
Reversal of Multidrug Resistance by Symmetrical Selenoesters in Colon Adenocarcinoma Cells
 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
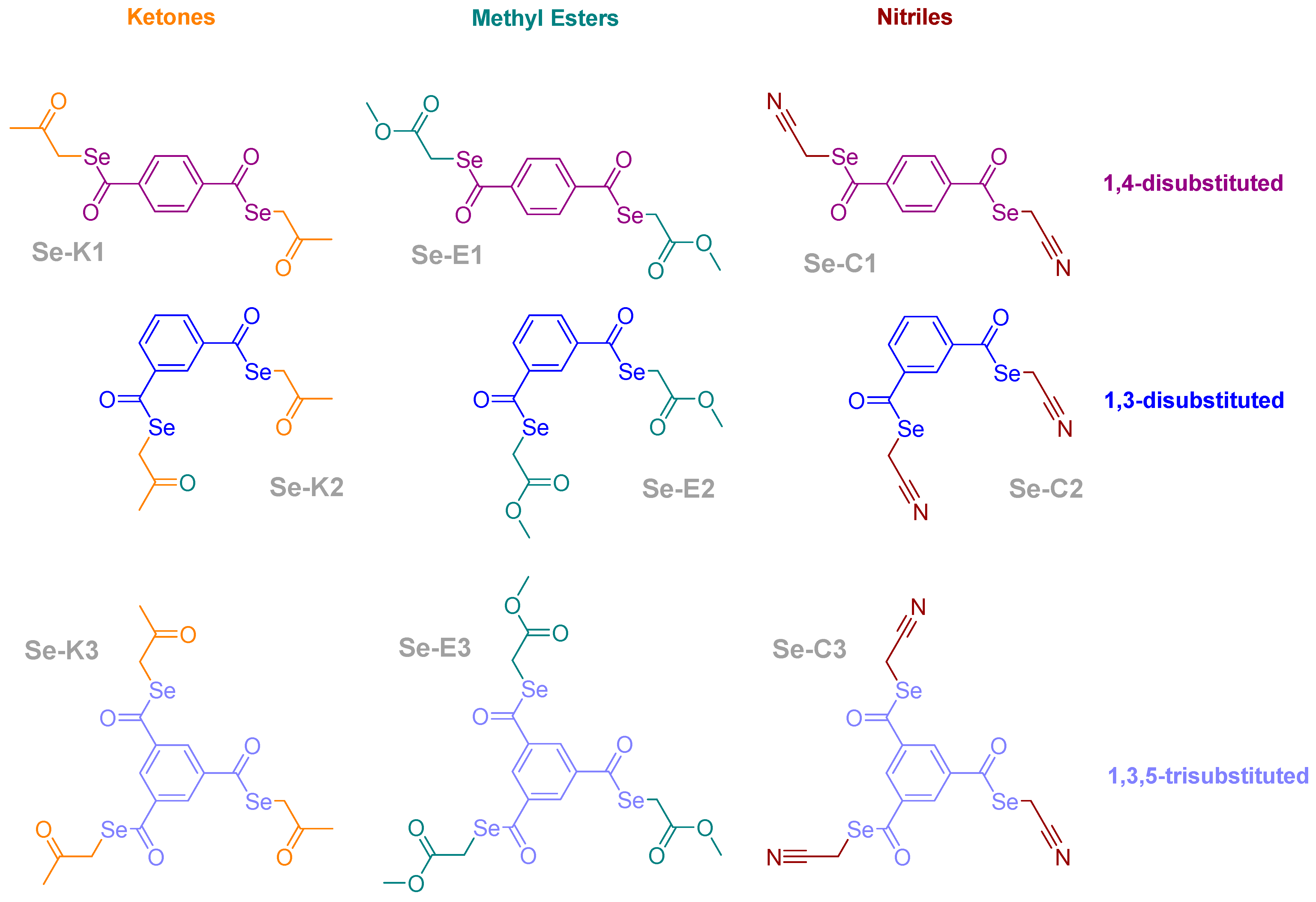
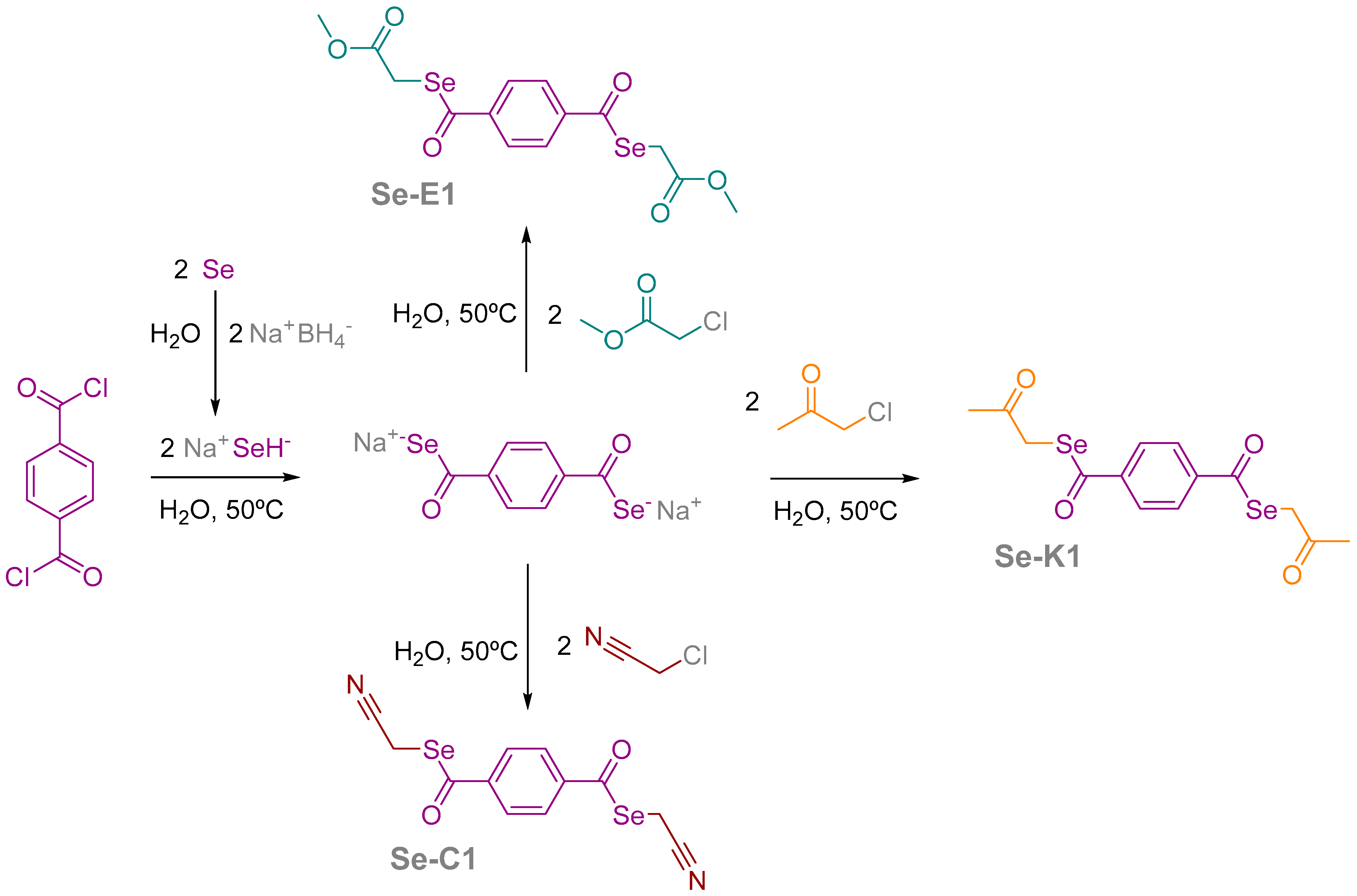
2.1. Compounds
2.2. Cell Cultures
2.3. Rhodamine 123 Accumulation Assay
2.4. Checkerboard Combination Assay
2.5. Apoptosis Assay
2.6. Preparation and Treatment of Cells for RNA Extraction
2.7. RNA Isolation and cDNA Library Preparation and Whole Transcriptome Sequencing
2.8. Analysis of Transcriptomic Data
3. Results
3.1. Combined Effect with Doxorubicin
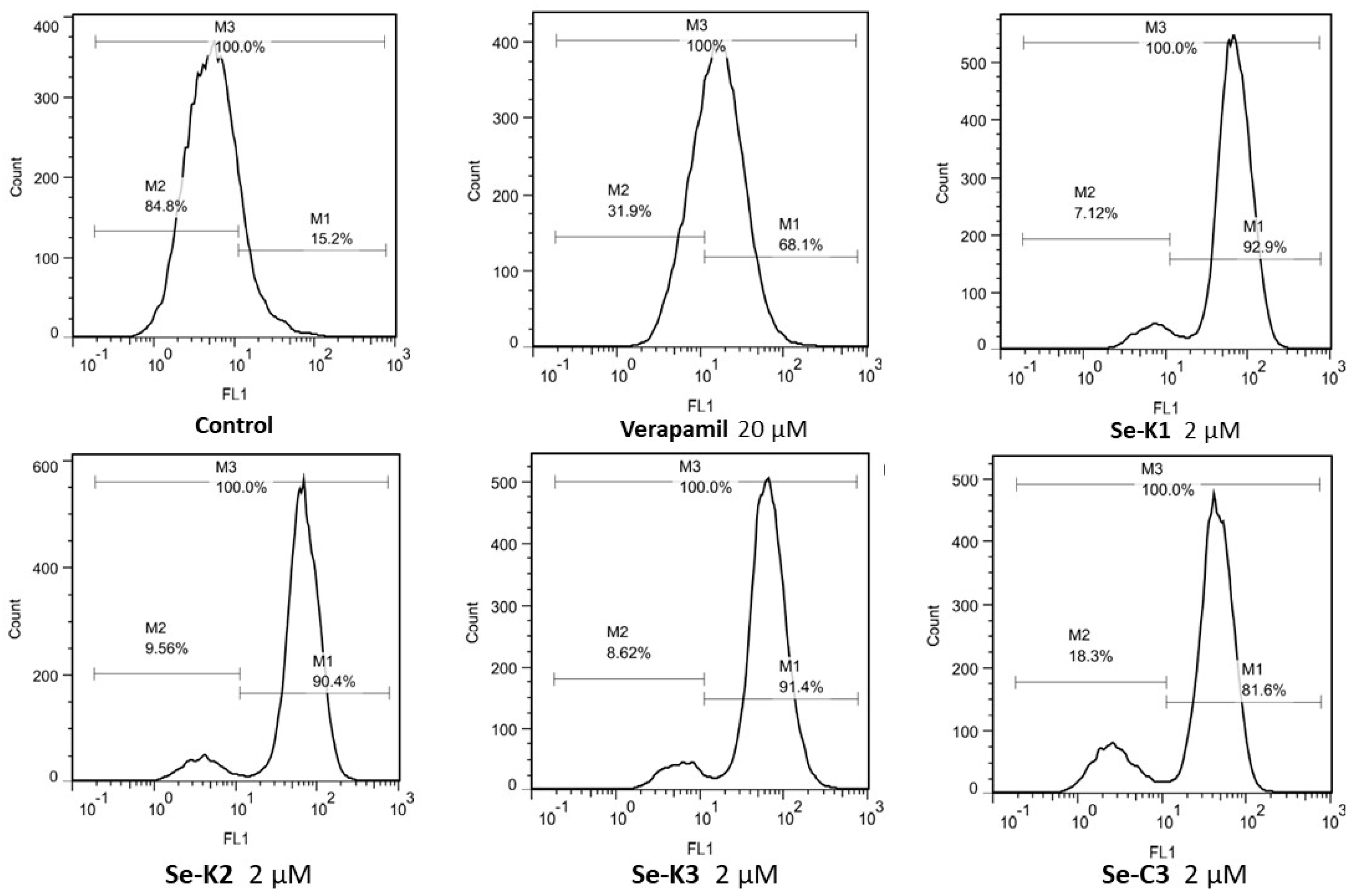
3.2. ABCB1 Inhibition
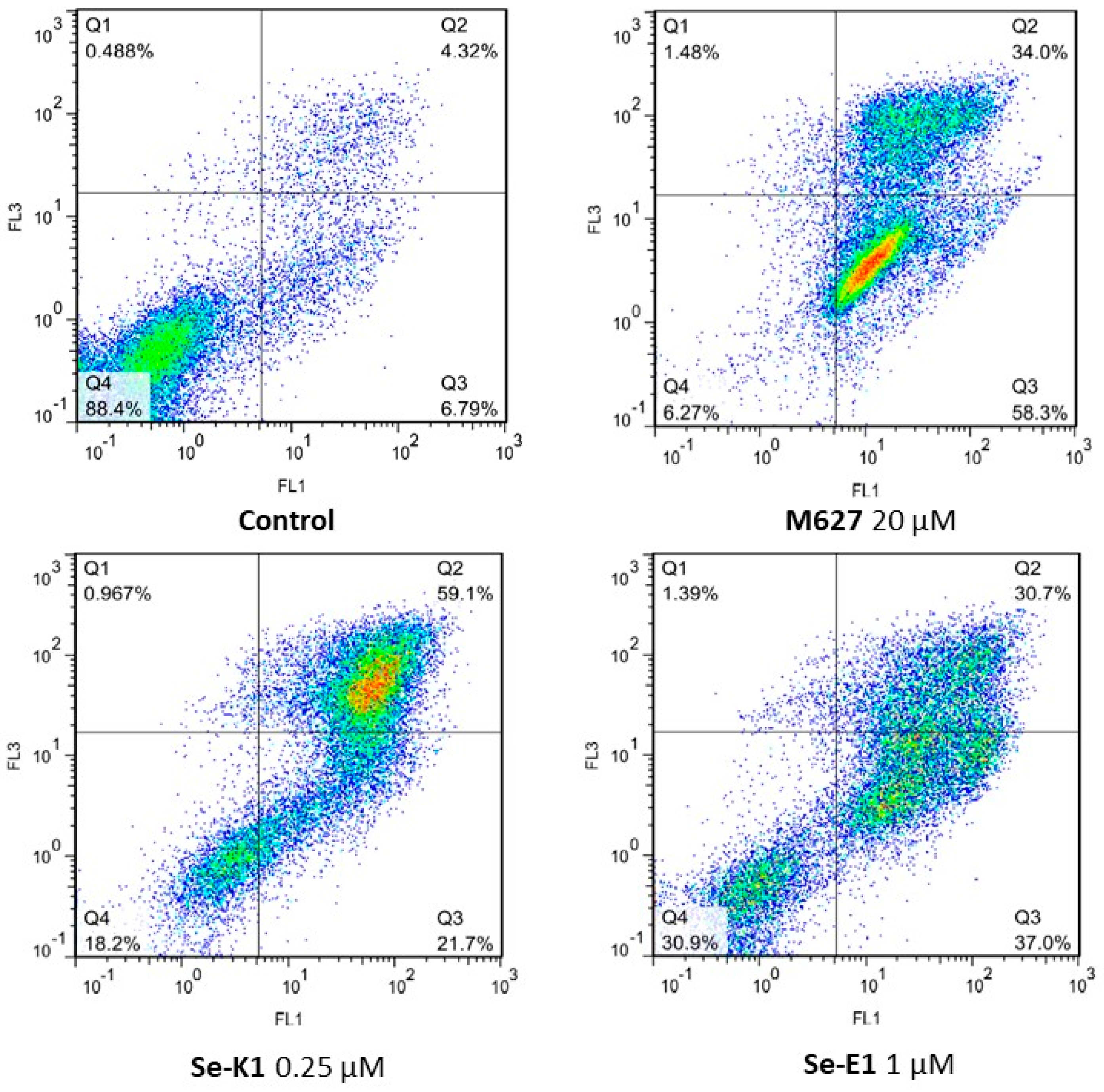
3.3. Apoptosis Assay
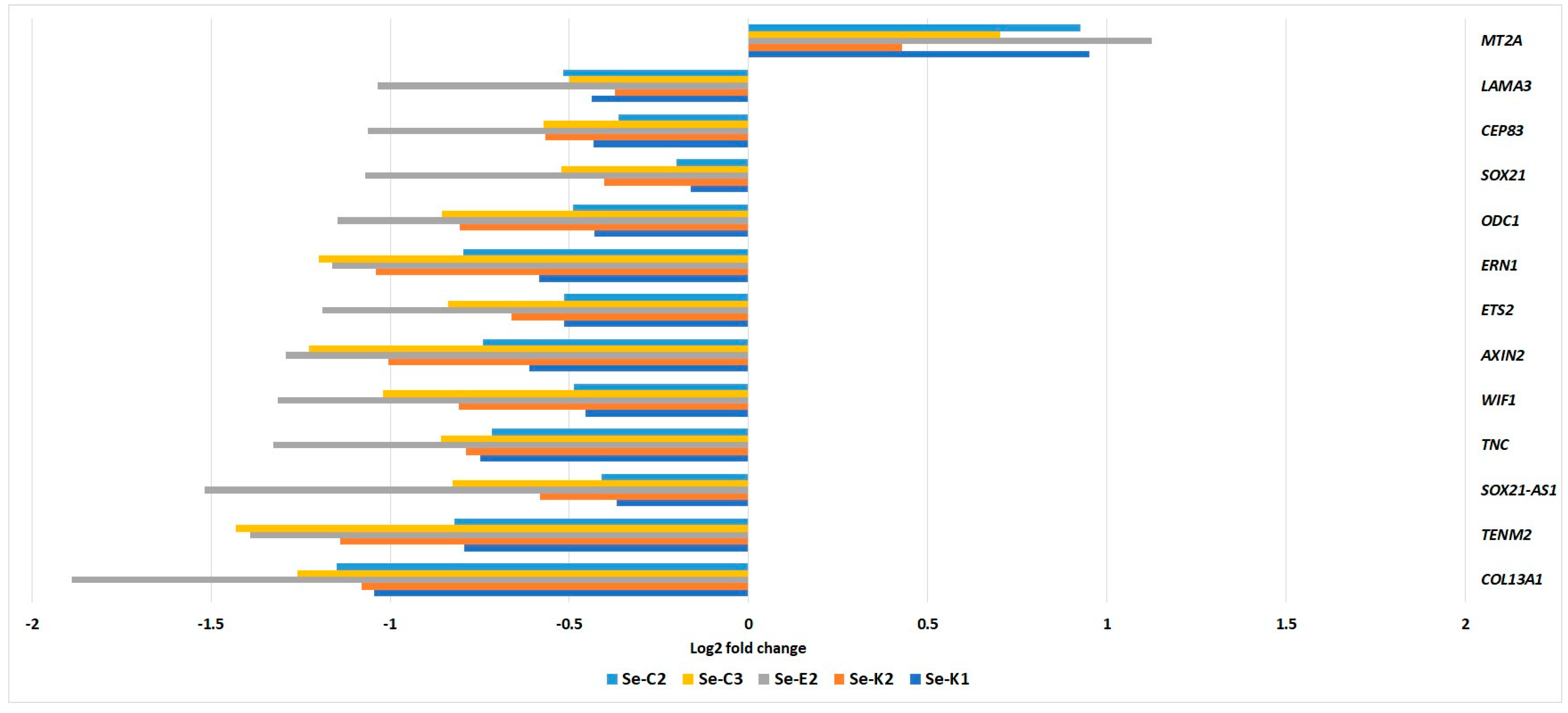
3.4. Transcriptome Analysis
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zugazagoitia, J.; Guedes, C.; Ponce, S.; Ferrer, I.; Molina-Pinelo, S.; Paz-Ares, L. Current Challenges in Cancer Treatment. Clin. Ther. 2016, 38, 1551–1566. [Google Scholar] [CrossRef] [PubMed]
- Auten, R.L.; Davis, J.M. Oxygen Toxicity and Reactive Oxygen Species: The Devil Is in the Details. Pediatr. Res. 2009, 66, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Cross, C.E. Oxygen-Derived Species: Their Relation to Human Disease and Environmental Stress. Environ. Health Perspect. 1994, 102, 5–12. [Google Scholar] [CrossRef]
