M.; Lambe, T.; Gilbert, S.C.; Krammer, F. Vaccination With Viral Vectors Expressing Chimeric Hemagglutinin, NP and M1 Antigens Protects Ferrets Against Influenza Virus Challenge. Front. Immunol. 2019, 10, 2005. [Google Scholar] [CrossRef]
- Fiers, W.; De Filette, M.; El Bakkouri, K.; Schepens, B.; Roose, K.; Schotsaert, M.; Birkett, A.; Saelens, X. M2e-based universal influenza A vaccine. Vaccine 2009, 27, 6280–6283. [Google Scholar] [CrossRef]
- Vitelli, A.; Quirion, M.R.; Lo, C.-Y.; Misplon, J.A.; Grabowska, A.K.; Pierantoni, A.; Ammendola, V.; Price, G.E.; Soboleski, M.R.; Cortese, R.; et al. Vaccination to Conserved Influenza Antigens in Mice Using a Novel Simian Adenovirus Vector, PanAd3, Derived from the Bonobo Pan paniscus. PLoS ONE 2013, 8, e55435. [Google Scholar] [CrossRef] [Green Version]
- Berthoud, T.K.; Hamill, M.; Lillie, P.J.; Hwenda, L.; Collins, K.A.; Ewer, K.J.; Milicic, A.; Poyntz, H.C.; Lambe, T.; Fletcher, H.A.; et al. Potent CD8+ T cell Immunogenicity in Humans of a Novel Heterosubtypic Influenza A Vaccine, MVA-NP+M1. Clin. Infect. Dis. 2011, 52, 1. [Google Scholar] [CrossRef]
- Grant, E.; Wu, C.; Chan, K.; Eckle, S.; Bharadwaj, M.; Zou, Q.M.; Kedzierska, K.; Chen, W. Nucleoprotein of influenza A virus is a major target of immunodominant CD8+ T-cell responses. Immunol. Cell Biol. 2013, 91, 184–194. [Google Scholar] [CrossRef]
- Grant, E.; Quiñones-Parra, S.M.; Clemens, E.B.; Kedzierska, K. Corrigendum to ‘Human influenza viruses and CD8+ T cell responses’ [Curr. Opin. Virol. 16 (2016) 132–142]. Curr. Opin. Virol. 2016, 19, 99. [Google Scholar] [CrossRef]
- Grant, E.J.; Quiñones-Parra, S.M.; Clemens, E.B.; Kedzierska, K. Human influenza viruses and CD8+ T cell responses. Curr. Opin. Virol. 2016, 16, 132–142. [Google Scholar] [CrossRef]
- Gage, E.; Van Hoeven, N.; Cauwelaert, N.D.; Larsen, S.E.; Erasmus, J.; Orr, M.T.; Coler, R.N. Memory CD4+T cells enhance B-cell responses to drifting influenza immunization. Eur. J. Immunol. 2018, 49, 266–276. [Google Scholar] [CrossRef] [Green Version]
- Jegaskanda, S.; Co, M.D.T.; Cruz, J.; Subbarao, K.; Ennis, F.A.; Terajima, M. Induction of H7N9-Cross-Reactive Antibody-Dependent Cellular Cytotoxicity Antibodies by Human Seasonal Influenza A Viruses that are Directed Toward the Nucleoprotein. J. Infect. Dis. 2016, 215, 818–823. [Google Scholar] [CrossRef]
- LaMere, M.W.; Lam, H.-T.; Moquin, A.; Haynes, L.; Lund, F.E.; Randall, T.D.; Kaminski, D.A. Contributions of Antinucleoprotein IgG to Heterosubtypic Immunity against Influenza Virus. J. Immunol. 2011, 186, 4331–4339. [Google Scholar] [CrossRef] [Green Version]
- LaMere, M.W.; Moquin, A.; Lee, F.E.-H.; Misra, R.S.; Blair, P.J.; Haynes, L.; Randall, T.D.; Lund, F.E.; Kaminski, D.A. Regulation of Antinucleoprotein IgG by Systemic Vaccination and Its Effect on Influenza Virus Clearance. J. Virol. 2011, 85, 5027–5035. [Google Scholar] [CrossRef] [Green Version]
