
Bioluminescence Imaging and ICP-MS Associated with SPION as a Tool for Hematopoietic Stem and Progenitor Cells Homing and Engraftment Evaluation

 ,
,  ,
,  , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement and Animal Model
2.2. In Vitro
2.2.1. Aspiration and Isolation of HSPC from Animals Treated with 5-FU
2.2.2. Immunophenotypic Characterization of Isolated Cells from Animals Treated with 5-FU
2.2.3. Effects of 5-FU in BM Cellularity
2.2.4. Transduction with Lentiviral Vectors for Luciferase Expression in HSPC
2.2.5. Labeling of HSPC with SPION
2.3. In Vivo
2.3.1. Bone Marrow Transplantation Model
2.3.2. Evaluation of Homing of HSPC Labeled with SPION by ICP-MS
Tissue Digestion
Iron Quantification by ICP-MS
2.3.3. Evaluation of HSPC Engraftment by BLI
2.3.4. Hematopoiesis Reconstitution
2.4. Statistical Analysis
3. Results
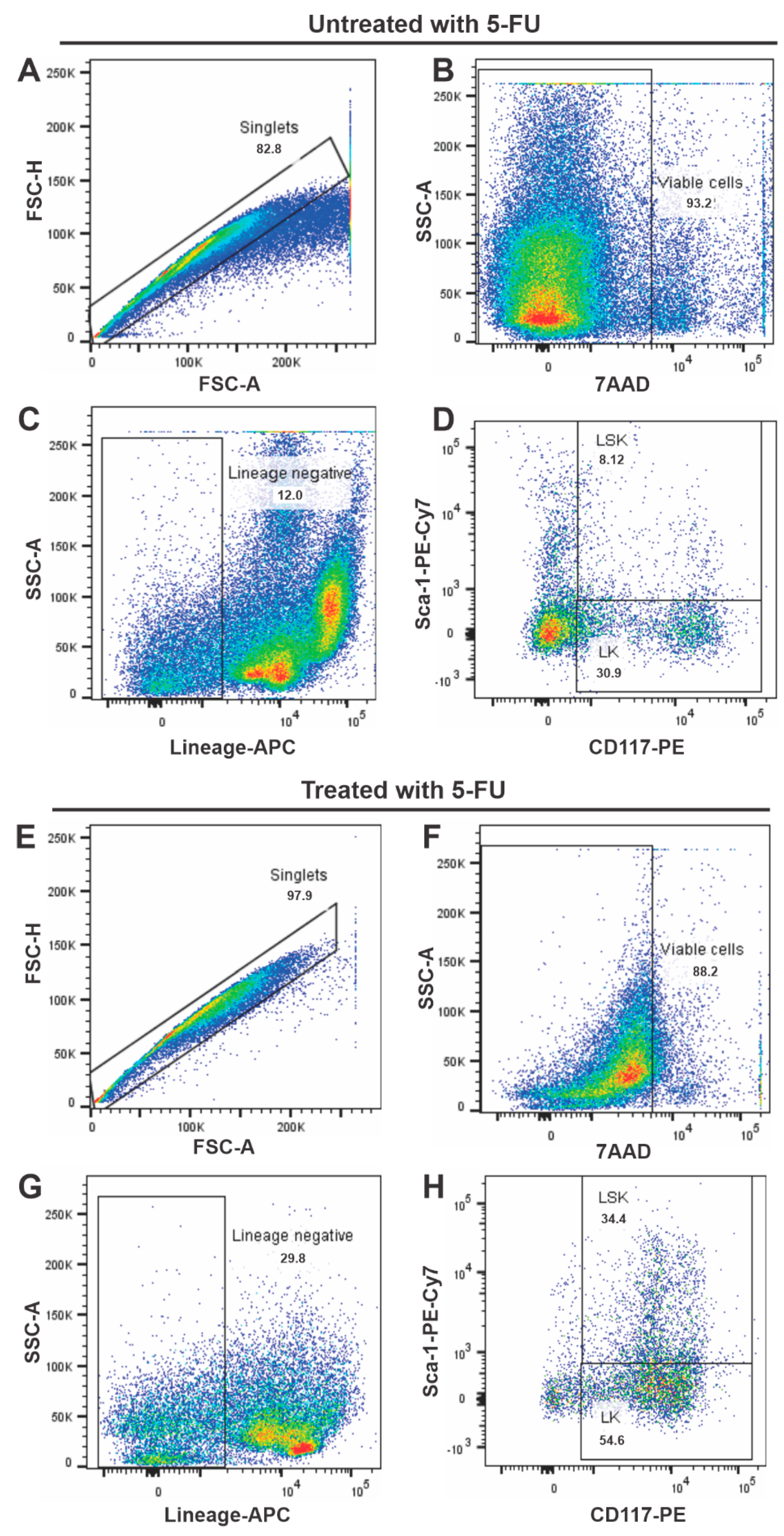
3.1. Immunophenotypic Characterization of Isolated Cells from Animals Treated with 5-FU
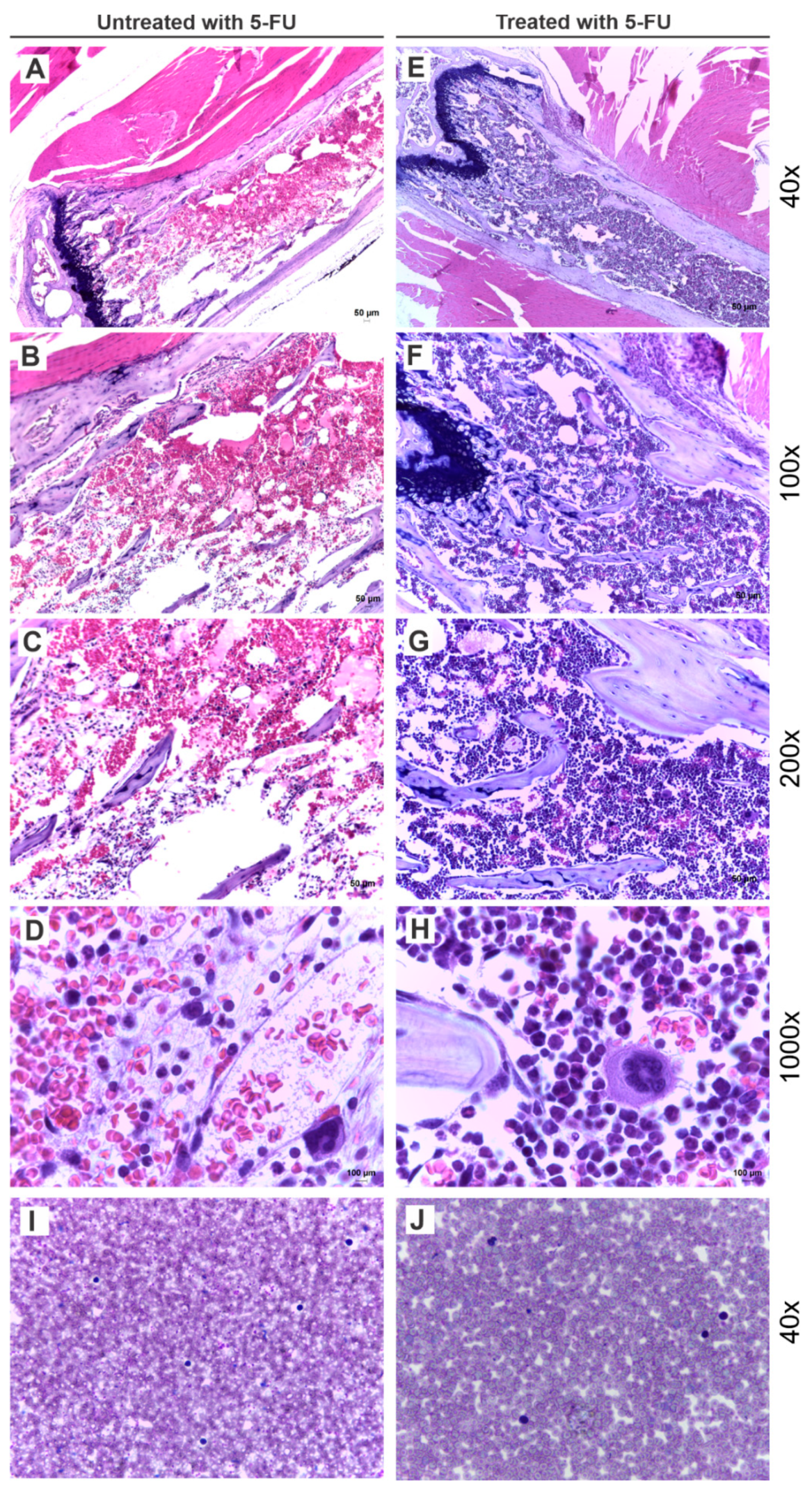
3.2. Effects of 5-FU on BM Cellularity
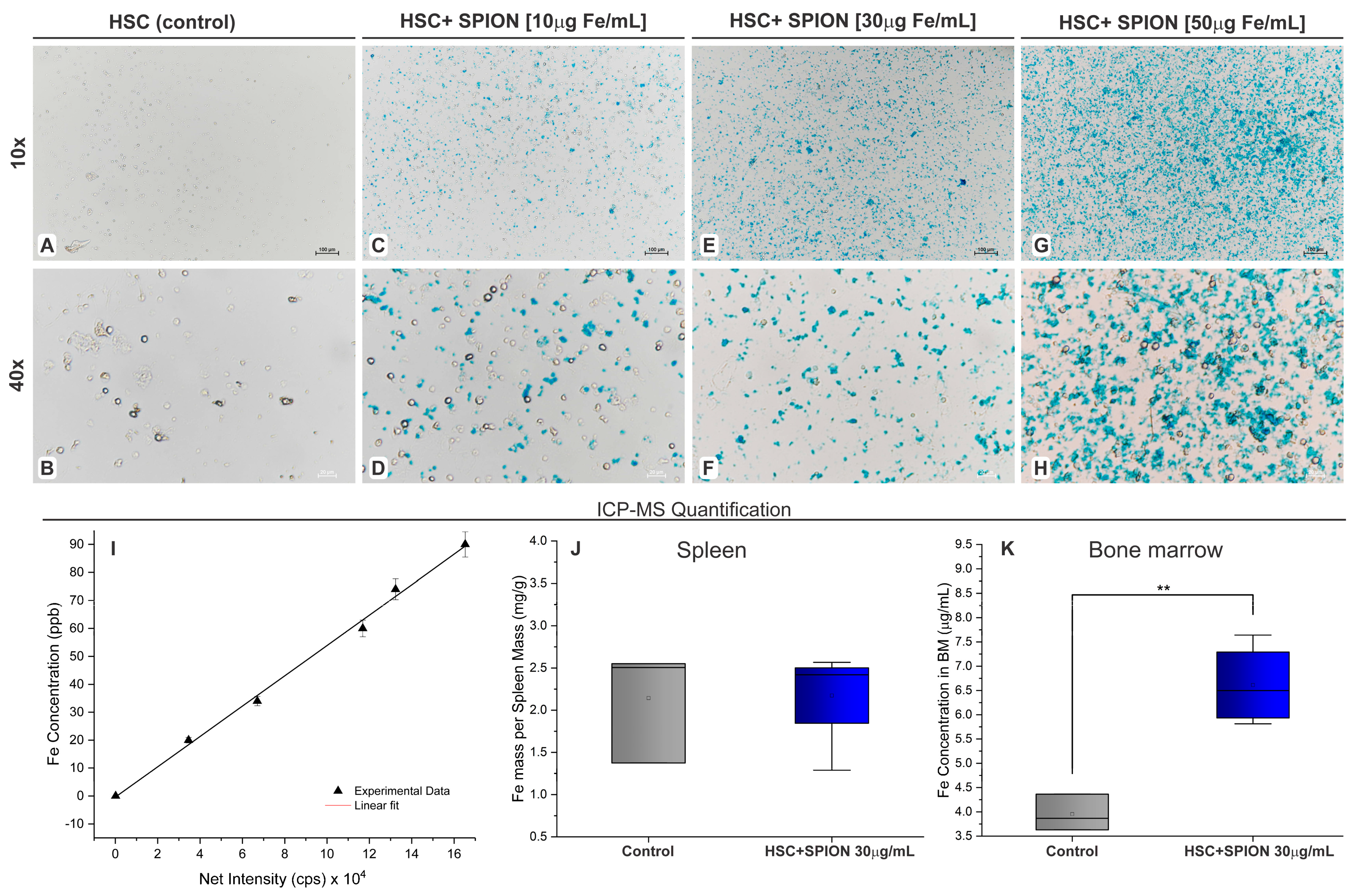
3.3. Evaluation of HSPC Labeled with SPION and Their In Vivo Migration
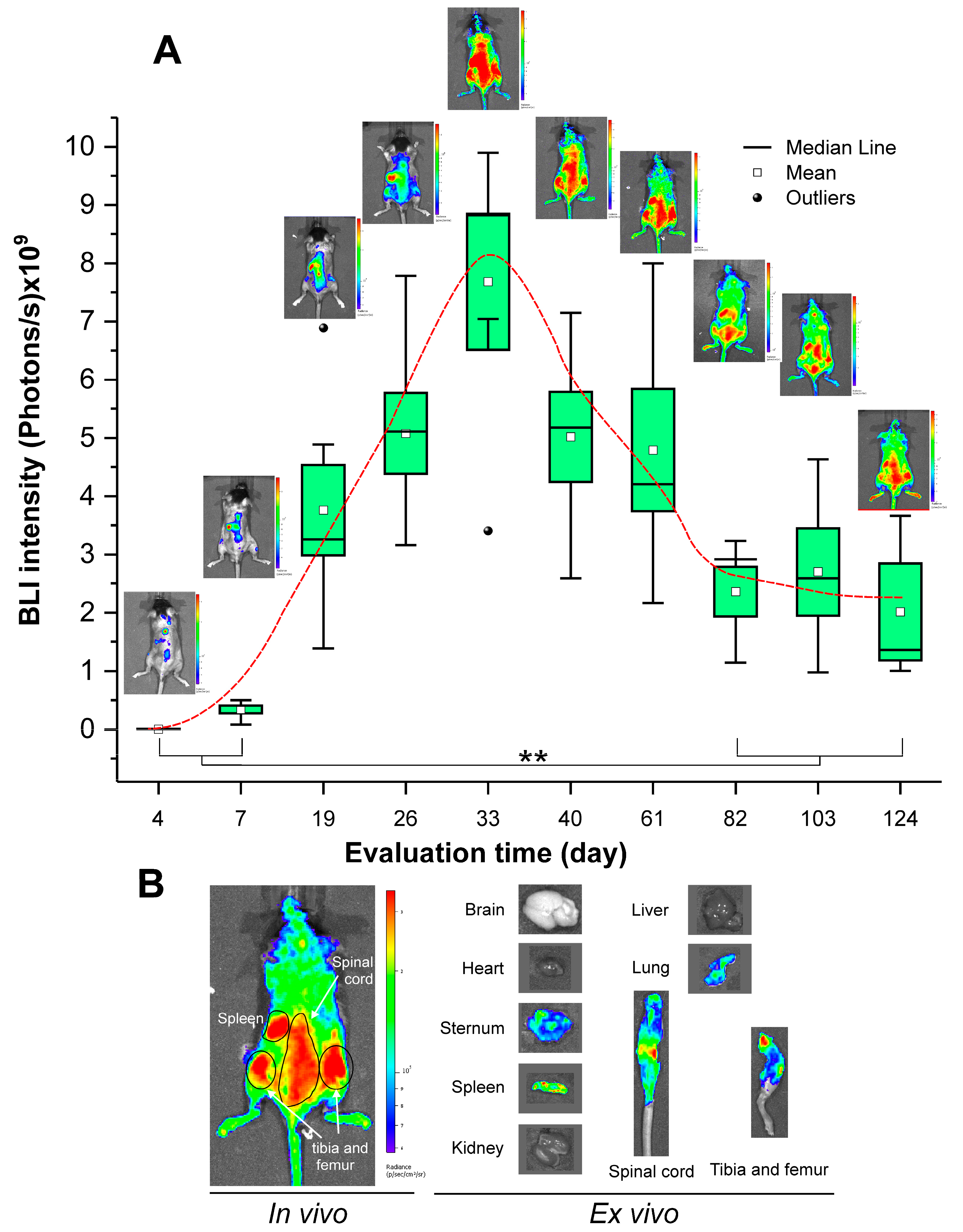
3.4. Evaluation of HSPC Engraftment by BLI
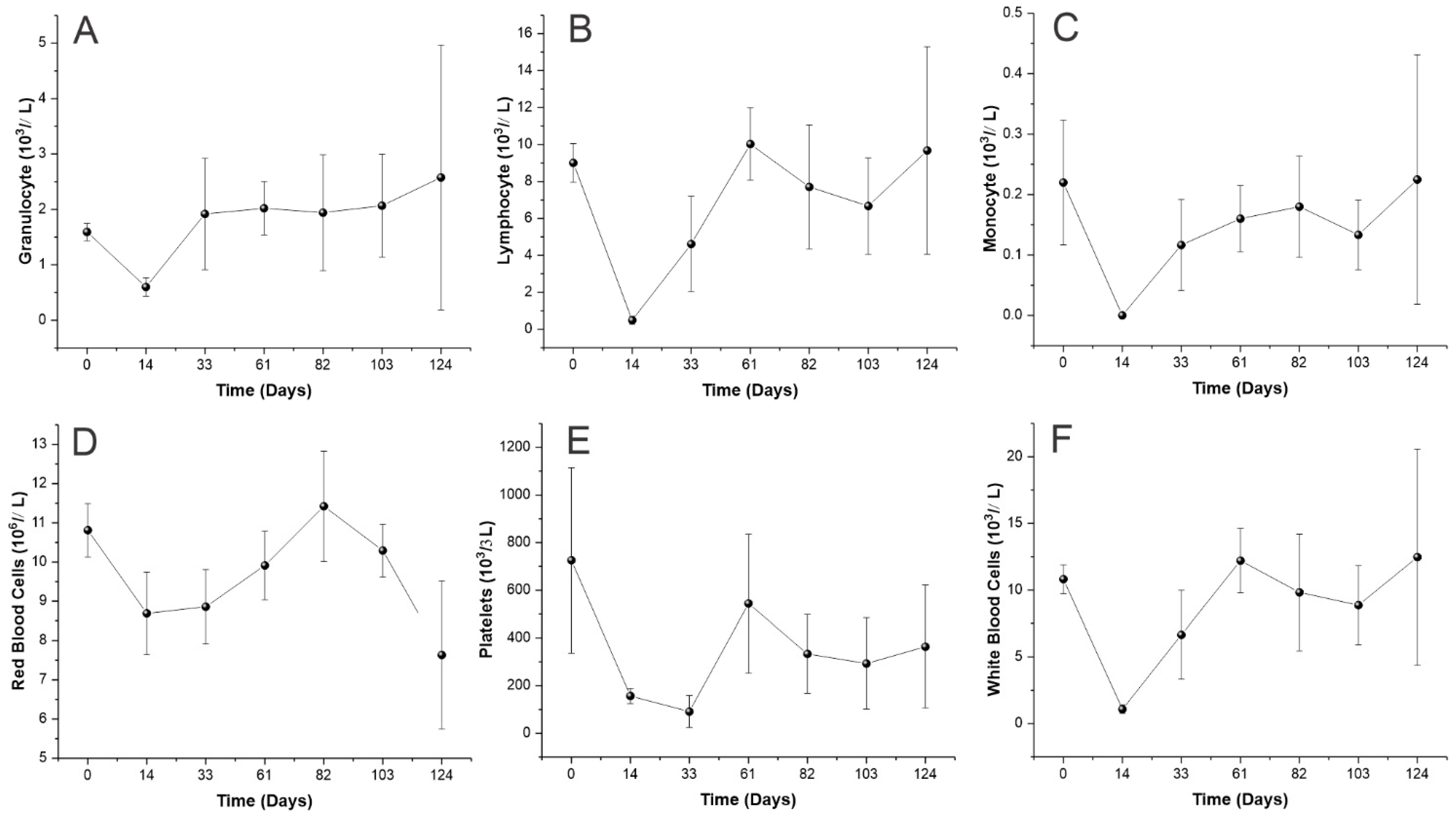
3.5. Hematopoiesis Reconstitution
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hawley, R.G.; Ramezani, A.; Hawley, T.S. Hematopoietic stem cells. Methods Enzymol. 2006, 419, 149–179. [Google Scholar] [CrossRef]
- Kandarakov, O.; Belyavsky, A.; Semenova, E. Bone Marrow Niches of Hematopoietic Stem and Progenitor Cells. Int. J. Mol. Sci. 2022, 23, 4462. [Google Scholar] [CrossRef] [PubMed]
- Castro, C.G.d., Jr.; Gregianin, L.J.; Brunetto, A.L. Transplante de medula óssea e transplante de sangue de cordão umbilical em pediatria. J. Pediatr. 2001, 77, 345–360. [Google Scholar] [CrossRef]
- Lee, J.Y.; Hong, S.H. Hematopoietic Stem Cells and Their Roles in Tissue Regeneration. Int. J. Stem. Cells 2020, 13, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Snowden, J.A.; Sánchez-Ortega, I.; Corbacioglu, S.; Basak, G.W.; Chabannon, C.; de la Camara, R.; Dolstra, H.; Duarte, R.F.; Glass, B.; Greco, R.; et al. Indications for haematopoietic cell transplantation for haematological diseases, solid tumours and immune disorders: Current practice in Europe, 2022. Bone Marrow Transplant. 2022, 57, 1217–1239. [Google Scholar] [CrossRef] [PubMed]