- Ray, P.D.; Huang, B.-W.; Tsuji, Y. Reactive Oxygen Species (ROS) Homeostasis and Redox Regulation in Cellular Signaling. Cell. Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef]
- Liou, G.-Y.; Storz, P. Reactive Oxygen Species in Cancer. Free Radic. Res. 2010, 44, 479–496. [Google Scholar] [CrossRef] [PubMed]
- Reczek, C.R.; Chandel, N.S. The Two Faces of Reactive Oxygen Species in Cancer. Annu. Rev. Cancer Biol. 2017, 1, 79–98. [Google Scholar] [CrossRef]
- Zhou, J. (Ed.) Multi-Drug Resistance in Cancer; Springer Protocols; Humana Press: New York, NY, USA, 2010; ISBN 978-1-60761-415-9. [Google Scholar]
- Ayob, A.Z.; Ramasamy, T.S. Cancer Stem Cells as Key Drivers of Tumour Progression. J. Biomed. Sci. 2018, 25, 20. [Google Scholar] [CrossRef]
- Cho, Y.; Kim, Y.K. Cancer Stem Cells as a Potential Target to Overcome Multidrug Resistance. Front. Oncol. 2020, 10, 764. [Google Scholar] [CrossRef]
- Wang, H.; Gao, Z.; Liu, X.; Agarwal, P.; Zhao, S.; Conroy, D.W.; Ji, G.; Yu, J.; Jaroniec, C.P.; Liu, Z.; et al. Targeted Production of Reactive Oxygen Species in Mitochondria to Overcome Cancer Drug Resistance. Nat. Commun. 2018, 9, 562. [Google Scholar] [CrossRef]
