
Relevance of the Extraction Stage on the Anti-Inflammatory Action of Fucoidans
 , , , ,
, , , ,
Abstract
:1. Introduction
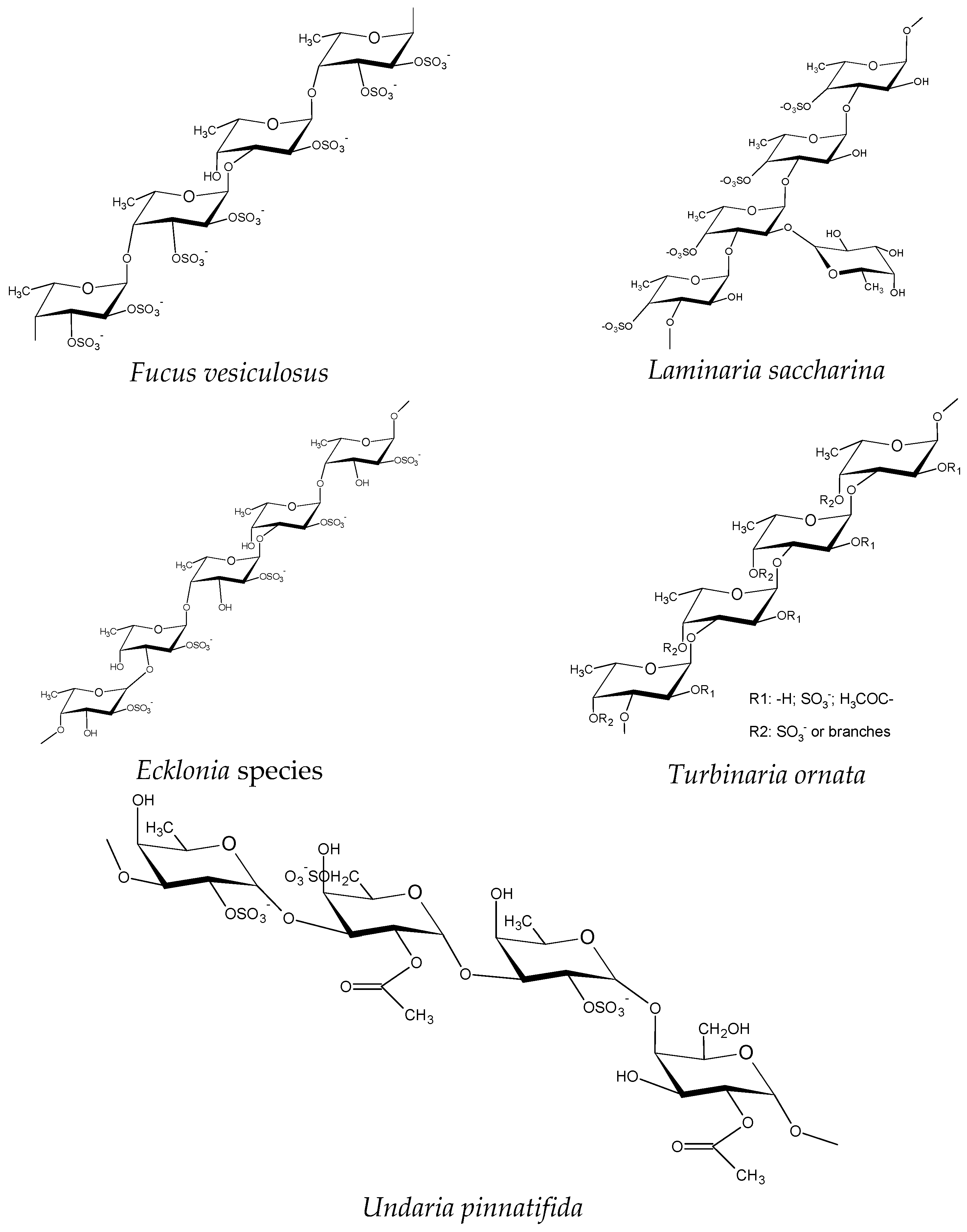
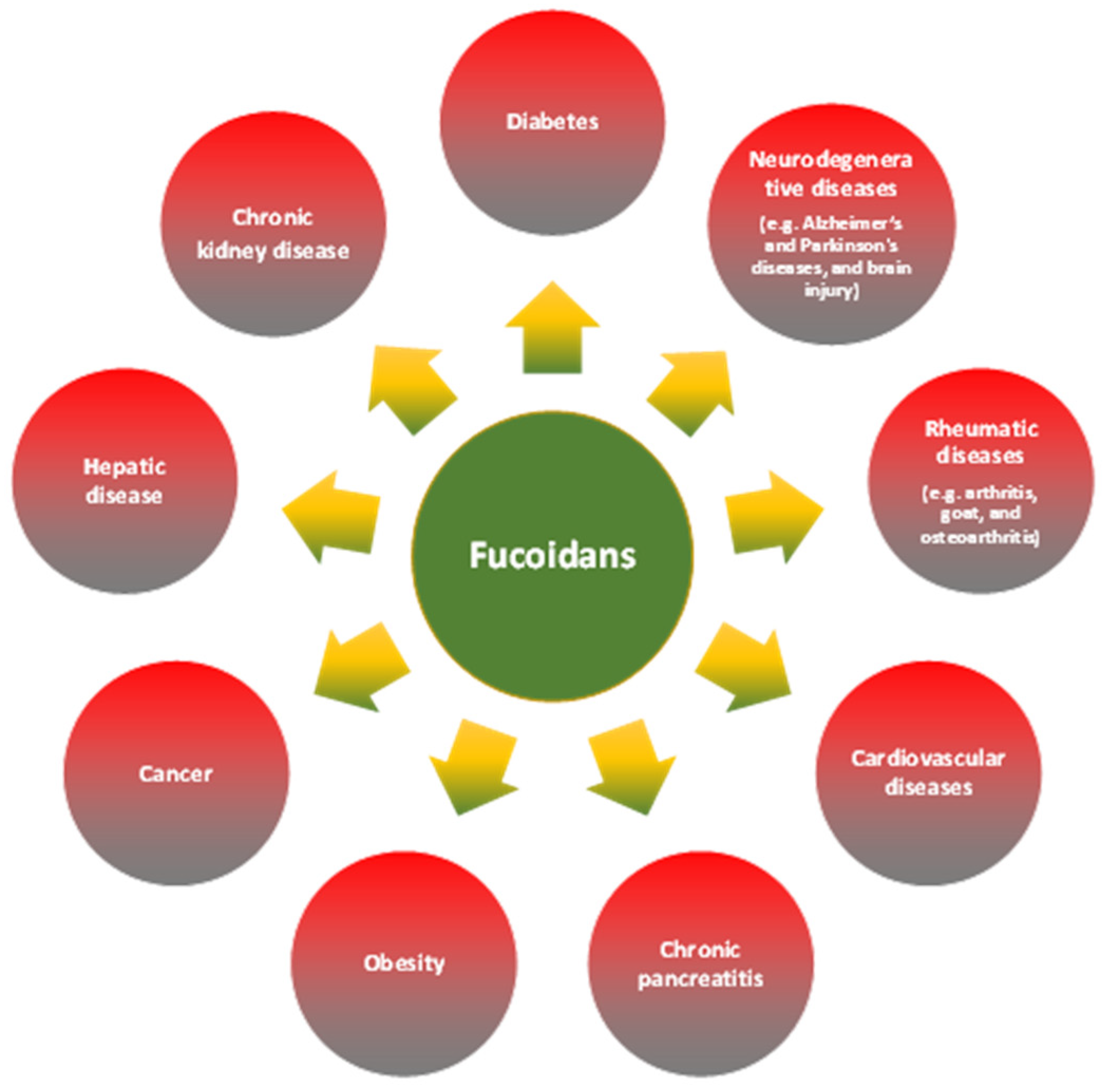
2. Fucoidans
3. Chronic Inflammation
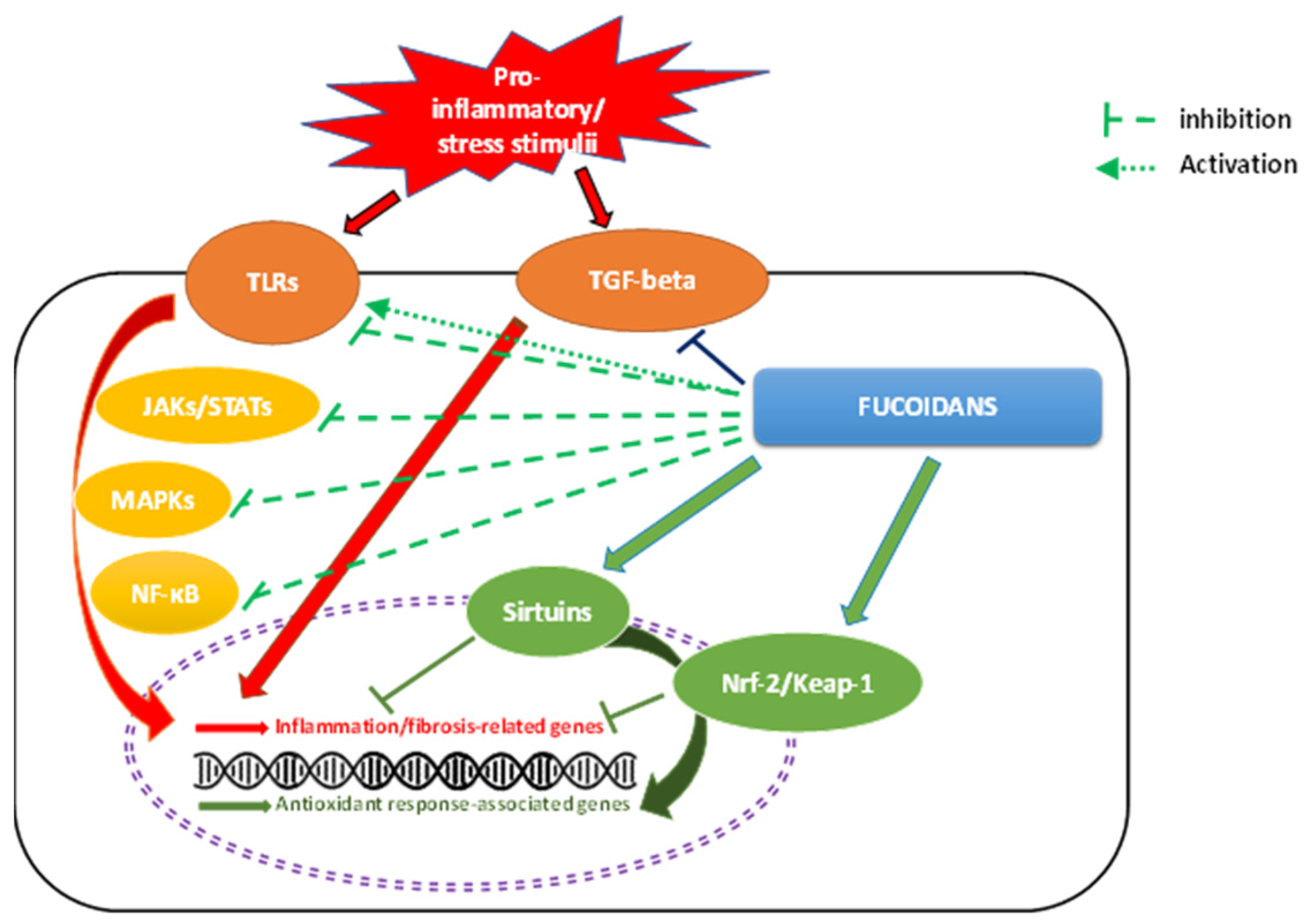
4. Mechanisms of Fucoidan Actions
4.1. NF-κB Signaling
4.2. MAPK Signaling Pathways
4.3. TLR Signaling Pathways
4.4. TGF-β1 Signaling Pathway
4.5. JAK–STAT Signaling Pathways
4.6. Nrf-2/Keap1 Signaling Pathway
4.7. Sirtuins
5. Major Fucoidan Features Influencing Anti-Inflammatory Activity
5.1. Effect of SCP Sulfate Content
5.2. Molecular Weight
5.3. Other Factors
6. Extraction and Purification
6.1. Conventional Extraction
6.1.1. Water Extraction
6.1.2. Alkaline Extraction

{kind=link}
{kind=link}
{kind=link}
| Aqueous Media | Seaweeds | References |
|---|---|---|
| Water, room temp., 2–24 h, 1–3 stages | Fucus vesiculosus,Sargassum stenophyllum | [44,140] |
| Water, 40–65 °C, 15 min,1 h, 1–2 stages | Sargassum cristaefolium, Sargassum wightii, Undaria pinnatifida | [12,126] |
| Water, 70–80 °C, 3–24 h, 1–3 stages | Ecklonia cava,Fucus vesiculosus,Hizikia fusiforme,Saccharina japonica | [18] |
| Water, 90–95 °C, 3–4 h | Chnoospora minima,Halimeda discoidea, Halimeda gracilis | [25] |
| Acidic Media | ||
| 0.05 M HCl, 25 °C, 2 h | Saccharina japonica | [135] |
| HCl (pH 2), 60–70 °C, 1–7 h, 1–9 stages | Dictyota dichotoma, Laminaria cichorioides, Padina sp., Sargassum binderi | [121,124,133,143] |
| 30% HCl, 100 °C, 15 min | Cladosiphon okamuranus | [144] |
| 0.01 M HCl or 0.001–0.1 M H2SO4, 60–80 °C | Fucus vesiculosus | [145] |
| 0.03–8 M HCl (pH 2), 90–100 °C, 0.25–4 h | Sargassum sp., Sargassum fulvellum | [131,146] |
| 0.15 M HCl, 45 °C, 3 h, 4 stages | Sargassum sp. | [141] |
| McIlvaine’s buffer solution (pH 4.0), 60 °C, 3 h | Sargassum sp. | [11] |
| HCl (pH 2), room temp., 24 h | Undaria pinnatifida | [147] |
| Alkaline Media | ||
| 1–2% CaCl2, room temp., 45–85 °C, 5–24 h, 1–6 stages | Ascophyllum nodosum, Fucus evanescens, Fucus dischitus, Fucus serratus, Fucus spiralis, Fucus vesiculosus, Laminaria digitata, Sargassum binderi, Sargassum sp., Undaria pinnatifida, | [117,118,124,129,132,133,141] |
| 4 M KOH, 10 mg NaBH4, room temp. | Sargassum stenophyllum | [142] |
6.1.3. Acid Extraction
6.2. Alternative Extraction Procedures
6.2.1. Ultrasound-Assisted Extraction
6.2.2. Microwave-Assisted Extraction
6.2.3. Enzyme-Assisted Extraction
6.2.4. Subcritical Water Extraction
7. Coatings and Micro- and Nanoencapsulation
8. Conclusions and Future Trends
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Pomin, V.H.; Mourão, P.A.S. Structure, Biology, Evolution, and Medical Importance of Sulfated Fucans and Galactans. Glycobiology 2008, 18, 1016–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holtkamp, A.D.; Kelly, S.; Ulber, R.; Lang, S. Fucoidans and Fucoidanases—Focus on Techniques for Molecular Structure Elucidation and Modification of Marine Polysaccharides. Appl. Microbiol. Biotechnol. 2009, 82, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kusaykin, M.; Bakunina, I.; Sova, V.; Ermakova, S.; Kuznetsova, T.; Besednova, N.; Zaporozhets, T.; Zvyagintseva, T. Structure, Biological Activity, and Enzymatic Transformation of Fucoidans from the Brown Seaweeds. Biotechnol. J. 2008, 3, 904–915. [Google Scholar] [CrossRef] [PubMed]
- Gideon, T.P.; Rengasamy, R. Toxicological Evaluation of Fucoidan from Cladosiphon okamuranus. J. Med. Food 2008, 11, 638–642. [Google Scholar] [CrossRef]
- Ha, J.-H.; Kwon, M.-C.; Han, J.-G.; Jin, L.; Jeong, H.S.; Choi, G.-P.; Park, U.-Y.; You, S.-G.; Lee, H.-Y. Enhancement of Immunomodulatory Activities of Low Molecular Weight Fucoidan Isolated from Hizikia fusiforme. Korean J. Food Sci. Technol. 2008, 40, 545–550. [Google Scholar]
- Usov, A.I.; Bilan, M.I. Fucoidans—Sulfated Polysaccharides of Brown Algae. Russ. Chem. Rev. 2009, 78, 785–799. [Google Scholar] [CrossRef]
- Apostolova, E.; Lukova, P.; Baldzhieva, A.; Katsarov, P.; Nikolova, M.; Iliev, I.; Peychev, L.; Trica, B.; Oancea, F.; Delattre, C.; et al. Immunomodulatory and Anti-inflammatory Effects of Fucoidan: A Review. Polymers 2020, 12, 2338. [Google Scholar] [CrossRef]
- Skriptsova, A.v.; Shevchenko, N.M.; Zvyagintseva, T.N.; Imbs, T.I. Monthly Changes in the Content and Monosaccharide Composition of Fucoidan from Undaria pinnatifida (Laminariales, Phaeophyta). J. Appl. Phycol. 2010, 22, 79–86. [Google Scholar] [CrossRef]
- Usov, A. Chemical Structures of Algal Polysaccharides. In Functional Ingredients from Algae for Foods and Nutraceuticals; Woodhead Publishing Limited: Cambridge, UK, 2013; pp. 23–86. [Google Scholar]
- Ananthi, S.; Raghavendran, H.R.; Sunil, A.G.; Gayathri, V.; Ramakrishnan, G.; Vasanthi, H.R. In Vitro Antioxidant and in Vivo Anti-Inflammatory Potential of Crude Polysaccharide from Turbinaria Ornata (Marine Brown Alga). Food Chem. Toxicol. 2010, 48, 187–192. [Google Scholar] [CrossRef]
- Hifney, A.F.; Fawzy, M.A.; Abdel-Gawad, K.M.; Gomaa, M. Industrial Optimization of Fucoidan Extraction from Sargassum sp. and Its Potential Antioxidant and Emulsifying Activities. Food Hydrocoll. 2016, 54, 77–88. [Google Scholar] [CrossRef]
- Yang, C.; Chung, D.; Shin, I.-S.; Lee, H.; Kim, J.; Lee, Y.; You, S. Effects of Molecular Weight and Hydrolysis Conditions on Anticancer Activity of Fucoidans from Sporophyll of Undaria pinnatifida. Int. J. Biol. Macromol. 2008, 43, 433–437. [Google Scholar] [CrossRef] [PubMed]
- Synytsya, A.; Kim, W.-J.; Kim, S.-M.; Pohl, R.; Synytsya, A.; Kvasnička, F.; Čopíková, J.; Park, Y. il Structure and Antitumour Activity of Fucoidan Isolated from Sporophyll of Korean Brown Seaweed Undaria pinnatifida. Carbohydr. Polym. 2010, 81, 41–48. [Google Scholar] [CrossRef]
- Foley, S.A.; Szegezdi, E.; Mulloy, B.; Samali, A.; Tuohy, M.G. An Unfractionated Fucoidan from Ascophyllum Nodosum: Extraction, Characterization, and Apoptotic Effects in vitro. J. Nat. Prod. 2011, 74, 1851–1861. [Google Scholar] [CrossRef] [PubMed]
- Santoyo, S.; Plaza, M.; Jaime, L.; Ibañez, E.; Reglero, G.; Señorans, J. Pressurized Liquids as an Alternative Green Process to Extract Antiviral Agents from the Edible Seaweed Himanthalia elongata. J. Appl. Phycol. 2011, 23, 909–917. [Google Scholar] [CrossRef] [Green Version]
- Luthuli, S.; Wu, S.; Cheng, Y.; Zheng, X.; Wu, M.; Tong, H. Therapeutic Effects of Fucoidan: A Review on Recent Studies. Mar. Drugs 2019, 17, 487. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.-M.; Kim, K.-N.; Lee, S.-H.; Ahn, G.; Cha, S.-H.; Kim, A.-D.; Yang, X.-D.; Kang, M.-C.; Jeon, Y.-J. Anti-Inflammatory Activity of Polysaccharide Purified from AMG-Assistant Extract of Ecklonia cava in LPS-Stimulated RAW 264.7 Macrophages. Carbohydr. Polym. 2011, 85, 80–85. [Google Scholar] [CrossRef]
- Lee, S.-H.; Ko, C.-I.; Ahn, G.; You, S.; Kim, J.-S.; Heu, M.S.; Kim, J.; Jee, Y.; Jeon, Y.-J. Molecular Characteristics and Anti-Inflammatory Activity of the Fucoidan Extracted from Ecklonia cava. Carbohydr. Polym. 2012, 89, 599–606. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.A.; Fernando, I.P.S.; Kim, E.-A.; Ahn, G.; Jee, Y.; Jeon, Y.-J. Anti-Inflammatory Activity of a Sulfated Polysaccharide Isolated from an Enzymatic Digest of Brown Seaweed Sargassum horneri in RAW 264.7 Cells. Nutr. Res. Pract. 2017, 11, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Liu, J.; Zhang, Y.; Song, J.; Zhang, Z.; Yang, Y.; Wu, M.; Tong, H. Structural Characterization and Antagonistic Effect against P-Selectin-Mediated Function of SFF-32, a Fucoidan Fraction from Sargassum fusiforme. J. Ethnopharmacol. 2022, 295, 115408. [Google Scholar] [CrossRef]
- Hwang, P.-A.; Chien, S.-Y.; Chan, Y.-L.; Lu, M.-K.; Wu, C.-H.; Kong, Z.-L.; Wu, C.-J. Inhibition of Lipopolysaccharide (LPS)-Induced Inflammatory Responses by Sargassum hemiphyllum Sulfated Polysaccharide Extract in RAW 264.7 Macrophage Cells. J. Agric. Food Chem. 2011, 59, 2062–2068. [Google Scholar] [CrossRef]
- Ahmad, T.; Eapen, M.S.; Ishaq, M.; Park, A.Y.; Karpiniec, S.S.; Stringer, D.N.; Sohal, S.S.; Fitton, J.H.; Guven, N.; Caruso, V.; et al. Anti-Inflammatory Activity of Fucoidan Extracts in vitro. Mar. Drugs 2021, 19, 702. [Google Scholar] [CrossRef]
- Wei, L.-Y.; Zhang, J.-K.; Zheng, L.; Chen, Y. The Functional Role of Sulforaphane in Intestinal Inflammation: A Review. Food Funct. 2022, 13, 514–529. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-J.; Yoon, K.-Y.; Lee, B.-Y. Low Molecular Weight Fucoidan from the Sporophyll of Undaria Pinnatifida Suppresses Inflammation by Promoting the Inhibition of Mitogen-Activated Protein Kinases and Oxidative Stress in RAW264.7 Cells. Fitoterapia 2012, 83, 1628–1635. [Google Scholar] [CrossRef] [PubMed]