- Lapidot, T.; Dar, A.; Kollet, O. How do stem cells find their way home? Blood 2005, 106, 1901–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suárez-Álvarez, B.; López-Vázquez, A.; López-Larrea, C. Mobilization and homing of hematopoietic stem cells. Adv. Exp. Med. Biol. 2012, 741, 152–170. [Google Scholar] [CrossRef]
- Broxmeyer, H.E.; Orschell, C.M.; Clapp, D.W.; Hangoc, G.; Cooper, S.; Plett, P.A.; Liles, W.C.; Li, X.; Graham-Evans, B.; Campbell, T.B.; et al. Rapid mobilization of murine and human hematopoietic stem and progenitor cells with AMD3100, a CXCR4 antagonist. J. Exp. Med. 2005, 201, 1307–1318. [Google Scholar] [CrossRef] [Green Version]
- Garrigós, M.M.; de Oliveira, F.A.; Nucci, M.P.; Nucci, L.P.; Alves, A.d.H.; Dias, O.F.M.; Gamarra, L.F. How mesenchymal stem cell cotransplantation with hematopoietic stem cells can improve engraftment in animal models. World J. Stem. Cells 2022, 14, 658–679. [Google Scholar] [CrossRef] [PubMed]
- Xaymardan, M.; Cimini, M.; Weisel, R.D.; Li, R.-K. 16-Bone Marrow Stem Cells: Properties and Pluripotency. In Principles of Regenerative Medicine; Atala, A., Lanza, R., Thomson, J.A., Nerem, R.M., Eds.; Academic Press: San Diego, CA, USA, 2008; pp. 268–283. [Google Scholar]
- Oliveira, F.A.; Nucci, M.P.; Filgueiras, I.S.; Ferreira, J.M.; Nucci, L.P.; Mamani, J.B.; Alvieri, F.; Souza, L.E.B.; Rego, G.N.A.; Kondo, A.T.; et al. Noninvasive Tracking of Hematopoietic Stem Cells in a Bone Marrow Transplant Model. Cells 2020, 9, 939. [Google Scholar] [CrossRef]
- Nucci, M.P.; Mamani, J.B.; Oliveira, F.A.; Filgueiras, I.S.; Alves, A.H.; Theinel, M.H.; Rodrigues, L.D.; Marti, L.; Gamarra, L.F. Optimization of Multimodal Nanoparticles Internalization Process in Mesenchymal Stem Cells for Cell Therapy Studies. Pharmaceutics 2022, 14, 1249. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Hu, Y.; Wang, J.; Gao, X.; Qian, X.; Tang, M. Superparamagnetic Iron Oxide Nanoparticles: Cytotoxicity, Metabolism, and Cellular Behavior in Biomedicine Applications. Int. J. Nanomed. 2021, 16, 6097–6113. [Google Scholar] [CrossRef] [PubMed]
- Jasmin; de Souza, G.T.; Louzada, R.A.; Rosado-de-Castro, P.H.; Mendez-Otero, R.; Campos de Carvalho, A.C. Tracking stem cells with superparamagnetic iron oxide nanoparticles: Perspectives and considerations. Int. J. Nanomed. 2017, 12, 779–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caspani, S.; Magalhães, R.; Araújo, J.P.; Sousa, C.T. Magnetic Nanomaterials as Contrast Agents for MRI. Materials 2020, 13, 2586. [Google Scholar] [CrossRef]