- Bodnar, M.; Konieczka, P.; Namiesnik, J. The Properties, Functions, and Use of Selenium Compounds in Living Organisms. J. Environ. Sci. Health Part C 2012, 30, 225–252. [Google Scholar] [CrossRef]
- Rayman, M.P. Selenium Intake, Status, and Health: A Complex Relationship. Hormones 2020, 19, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Roman, M.; Jitaru, P.; Barbante, C. Selenium Biochemistry and Its Role for Human Health. Metallomics 2014, 6, 25–54. [Google Scholar] [CrossRef]
- Razaghi, A.; Poorebrahim, M.; Sarhan, D.; Björnstedt, M. Selenium Stimulates the Antitumour Immunity: Insights to Future Research. Eur. J. Cancer 2021, 155, 256–267. [Google Scholar] [CrossRef]
- Kieliszek, M.; Lipinski, B.; Błażejak, S. Application of Sodium Selenite in the Prevention and Treatment of Cancers. Cells 2017, 6, 39. [Google Scholar] [CrossRef]
- Redman, C.; Scott, J.A.; Baines, A.T.; Basye, J.L.; Clark, L.C.; Calley, C.; Roe, D.; Payne, C.M.; Nelson, M.A. Inhibitory Effect of Selenomethionine on the Growth of Three Selected Human Tumor Cell Lines. Cancer Lett. 1998, 125, 103–110. [Google Scholar] [CrossRef]
- Hatfield, D.L.; Tsuji, P.A.; Carlson, B.A.; Gladyshev, V.N. Selenium and Selenocysteine: Roles in Cancer, Health, and Development. Trends Biochem. Sci. 2014, 39, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Varlamova, E.G.; Turovsky, E.A. The main cytotoxic effects of methylseleninic acid on various cancer cells. Int. J. Mol. Sci. 2021, 22, 6614. [Google Scholar] [CrossRef] [PubMed]