- Cullen, J.G.; McQuilten, H.A.; Quinn, K.M.; Olshansky, M.; Russ, B.E.; Morey, A.; Wei, S.; Prier, J.E.; La Gruta, N.L.; Doherty, P.C.; et al. CD4+ T help promotes influenza virus-specific CD8+ T cell memory by limiting metabolic dysfunction. Proc. Natl. Acad. Sci. USA 2019, 116, 4481–4488. [Google Scholar] [CrossRef] [Green Version]
- Boonnak, K.; Subbarao, K. Memory CD4+ T cells: Beyond “helper” functions. J. Clin. Investig. 2012, 122, 2768–2770. [Google Scholar] [CrossRef]
- McKinstry, K.K.; Dutton, R.W.; Swain, S.L.; Strutt, T.M. Memory CD4 T Cell-Mediated Immunity against Influenza A Virus: More than a Little Helpful. Arch. Immunol. Ther. Exp. 2013, 61, 341–353. [Google Scholar] [CrossRef] [Green Version]
- McKinstry, K.K.; Strutt, T.M.; Kuang, Y.; Brown, D.M.; Sell, S.; Dutton, R.W.; Swain, S.L. Memory CD4+ T cells protect against influenza through multiple synergizing mechanisms. J. Clin. Investig. 2012, 122, 2847–2856. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, T.M.; Li, C.K.F.; Chui, C.S.C.; Huang, A.K.Y.; Perkins, M.; Liebner, J.C.; Lambkin-Williams, R.; Gilbert, A.; Oxford, J.; Nicholas, B.; et al. Preexisting influenza-specific CD4+ T cells correlate with disease protection against influenza challenge in humans. Nat. Med. 2012, 18, 274–280. [Google Scholar] [CrossRef]
- Rudicell, R.S.; Garinot, M.; Kanekiyo, M.; Kamp, H.D.; Swanson, K.; Chou, T.-H.; Dai, S.; Bedel, O.; Simard, D.; Gillespie, R.A.; et al. Comparison of adjuvants to optimize influenza neutralizing antibody responses. Vaccine 2019, 37, 6208–6220. [Google Scholar] [CrossRef]
- Bernasconi, V.; Norling, K.; Bally, M.; Höök, F.; Lycke, N.Y. Mucosal Vaccine Development Based on Liposome Technology. J. Immunol. Res. 2016, 2016, 5482087. [Google Scholar] [CrossRef]
- Lycke, N. Recent progress in mucosal vaccine development: Potential and limitations. Nat. Rev. Immunol. 2012, 12, 592–605. [Google Scholar] [CrossRef]
- Rose, M.A.; Zielen, S.; Baumann, U. Mucosal immunity and nasal influenza vaccination. Expert Rev. Vaccines 2012, 11, 595–607. [Google Scholar] [CrossRef]
- Lau, Y.-F.; Wright, A.R.; Subbarao, K. The Contribution of Systemic and Pulmonary Immune Effectors to Vaccine-Induced Protection from H5N1 Influenza Virus Infection. J. Virol. 2012, 86, 5089–5098. [Google Scholar] [CrossRef] [Green Version]
- Aevermann, B.D.; Pickett, B.E.; Kumar, S.; Klem, E.B.; Agnihothram, S.; Askovich, P.S.; Bankhead, A.; Bolles, M.; Carter, V.; Chang, J.; et al. A comprehensive collection of systems biology data characterizing the host response to viral infection. Sci. Data 2014, 1, 140033. [Google Scholar] [CrossRef] [Green Version]
- Askovich, P.S.; Sanders, C.J.; Rosenberger, C.M.; Diercks, A.H.; Dash, P.; Navarro, G.; Vogel, P.; Doherty, P.C.; Thomas, P.G.; Aderem, A. Differential Host Response, Rather Than Early Viral Replication Efficiency, Correlates with Pathogenicity Caused by Influenza Viruses. PLoS ONE 2013, 8, e74863. [Google Scholar] [CrossRef]
- Bao, J.; Cui, D.; Wang, X.; Zou, Q.; Zhao, D.; Zheng, S.; Yu, F.; Huang, L.; Dong, Y.; Yang, X.; et al. Decreased Frequencies of Th17 and Tc17 Cells in Patients Infected with Avian Influenza A (H7N9) Virus. J. Immunol. Res. 2019, 2019, 1418251. [Google Scholar] [CrossRef] [Green Version]