- Fernando, I.S.; Sanjeewa, K.A.; Samarakoon, K.W.; Lee, W.W.; Kim, H.-S.; Kang, N.; Ranasinghe, P.; Lee, H.-S.; Jeon, Y.-J. A Fucoidan Fraction Purified from Chnoospora minima; a Potential Inhibitor of LPS-Induced Inflammatory Responses. Int. J. Biol. Macromol. 2017, 104, 1185–1193. [Google Scholar] [CrossRef] [PubMed]
- Lin, E.-T.; Lee, Y.-C.; Wang, H.-M.; Chiu, C.-Y.; Chang, Y.-K.; Huang, C.-Y.; Chang, C.-C.; Tsai, P.-C.; Chang, J.-S. Efficient Fucoidan Extraction and Purification from Sargassum cristaefolium and Preclinical Dermal Biological Activity Assessments of the Purified Fucoidans. J. Taiwan Inst. Chem. Eng. 2022, 137, 104294. [Google Scholar] [CrossRef]
- Wu, G.-J.; Shiu, S.-M.; Hsieh, M.-C.; Tsai, G.-J. Anti-Inflammatory Activity of a Sulfated Polysaccharide from the Brown Alga Sargassum cristaefolium. Food Hydrocoll. 2016, 53, 16–23. [Google Scholar] [CrossRef]
- Lee, S.M.; Son, K.-N.; Shah, D.; Ali, M.; Balasubramaniam, A.; Shukla, D.; Aakalu, V.K. Histatin-1 Attenuates LPS-Induced Inflammatory Signaling in RAW264.7 Macrophages. Int. J. Mol. Sci. 2021, 22, 7856. [Google Scholar] [CrossRef]
- Huwait, E.; Ayoub, M.; Karim, S. Investigation of the Molecular Mechanisms Underlying the Antiatherogenic Actions of Kaempferol in Human THP-1 Macrophages. Int. J. Mol. Sci. 2022, 23, 7461. [Google Scholar] [CrossRef]
- Phull, A.-R.; Majid, M.; Haq, I.; Khan, M.R.; Kim, S.J. In vitro and in vivo Evaluation of Anti-Arthritic, Antioxidant Efficacy of Fucoidan from Undaria pinnatifida (Harvey) Suringar. Int. J. Biol. Macromol. 2017, 97, 468–480. [Google Scholar] [CrossRef]
- Li, X.; Xin, S.; Zheng, X.; Lou, L.; Ye, S.; Li, S.; Wu, Q.; Ding, Q.; Ji, L.; Nan, C.; et al. Inhibition of the Occurrence and Development of Inflammation-Related Colorectal Cancer by Fucoidan Extracted from Sargassum fusiforme. J. Agric. Food Chem. 2022, 70, 9463–9476. [Google Scholar] [CrossRef]
- Ponce, N.M.A.; Stortz, C.A. A Comprehensive and Comparative Analysis of the Fucoidan Compositional Data Across the Phaeophyceae. Front. Plant. Sci. 2020, 11, 556312. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ni, L.; Fu, X.; Wang, L.; Duan, D.; Huang, L.; Xu, J.; Gao, X. Molecular Mechanism of Anti-Inflammatory Activities of a Novel Sulfated Galactofucan from Saccharina japonica. Mar. Drugs 2021, 19, 430. [Google Scholar] [CrossRef] [PubMed]
- Zayed, A.; El-Aasr, M.; Ibrahim, A.R.S.; Ulber, R. Fucoidan Characterization: Determination of Purity and Physicochemical and Chemical Properties. Mar. Drugs 2020, 18, 571. [Google Scholar] [CrossRef] [PubMed]
- Phull, A.R.; Kim, S.J. Fucoidan as Bio-Functional Molecule: Insights into the Anti-Inflammatory Potential and Associated Molecular Mechanisms. J. Funct. Foods 2017, 38, 415–426. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.A.; Herath, K.H.I.N.M.; Yang, H.-W.; Choi, C.S.; Jeon, Y.-J. Anti-Inflammatory Mechanisms of Fucoidans to Treat Inflammatory Diseases: A Review. Mar. Drugs 2021, 19, 678. [Google Scholar] [CrossRef] [PubMed]
- Holdt, S.L.; Kraan, S. Bioactive Compounds in Seaweed: Functional Food Applications and Legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Skriptsova, A.V.; Shevchenko, N.M.; Tarbeeva, D.V.; Zvyagintseva, T.N. Comparative Study of Polysaccharides from Reproductive and Sterile Tissues of Five Brown Seaweeds. Mar. Biotechnol. 2012, 14, 304–311. [Google Scholar] [CrossRef]
- Lim, S.J.; Wan mustapha, W. Extraction of Sulfated Polysaccharides (Fucoidan) From Brown Seaweed. In Seaweed Polysaccharides: Isolation, Biological and Biomedical Applications; Elsevier: Amsterdam, The Netherlands, 2017; pp. 27–46. [Google Scholar]
- Lahrsen, E.; Schoenfeld, A.K.; Alban, S. Size-Dependent Pharmacological Activities of Differently Degraded Fucoidan Fractions from Fucus Vesiculosus. Carbohydr. Polym. 2018, 189, 162–168. [Google Scholar] [CrossRef]
- Hemmingson, J.A.; Falshaw, R.; Furneaux, R.H.; Thompson, K. Structure and Antiviral Activity of the Galactofucan Sulfates Extracted from Undaria pinnatifida (Phaeophyta). J. Appl. Phycol. 2006, 18, 185. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Q.; Zhang, Z.; Song, H.; Li, P. Potential Antioxidant and Anticoagulant Capacity of Low Molecular Weight Fucoidan Fractions Extracted from Laminaria japonica. Int. J. Biol. Macromol. 2010, 46, 6–12. [Google Scholar] [CrossRef]
- De Paiva, A.A.O.; Castro, A.J.G.; Nascimento, M.S.; Will, L.S.E.P.; Santos, N.D.; Araújo, R.M.; Xavier, C.A.C.; Rocha, F.A.; Leite, E.L. Antioxidant and Anti-Inflammatory Effect of Polysaccharides from Lobophora variegata on Zymosan-Induced Arthritis in Rats. Int. Immunopharmacol. 2011, 11, 1241–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, M.E.R.; Cardoso, M.A.; Noseda, M.D.; Cerezo, A.S. Structural Studies on Fucoidans from the Brown Seaweed Sargassum stenophyllum. Carbohydr. Res. 2001, 333, 281–293. [Google Scholar] [CrossRef]
- Ale, M.T.; Mikkelsen, J.D.; Meyer, A.S. Important Determinants for Fucoidan Bioactivity: A Critical Review of Structure-Function Relations and Extraction Methods for Fucose-Containing Sulfated Polysaccharides from Brown Seaweeds. Mar. Drugs 2011, 9, 2106–2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varela, M.L.; Mogildea, M.; Moreno, I.; Lopes, A. Acute Inflammation and Metabolism. Inflammation 2018, 41, 1115–1127. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Origin and Physiological Roles of Inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic Inflammation in the Etiology of Disease across the Life Span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef]
- Arteaga-Henríquez, G.; Lugo-Marín, J.; Gisbert, L.; Setién-Ramos, I.; Martínez-Gallo, M.; Pujol-Borrell, R.; Ramos-Quiroga, J.A. Activation of the Monocyte/Macrophage System and Abnormal Blood Levels of Lymphocyte Subpopulations in Individuals with Autism Spectrum Disorder: A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2022, 23, 14329. [Google Scholar] [CrossRef]
- Zizmare, L.; Mehling, R.; Gonzalez-Menendez, I.; Lonati, C.; Quintanilla-Martinez, L.; Pichler, B.J.; Kneilling, M.; Trautwein, C. Acute and Chronic Inflammation Alter Immunometabolism in a Cutaneous Delayed-Type Hypersensitivity Reaction (DTHR) Mouse Model. Commun. Biol. 2022, 5, 1250. [Google Scholar] [CrossRef]
- Wynn, T.A.; Ramalingam, T.R. Mechanisms of Fibrosis: Therapeutic Translation for Fibrotic Disease. Nat. Med. 2012, 18, 1028–1040. [Google Scholar] [CrossRef] [Green Version]
- Öz, H.H.; Cheng, E.C.; di Pietro, C.; Tebaldi, T.; Biancon, G.; Zeiss, C.; Zhang, P.X.; Huang, P.H.; Esquibies, S.S.; Britto, C.J.; et al. Recruited Monocytes/Macrophages Drive Pulmonary Neutrophilic Inflammation and Irreversible Lung Tissue Remodeling in Cystic Fibrosis. Cell. Rep. 2022, 41, 111797. [Google Scholar] [CrossRef]
- Zuo, L.; Prather, E.R.; Stetskiv, M.; Garrison, D.E.; Meade, J.R.; Peace, T.I.; Zhou, T. Inflammaging and Oxidative Stress in Human Diseases: From Molecular Mechanisms to Novel Treatments. Int. J. Mol. Sci. 2019, 20, 4472. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Tu, Y.; Long, Z.; Liu, J.; Kong, D.; Peng, J.; Wu, H.; Zheng, G.; Zhao, J.; Chen, Y.; et al. Reactive Oxygen Species Bridge the Gap between Chronic Inflammation and Tumor Development. Oxid. Med. Cell. Longev. 2022, 2022, 2606928. [Google Scholar] [CrossRef] [PubMed]
- Kiani, A.K.; Bonetti, G.; Donato, K.; Bertelli, M. Dietary Supplements for Intestinal Inflammation. J. Prev. Med. Hyg. 2022, 63, E214–E220. [Google Scholar] [PubMed]
- Andrianova, I.A.; Khabirova, A.I.; Ponomareva, A.A.; Peshkova, A.D.; Evtugina, N.G.; le Minh, G.; Sibgatullin, T.B.; Weisel, J.W.; Litvinov, R.I. Chronic Immune Platelet Activation Is Followed by Platelet Refractoriness and Impaired Contractility. Int. J. Mol. Sci. 2022, 23, 7336. [Google Scholar] [CrossRef] [PubMed]
- Vieceli Dalla Sega, F.; Cimaglia, P.; Manfrini, M.; Fortini, F.; Marracino, L.; Bernucci, D.; Pompei, G.; Scala, A.; Trichilo, M.; de Carolis, B.; et al. Circulating Biomarkers of Endothelial Dysfunction and Inflammation in Predicting Clinical Outcomes in Diabetic Patients with Critical Limb Ischemia. Int. J. Mol. Sci. 2022, 23, 10641. [Google Scholar] [CrossRef]
- Vahid, F.; Jalili, M.; Rahmani, W.; Nasiri, Z.; Bohn, T. A Higher Healthy Eating Index Is Associated with Decreased Markers of Inflammation and Lower Odds for Being Overweight/Obese Based on a Case-Control Study. Nutrients 2022, 14, 5127. [Google Scholar] [CrossRef]
- Vargas-Mendoza, N.; Morales-González, Á.; Osiris Madrigal-Santillán, E.; Angeles-Valencia, M.; Anguiano-Robledo, L.; Ligia González-López, L.; Sosa-Gómez, A.; Fregoso-Aguilar, T.; Esquivel-Chirino, C.; Anahí Ruiz-Velazco-Benítez, Y.; et al. Review Article Phytochemicals and Modulation of Exercise-Induced Oxidative Stress: A Novel Overview of Antioxidants. Am. J. Transl. Res. 2022, 14, 8292–8314. [Google Scholar]