- Marć, M.A.; Kincses, A.; Rácz, B.; Nasim, M.J.; Sarfraz, M.; Lázaro-Milla, C.; Domínguez-Álvarez, E.; Jacob, C.; Spengler, G.; Almendros, P. Antimicrobial, Anticancer and Multidrug-Resistant Reversing Activity of Novel Oxygen-, Sulfur- and Selenoflavones and Bioisosteric Analogues. Pharmaceuticals 2020, 13, 453. [Google Scholar] [CrossRef] [PubMed]
- Kharma, A.; Misak, A.; Grman, M.; Brezova, V.; Kurakova, L.; Baráth, P.; Jacob, C.; Chovanec, M.; Ondrias, K.; Domínguez-Álvarez, E. Release of Reactive Selenium Species from Phthalic Selenoanhydride in the Presence of Hydrogen Sulfide and Glutathione with Implications for Cancer Research. New J. Chem. 2019, 43, 11771–11783. [Google Scholar] [CrossRef]
- Santi, C.; Scimmi, C.; Sancineto, L. Ebselen and Analogues: Pharmacological Properties and Synthetic Strategies for Their Preparation. Molecules 2021, 26, 4230. [Google Scholar] [CrossRef] [PubMed]
- Gandin, V.; Khalkar, P.; Braude, J.; Fernandes, A.P. Organic Selenium Compounds as Potential Chemotherapeutic Agents for Improved Cancer Treatment. Free Radic. Biol. Med. 2018, 127, 80–97. [Google Scholar] [CrossRef]