- Cargnelutti, D.E.; Sanchez, M.; Alvarez, P.; Boado, L.; Mattion, N.; Scodeller, E.A. Enhancement of Th1 immune responses to recombinant influenza nucleoprotein by Ribi adjuvant. New Microbiol. 2013, 36, 145–151. [Google Scholar]
- Cargnelutti, D.E.; Sanchez, M.V.; Alvarez, P.; Boado, L.; Glikmann, G.; Mattion, N.; Scodeller, E.A. Improved Immune Response to Recombinant Influenza Nucleoprotein Formulated with ISCOMATRIX. J. Microbiol. Biotechnol. 2012, 22, 416–421. [Google Scholar] [CrossRef] [Green Version]
- Switalla, S.; Lauenstein, L.; Prenzler, F.; Knothe, S.; Förster, C.; Fieguth, H.-G.; Pfennig, O.; Schaumann, F.; Martin, C.; Guzman, C.; et al. Natural innate cytokine response to immunomodulators and adjuvants in human precision-cut lung slices. Toxicol. Appl. Pharmacol. 2010, 246, 107–115. [Google Scholar] [CrossRef]
- Borsutzky, S.; Fiorelli, V.; Ebensen, T.; Tripiciano, A.; Rharbaoui, F.; Scoglio, A.; Link, C.; Nappi, F.; Morr, M.; Buttò, S.; et al. Efficient mucosal delivery of the HIV-1 Tat protein using the synthetic lipopeptide MALP-2 as adjuvant. Eur. J. Immunol. 2003, 33, 1548–1556. [Google Scholar] [CrossRef]
- Link, C.; Gavioli, R.; Ebensen, T.; Canella, A.; Reinhard, E.; Guzmán, C.A. The Toll-like receptor ligand MALP-2 stimulates dendritic cell maturation and modulates proteasome composition and activity. Eur. J. Immunol. 2004, 34, 899–907. [Google Scholar] [CrossRef]
- Becker, P.D.; Fiorentini, S.; Link, C.; Tosti, G.; Ebensen, T.; Caruso, A.; Guzmán, C.A. The HIV-1 matrix protein p17 can be efficiently delivered by intranasal route in mice using the TLR 2/6 agonist MALP-2 as mucosal adjuvant. Vaccine 2006, 24, 5269–5276. [Google Scholar] [CrossRef]
- Borsutzky, S.; Ebensen, T.; Link, C.; Becker, P.D.; Fiorelli, V.; Cafaro, A.; Ensoli, B.; Guzmán, C.A. Efficient systemic and mucosal responses against the HIV-1 Tat protein by prime/boost vaccination using the lipopeptide MALP-2 as adjuvant. Vaccine 2006, 24, 2049–2056. [Google Scholar] [CrossRef]
- Rharbaoui, F.; Drabner, B.; Borsutzky, S.; Winckler, U.; Morr, M.; Ensoli, B.; Mühlradt, P.F.; Guzmán, C.A. The Mycoplasma-derived lipopeptide MALP-2 is a potent mucosal adjuvant. Eur. J. Immunol. 2002, 32, 2857–2865. [Google Scholar] [CrossRef]
- Palma, C.; Iona, E.; Ebensen, T.; Guzmán, C.; Cassone, A. The Toll-like Receptor 2/6 Ligand MALP-2 Reduces the Viability of Mycobacterium tuberculosis in Murine Macrophages. Open Microbiol. J. 2009, 3, 47–52. [Google Scholar] [CrossRef]
- Schulze, K.; Ebensen, T.; Chandrudu, S.; Skwarczynski, M.; Toth, I.; Olive, C.; Guzman, C.A. Bivalent mucosal peptide vaccines administered using the LCP carrier system stimulate protective immune responses against Streptococcus pyogenes infection. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 2463–2474. [Google Scholar] [CrossRef]
- Pandey, S.P.; Chandel, H.S.; Srivastava, S.; Selvaraj, S.; Jha, M.K.; Shukla, D.; Ebensen, T.; Guzman, C.A.; Saha, B. Pegylated Bisacycloxypropylcysteine, a Diacylated Lipopeptide Ligand of TLR6, Plays a Host-Protective Role against Experimental Leishmania major Infection. J. Immunol. 2014, 193, 3632–3643. [Google Scholar] [CrossRef] [Green Version]