- Du, Y.; Zhu, Y.J.; Zhou, Y.X.; Ding, J.; Liu, J.Y. Metformin in Therapeutic Applications in Human Diseases: Its Mechanism of Action and Clinical Study. Mol. Biomed. 2022, 3, 41. [Google Scholar] [CrossRef]
- Martin, J.R.; Yu, M.; Erstad, B.L. Adverse Effects of Nonsteroidal Anti-Inflammatory Drugs in Critically Ill Patients: A Scoping Review. Am. J. Health-Syst. Pharm. 2022. ahead of print. [Google Scholar] [CrossRef]
- Ward, N.C.; Watts, G.F.; Eckel, R.H. Statin Toxicity: Mechanistic Insights and Clinical Implications. Circ. Res. 2019, 124, 328–350. [Google Scholar] [CrossRef]
- Negreanu-Pirjol, B.-S.; Negreanu-Pirjol, T.; Popoviciu, D.R.; Anton, R.-E.; Prelipcean, A.-M. Marine Bioactive Compounds Derived from Macroalgae as New Potential Players in Drug Delivery Systems: A Review. Pharmaceutics 2022, 14, 1781. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Q.; Han, X.; Ma, Y.; Zhang, Z.; Zhao, L.; Guan, F.; Ma, S. Fucoidan: A Promising Agent for Brain Injury and Neurodegenerative Disease Intervention. Food Funct. 2021, 12, 3820–3830. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Kawaguchi, M.; Kitamura, K.; Narumiya, S.; Kawamura, M.; Tengan, I.; Nishimoto, S.; Hanamure, Y.; Majima, Y.; Tsubura, S.; et al. An Exploratory Study on the Anti-Inflammatory Effects of Fucoidan in Relation to Quality of Life in Advanced Cancer Patients. Integr. Cancer Ther. 2018, 17, 282–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahan, M.S.; Hasan, A.; Rahman, M.D.H.; Meem, K.N.; Moni, A.; Hannan, M.A.; Uddin, M.J. Protective Effects of Fucoidan against Kidney Diseases: Pharmacological Insights and Future Perspectives. Int. J. Biol. Macromol. 2022, 209, 2119–2129. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Xu, J.; Ge, K.; Tian, Q.; Zhao, P.; Guo, Y. Anti-Inflammatory Effect of Low Molecular Weight Fucoidan from Saccharina japonica on Atherosclerosis in ApoE-Knockout Mice. Int. J. Biol. Macromol. 2018, 118, 365–374. [Google Scholar] [CrossRef]
- Pomin, V.H. Sulfated Glycans in Inflammation. Eur J Med Chem 2015, 92, 353–369. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-ΚB Signaling in Inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Yang, X.; Yuan, P.; Cai, S.; Bao, J.; Zhao, Y.; Aimaier, A.; Aipire, A.; Lu, J.; Li, J. In vitro and in vivo Dendritic Cell Immune Stimulation Effect of Low Molecular Weight Fucoidan from New Zealand Undaria pinnatifida. Mar. Drugs 2022, 20, 197. [Google Scholar] [CrossRef]
- Wang, L.; Cui, Y.R.; Wang, K.; Fu, X.; Xu, J.; Gao, X.; Jeon, Y.-J. Anti-Inflammatory Effect of Fucoidan Isolated from Fermented Sargassum fusiforme in in vitro and in vivo Models. Int. J. Biol. Macromol. 2022, 222, 2065–2071. [Google Scholar] [CrossRef]
- Liyanage, N.M.; Lee, H.-G.; Nagahawatta, D.P.; Jayawardhana, H.H.A.C.K.; Ryu, B.; Jeon, Y.-J. Characterization and Therapeutic Effect of Sargassum coreanum Fucoidan That Inhibits Lipopolysaccharide-Induced Inflammation in RAW 264.7 Macrophages by Blocking NF-ΚB Signaling. Int. J. Biol. Macromol. 2022, 223, 500–510. [Google Scholar] [CrossRef]
- Vaamonde-García, C.; Flórez-Fernández, N.; Torres, M.D.; Lamas-Vázquez, M.J.; Blanco, F.J.; Domínguez, H.; Meijide-Faílde, R. Study of Fucoidans as Natural Biomolecules for Therapeutical Applications in Osteoarthritis. Carbohydr. Polym. 2021, 258, 117692. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Guo, C.; Wang, Y.; Su, M.; Huang, W.; Lai, K.P. Preclinical Insights into Fucoidan as a Nutraceutical Compound against Perfluorooctanoic Acid-Associated Obesity via Targeting Endoplasmic Reticulum Stress. Front. Nutr. 2022, 9, 950130. [Google Scholar] [CrossRef] [PubMed]
- El-Far, Y.M.; Khodir, A.E.; Emarah, Z.A.; Ebrahim, M.A.; Al-Gayyar, M.M.H. Fucoidan Ameliorates Hepatocellular Carcinoma Induced in Rats: Effect on MiR143 and Inflammation. Nutr. Cancer 2021, 73, 1498–1510. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, Y.; Wang, Z.; Guo, L.; Li, X. Fucoidan Mitigated Diabetic Nephropathy through the Downregulation of PKC and Modulation of NF-ΚB Signaling Pathway: In vitro and in vivo Investigations. Phytother. Res. 2021, 35, 2133–2144. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Zhu, M.; Wu, J.; Yu, T.; Chen, Y.; Ye, X.; Li, S.; Xu, N. Correction: Fucoidans from Cucumaria frondosa Ameliorate Renal Interstitial Fibrosis via Inhibition of the PI3K/Akt/NF-ΚB Signaling Pathway. Food Funct. 2022, 13, 3776–3777. [Google Scholar] [CrossRef] [PubMed]
- Soares-Silva, M.; Diniz, F.F.; Gomes, G.N.; Bahia, D. The Mitogen-Activated Protein Kinase (MAPK) Pathway: Role in Immune Evasion by Trypanosomatids. Front. Microbiol. 2016, 7, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, D.K. MAP Kinase Pathways. Cold Spring Harb. Perspect. Biol. 2012, 4, a011254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jayasinghe, A.M.K.; Kirindage, K.G.I.S.; Fernando, I.P.S.; Han, E.J.; Oh, G.W.; Jung, W.K.; Ahn, G. Fucoidan Isolated from Sargassum confusum Suppresses Inflammatory Responses and Oxidative Stress in TNF-α/IFN-γ-Stimulated HaCaT Keratinocytes by Activating Nrf2/HO-1 Signaling Pathway. Mar. Drugs 2022, 20, 117. [Google Scholar] [CrossRef] [PubMed]
- Nagahawatta, D.P.; Liyanage, N.M.; Jayawardhana, H.H.A.C.K.; Lee, H.-G.; Jayawardena, T.U.; Jeon, Y.-J. Anti-Fine Dust Effect of Fucoidan Extracted from Ecklonia maxima Leaves in Macrophages via Inhibiting Inflammatory Signaling Pathways. Mar. Drugs 2022, 20, 413. [Google Scholar] [CrossRef]
- Do, H.; Kang, N.-S.; Pyo, S.; Billiar, T.; Sohn, E.-H. Differential Regulation by Fucoidan of IFN-Gamma-Induced NO Production in Glial Cells and Macrophages. J. Cell. Biochem. 2010, 111, 1337–1345. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-like Receptor Signaling Pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, Y.; Iwaihara, Y.; Bak, J.; Nakano, H.; Takeuchi, S.; Takeuchi, H.; Matsui, T.; Tachikawa, D. The Cooperative Induction of Macrophage Activation by Fucoidan Derived from Cladosiphon okamuranus and β-Glucan Derived from Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2019, 516, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Makarenkova, I.D.; Logunov, D.Y.; Tukhvatulin, A.I.; Semenova, I.B.; Besednova, N.N.; Zvyagintseva, T.N. Immunology and Microbiology Interactions between Sulfated Polysaccharides from Sea Brown Algae and Toll-Like Receptors on HEK293 Eukaryotic Cells in vitro. Bull. Exp. Biol. Med. 2012, 154, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, L.; Yan, C.; Ai, C.; Wen, C.; Guo, X.; Song, S. Two Ascophyllum nodosum Fucoidans with Different Molecular Weights Inhibit Inflammation via Blocking of TLR/NF-κB Signaling Pathway Discriminately. Foods 2022, 11, 2381. [Google Scholar] [CrossRef]
- Stolfi, C.; Troncone, E.; Marafini, I.; Monteleone, G. Role of Tgf-Beta and Smad7 in Gut Inflammation, Fibrosis and Cancer. Biomolecules 2021, 11, 17. [Google Scholar] [CrossRef]
- Yoshimura, A.; Wakabayashi, Y.; Mori, T. Cellular and Molecular Basis for the Regulation of Inflammation by TGF-β. J. Biochem. 2010, 147, 781–792. [Google Scholar] [CrossRef]
- Han, G.; Li, F.; Singh, T.P.; Wolf, P.; Wang, X.-J. The Pro-Inflammatory Role of TGFβ1: A Paradox? Int. J. Biol. Sci. 2012, 8, 228–235. [Google Scholar] [CrossRef] [Green Version]
- Piñeiro-Ramil, M.; Flórez-Fernández, N.; Ramil-Gómez, O.; Torres, M.D.; Dominguez, H.; Blanco, F.J.; Meijide-faílde, R.; Vaamonde-García, C. Antifibrotic Effect of Brown Algae-Derived Fucoidans on Osteoarthritic Fibroblast-like Synoviocytes. Carbohydr. Polym. 2022, 282, 119134. [Google Scholar] [CrossRef]
- Hong, S.-W.; Jung, K.H.; Lee, H.-S.; Zheng, H.-M.; Choi, M.-J.; Lee, C.; Hong, S.-S. Suppression by Fucoidan of Liver Fibrogenesis via the TGF-β/Smad Pathway in Protecting against Oxidative Stress. Biosci. Biotechnol. Biochem. 2011, 75, 833–840. [Google Scholar] [CrossRef]
- Yu, W.-C.; Huang, R.-Y.; Chou, T.-C. Oligo-Fucoidan Improves Diabetes-Induced Renal Fibrosis via Activation of Sirt-1, GLP-1R, and Nrf2/HO-1: An in vitro and in vivo Study. Nutrients 2020, 12, 3068. [Google Scholar] [CrossRef]
- Li, X.; Wu, N.; Chen, Y.; Tan, J.; Wang, J.; Geng, L.; Qin, Y.; Zhang, Q. Degradation of Different Molecular Weight Fucoidans and Their Inhibition of TGF-Β1 Induced Epithelial–Mesenchymal Transition in Mouse Renal Tubular Epithelial Cells. Int. J. Biol. Macromol. 2020, 151, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Deng, M.; He, X.; Li, M.; Li, J.; He, P.; Liu, H.; Li, M.; Zhang, Z.; He, Q. Fucoidan-Functionalized Activated Platelet-Hitchhiking Micelles Simultaneously Track Tumor Cells and Remodel the Immunosuppressive Microenvironment for Efficient Metastatic Cancer Treatment. Acta Pharm. Sin. B 2022, 12, 467–482. [Google Scholar] [CrossRef] [PubMed]