- Collery, P. Strategies for the Development of Selenium-Based Anticancer Drugs. J. Trace Elem. Med. Biol. 2018, 50, 498–507. [Google Scholar] [CrossRef]
- Domínguez-Álvarez, E.; Rácz, B.; Marć, M.A.; Nasim, M.J.; Szemerédi, N.; Viktorová, J.; Jacob, C.; Spengler, G. Selenium and Tellurium in the Development of Novel Small Molecules and Nanoparticles as Cancer Multidrug Resistance Reversal Agents. Drug Resist. Updates 2022, 63, 100844. [Google Scholar] [CrossRef] [PubMed]
- Gajdács, M.; Spengler, G.; Sanmartín, C.; Marć, M.A.; Handzlik, J.; Domínguez-Álvarez, E. Selenoesters and Selenoanhydrides as Novel Multidrug Resistance Reversing Agents: A Confirmation Study in a Colon Cancer MDR Cell Line. Bioorg. Med. Chem. Lett. 2017, 27, 797–802. [Google Scholar] [CrossRef]
- Marć, M.A.; Domínguez-Álvarez, E.; Latacz, G.; Doroz-Płonka, A.; Sanmartín, C.; Spengler, G.; Handzlik, J. Pharmaceutical and Safety Profile Evaluation of Novel Selenocompounds with Noteworthy Anticancer Activity. Pharmaceutics 2022, 14, 367. [Google Scholar] [CrossRef]
- Jamier, V.; Ba, L.A.; Jacob, C. Selenium- and Tellurium-Containing Multifunctional Redox Agents as Biochemical Redox Modulators with Selective Cytotoxicity. Chem. Eur. J. 2010, 16, 10920–10928. [Google Scholar] [CrossRef] [PubMed]