- Du, J.; Zhang, Y.; Jin, Z.; Wu, H.a.; Cang, J.; Shen, Y.; Miao, F.; Zhang, A.; Zhang, Y.; Zhang, J.; et al. Targeted NIRF/MR dual-mode imaging of breast cancer brain metastasis using BRBP1-functionalized ultra-small iron oxide nanoparticles. Mater. Sci. Eng. C 2020, 116, 111188. [Google Scholar] [CrossRef]
- Groult, H.; Ruiz-Cabello, J.; Pellico, J.; Lechuga-Vieco, A.V.; Bhavesh, R.; Zamai, M.; Almarza, E.; Martín-Padura, I.; Cantelar, E.; Martínez-Alcázar, M.P.; et al. Parallel multifunctionalization of nanoparticles: A one-step modular approach for in vivo imaging. Bioconjug. Chem. 2015, 26, 153–160. [Google Scholar] [CrossRef]
- Ghosh, S.; Ghosh, I.; Chakrabarti, M.; Mukherjee, A. Genotoxicity and biocompatibility of superparamagnetic iron oxide nanoparticles: Influence of surface modification on biodistribution, retention, DNA damage and oxidative stress. Food Chem. Toxicol. 2020, 136, 110989. [Google Scholar] [CrossRef]
- Becker, J.S.; Jakubowski, N. The synergy of elemental and biomolecular mass spectrometry: New analytical strategies in life sciences. Chem. Soc. Rev. 2009, 38, 1969–1983. [Google Scholar] [CrossRef]
- Souza, T.K.F.; Nucci, M.P.; Mamani, J.B.; da Silva, H.R.; Fantacini, D.M.C.; de Souza, L.E.B.; Picanço-Castro, V.; Covas, D.T.; Vidoto, E.L.; Tannús, A.; et al. Image and motor behavior for monitoring tumor growth in C6 glioma model. PLoS ONE 2018, 13, e0201453. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, H.; Kutner, R.H.; Bazan, N.G.; Reiser, J. Simplified lentivirus vector production in protein-free media using polyethylenimine-mediated transfection. J. Virol. Methods 2009, 157, 113–121. [Google Scholar] [CrossRef]
- da Silva, H.R.; Mamani, J.B.; Nucci, M.P.; Nucci, L.P.; Kondo, A.T.; Fantacini, D.M.C.; de Souza, L.E.B.; Picanço-Castro, V.; Covas, D.T.; Kutner, J.M.; et al. Triple-modal imaging of stem-cells labeled with multimodal nanoparticles, applied in a stroke model. World J. Stem. Cells 2019, 11, 100–123. [Google Scholar] [CrossRef] [PubMed]
- Mezzanotte, L.; van ‘t Root, M.; Karatas, H.; Goun, E.A.; Löwik, C.W.G.M. In Vivo Molecular Bioluminescence Imaging: New Tools and Applications. Trends Biotechnol. 2017, 35, 640–652. [Google Scholar] [CrossRef] [PubMed]
- Camargo, F.D.; Chambers, S.M.; Drew, E.; McNagny, K.M.; Goodell, M.A. Hematopoietic stem cells do not engraft with absolute efficiencies. Blood 2006, 107, 501–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caocci, G.; Greco, M.; La Nasa, G. Bone Marrow Homing and Engraftment Defects of Human Hematopoietic Stem and Progenitor Cells. Mediterr. J. Hematol. Infect Dis. 2017, 9, e2017032. [Google Scholar] [CrossRef] [Green Version]