- Seif, F.; Khoshmirsafa, M.; Aazami, H.; Mohsenzadegan, M.; Sedighi, G.; Bahar, M. The Role of JAK-STAT Signaling Pathway and Its Regulators in the Fate of T Helper Cells. Cell. Commun. Signal 2017, 15, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, D.A. The JAK/STAT Pathway. Cold Spring Harb. Perspect. Biol. 2012, 4, a011205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Do, H.; Pyo, S.; Sohn, E.-H. Suppression of INOS Expression by Fucoidan Is Mediated by Regulation of P38 MAPK, JAK/STAT, AP-1 and IRF-1, and Depends on up-Regulation of Scavenger Receptor B1 Expression in TNF-α- and IFN-γ-Stimulated C6 Glioma Cells. J. Nutr. Biochem. 2010, 21, 671–679. [Google Scholar] [CrossRef]
- Sun, Y.; Xu, M.; Wang, C.; Guan, S.; Wang, L.; Cong, B.; Zhu, W.; Xu, Y. Low-Molecular-Weight Fucoidan Bidirectionally Regulates Lipid Uptake and Cholesterol Efflux through the P38 MAPK Phosphorylation. Int. J. Biol. Macromol. 2022, 220, 371–384. [Google Scholar] [CrossRef]
- Ye, J.; Chen, D.; Ye, Z.; Huang, Y.; Zhang, N.; Lui, E.M.K.; Xue, C.; Xiao, M. Fucoidan Isolated from Saccharina japonica Inhibits LPS-Induced Inflammation in Macrophages via Blocking NF-ΚB, MAPK and JAK-STAT Pathways. Mar. Drugs 2020, 18, 328. [Google Scholar] [CrossRef]
- Yang, J.; Yang, X.; Pan, W.; Wang, M.; Lu, Y.; Zhang, J.; Fang, Z.; Zhang, X.; Ji, Y.; Bei, J.X.; et al. Fucoidan-Supplemented Diet Potentiates Immune Checkpoint Blockage by Enhancing Antitumor Immunity. Front. Cell. Dev. Biol. 2021, 9, 733246. [Google Scholar] [CrossRef]
- Vaamonde-Garcia, C.; Courties, A.; Pigenet, A.; Laiguillon, M.C.; Sautet, A.; Houard, X.; Kerdine-Römer, S.; Meijide, R.; Berenbaum, F.; Sellam, J. The Nuclear Factor-Erythroid 2-Related Factor/Heme Oxygenase-1 Axis Is Critical for the Inflammatory Features of Type 2 Diabetes–Associated Osteoarthritis. J. Biol. Chem. 2017, 292, 14505–14515. [Google Scholar] [CrossRef] [Green Version]
- Davinelli, S.; Medoro, A.; Intrieri, M.; Saso, L.; Scapagnini, G.; Kang, J.X. Targeting NRF2–KEAP1 Axis by Omega-3 Fatty Acids and Their Derivatives: Emerging Opportunities against Aging and Diseases. Free Radic. Biol. Med. 2022, 193, 736–750. [Google Scholar] [CrossRef]
- Ma, Z.; Yang, Z.; Feng, X.; Deng, J.; He, C.; Li, R.; Zhao, Y.; Ge, Y.; Zhang, Y.; Song, C.; et al. The Emerging Evidence for a Protective Role of Fucoidan from Laminaria japonica in Chronic Kidney Disease-Triggered Cognitive Dysfunction. Mar. Drugs 2022, 20, 258. [Google Scholar] [CrossRef] [PubMed]
- Vaamonde-García, C.; Capelo-Mera, E.; Flórez-Fernández, N.; Torres, M.D.; Rivas-Murias, B.; Mejide-Faílde, R.; Blanco, F.J.; Domínguez, H. In vitro Study of the Therapeutic Potential of Brown Crude Fucoidans in Osteoarthritis Treatment. Int. J. Mol. Sci. 2022, 23, 14236. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Tian, Y.; Sui, Y.; Zhao, H.; Gao, H.; Liang, H.; Qiu, X.; Sun, Z.; Zhang, Y.; Qin, Y. Protective Effect of Fucoidan against Iron Overload and Ferroptosis-Induced Liver Injury in Rats Exposed to Alcohol. Biomed. Pharmacother. 2022, 153, 113402. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, Y.; Wang, Y.; Chao, Y.; Zhang, J.; Jia, Y.; Tie, J.; Hu, D. Regulation of SIRT1 and Its Roles in Inflammation. Front. Immunol. 2022, 13, 831168. [Google Scholar] [CrossRef]
- Vachharajani, V.T.; Liu, T.; Wang, X.; Hoth, J.J.; Yoza, B.K.; McCall, C.E. Sirtuins Link Inflammation and Metabolism. J. Immunol. Res. 2016, 2016, 8167273. [Google Scholar] [CrossRef] [Green Version]
- Grabowska, W.; Sikora, E.; Bielak-Zmijewska, A. Sirtuins, a Promising Target in Slowing down the Ageing Process. Biogerontology 2017, 18, 447–476. [Google Scholar] [CrossRef] [Green Version]
- Akter, R.; Afrose, A.; Rahman, M.R.; Chowdhury, R.; Nirzhor, S.S.R.; Khan, R.I.; Kabir, M.T. A Comprehensive Analysis into the Therapeutic Application of Natural Products as SIRT6 Modulators in Alzheimer’s Disease, Aging, Cancer, Inflammation, and Diabetes. Int. J. Mol. Sci. 2021, 22, 4180. [Google Scholar] [CrossRef]
- Zheng, Y.; Liu, T.; Wang, Z.; Xu, Y.; Zhang, Q.; Luo, D. Low Molecular Weight Fucoidan Attenuates Liver Injury via SIRT1/AMPK/PGC1α Axis in Db/Db Mice. Int. J. Biol. Macromol. 2018, 112, 929–936. [Google Scholar] [CrossRef]
- Dimitrova-Shumkovska, J.; Krstanoski, L.; Veenman, L. Potential Beneficial Actions of Fucoidan in Brain and Liver Injury, Disease, and Intoxication—Potential Implication of Sirtuins. Mar. Drugs 2020, 18, 242. [Google Scholar] [CrossRef]
- Kuznetsova, T.A.; Ivanushko, L.A.; Persiyanova, E.v.; Ermakova, S.P.; Besednova, N.N. Markers of Systemic Inflammation in Experimental Dyslipidemia Induced by P-407: Modulation with Fucoidan from Brown Alga Fucus evanescens. Bull. Exp. Biol. Med. 2019, 166, 766–769. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Wang, S.-H.; Huang, C.-Y.; Dong, C.-D.; Huang, C.-Y.; Chang, C.-C.; Chang, J.-S. Effect of Molecular Mass and Sulfate Content of Fucoidan from Sargassum siliquosum on Antioxidant, Anti-Lipogenesis, and Anti-Inflammatory Activity. J. Biosci. Bioeng. 2021, 132, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Rioux, L.-E.; Turgeon, S.L.; Beaulieu, M. Characterization of Polysaccharides Extracted from Brown Seaweeds. Carbohydr. Polym. 2007, 69, 530–537. [Google Scholar] [CrossRef]
- Park, S.-B.; Chun, K.-R.; Kim, J.-K.; Suk, K.; Jung, Y.-M.; Lee, W.-H. The Differential Effect of High and Low Molecular Weight Fucoidans on the Severity of Collagen-Induced Arthritis in Mice. Phytother. Res. 2010, 24, 1384–1391. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.-R.; Hsu, K.-T.; Li, T.-L.; Chan, Y.-L.; Wu, C.-J. Topical Application of Fucoidan Derived from Cladosiphon okamuranus Alleviates Atopic Dermatitis Symptoms through Immunomodulation. Int. Immunopharmacol. 2021, 101, 108362. [Google Scholar] [CrossRef]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I.; et al. A Comparative Study of the Anti-Inflammatory, Anticoagulant, Antiangiogenic, and Antiadhesive Activities of Nine Different Fucoidans from Brown Seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [CrossRef] [Green Version]
- Bilan, M.I.; Grachev, A.A.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. Structure of a Fucoidan from the Brown Seaweed Fucus serratus L. Carbohydr. Res. 2006, 341, 238–245. [Google Scholar] [CrossRef]
- Liu, L.; Heinrich, M.; Myers, S.; Dworjanyn, S.A. Towards a Better Understanding of Medicinal Uses of the Brown Seaweed Sargassum in Traditional Chinese Medicine: A Phytochemical and Pharmacological Review. J. Ethnopharmacol. 2012, 142, 591–619. [Google Scholar] [CrossRef]
- Ale, M.T.; Meyer, A.S. Fucoidans from Brown Seaweeds: An Update on Structures, Extraction Techniques and Use of Enzymes as Tools for Structural Elucidation. RSC Adv. 2013, 3, 8131–8141. [Google Scholar] [CrossRef] [Green Version]
- Anastyuk, S.D.; Shevchenko, N.M.; Nazarenko, E.L.; Imbs, T.I.; Gorbach, V.I.; Dmitrenok, P.S.; Zvyagintseva, T.N. Structural Analysis of a Highly Sulfated Fucan from the Brown Alga Laminaria cichorioides by Tandem MALDI and ESI Mass Spectrometry. Carbohydr. Res. 2010, 345, 2206–2212. [Google Scholar] [CrossRef]
- Flórez-Fernández, N.; González-Muñoz, M.J.; Domínguez, H. Feasibility of Posthydrolysis Processing of Hydrothermal Extracts from Sargassum muticum. Algal Res. 2017, 27, 73–81. [Google Scholar] [CrossRef]
- Hahn, T.; Lang, S.; Ulber, R.; Muffler, K. Novel Procedures for the Extraction of Fucoidan from Brown Algae. Process Biochem. 2012, 47, 1691–1698. [Google Scholar] [CrossRef]
- Imbs, T.I.; Skriptsova, A.v.; Zvyagintseva, T.N. Antioxidant Activity of Fucose-Containing Sulfated Polysaccharides Obtained from Fucus evanescens by Different Extraction Methods. J. Appl. Phycol. 2015, 27, 545–553. [Google Scholar] [CrossRef]
- Anastyuk, S.D.; Imbs, T.I.; Dmitrenok, P.S.; Zvyagintseva, T.N. Rapid Mass Spectrometric Analysis of a Novel Fucoidan, Extracted from the Brown Alga Coccophora langsdorfii. Sci. World J. 2014, 2014, 972450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinoth Kumar, T.; Lakshmanasenthil, S.; Geetharamani, D.; Marudhupandi, T.; Suja, G.; Suganya, P.; Kumar, T.V.; Lakshmanasenthil, S.; Geetharamani, D.; Marudhupandi, T.; et al. Fucoidan—A α-d-Glucosidase Inhibitor from Sargassum wightii with Relevance to Type 2 Diabetes Mellitus Therapy. Int. J. Biol. Macromol. 2015, 72, 1044–1047. [Google Scholar] [CrossRef]