- Krishnaswamy, J.K.; Jirmo, A.C.; Baru, A.M.; Ebensen, T.; Guzmán, C.A.; Sparwasser, T.; Behrens, G.M.N. Toll-Like Receptor–2 Agonist–Allergen Coupling Efficiently Redirects Th2 Cell Responses and Inhibits Allergic Airway Eosinophilia. Am. J. Respir. Cell Mol. Biol. 2012, 47, 852–863. [Google Scholar] [CrossRef]
- Knothe, S.; Mutschler, V.; Rochlitzer, S.; Winkler, C.; Ebensen, T.; Guzman, C.; Hohlfeld, J.; Braun, A.; Muller, M. Local treatment with BPPcysMPEG reduces allergic airway inflammation in sensitized mice. Immunobiology 2011, 216, 110–117. [Google Scholar] [CrossRef]
- Prajeeth, C.K.; Jirmo, A.C.; Krishnaswamy, J.K.; Ebensen, T.; Guzman, C.A.; Weiss, S.; Constabel, H.; Schmidt, R.E.; Behrens, G.M.N. The synthetic TLR2 agonist BPPcysMPEG leads to efficient cross-priming against co-administered and linked antigens. Eur. J. Immunol. 2010, 40, 1272–1283. [Google Scholar] [CrossRef]
- Xu, D.; Horst, K.; Wang, W.; Luo, P.; Shi, Y.; Tschernig, T.; Greven, J.; Hildebrand, F. The Influence of Macrophage-Activating Lipopeptide 2 (MALP-2) on Local and Systemic Inflammatory Response in a Murine Two-Hit Model of Hemorrhagic Shock and Subsequent Sepsis. Inflammation 2021, 44, 481–492. [Google Scholar] [CrossRef]
- Metzger, J.; Wiesmüller, K.-H.; Schaude, R.; Bessler, W.G.; Jung, G. Synthesis of novel immunologically active tripalmitoyl-S-glycerylcysteinyl lipopeptides as useful intermediates for immunogen preparations. Int. J. Pept. Protein Res. 1991, 37, 46–57. [Google Scholar] [CrossRef]
- Mühlradt, P.F.; Kieß, M.; Meyer, H.; Süssmuth, R.; Jung, G. Isolation, Structure Elucidation, and Synthesis of a Macrophage Stimulatory Lipopeptide from Mycoplasma fermentans Acting at Picomolar Concentration. J. Exp. Med. 1997, 185, 1951–1958. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, B.; Błażejewska, P.; Heßmann, M.; Bruder, D.; Geffers, R.; Mauel, S.; Gruber, A.D.; Schughart, K. Host Genetic Background Strongly Influences the Response to Influenza A Virus Infections. PLoS ONE 2009, 4, e4857. [Google Scholar] [CrossRef] [Green Version]
- Madhun, A.S.; Haaheim, L.R.; Nøstbakken, J.K.; Ebensen, T.; Chichester, J.; Yusibov, V.; Guzman, C.A.; Cox, R.J. Intranasal c-di-GMP-adjuvanted plant-derived H5 influenza vaccine induces multifunctional Th1 CD4+ cells and strong mucosal and systemic antibody responses in mice. Vaccine 2011, 29, 4973–4982. [Google Scholar] [CrossRef]
- Pedersen, G.K.; Ebensen, T.; Gjeraker, I.H.; Svindland, S.; Bredholt, G.; Guzmán, C.A.; Cox, R.J. Evaluation of the Sublingual Route for Administration of Influenza H5N1 Virosomes in Combination with the Bacterial Second Messenger c-di-GMP. PLoS ONE 2011, 6, e26973. [Google Scholar] [CrossRef] [Green Version]