- Sanmartín, C.; Font, M.; Palop, J. Molecular Symmetry: A Structural Property Frequently Present in New Cytotoxic and Proapoptotic Drugs. Mini-Rev. Med. Chem. 2006, 6, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Álvarez, E.; Plano, D.; Font, M.; Calvo, A.; Prior, C.; Jacob, C.; Palop, J.A.; Sanmartín, C. Synthesis and Antiproliferative Activity of Novel Selenoester Derivatives. Eur. J. Med. Chem. 2014, 73, 153–166. [Google Scholar] [CrossRef]
- Domínguez-Álvarez, E.; Spengler, G.; Jacob, C.; Sanmartín, C. Selenoester-Containing Compounds for Use in the Treatment of Microbial Infections or Colorectal Cancer. European Patent EP3628659A1, 28 September 2018. [Google Scholar]
- Szemerédi, N.; Dobiasová, S.; Salardón-Jiménez, N.; Kincses, A.; Nové, M.; Habibullah, G.; Sevilla-Hernández, C.; Benito-Lama, M.; Alonso-Martínez, F.-J.; Viktorová, J.; et al. Cyano- and Ketone-Containing Selenoesters as Multi-Target Compounds against Resistant Cancers. Cancers 2021, 13, 4563. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.-C. Theoretical Basis, Experimental Design, and Computerized Simulation of Synergism and Antagonism in Drug Combination Studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef]