- Costa, L.S.; Fidelis, G.P.; Cordeiro, S.L.; Oliveira, R.M.; Sabry, D.A.; Camara, R.B.; Nobre, L.T.D.B.; Costa, M.S.S.P.; Almeida-Lima, J.; Farias, E.H.C.; et al. Biological Activities of Sulfated Polysaccharides from Tropical Seaweeds. Biomed. Pharmacother. 2010, 64, 21–28. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Sanjeewa, K.A.; Samarakoon, K.W.; Lee, W.W.; Kim, H.-S.; Kim, E.-A.; Gunasekara, U.K.D.S.S.; Abeytunga, D.T.U.; Nanayakkara, C.; de Silva, E.D. FTIR Characterization and Antioxidant Activity of Water Soluble Crude Polysaccharides of Sri Lankan Marine Algae. Algae 2017, 32, 75–86. [Google Scholar] [CrossRef] [Green Version]
- Bilan, M.I.; Grachev, A.A.; Ustuzhanina, N.E.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. Structure of a Fucoidan from the Brown Seaweed Fucus evanescens C.Ag. Carbohydr. Res. 2002, 337, 719–730. [Google Scholar] [CrossRef]
- Bilan, M.I.; Grachev, A.A.; Ustuzhanina, N.E.; Shashkov, A.S.; Nifantiev, N.E.; Usov, A.I. A Highly Regular Fraction of a Fucoidan from the Brown Seaweed Fucus distichus L. Carbohydr. Res. 2004, 339, 511–517. [Google Scholar] [CrossRef]
- Ale, M.T.; Mikkelsen, J.; Meyer, A.S. Designed Optimization of a Single-Step Extraction of Fucose-Containing Sulfated Polysaccharides from Sargassum sp. J. Appl. Phycol. 2012, 24, 715–723. [Google Scholar] [CrossRef]
- Mak, W.; Hamid, N.; Liu, T.; Lu, J.; White, W.L. Fucoidan from New Zealand Undaria pinnatifida: Monthly Variations and Determination of Antioxidant Activities. Carbohydr. Polym. 2013, 95, 606–614. [Google Scholar] [CrossRef]
- Lim, S.J.; Aida, W.M.W.; Maskat, M.Y.; Mamot, S.; Ropien, J.; Mohd, D.M.; Wan Aida, W.M.; Maskat, M.Y.; Mamot, S.; Ropien, J.; et al. Isolation and Antioxidant Capacity of Fucoidan from Selected Malaysian Seaweeds. Food Hydrocoll. 2014, 42, 280–288. [Google Scholar] [CrossRef]
- Quitain, A.T.; Kai, T.; Sasaki, M.; Goto, M. Microwave–Hydrothermal Extraction and Degradation of Fucoidan from Supercritical Carbon Dioxide Deoiled Undaria pinnatifida. Ind. Eng. Chem. Res. 2013, 52, 7940–7946. [Google Scholar] [CrossRef]
- Saravana, P.S.; Cho, Y.-J.; Park, Y.-B.; Woo, H.-C.; Chun, B.-S. Structural, Antioxidant, and Emulsifying Activities of Fucoidan from Saccharina japonica Using Pressurized Liquid Extraction. Carbohydr. Polym. 2016, 153, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Jayawardena, T.U.; Fernando, I.P.S.; Lee, W.W.; Sanjeewa, K.K.A.; Kim, H.-S.; Lee, D.-S.; Jeon, Y.-J. Isolation and Purification of Fucoidan Fraction in Turbinaria ornata from the Maldives; Inflammation Inhibitory Potential under LPS Stimulated Conditions in in-vitro and in-vivo Models. Int. J. Biol. Macromol. 2019, 131, 614–623. [Google Scholar] [CrossRef]
- Jayawardena, T.U.; Sanjeewa, K.K.A.; Nagahawatta, D.P.; Lee, H.-G.; Lu, Y.-A.; Vaas, A.P.J.P.; Abeytunga, D.T.U.; Nanayakkara, C.M.; Lee, D.-S.; Jeon, Y.-J. Anti-Inflammatory Effects of Sulfated Polysaccharide from Sargassum swartzii in Macrophages via Blocking TLR/NF-Κb Signal Transduction. Mar. Drugs 2020, 18, 601. [Google Scholar] [CrossRef]
- Obluchinskaya, E.D.; Pozharitskaya, O.N.; Flisyuk, E.V.; Shikov, A.N. Formulation, Optimization and in vivo Evaluation of Fucoidan-Based Cream with Anti-Inflammatory Properties. Mar. Drugs 2021, 19, 643. [Google Scholar] [CrossRef]
- Garcia-Vaquero, M.; Rajauria, G.; O’Doherty, J.; Sweeney, T. Polysaccharides from Macroalgae: Recent Advances, Innovative Technologies and Challenges in Extraction and Purification. Food Res. Int. 2017, 99, 1011–1020. [Google Scholar] [CrossRef] [Green Version]
- Fujimura, T.; Shibuya, Y.; Moriwaki, S.; Tsukahara, K.; Kitahara, T.; Sano, T.; Nishizawa, Y.; Takema, Y. Fucoidan Is the Active Component of Fucus vesiculosus That Promotes Contraction of Fibroblast-Populated Collagen Gels. Biol. Pharm. Bull. 2000, 23, 1180–1184. [Google Scholar] [CrossRef] [Green Version]
- Baba, B.M.; Mustapha, W.A.W.; Joe, L.S.; Ibrahim, K.; Badri, K.H.; Jumali, M.H.H.; Noorani, M.S.M.; Ibrahim, N.; Rasol, N.H.A.; Yaacob, W.Z.W. Effects of Extraction Solvent on Fucose Content in Fucoidan Extracted from Brown Seaweed (Sargassum sp.) from Pulau Langkawi, Kedah, Malaysia. AIP Conf. Proc. 2016, 1784, 30045. [Google Scholar] [CrossRef]
- Dias, P.F.; Siqueira, J.M.; Maraschin, M.; Ferreira, A.G.; Gagliardi, A.R.; Ribeiro-do-Valle, R.M. A Polysaccharide Isolated from the Brown Seaweed Sargassum stenophyllum Exerts Antivasculogenic Effects Evidenced by Modified Morphogenesis. Microvasc. Res. 2008, 75, 34–44. [Google Scholar] [CrossRef]
- Rabanal, M.; Ponce, N.M.A.; Navarro, D.A.; Gómez, R.M.; Stortz, C.A. The System of Fucoidans from the Brown Seaweed Dictyota dichotoma: Chemical Analysis and Antiviral Activity. Carbohydr. Polym. 2014, 101, 804–811. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, M.; Shibata, H.; Kimura-Takagi, I.; Hashimoto, S.; Kimura, K.; Makino, T.; Aiyama, R.; Ueyama, S.; Yokokura, T. Structural Study of Fucoidan from Cladosiphon okamuranus TOKIDA. Glycoconj. J. 1999, 16, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Pielesz, A.; Biniaś, W.; Paluch, J. Mild Acid Hydrolysis of Fucoidan: Characterization by Electrophoresis and FT-Raman Spectroscopy. Carbohydr. Res. 2011, 346, 1937–1944. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.-S.S.; Athukorala, Y.; Jeon, Y.-J.J.; Senevirathne, M.; Rha, C.K.; Kim, S.-H.H. Antioxidant Activity of Sulfated Polysaccharides Isolated from Sargassum fulvellum. Prev. Nutr. Food Sci. 2007, 12, 65–73. [Google Scholar] [CrossRef]
- Song, K.-M.; Ha, S.J.; Lee, J.-E.; Kim, S.-H.; Kim, Y.H.; Kim, Y.; Hong, S.P.; Jung, S.K.; Lee, N.H. High Yield Ultrasonication Extraction Method for Undaria pinnatifida Sporophyll and Its Anti-Inflammatory Properties Associated with AP-1 Pathway Suppression. LWT-Food Sci. Technol. 2015, 64, 1315–1322. [Google Scholar] [CrossRef]
- Lorbeer, A.J.; Lahnstein, J.; Fincher, G.B.; Su, P.; Zhang, W. Kinetics of Conventional and Microwave-Assisted Fucoidan Extractions from the Brown Alga, Ecklonia radiata. J. Appl. Phycol. 2015, 27, 2079–2087. [Google Scholar] [CrossRef]
- Menshova, R.v.; Anastyuk, S.D.; Ermakova, S.P.; Shevchenko, N.M.; Isakov, V.I.; Zvyagintseva, T.N. Structure and Anticancer Activity in Vitro of Sulfated Galactofucan from Brown Alga Alaria angusta. Carbohydr. Polym. 2015, 132, 118–125. [Google Scholar] [CrossRef]
- Yang, C.; Chung, D.; You, S. Determination of Physicochemical Properties of Sulphated Fucans from Sporophyll of Undaria pinnatifida Using Light Scattering Technique. Food Chem. 2008, 111, 503–507. [Google Scholar] [CrossRef]
- Wang, J.; Liu, L.; Zhang, Q.; Zhang, Z.; Qi, H.; Li, P. Synthesized Oversulphated, Acetylated and Benzoylated Derivatives of Fucoidan Extracted from Laminaria japonica and Their Potential Antioxidant Activity in vitro. Food Chem. 2009, 114, 1285–1290. [Google Scholar] [CrossRef]
- Bendicho, C.; Lavilla, I. Ultrasound Extractions. In Reference Module in Chemistry, Molecular Sciences and Chemical Engineering; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Vinatoru, M. An Overview of the Ultrasonically Assisted Extraction of Bioactive Principles from Herbs. Ultrason. Sonochem. 2001, 8, 303–313. [Google Scholar] [CrossRef]
- Vardanega, R.; Santos, D.T.; de Almeida, M.A. Intensification of Bioactive Compounds Extraction from Medicinal Plants Using Ultrasonic Irradiation. Pharmacogn. Rev. 2014, 8, 88–95. [Google Scholar] [PubMed] [Green Version]
- Lavilla, I.; Bendicho, C. Chapter 11—Fundamentals of Ultrasound-Assisted Extraction. In Water Extraction of Bioactive Compounds: From Plants to Drug Development; Dominguez González, H., González Muñoz, M.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 291–316. [Google Scholar]
- Kadam, S.U.; Tiwari, B.K.; Smyth, T.J.; O’Donnell, C.P. Optimization of Ultrasound Assisted Extraction of Bioactive Components from Brown Seaweed Ascophyllum nodosum Using Response Surface Methodology. Ultrason. Sonochem. 2015, 23, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Sugiono; Widjanarko, S.; Soehono, L. Extraction Optimization by Response Surface Methodology and Characterization of Fucoidan from Brown Seaweed Sargassum polycystum. Int. J. Chemtech. Res. 2014, 6, 195–205. [Google Scholar]
- Flórez-Fernández, N.; López-García, M.; González-Muñoz, M.J.; Vilariño, J.M.L.; Domínguez, H. Ultrasound-Assisted Extraction of Fucoidan from Sargassum muticum. J. Appl. Phycol. 2017, 29, 1553–1561. [Google Scholar] [CrossRef]
- Sánchez-Ávila, N.; Priego-Capote, F.; Ruiz-Jiménez, J.; Luque de Castro, M.D. Fast and Selective Determination of Triterpenic Compounds in Olive Leaves by Liquid Chromatography-Tandem Mass Spectrometry with Multiple Reaction Monitoring after Microwave-Assisted Extraction. Talanta 2009, 78, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Macquarrie, D. Microwave Assisted Extraction of Sulfated Polysaccharides (Fucoidan) from Ascophyllum nodosum and Its Antioxidant Activity. Carbohydr. Polym. 2015, 129, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Jasso, R.M.; Mussatto, S.I.; Pastrana, L.; Aguilar, C.N.; Teixeira, J.A. Microwave-Assisted Extraction of Sulfated Polysaccharides (Fucoidan) from Brown Seaweed. Carbohydr. Polym. 2011, 86, 1137–1144. [Google Scholar] [CrossRef] [Green Version]