- Carragher, D.M.; Kaminski, D.A.; Moquin, A.; Hartson, L.; Randall, T.D. A Novel Role for Non-Neutralizing Antibodies against Nucleoprotein in Facilitating Resistance to Influenza Virus. J. Immunol. 2008, 181, 4168–4176. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, M.; Ebensen, T.; Schulze, K.; Cargnelutti, D.E.; Blazejewska, P.; Scodeller, E.A.; Guzmán, C.A. Intranasal Delivery of Influenza rNP Adjuvanted with c-di-AMP Induces Strong Humoral and Cellular Immune Responses and Provides Protection against Virus Challenge. PLoS ONE 2014, 9, e104824. [Google Scholar] [CrossRef]
- Zheng, M.; Liu, F.; Shen, Y.; Wang, S.; Xu, W.; Fang, F.; Sun, B.; Xie, Z.; Chen, Z. Cross-protection against influenza virus infection by intranasal administration of nucleoprotein-based vaccine with compound 48/80 adjuvant. Hum. Vaccines Immunother. 2015, 11, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Derrick, S.C.; Yabe, I.M.; Yang, A.; Morris, S.L. Vaccine-induced anti-tuberculosis protective immunity in mice correlates with the magnitude and quality of multifunctional CD4 T cells. Vaccine 2011, 29, 2902–2909. [Google Scholar] [CrossRef]
- Seder, R.A.; Darrah, P.A.; Roederer, M. T cell quality in memory and protection: Implications for vaccine design. Nat. Rev. Immunol. 2008, 8, 247–258. [Google Scholar] [CrossRef]
- Thakur, A.; Pedersen, L.E.; Jungersen, G. Immune markers and correlates of protection for vaccine induced immune responses. Vaccine 2012, 30, 4907–4920. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.A.; Li, M.O. TGF-β: Guardian of T Cell Function. J. Immunol. 2013, 191, 3973–3979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbahesh, H.; Gerlach, T.; Saletti, G.; Rimmelzwaan, G.F. Response Modifiers: Tweaking the Immune Response Against Influenza A Virus. Front. Immunol. 2019, 10, 809. [Google Scholar] [CrossRef]
- Sarawar, S.; Gabaglia, C.R.; Sanchez, A.; Hatta, Y.; Dias, P.; Neumann, G.; Kawaoka, Y.; Bilsel, P. Longevity and Mechanism of Heterosubtypic Protection Induced by M2SR (M2-Deficient Single-Replication) Live Influenza Virus Vaccine in Mice. Vaccines 2022, 10, 2131. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, S. Heterosubtypic T cell Immunity to Influenza in Humans: Challenges for Universal T cell Influenza Vaccines. Front. Immunol. 2016, 7, 195. [Google Scholar] [CrossRef] [Green Version]
- Rudolph, W.; Ben Yedidia, T. A universal influenza vaccine: Where are we in the pursuit of this “Holy Grail”? Hum. Vaccines 2011, 7, 10–11. [Google Scholar] [CrossRef]
- Sah, P.; Alfaro-Murillo, J.A.; Fitzpatrick, M.C.; Neuzil, K.M.; Meyers, L.A.; Singer, B.H.; Galvani, A.P. Future epidemiological and economic impacts of universal influenza vaccines. Proc. Natl. Acad. Sci. USA 2019, 116, 22409. [Google Scholar] [CrossRef] [Green Version]
- Calzas, C.; Chevalier, C. Innovative Mucosal Vaccine Formulations Against Influenza A Virus Infections. Front. Immunol. 2019, 10, 1605. [Google Scholar] [CrossRef] [Green Version]
- Sridhar, S.; Brokstad, K.A.; Cox, R.J. Influenza Vaccination Strategies: Comparing Inactivated and Live Attenuated Influenza Vaccines. Vaccines 2015, 3, 373–389. [Google Scholar] [CrossRef] [PubMed]