- Duran-Struuck, R.; Dysko, R.C. Principles of bone marrow transplantation (BMT): Providing optimal veterinary and husbandry care to irradiated mice in BMT studies. J. Am. Assoc. Lab. Anim. Sci. 2009, 48, 11–22. [Google Scholar]
- Park, E.; Evans, M.A.; Doviak, H.; Horitani, K.; Ogawa, H.; Yura, Y.; Wang, Y.; Sano, S.; Walsh, K. Bone Marrow Transplantation Procedures in Mice to Study Clonal Hematopoiesis. J. Vis. Exp. 2021. [Google Scholar] [CrossRef]
- Rando, A.; de la Torre, M.; Martinez-Muriana, A.; Zaragoza, P.; Musaro, A.; Hernández, S.; Navarro, X.; Toivonen, J.M.; Osta, R. Chemotherapeutic agent 5-fluorouracil increases survival of SOD1 mouse model of ALS. PLoS ONE 2019, 14, e0210752. [Google Scholar] [CrossRef] [Green Version]
- Shaikh, A.; Bhartiya, D.; Kapoor, S.; Nimkar, H. Delineating the effects of 5-fluorouracil and follicle-stimulating hormone on mouse bone marrow stem/progenitor cells. Stem. Cell Res. Ther. 2016, 7, 59. [Google Scholar] [CrossRef] [Green Version]
- Focaccetti, C.; Bruno, A.; Magnani, E.; Bartolini, D.; Principi, E.; Dallaglio, K.; Bucci, E.O.; Finzi, G.; Sessa, F.; Noonan, D.M.; et al. Effects of 5-fluorouracil on morphology, cell cycle, proliferation, apoptosis, autophagy and ROS production in endothelial cells and cardiomyocytes. PLoS ONE 2015, 10, e0115686. [Google Scholar] [CrossRef]
- Sara, J.D.; Kaur, J.; Khodadadi, R.; Rehman, M.; Lobo, R.; Chakrabarti, S.; Herrmann, J.; Lerman, A.; Grothey, A. 5-fluorouracil and cardiotoxicity: A review. Ther. Adv. Med. Oncol. 2018, 10, 1758835918780140. [Google Scholar] [CrossRef] [Green Version]
- Sethy, C.; Kundu, C.N. 5-Fluorouracil (5-FU) resistance and the new strategy to enhance the sensitivity against cancer: Implication of DNA repair inhibition. Biomed. Pharmacother. 2021, 137, 111285. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.Y.; Hu, W.; Naramura, M.; Park, C.Y. High c-Kit expression identifies hematopoietic stem cells with impaired self-renewal and megakaryocytic bias. J. Exp. Med. 2014, 211, 217–231. [Google Scholar] [CrossRef] [Green Version]
- Szilvassy, S.J.; Cory, S. Phenotypic and functional characterization of competitive long-term repopulating hematopoietic stem cells enriched from 5-fluorouracil-treated murine marrow. Blood 1993, 81, 2310–2320. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.-Y.; Kim, J.-A.; Oh, I.-H. 5-Fluorouracil Treatment Leads to Activation of Stem Cell Niche By Reconstructing Mesenchymal Stromal Cells and Exert a Distinct Microenvironmental Impact on Normal and Leukemic Cells. Blood 2015, 126, 1197. [Google Scholar] [CrossRef]