- Navarro, D.A.; Flores, M.L.; Stortz, C.A. Microwave-Assisted Desulfation of Sulfated Polysaccharides. Carbohydr. Polym. 2007, 69, 742–747. [Google Scholar] [CrossRef]
- Li, B.; Liu, S.; Xing, R.; Li, K.; Li, R.; Qin, Y.; Wang, X.; Wei, Z.; Li, P. Degradation of Sulfated Polysaccharides from Enteromorpha prolifera and Their Antioxidant Activities. Carbohydr. Polym. 2013, 92, 1991–1996. [Google Scholar] [CrossRef]
- Mandal, V.; Mohan, Y.; Hemalatha, S. Microwave Assisted Extraction—An Innovative and Promising Extraction Tool for Medicinal Plant Research. Pharmacogn. Rev. 2007, 1, 7–18. [Google Scholar]
- Flórez, N.; Conde, E.; Domínguez, H. Microwave Assisted Water Extraction of Plant Compounds. J. Chem. Technol. Biotechnol. 2015, 90, 590–607. [Google Scholar] [CrossRef]
- Chémat, F.; Cravotto, G. Microwave-Assisted Extraction for Bioactive Compounds: Theory and Practice; Springer: New York, NY, USA, 2013. [Google Scholar]
- Wijesinghe, W.A.J.P.; Jeon, Y.-J. Biological Activities and Potential Industrial Applications of Fucose Rich Sulfated Polysaccharides and Fucoidans Isolated from Brown Seaweeds: A Review. Carbohydr. Polym. 2012, 88, 13–20. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Dias, M.K.H.M.; Madusanka, D.M.D.; Han, E.J.; Kim, M.J.; Heo, S.-J.; Lee, K.; Cheong, S.H.; Ahn, G. Low Molecular Weight Fucoidan Fraction Ameliorates Inflammation and Deterioration of Skin Barrier in Fine-Dust Stimulated Keratinocytes. Int. J. Biol. Macromol. 2021, 168, 620–630. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, V.P.; Queiroz, K.C.S.; Cardoso, M.L.; Monteiro, G.R.G.; Oliveira, F.W.; Chavante, S.F.; Guimaraes, L.A.; Rocha, H.A.O.; Leite, E.L. Sulfated Galactofucan from Lobophora variegata: Anticoagulant and Anti-Inflammatory Properties. Biochemistry 2008, 73, 1018–1024. [Google Scholar] [CrossRef]
- Charoensiddhi, S.; Lorbeer, A.J.; Lahnstein, J.; Bulone, V.; Franco, C.M.M.; Zhang, W. Enzyme-Assisted Extraction of Carbohydrates from the Brown Alga Ecklonia radiata: Effect of Enzyme Type, PH and Buffer on Sugar Yield and Molecular Weight Profiles. Process Biochem. 2016, 51, 1503–1510. [Google Scholar] [CrossRef]
- Rodrigues, D.; Sousa, S.; Silva, A.; Amorim, M.; Pereira, L.; Rocha-Santos, T.A.P.; Gomes, A.M.P.; Duarte, A.C.; Freitas, A.C. Impact of Enzyme- and Ultrasound-Assisted Extraction Methods on Biological Properties of Red, Brown, and Green Seaweeds from the Central West Coast of Portugal. J. Agric. Food Chem. 2015, 63, 3177–3188. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Duan, L.; Li, X.; Yang, W.; Huang, T.; Kong, M.; Guan, F.; Ma, S. Fucoidan Ameliorates LPS-Induced Neuronal Cell Damage and Cognitive Impairment in Mice. Int. J. Biol. Macromol. 2022, 222, 759–771. [Google Scholar] [CrossRef]
- Hwang, P.A.; Phan, N.N.; Lu, W.J.; Hieu, B.T.N.; Lin, Y.C. Low-Molecular-Weight Fucoidan and High-Stability Fucoxanthin from Brown Seaweed Exert Prebiotics and Anti-Inflammatory Activities in Caco-2 Cells. Food Nutr. Res. 2016, 60, 32033. [Google Scholar] [CrossRef] [Green Version]
- Wijesinghe, W.A.J.P.; Jeon, Y.-J. Enzyme-Assistant Extraction (EAE) of Bioactive Components: A Useful Approach for Recovery of Industrially Important Metabolites from Seaweeds: A Review. Fitoterapia 2012, 83, 6–12. [Google Scholar] [CrossRef]
- Kim, K.-N.; Heo, S.-J.; Song, C.B.; Lee, J.; Heo, M.-S.; Yeo, I.-K.; Kang, K.A.; Hyun, J.W.; Jeon, Y.-J. Protective Effect of Ecklonia cava Enzymatic Extracts on Hydrogen Peroxide-Induced Cell Damage. Process Biochem. 2006, 41, 2393–2401. [Google Scholar] [CrossRef]
- Athukorala, Y.; Kim, K.N.; Jeon, Y.J. Antiproliferative and Antioxidant Properties of an Enzymatic Hydrolysate from Brown Alga, Ecklonia cava. Food Chem. Toxicol. 2006, 44, 1065–1074. [Google Scholar] [CrossRef] [PubMed]
- Olivares-Molina, A.; Fernández, K. Comparison of Different Extraction Techniques for Obtaining Extracts from Brown Seaweeds and Their Potential Effects as Angiotensin I-Converting Enzyme (ACE) Inhibitors. J. Appl. Phycol. 2016, 28, 1295–1302. [Google Scholar] [CrossRef]
- Kim, Y.-I.; Oh, W.-S.; Song, P.H.; Yun, S.; Kwon, Y.-S.; Lee, Y.J.; Ku, S.-K.; Song, C.-H.; Oh, T.-H. Anti-Photoaging Effects of Low Molecular-Weight Fucoidan on Ultraviolet B-Irradiated Mice. Mar. Drugs 2018, 16, 286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogalinski, T.; Liu, K.; Albrecht, T.; Brunner, G. Hydrolysis Kinetics of Biopolymers in Subcritical Water. J. Supercrit. Fluids 2008, 46, 335–341. [Google Scholar] [CrossRef]
- Balboa, M.E.; Rivas, S.; Moure, A.; Domínguez, H.; Parajó, J.C. Simultaneous Extraction and Depolymerization of Fucoidan from Sargassum muticum in Aqueous Media. Mar. Drugs 2013, 11, 4612–4627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Meizoso, I.; Marin, F.R.; Herrero, M.; Señorans, F.J.; Reglero, G.; Cifuentes, A.; Ibáñez, E. Subcritical Water Extraction of Nutraceuticals with Antioxidant Activity from Oregano. Chemical and Functional Characterization. J. Pharm. Biomed. Anal. 2006, 41, 1560–1565. [Google Scholar] [CrossRef] [Green Version]
- Queffelec, J.; Flórez-Fernández, N.; Dominguez, H.; Torres, M.D. Microwave Hydrothermal Processing of Undaria pinnatifida for Bioactive Peptides. Bioresour. Technol. 2021, 342, 125882. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Q.; Zhang, Z.; Li, Z. Antioxidant Activity of Sulfated Polysaccharide Fractions Extracted from Laminaria japonica. Int. J. Biol. Macromol. 2008, 42, 127–132. [Google Scholar] [CrossRef]
- Cernadas, H.; Flórez-Fernández, N.; González-Muñoz, M.J.; Domínguez, H.; Torres, M.D. Retrieving of High-Value Biomolecules from Edible Himanthalia elongata Brown Seaweed Using Hydrothermal Processing. Food Bioprod. Process. 2019, 7, 275–286. [Google Scholar] [CrossRef]
- Gan, A.; Baroutian, S. Subcritical Water Extraction for Recovery of Phenolics and Fucoidan from New Zealand Wakame (Undaria pinnatifida) Seaweed. J. Supercrit. Fluids. 2022, 190, 105732. [Google Scholar] [CrossRef]
- Morimoto, M.; Takatori, M.; Hayashi, T.; Mori, D.; Takashima, O.; Yoshida, S.; Sato, K.; Kawamoto, H.; Tamura, J.; Izawa, H. Depolymerization of Sulfated Polysaccharides under Hydrothermal Conditions. Carbohydr. Res. 2014, 384, 56–60. [Google Scholar] [CrossRef]
- Shanura Fernando, I.P.; Asanka Sanjeewa, K.K.; Samarakoon, K.W.; Kim, H.-S.; Gunasekara, U.K.D.S.S.; Park, Y.-J.; Abeytunga, D.T.U.; Lee, W.W.; Jeon, Y.-J. The Potential of Fucoidans from Chnoospora minima and Sargassum polycystum in Cosmetics: Antioxidant, Anti-Inflammatory, Skin-Whitening, and Antiwrinkle Activities. J. Appl. Phycol. 2018, 30, 3223–3232. [Google Scholar] [CrossRef]
- Lee, S.-H.; Ko, C.-I.; Jee, Y.; Jeong, Y.; Kim, M.; Kim, J.-S.; Jeon, Y.-J. Anti-Inflammatory Effect of Fucoidan Extracted from Ecklonia cava in Zebrafish Model. Carbohydr. Polym. 2013, 92, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Oh, J.-Y.; Lee, W.; Jeon, Y.-J. Fucoidan Isolated from Hizikia fusiforme Suppresses Ultraviolet B-Induced Photodamage by down-Regulating the Expressions of Matrix Metalloproteinases and pro-Inflammatory Cytokines via Inhibiting NF-ΚB, AP-1, and MAPK Signaling Pathways. Int. J. Biol. Macromol. 2021, 166, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Ren, D.; Song, Y.; Wu, L.; He, Y.; Peng, Y.; Zhou, H.; Liu, S.; Cong, H.; Zhang, Z.; et al. Gastric Protective Activities of Fucoidan from Brown Alga Kjellmaniella crassifolia through the NF-ΚB Signaling Pathway. Int. J. Biol. Macromol. 2020, 149, 893–900. [Google Scholar] [CrossRef]
- Asanka Sanjeewa, K.K.; Jayawardena, T.U.; Kim, H.-S.; Kim, S.-Y.; Shanura Fernando, I.P.; Wang, L.; Abetunga, D.T.U.; Kim, W.-S.; Lee, D.-S.; Jeon, Y.-J. Fucoidan Isolated from Padina commersonii Inhibit LPS-Induced Inflammation in Macrophages Blocking TLR/NF-ΚB Signal Pathway. Carbohydr. Polym. 2019, 224, 115195. [Google Scholar] [CrossRef]
- Ni, L.; Wang, L.; Fu, X.; Duan, D.; Jeon, Y.-J.; Xu, J.; Gao, X. In vitro and in vivo Anti-Inflammatory Activities of a Fucose-Rich Fucoidan Isolated from Saccharina japonica. Int. J. Biol. Macromol. 2020, 156, 717–729. [Google Scholar] [CrossRef]
- Hwang, P.-A.; Hung, Y.-L.; Chien, S.-Y. Inhibitory Activity of Sargassum hemiphyllum Sulfated Polysaccharide in Arachidonic Acid-Induced Animal Models of Inflammation. J. Food Drug. Anal. 2015, 23, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Luo, D.; Wang, Z.; Nie, K. Structural Characterization of a Novel Polysaccharide from Sargassum thunbergii and Its Antioxidant and Anti-Inflammation Effects. PLoS ONE 2019, 14, e0223198. [Google Scholar] [CrossRef]