- McMichael, A.J.; Gotch, F.M.; Noble, G.R.; Beare, P.A.S. Cytotoxic T cell Immunity to Influenza. N. Engl. J. Med. 1983, 309, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.M.; Askonas, B.A. Influenza nucleoprotein-specific cytotoxic T cell clones are protective in vivo. Immunology 1986, 58, 417–420. [Google Scholar] [PubMed]
- Yewdell, J.W.; Bennink, J.R.; Smith, G.L.; Moss, B. Influenza A virus nucleoprotein is a major target antigen for cross-reactive anti-influenza A virus cytotoxic T lymphocytes. Proc. Natl. Acad. Sci. USA 1985, 82, 1785–1789. [Google Scholar] [CrossRef] [Green Version]
- Antrobus, R.D.; Lillie, P.; Berthoud, T.K.; Spencer, A.; McLaren, J.; Ladell, K.; Lambe, T.; Milicic, A.; Price, D.; Hill, A.V.S.; et al. A T Cell-Inducing Influenza Vaccine for the Elderly: Safety and Immunogenicity of MVA-NP+M1 in Adults Aged over 50 Years. PLoS ONE 2012, 7, e48322. [Google Scholar] [CrossRef]
- Lambe, T.; Carey, J.B.; Li, Y.; Spencer, A.J.; van Laarhoven, A.; Mullarkey, C.E.; Vrdoljak, A.; Moore, A.C.; Gilbert, S.C. Immunity Against Heterosubtypic Influenza Virus Induced By Adenovirus And MVA Expressing Nucleoprotein And Matrix Protein-1. Sci. Rep. 2013, 3, 1443. [Google Scholar] [CrossRef] [Green Version]
- Price, G.E.; Lo, C.-Y.; Misplon, J.A.; Epstein, S.L. Mucosal Immunization with a Candidate Universal Influenza Vaccine Reduces Virus Transmission in a Mouse Model. J. Virol. 2014, 88, 6019–6030. [Google Scholar] [CrossRef] [Green Version]
- Retamal-Díaz, A.; Riquelme-Neira, R.; Sáez, D.; Rivera, A.; Fernández, P.; Cabrera, A.; Guzmán, C.A.; Oñate, Á. Use of S-[2,3-Bispalmitoyiloxy-(2R)-Propyl]-R-Cysteinyl-Amido-Monomethoxy Polyethylene Glycol as an Adjuvant Improved Protective Immunity Associated with a DNA Vaccine Encoding Cu,Zn Superoxide Dismutase of Brucella abortus in Mice. Clin. Vaccine Immunol. 2014, 21, 1474–1480. [Google Scholar] [CrossRef] [Green Version]
- McLain, L.; Dimmock, N.J. Protection of Mice from Lethal Influenza by Adoptive Transfer of Non-neutralizing Haemagglutination-inhibiting IgG Obtained from the Lungs of Infected Animals Treated with Defective Interfering Virus. J. Gen. Virol. 1989, 70, 2615–2624. [Google Scholar] [CrossRef]
- Jegaskanda, S.; Reading, P.C.; Kent, S.J. Influenza-Specific Antibody-Dependent Cellular Cytotoxicity: Toward a Universal Influenza Vaccine. J. Immunol. 2014, 193, 469–475. [Google Scholar] [CrossRef] [Green Version]
- Bodewes, R.; Geelhoed-Mieras, M.M.; Wrammert, J.; Ahmed, R.; Wilson, P.C.; Fouchier, R.A.M.; Osterhaus, A.D.M.E.; Rimmelzwaan, G.F. In Vitro Assessment of the Immunological Significance of a Human Monoclonal Antibody Directed to the Influenza A Virus Nucleoprotein. Clin. Vaccine Immunol. 2013, 20, 1333–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renegar, K.B.; Small, P.A., Jr.; Boykins, L.G.; Wright, P.F. Role of IgA versus IgG in the Control of Influenza Viral Infection in the Murine Respiratory Tract. J. Immunol. 2004, 173, 1978–1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, S.; Miyata, K.; Matsuo, K.; Asanuma, H.; Takahashi, H.; Nakajima, K.; Suzuki, Y.; Aizawa, C.; Kurata, T. Acceleration of influenza virus clearance by Th1 cells in the nasal site of mice immunized intranasally with adjuvant-combined recombinant nucleoprotein. J. Immunol. 1996, 156, 3892–3900. [Google Scholar] [CrossRef] [PubMed]