- Edmundson, M.; Thanh, N.T.; Song, B. Nanoparticles based stem cell tracking in regenerative medicine. Theranostics 2013, 3, 573–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, F.A.; Nucci, M.P.; Mamani, J.B.; Alves, A.H.; Rego, G.N.A.; Kondo, A.T.; Hamerschlak, N.; Junqueira, M.S.; de Souza, L.E.B.; Gamarra, L.F. Multimodal Tracking of Hematopoietic Stem Cells from Young and Old Mice Labeled with Magnetic–Fluorescent Nanoparticles and Their Grafting by Bioluminescence in a Bone Marrow Transplant Model. Biomedicines 2021, 9, 752. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.; Nan, X.; Wang, Z.; Gao, L.; Xie, L.; Zou, C.; Wan, Q.; Pan, D.; Beauchamp, N.; Yang, X.; et al. Stem Cell Labeling with Superparamagnetic Iron Oxide Nanoparticles Using Focused Ultrasound and Magnetic Resonance Imaging Tracking. J. Nanosci. Nanotechnol. 2015, 15, 2605–2612. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-C.V.; Ku, M.-C.; Jayaseema, D.M.; Lai, J.-S.; Hueng, D.-Y.; Chang, C. Simple SPION Incubation as an Efficient Intracellular Labeling Method for Tracking Neural Progenitor Cells Using MRI. PLoS ONE 2013, 8, e56125. [Google Scholar] [CrossRef] [Green Version]
- Mamani, J.B.; Souza, T.K.F.; Nucci, M.P.; Oliveira, F.A.; Nucci, L.P.; Alves, A.H.; Rego, G.N.A.; Marti, L.; Gamarra, L.F. In Vitro Evaluation of Hyperthermia Magnetic Technique Indicating the Best Strategy for Internalization of Magnetic Nanoparticles Applied in Glioblastoma Tumor Cells. Pharmaceutics 2021, 13, 1219. [Google Scholar] [CrossRef]
- Bengtsson, N.E.; Kim, S.; Lin, L.; Walter, G.A.; Scott, E.W. Ultra-high-field MRI real-time imaging of HSC engraftment of the bone marrow niche. Leukemia 2011, 25, 1223–1231. [Google Scholar] [CrossRef] [Green Version]
- Niemeyer, M.; Oostendorp, R.A.J.; Kremer, M.; Hippauf, S.; Jacobs, V.R.; Baurecht, H.; Ludwig, G.; Piontek, G.; Bekker-Ruz, V.; Timmer, S.; et al. Non-invasive tracking of human haemopoietic CD34+ stem cells in vivo in immunodeficient mice by using magnetic resonance imaging. Eur. Radiol. 2010, 20, 2184–2193. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Xu, H.; Han, Y.; Dai, W.; Hao, W.; Wang, C.; Ning, G.; Xu, H.; Cao, J.