- Hai-lan, C.; Hong-lian, T.; Jian, Y.; Manling, S.; Heyu, F.; Na, K.; Wenyue, H.; Si-yu, C.; Ying-yi, W.; Ting-jun, H. Inhibitory Effect of Polysaccharide of Sargassum weizhouense on PCV2 Induced Inflammation in Mice by Suppressing Histone Acetylation. Biomed. Pharmacother. 2019, 112, 108741. [Google Scholar] [CrossRef]
- Manikandan, R.; Parimalanandhini, D.; Mahalakshmi, K.; Beulaja, M.; Arumugam, M.; Janarthanan, S.; Palanisamy, S.; You, S.; Prabhu, N.M. Studies on Isolation, Characterization of Fucoidan from Brown Algae Turbinaria decurrens and Evaluation of It’s in vivo and in vitro Anti-Inflammatory Activities. Int. J. Biol. Macromol. 2020, 160, 1263–1276. [Google Scholar] [CrossRef] [PubMed]
- Wardani, G.; Nugraha, J.; Mustafa, M.R.; Sudjarwo, S.A. Antioxidative Stress and Anti-Inflammatory Activity of Fucoidan Nanoparticles against Nephropathy of Streptozotocin-Induced Diabetes in Rats. Evid.-Based Complement. Altern. Med. 2022, 2022, 3405871. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; Jeong, S.; Lee, Y.; Jeon, C.; Kwon, G.; Kim, S.; Lee, D. H2O2-Activatable Antioxidant Polymeric Prodrug Nanoparticles for the Prevention of Renal Ischemia/Reperfusion Injury. Biomacromolecules 2022, 23, 3810–3821. [Google Scholar] [CrossRef] [PubMed]
- Sumayya, A.S.; Muraleedhara Kurup, G. In Vitro Anti-Inflammatory Potential of Marine Macromolecules Cross-Linked Bio-Composite Scaffold on LPS Stimulated RAW 264.7 Macrophage Cells for Cartilage Tissue Engineering Applications. J. Biomater. Sci. Polym. Ed. 2021, 32, 1040–1056. [Google Scholar] [CrossRef]
- Liu, Y.; Qi, X.; Wang, Y.; Li, M.; Yuan, Q.; Zhao, Z. Inflammation-Targeted Cannabidiol-Loaded Nanomicelles for Enhanced Oral Mucositis Treatment. Drug Deliv. 2022, 29, 1272–1281. [Google Scholar] [CrossRef]
- Hao, Y.; Zheng, W.; Sun, Z.; Zhang, D.; Sui, K.; Shen, P.; Li, P.; Zhou, Q. Marine Polysaccharide-Based Composite Hydrogels Containing Fucoidan: Preparation, Physicochemical Characterization, and Biocompatible Evaluation. Int. J. Biol. Macromol. 2021, 183, 1978–1986. [Google Scholar] [CrossRef]
- Yu, Q.; Han, F.; Yuan, Z.; Zhu, Z.; Liu, C.; Tu, Z.; Guo, Q.; Zhao, R.; Zhang, W.; Wang, H.; et al. Fucoidan-Loaded Nanofibrous Scaffolds Promote Annulus Fibrosus Repair by Ameliorating the Inflammatory and Oxidative Microenvironments in Degenerative Intervertebral Discs. Acta Biomater. 2022, 148, 73–89. [Google Scholar] [CrossRef]
- Chen, B.-R.; Hsu, K.-T.; Hsu, W.-H.; Lee, B.-H.; Li, T.-L.; Chan, Y.-L.; Wu, C.-J. Immunomodulation and Mechanisms of Fucoidan from Cladosiphon okamuranus Ameliorates Atopic Dermatitis Symptoms. Int. J. Biol. Macromol. 2021, 189, 537–543. [Google Scholar] [CrossRef]



| Extraction Technology Advantages Disadvantages | Challenges | Effects on Fucoidan Features and Anti-Inflammatory Properties |
|---|---|---|
| Conventional solvent extraction | ||
| Simple and established | Prolonged time High temperatures High energy consumption Chemicals Degradation of products | Excessive degradation under severe conditions Desulfation and depolymerization |
| Subcritical water extraction | ||
| Short processing time High extraction yields Selectivity by modulating operational conditions | High capital cost Degradation of products Undesirable byproducts | Simultaneous extraction and depolymerization Maintains structure and sulfation Further processing with other technologies |
| Enzyme assisted extraction | ||
| Mild conditions Low energy requirements Selective Enhanced extraction yields | Slow process Availability and cost of enzymes Enzyme recycling and reuse strategies | Depolymerization Maintains sulfation degree |
| Ultrasound assisted extraction | ||
| Simplicity Mild temperatures Short time Decreased use of solvent Scalable Reduced equipment size Low energy, costs, and risks | Localized heating and non-uniform conditions Radical formation and possible degradation Noise pollution | Reduction in molecular weight |
| Microwave assisted extraction | ||
| Rapid heating Short extraction times Increased yield | High energy needs Thermal degradation of some components | Degradation of structure sulfate |
| Seaweed/Processing | Model System and Anti-Inflammatory Actions | Reference |
|---|---|---|
| Chooospora minima Pre-Ext: 95% E; 10% FMD in 95% E, 8 h, 37 °C, E wash Ext: Celluclast (0.5% E/S), pH 4.5, 50 °C, 24 h Alcalase (0.5% E/S), pH 8.0, 50 °C, 24 h Neutr., 5 M CaCl2 alginate pptn; conc., 95% E pptn Purif: AEC 63.5 % carboh.; 34.1% sulf.; 0.2% prot.; 0.4% polyph. | LPS-stimulated RAW cells Reduced activity of iNOS and NO levels. Downregulation of PGE2, reduction in COX-2 levels. Downregulation of pro-inflammatory cytokines TNF-α, IL1β, and IL6 | [187] |
| Ecklonia maxima Pre-Ext: 95% E, 10% FMD in 95% E, 8 h, 37 °C, E washing Ext: Celluclast (0.5% E/S), pH 4.5, 50 °C, 24 h Alcalase (0.5% E/S), pH 8.0, 50 °C, 24 h Neutraliz., 5 M CaCl2 alginate pptn; conc.; 95% E pptn Purif: AEC 51.4% carbohyd.; 39.8 % sulf., 0.5% prot.; 0.3 % polyph. 81.8% Fuc, 0.33 % Rha, 0.04% Ara, 14.7%, Gal, 3.1% Man | Particulate matter-stimulated RAW 264.7 cells Suppressed production of NO, prostaglandin E2, and pro-inflammatory cytokines (TNF-α, IL-6, and IL-1β) by inhibiting the NF-κB and MAPK signaling pathways | [112] |
| Ecklonia cava Ext: Celluclast, 1%, 50 °C, 24 h E pptn; C; 4 M CaCl2 alginate pptn; cetylpyridinium chloride pptn, resolubilized with 3 M CaCl2; E pptn; Dl, FD 51.8% carbohyd.; 20.1 % sulf.; 11.3% uronic ac.; 8.9% protein 61.1% Fuc, 3.9% Rha; 27.2 Gal; 0.8 % Glu; 7.0% Xyl | Tail-cutting-induced and LPS-induced zebrafish Inhibition of ROS and NO generation | [188] |
| Ecklonia cava Ext: Glucoamylase, optimal pH and T, 1% (v/w), 24 h UF (50 kDa), E pptn Purif: AEC and GPC Fuc:Rha:Gal:Glu:Man:Xyl ratio 82.1:0.3:12.2:0.22.2:2.2 Sulf: Total sugars 0.92:1.38, 103 kDa | LPS-stimulated RAW 264.7 cells Inhibited NO and PGE2 production, and suppressed iNOS and COX-2 expression | [17] |
| Fucus vesiculosus Pre-Ext: MC:E Ext: US, 5% E, 40 °C, pH 3-4, 4 h; centr; Dl; FD 79.5% carbohyd., 27.0% sulf., 0.7% uronic ac. Fuc:Gluc:Gal:Xyl:Man:Ara (mol) 1.0:0.16:0.05:0.09:0.03:0.03; 735 kDa | Topical application in rats Inhibited carrageenan-induced edema with comparable efficacy to diclofenac gel | [138] |
| Hizikia fusiforme Ext: Celluclast (5% enzyme), pH 4.5, 50 °C, 24 h; E pptn Pur: AEC; Dl, FD 71.8% carbohyd., 27.2% sulf.; 102.67 kDa 79.2% Fuc, 18.1% Man, 2.1 % Rha, 0.4% Arab, 0.2% Glu | UVB-induced photodamage in vitro in human dermal fibroblasts and in vivo in zebrafish Improved collagen synthesis, inhibited intracellular collagenase, and reduced expression of matrix metalloproteinases and pro-inflammatory cytokines | [189] |
| Kjellmaniella crassifolia Ext: cellulase and pectinase; Filtr, E pptn, washed, FD | Gastric protective effects. Suppressed aspirin-induced NF-κB activation via stabilization of IκB-α | [190] |
| Laminaria japonica Ext: 30 mM H2O2 and ascorbate 1:1, 2 h Dl (3.6 kDa); E pptn 28.7% fuc; 3.6% uronic ac., 30.1% sulf., 6.5 kDa | Non-alcoholic fatty-liver disease in obese diabetic rats Reduced expression of TNF-α, MCP-1, and NF-κB. Down-regulation of pro-inflammatory cytokines and transcription factors, and up-regulation of adiponectin | [110] |
| Lobophora variegata Pre-Ext: A, 60 °C, 18 h Ext: Maxatase, A (1 vol) fractionation, molec. sieving, IEC; MeOH pptn, D, Fuc:Gal:Sulf (molar) 1:3:2 | Ear swelling caused by croton oil Inhibited leukocyte migration to the inflammation site | [169] |
| Padina commersonii Frozen samples Ext: Celluclast Purif: DEAE-cellulose; Dl | LPS induction in RAW 264.7 macrophages Inhibited TLR2/4 and MyD88 transcriptional activities. Reduced the transcriptional activities of NF-κB signal transduction. Inhibited cytokine and NO secretion | [191] |
| Saccharina japonica Pre-Ext: 95% E, 2 h, 40 °C Ext: W (1:30, w/v), 120 °C, 2 h, C, 2% CaCl alginate pptn; C, conc.; Dl (3.5 kDa), FD Purif: AEC (DEAE) 56.5 % polysacch., 30.7% sulfate 79.5 % Fuc; 16.8 % Gal; 0.8 % Rha, 1.1% Xyl, 1.8% Man | LPS-induced RAW264.7 cells Decreased the production of NO, TNF-α, IL-1β and IL-6. Down-regulated expression of MAPK and NF-κB pathways. in vivo LPS-induced zebrafish Reduced cell death rate and production of NO and ROS | [192] |
| Saccharina japonica Pre-Ext: 95% E, 40 °C, 2 h Ext: W, 120 °C, 2 h C, 2% CaCl2 alginate pptn, conc., Dl, FD Purif: AEC, Fuc:Gal 79.2:20.8; 11.46% sulfate | RAW264.7 cells Reduced NO. Down-regulated MAPK (p38, ENK and JNK) and NF-κB (p65 and IKKα/IKKβ) pathways. Zebrafish Reduced cell death rate, inhibited NO, and decreased ROS | [33] |