- Mazanec, M.B.; Coudret, C.L.; Fletcher, D.R. Intracellular neutralization of influenza virus by immunoglobulin A anti-hemagglutinin monoclonal antibodies. J. Virol. 1995, 69, 1339–1343. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.; Zhang, Y.; Li, Q.; Chen, Y.; He, B.; Yang, J.; Tu, H.; Lei, L.; Yan, H. Matrix Protein-Specific IgA Antibody Inhibits Measles Virus Replication by Intracellular Neutralization. J. Virol. 2011, 85, 11090–11097. [Google Scholar] [CrossRef] [Green Version]
- Arias, M.A.; Van Roey, G.A.; Tregoning, J.S.; Moutaftsi, M.; Coler, R.N.; Windish, H.P.; Reed, S.G.; Carter, D.; Shattock, R.J. Glucopyranosyl Lipid Adjuvant (GLA), a Synthetic TLR4 Agonist, Promotes Potent Systemic and Mucosal Responses to Intranasal Immunization with HIVgp140. PLoS ONE 2012, 7, e41144. [Google Scholar] [CrossRef] [Green Version]
- Brereton, C.F.; Sutton, C.E.; Ross, P.J.; Iwakura, Y.; Pizza, M.; Rappuoli, R.; Lavelle, E.C.; Mills, K.H.G. Escherichia coli Heat-Labile Enterotoxin Promotes Protective Th17 Responses against Infection by Driving Innate IL-1 and IL-23 Production. J. Immunol. 2011, 186, 5896–5906. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Huang, J.; Duan, B.; Traficante, D.C.; Hong, H.; Risech, M.; Lory, S.; Priebe, G.P. Th17-stimulating Protein Vaccines Confer Protection against Pseudomonas aeruginosa Pneumonia. Am. J. Respir. Crit. Care Med. 2012, 186, 420–427. [Google Scholar] [CrossRef] [Green Version]
- Happel, K.I.; Dubin, P.J.; Zheng, M.; Ghilardi, N.; Lockhart, C.; Quinton, L.; Odden, A.R.; Shellito, J.E.; Bagby, G.J.; Nelson, S.; et al. Divergent roles of IL-23 and IL-12 in host defense against Klebsiella pneumoniae. J. Exp. Med. 2005, 202, 761–769. [Google Scholar] [CrossRef]
- Khader, S.A.; Bell, G.K.; Pearl, J.E.; Fountain, J.J.; Rangel-Moreno, J.; Cilley, G.E.; Shen, F.; Eaton, S.M.; Gaffen, S.L.; Swain, S.L.; et al. IL-23 and IL-17 in the establishment of protective pulmonary CD4+ T cell responses after vaccination and during Mycobacterium tuberculosis challenge. Nat. Immunol. 2007, 8, 369–377. [Google Scholar] [CrossRef]
- Hamada, H.; Garcia-Hernandez, M.D.L.L.; Reome, J.B.; Misra, S.K.; Strutt, T.M.; McKinstry, K.K.; Cooper, A.M.; Swain, S.L.; Dutton, R.W. Tc17, a Unique Subset of CD8 T Cells That Can Protect against Lethal Influenza Challenge. J. Immunol. 2009, 182, 3469–3481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Chan, C.C.; Yang, M.; Deng, J.; Poon, V.K.; Leung, V.H.; Ko, K.-H.; Zhou, J.; Yuen, K.Y.; Zheng, B.-J.; et al. A critical role of IL-17 in modulating the B cell response during H5N1 influenza virus infection. Cell Mol. Immunol. 2011, 8, 462–468. [Google Scholar] [CrossRef] [Green Version]