-M. The internalization pathway, metabolic fate and biological effect of superparamagnetic iron oxide nanoparticles in the macrophage-like RAW264.7 cell. Sci. China Life Sci. 2011, 54, 793–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atilla, E.; Toprak, S.K.; Demirer, T. Current Review of Iron Overload and Related Complications in Hematopoietic Stem Cell Transplantation. Turk J. Haematol. 2017, 34, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rodea, G.E.; Montiel-Infante, F.X.; Cruz-Córdova, A.; Saldaña-Ahuactzi, Z.; Ochoa, S.A.; Espinosa-Mazariego, K.; Hernández-Castro, R.; Xicohtencatl-Cortes, J. Tracking Bioluminescent ETEC during In vivo BALB/c Mouse Colonization. Front. Cell Infect. Microbiol. 2017, 7, 187. [Google Scholar] [CrossRef] [Green Version]
- Oh, E.J.; Lee, H.W.; Kalimuthu, S.; Kim, T.J.; Kim, H.M.; Baek, S.H.; Zhu, L.; Oh, J.M.; Son, S.H.; Chung, H.Y.; et al. In vivo migration of mesenchymal stem cells to burn injury sites and their therapeutic effects in a living mouse model. J. Control. Release 2018, 279, 79–88. [Google Scholar] [CrossRef]
- Kim, J.E.; Kalimuthu, S.; Ahn, B.C. In vivo cell tracking with bioluminescence imaging. Nucl. Med. Mol. Imaging 2015, 49, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Bernau, K.; Lewis, C.M.; Petelinsek, A.M.; Benink, H.A.; Zimprich, C.A.; Meyerand, M.E.; Suzuki, M.; Svendsen, C.N. In vivo tracking of human neural progenitor cells in the rat brain using bioluminescence imaging. J. Neurosci. Methods 2014, 228, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Gobbo, O.L.; Wetterling, F.; Vaes, P.; Teughels, S.; Markos, F.; Edge, D.; Shortt, C.M.; Crosbie-Staunton, K.; Radomski, M.W.; Volkov, Y.; et al. Biodistribution and pharmacokinetic studies of SPION using particle electron paramagnetic resonance, MRI and ICP-MS. Nanomedicine 2015, 10, 1751–1760. [Google Scholar] [CrossRef] [Green Version]
- Lanzkron, S.M.; Collector, M.I.; Sharkis, S.J. Hematopoietic Stem Cell Tracking In Vivo: A Comparison of Short-Term and Long-Term Repopulating Cells. Blood 1999, 93, 1916–1921. [Google Scholar] [CrossRef]
- Maxwell, D.J.; Bonde, J.; Hess, D.A.; Hohm, S.A.; Lahey, R.; Zhou, P.; Creer, M.H.; Piwnica-Worms, D.; Nolta, J.A. Fluorophore-Conjugated Iron Oxide Nanoparticle Labeling and Analysis of Engrafting Human Hematopoietic Stem Cells. Stem Cells 2007, 26, 517–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavey, T.; Latour, C.; Island, M.-L.; Leroyer, P.; Guggenbuhl, P.; Coppin, H.; Roth, M.