| Sargassum cristaefolium Pre-Ext: (Compressional-puffing) 99% E, 25°C, 4 h Ext: SWE, 121°C, 20 min, Dl Pur: AEC, Dl, FD Depol: 1 M H2O2, 60 °C, 1 h, C, filtr., 3.2 kDa | LPS-stimulated RAW264.7 and HaCaT cells Inhibited COX2 and p38 UVB-induced damage in Hs68 cells and in rats After stimulation with LPS, TNF-α, and IFN-γ, improved redness and swelling caused by UVB irradiation | [26] |
| Sargassum cristaefolium Pre-Ext: E defatted with ethanol Ext: W, 100 °C, 1 h 95% E pptn; 75% E, 2 stages; FD | LPS-induced RAW264.7 cells Suppression of induced p38, ERK1/2, and JNK phosphorylation. Inhibited NO secretion. Down-regulated iNOS expression by inhibiting MAPK and NF-κB pathways | [27] |
| Sargassum fusiforme Pre-Ext: 80% E Ext: 0.01 M HCl 4 M CaCl2 alginate pptn., Dl, Conc, 95% E ppt, deproteinizat. Purif: AEC 64.1 % carbohyd., 28.2% uronic ac., 5.4% sulf. Man:Fuc:Rha:GlcA:Xyl:Gal:Glu (mol) 26.9: 21.5: 18.5: 9.9: 9.7: 7.7: 5.8 | Binding of P-selectin to HL-60 cells Disrupted P-selectin-mediated cell adhesion and rolling and blocked the interaction between P-selectin and its physiological ligand PSGL-1 | [20] |
| Sargassum fusiforme Pre-Ext: 90% E reflux Ext: W, 70 °C, 2 h, 2 stages, C, conc., UF, FD Depol: HCl (pH 2.0), neutr., E pptn; A and E washing; D 98.1% purity, 22% Fuc, 22.8% sulfate | LPS in normal human colonic epithelial cells in vivo experiments in mice Restored the diversity of gut microbial composition. Reduced incidence of tumors in mice. Decreased TNF-α, IL-6, and IL-1β expression levels | [31] |
| Sargassum fusiforme Ext: 0.4% citric ac., 60 °C, 120 min, homogeniz., neutr., steriliz., inoculated 5% Lactobacillus rhamnosus, 37 °C, 48 h, C, E pptn Purif: AEC 71.8% carbohydrate, 27.2% sulfate, 102.7 kDa 79.2% Fuc, 2.1% Rha, 0.2% Glu, 18.1 % Man, 0.4% Ara | LPS-stimulated RAW 264.7 cells Inhibited TNF-α, NO, PGE2, IL-1β, and IL-6 production. Improved viability. Suppressed the expression of COX-2 and iNOS by regulating the NF-κB pathway in vivo zebrafish Reduced ROS | [189] |
| Sargassum horneri Pre-Ext: 95% E; 10% FMD in E; 3 h; 95% E washing Celluclast, 50 °C, 8 h FD, Centr, E pptn Pur: AEC; Dl sulfated mannofucans 45 kDa | Fine-dust on skin inflammation in HaCaT keratinocytes Recovery of skin barrier dysfunction. Lowered ROS levels. Down-regulated TNF-α, IL-1β, -5, -6, -8, -13, interferon-γ, and chemokines. Inhibited mitogen-activated protein kinase and NF-κB pathways | [168] |
| Sargassum hemiphyllum Ext: W, 100 °C, 30 min C, FD | Lipopolysaccharide-activated RAW 264.7 cells Reduced secretion profiles of IL-1β, IL-6, TNF-α, and NO. Down-regulated NF-κB (p65) in nucleus | [21] |
| Sargassum hemiphyllum Ext: W, 100 °C, 30 min C, FD 95% E pptn., C, FD | Aarachidonic acid-induced ear inflammatory in mice Decreased ear swelling and erythema. Decreased production of myeloperoxidase, NO, IL-1β, IL-6, and TNF-α. Reduced area of neutrophilic infiltration in ears | [193] |
| Sargassum muticum Ext.: W, heating up to 170 °C Alginate pptn Fuc:Gal + Xyl + Man:Glu1:0.94:0.24 | Reduced IL-6 production stimulated by IL-1β. Up-regulated Nrf-2 levels and the expression of its transcriptional target genes HO-1 and SOD-2. No attenuation of chondrocyte senescence | [104] |
| Sargassum patens Ext: crude enzyme from Shewanella oneidensis PKA 1008, LRS: 50; Seaweed:crude enzyme ratio 1:1(v/v), 30°C, 48 h | LPS-induced RAW 264.7 cells Inhibited secretion of IL-6, IL-1β, and TNF-α cytokines | [28] |
| Sargassum polycystum Pre-Ext: 95% E; 10% FMD in 95% E, 8 h, 37 °C, E washing Ext: Celluclast (0.5% E/S), pH 4.5, 50 °C, 24 h. Alcalase (0.5% E/S), pH 8.0, 50 °C, 24 h Neutraliz., 5 M CaCl2 alginate pptn, concent., 95% E pptn 75% E washing; C 62.9% carbohyd.; 27.5% sulf; 0.1% prot; 3.4% polyp., 0.5% ash | RAW cells Reduce the NO levels related to the reduced production or activity of iNOS. Down-regulated PGE2, reduced COX-2, TNF-α, IL1β, and IL6 levels | [187] |
| Sargassum siliquosum Pre-Ext: (HTHPP), 95% E, RT, 4 h Ext: MAE: W, LSR 15; MAE, 750 W, 10 min, F; Dl; 95% E pptn; IE pptn; C, Dl; 2% CaCl2 alginate pptn Purif: AEC; Dl, F, FD Depol: 0.1 M H2O2, 60 °C, 60 min 64.5% carbohyd., 19.5% sulf., 6.1% uronic ac., 9.9% phen. + prot., 31.5 and 3.2 kDa | LPS-stimulated RAW264.7 macrophage cells Suppressed TNF-α production | [113] |
| Sargassum swartzii Pr-Ext: 95% E, 3 stages; 10% FMD in 95% E Ext: Celluclast (0.5%), pH 4.5, 50 °C, 24 h; Filtr, Alcalase, pH 8.0, 50 °C, 24 h; neutr.; CaCl2 alginate pptn; neutraliz.; conc. FD, Pur: IEC 61% carbohyd., 34% sulf.; 0.4 % prot.; 0.3% polyphenol Fuc:Gal: Glu:Others ratio 82.5:3.2:1.3:13.0 | LPS-stimulated RAW 264.7 macrophages Decreased NO production, acted on mediators such as iNOS, COX-2, and pro-inflammatory cytokines (TNF-α, IL-6, and IL-1β) Suppressed TLR-mediated MyD88, IKK complex, ultimately hindering NF-κB and MAPK activation | [137] |
| Sargasum thunbergii Ext: hot-water extraction, 0.5 M NaOH, 4 °C, 10 h; Amylase, pH 2.0, 37 °C, 2 h; conc.; E pptn, D; Deproteinization Purif: AEC; D, FD; 98.9% carbohyd. Fuc:Gal (mol) 1.2:1, 373 kDa | LPS-stimulated RAW 264.7 mouse macrophage cells Reduced TNF-α, IL-6, and COX-2 mRNA expression | [194] |
| Sargassum weizhouense Pre-Ext: 95% E, 80 °C, 8 h Ext: 1% papain, 50 °C, 1.5 h; filtr; conc., 95% E, 100% E | PCV2 infection Inhibited histone acetylation and the production of inflammatory cytokines, improving the resistance of the host | [195] |
| Turbinaria ornata Pre-Ext: 95% E; 90% E, 10% FMD, 8 h Celluclast 0.5%, pH 4.5, 50 °C, 24 h Alcalase 0.5% E/S, pH 8.0, <50 °C, 24 h CaCl2 alginate pptn; neutraliz; 95% E pptn AEC, 60.3% carboh., 38.3% sulf., 0.24% prot., 0.25% polyph | LPS-treated RAW 264.7 macrophages Inhibited NO production. Down-regulated expression of iNOS and COX-2 Zebra fish embryos Reduced the NO, ROS, and cell death levels; down-regulated inflammatory mediators, iNOS, and COX-2 | [136] |
| Undaria pinnatifida Comercial fucoidans hydrolyzed CuAc pH 7.5, 60 °C, 9% H2O2, 5 h Cu removal, neutralization, Dl (1 kDa), FD LMWF (5–30 kDa) | LPS RAW264.7 cells Regulated signaling pathways, attenuated IL-1β, IL-1, and TNF-α, and the degradation of phosphorylated p38 MAPK, ERK1/2 and JNK. Blocked NO and ROS. Inhibited iNOS and COX-2 | [24] |
| Turbinaria decurrens Pre-Ext.: 85% E, RT, 12 h W, 65 °C, 1 h C, 1% CaCl2 alginate pptn; 99% E sequential pptn Pur: dW heated with 3.0 M HCl, 3 h, cooled, C, neutr., E pptn, W washing, FD 54.8% polys., 23.5% sulf, 3.4 % uronic, 2.7% prot. 9.3% Fuc, 12.6% Gal, 9.6% Man, 6.4% Rha, 11.4% Xyl | Reduced LPS-induced cytotoxicity in IC-21 macrophages Formalin-induced paw edema in mouse model Decreased the MDA and increased SOD, CAT, GPx, GST, and GSH activity. Retained p65/NF-κB transcription factor. Down-regulated expression of pro-inflammatory mediators such as IL-1β, COX-2, and MMP-9 | [196] |
| Undaria pinnatifida Ext: Triton solubilized; hot, acidic water or hot alkaline water Depol: W, copper acetate monohydrate, 9% (v/v) H2O2, 60 °C, 5 h, Cu removal, neutr., diafiltration (1 kDa), FD AEC (DEAE), conc., Dl, LMWF, 1 kDa | LPS-stimulated spleen cells Suppressed the production of IFN-γ Collagen-induced arthritis mice model; spleen cells Inhibited Th1-mediated responses, reduced collagen-specific IgG2a levels in serum | [115] |
| Undaria pinnatifida Pre-Ext: 85% EtOH, 70 °C, 2 h Ext: HCl (pH 2), RT, 24 h. 582.5 kDa or US, 80% amplitude, 6 h, HCl (pH 2) neutr., Dl (3.5 kDa), FD, 390.6 kDa | LPS-induced inflammation in Raw 264.7 cells Suppressed iNOS and COX-2, and JNK1/2 and p38 phosphorylation | [147] |
| Undaria pinnatifida Ext: MAE, W, 160 °C Alginate pptn Fuc:Gal + Xyl + Man:Glu 1:0.93:0.24 | Reduced IL-6 production stimulated by IL-1β. Up-regulated Nrf-2 levels and the expression of its transcriptional target genes HO-1 and SOD-2. No attenuation of chondrocyte senescence | [104] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flórez-Fernández, N.; Vaamonde-García, C.; Torres, M.D.; Buján, M.; Muíños, A.; Muiños, A.; Lamas-Vázquez, M.J.; Meijide-Faílde, R.; Blanco, F.J.; Domínguez, H. Relevance of the Extraction Stage on the Anti-Inflammatory Action of Fucoidans. Pharmaceutics 2023, 15, 808. https://doi.org/10.3390/pharmaceutics15030808
Flórez-Fernández N, Vaamonde-García C, Torres MD, Buján M, Muíños A, Muiños A, Lamas-Vázquez MJ, Meijide-Faílde R, Blanco FJ, Domínguez H. Relevance of the Extraction Stage on the Anti-Inflammatory Action of Fucoidans. Pharmaceutics. 2023; 15(3):808. https://doi.org/10.3390/pharmaceutics15030808
Chicago/Turabian StyleFlórez-Fernández, Noelia, Carlos Vaamonde-García, Maria Dolores Torres, Manuela Buján, Alexandra Muíños, Antonio Muiños, María J. Lamas-Vázquez, Rosa Meijide-Faílde, Francisco J. Blanco, and Herminia Domínguez. 2023. "Relevance of the Extraction Stage on the Anti-Inflammatory Action of Fucoidans" Pharmaceutics 15, no. 3: 808. https://doi.org/10.3390/pharmaceutics15030808