- Gopal, R.; Rangel-Moreno, J.; Junecko, B.A.F.; Mallon, D.J.; Chen, K.; Pociask, D.A.; Connell, T.D.; Reinhart, T.A.; Alcorn, J.F.; Ross, T.; et al. Mucosal Pre-Exposure to Th17-Inducing Adjuvants Exacerbates Pathology after Influenza Infection. Am. J. Pathol. 2014, 184, 55–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egarnes, B.; Gosselin, J. Contribution of Regulatory T Cells in Nucleotide-Binding Oligomerization Domain 2 Response to Influenza Virus Infection. Front. Immunol. 2018, 9, 132. [Google Scholar] [CrossRef] [PubMed]
- Betts, R.J.; Prabhu, N.; Ho, A.W.S.; Lew, F.C.; Hutchinson, P.E.; Rotzschke, O.; Macary, P.A.; Kemeny, D.M. Influenza A Virus Infection Results in a Robust, Antigen-Responsive, and Widely Disseminated Foxp3+ Regulatory T Cell Response. J. Virol. 2012, 86, 2817–2825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, D.M.; Lee, S.; Garcia-Hernandez, M.D.L.L.; Swain, S.L. Multifunctional CD4 Cells Expressing Gamma Interferon and Perforin Mediate Protection against Lethal Influenza Virus Infection. J. Virol. 2012, 86, 6792–6803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trieu, M.-C.; Zhou, F.; Lartey, S.L.; Sridhar, S.; Mjaaland, S.; Cox, R.J. Augmented CD4+ T cell and humoral responses after repeated annual influenza vaccination with the same vaccine component A/H1N1pdm09 over 5 years. NPJ Vaccines 2018, 3, 37. [Google Scholar] [CrossRef] [Green Version]
- Folegatti, P.M.; Bellamy, D.; Flaxman, A.; Mair, C.; Ellis, C.; Ramon, R.L.; Lopez, F.R.; Mitton, C.; Baker, M.; Poulton, I.; et al. Safety and Immunogenicity of the Heterosubtypic Influenza A Vaccine MVA-NP+M1 Manufactured on the AGE1.CR.pIX Avian Cell Line. Vaccines 2019, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Withanage, K.; De Coster, I.; Cools, N.; Viviani, S.; Tourneur, J.; Chevandier, M.; Lambiel, M.; Willems, P.; Le Vert, A.; Nicolas, F.; et al. Phase 1 Randomized, Placebo-Controlled, Dose-Escalating Study to Evaluate OVX836, a Nucleoprotein-Based Influenza Vaccine: Intramuscular Results. J. Infect. Dis. 2021, 226, 119–127. [Google Scholar] [CrossRef]
- Park, J.; Lee, H. Delivery Routes for COVID-19 Vaccines. Vaccines 2021, 9, 524. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanchez, M.V.; Ebensen, T.; Schulze, K.; Cargnelutti, D.E.; Scodeller, E.A.; Guzmán, C.A. Protective Efficacy of a Mucosal Influenza Vaccine Formulation Based on the Recombinant Nucleoprotein Co-Administered with a TLR2/6 Agonist BPPcysMPEG. Pharmaceutics 2023, 15, 912. https://doi.org/10.3390/pharmaceutics15030912
Sanchez MV, Ebensen T, Schulze K, Cargnelutti DE, Scodeller EA, Guzmán CA. Protective Efficacy of a Mucosal Influenza Vaccine Formulation Based on the Recombinant Nucleoprotein Co-Administered with a TLR2/6 Agonist BPPcysMPEG. Pharmaceutics. 2023; 15(3):912. https://doi.org/10.3390/pharmaceutics15030912
Chicago/Turabian StyleSanchez, Maria Victoria, Thomas Ebensen, Kai Schulze, Diego Esteban Cargnelutti, Eduardo A. Scodeller, and Carlos A. Guzmán. 2023. "Protective Efficacy of a Mucosal Influenza Vaccine Formulation Based on the Recombinant Nucleoprotein Co-Administered with a TLR2/6 Agonist BPPcysMPEG" Pharmaceutics 15, no. 3: 912. https://doi.org/10.3390/pharmaceutics15030912