-P.; Bendavid, C.; Brissot, P.; Ropert, M.; et al. Spleen iron, molybdenum, and manganese concentrations are coregulated in hepcidin-deficient and secondary iron overload models in mice. FASEB J. 2019, 33, 11072–11081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Chen, L.; Li, R.; Dan, M.; Liu, H.; Wang, X.; Wu, X.; Liu, Y.; Xu, L.; Xie, L. Bio-distribution and bio-availability of silver and gold in rat tissues with silver/gold nanorod administration. RSC Adv. 2018, 8, 12260–12268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonavane, G.; Tomoda, K.; Makino, K. Biodistribution of colloidal gold nanoparticles after intravenous administration: Effect of particle size. Colloids Surf. B Biointerfaces 2008, 66, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Di Gregorio, E.; Iani, R.; Ferrauto, G.; Nuzzi, R.; Aime, S.; Gianolio, E. Gd accumulation in tissues of healthy mice upon repeated administrations of Gadodiamide and Gadoteridol. J. Trace Elem. Med. Biol. 2018, 48, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Narmani, A.; Farhood, B.; Haghi-Aminjan, H.; Mortezazadeh, T.; Aliasgharzadeh, A.; Mohseni, M.; Najafi, M.; Abbasi, H. Gadolinium nanoparticles as diagnostic and therapeutic agents: Their delivery systems in magnetic resonance imaging and neutron capture therapy. J. Drug Deliv. Sci. Technol. 2018, 44, 457–466. [Google Scholar] [CrossRef]
- Kim, S.J.; Lewis, B.; Steiner, M.S.; Bissa, U.V.; Dose, C.; Frank, J.A. Superparamagnetic iron oxide nanoparticles for direct labeling of stem cells and in vivo MRI tracking. Contrast Media Mol. Imaging 2016, 11, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Comenge, J.; Sharkey, J.; Fragueiro, O.; Wilm, B.; Brust, M.; Murray, P.; Levy, R.; Plagge, A. Multimodal cell tracking from systemic administration to tumour growth by combining gold nanorods and reporter genes. eLife 2018, 7, e33140. [Google Scholar] [CrossRef]
- Higuchi, T.; Anton, M.; Dumler, K.; Seidl, S.; Pelisek, J.; Saraste, A.; Welling, A.; Hofmann, F.; Oostendorp, R.A.; Gansbacher, B.; et al. Combined reporter gene PET and iron oxide MRI for monitoring survival and localization of transplanted cells in the rat heart. J. Nucl. Med. 2009, 50, 1088–1094. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Rosol, M.; Ge, S.; Peterson, D.; McNamara, G.; Pollack, H.; Kohn, D.B.; Nelson, M.D.; Crooks, G.M. Dynamic tracking of human hematopoietic stem cell engraftment using in vivo bioluminescence imaging. Blood 2003, 102, 3478–3482. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.-A.; Wagers, A.J.; Beilhack, A.; Dusich, J.; Bachmann, M.H.; Negrin, R.S.; Weissman, I.L.; Contag, C.H. Shifting foci of hematopoiesis during reconstitution from single stem cells. Proc. Natl. Acad. Sci. USA 2004, 101, 221–226. [Google Scholar] [CrossRef] [Green Version]
- Báječný, M.; Chen, C.-L.; Faltusová, K.; Heizer, T.; Szikszai, K.; Páral, P.; Šefc, L.; Nečas, E. Hematopoiesis Remains Permissive to Bone Marrow Transplantation After Expansion of Progenitors and Resumption of Blood Cell Production. Front. Cell Dev. Biol. 2021, 9, 660617. [Google Scholar] [CrossRef] [PubMed]
- Liebler, J.M.; Lutzko, C.; Banfalvi, A.; Senadheera, D.; Aghamohammadi, N.; Crandall, E.D.; Borok, Z. Retention of human bone marrow-derived cells in murine lungs following bleomycin-induced lung injury. Am. J. Physiol. Lung Cell Mol. Physiol. 2008, 295, L285–L292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwarteng, E.O.; Heinonen, K.M. Competitive Transplants to Evaluate Hematopoietic Stem Cell Fitness. J. Vis. Exp. 2016. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Javed, S.; Jain, S.K.; Majumdar, S.S.; Mukhopadhyay, A. Donor hematopoietic stem cells confer long-term marrow reconstitution by self-renewal divisions exceeding to that of host cells. PLoS ONE 2012, 7, e50693. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garrigós, M.M.; Oliveira, F.A.; Nucci, M.P.; Mamani, J.B.; Dias, O.F.M.; Rego, G.N.A.; Junqueira, M.S.; Costa, C.J.S.; Silva, L.R.R.; Alves, A.H.; et al. Bioluminescence Imaging and ICP-MS Associated with SPION as a Tool for Hematopoietic Stem and Progenitor Cells Homing and Engraftment Evaluation. Pharmaceutics 2023, 15, 828. https://doi.org/10.3390/pharmaceutics15030828
Garrigós MM, Oliveira FA, Nucci MP, Mamani JB, Dias OFM, Rego GNA, Junqueira MS, Costa CJS, Silva LRR, Alves AH, et al. Bioluminescence Imaging and ICP-MS Associated with SPION as a Tool for Hematopoietic Stem and Progenitor Cells Homing and Engraftment Evaluation. Pharmaceutics. 2023; 15(3):828. https://doi.org/10.3390/pharmaceutics15030828
Chicago/Turabian StyleGarrigós, Murilo M., Fernando A. Oliveira, Mariana P. Nucci, Javier B. Mamani, Olívia F. M. Dias, Gabriel N. A. Rego, Mara S. Junqueira, Cícero J. S. Costa, Lucas R. R. Silva, Arielly H. Alves, and et al. 2023. "Bioluminescence Imaging and ICP-MS Associated with SPION as a Tool for Hematopoietic Stem and Progenitor Cells Homing and Engraftment Evaluation" Pharmaceutics 15, no. 3: 828. https://doi.org/10.3390/pharmaceutics15030828