- Chou, T.-C. Drug Combination Studies and Their Synergy Quantification Using the Chou-Talalay Method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize Analysis Results for Multiple Tools and Samples in a Single Report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet. J. 2011, 17, 10. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential Expression Analysis of Multifactor RNA-Seq Experiments with Respect to Biological Variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef]
- Chen, Y.; Lun, A.T.L.; Smyth, G.K. From Reads to Genes to Pathways: Differential Expression Analysis of RNA-Seq Experiments Using Rsubread and the EdgeR Quasi-Likelihood Pipeline. F1000Research 2016, 5, 1438. [Google Scholar] [CrossRef]
- Zu, Y.; Yang, Z.; Tang, S.; Han, Y.; Ma, J. Effects of P-Glycoprotein and Its Inhibitors on Apoptosis in K562 Cells. Molecules 2014, 19, 13061–13075. [Google Scholar] [CrossRef]
- Turella, P.; Filomeni, G.; Dupuis, M.L.; Ciriolo, M.R.; Molinari, A.; De Maria, F.; Tombesi, M.; Cianfriglia, M.; Federici, G.; Ricci, G.; et al. A Strong Glutathione S-Transferase Inhibitor Overcomes the P-Glycoprotein-Mediated Resistance in Tumor Cells. J. Biol. Chem. 2006, 281, 23725–23732. [Google Scholar] [CrossRef]
- Ling, X.-B.; Wei, H.-W.; Wang, J.; Kong, Y.-Q.; Wu, Y.-Y.; Guo, J.-L.; Li, T.-F.; Li, J.-K. Mammalian Metallothionein-2A and Oxidative Stress. Int. J. Mol. Sci. 2016, 17, 1483. [Google Scholar] [CrossRef]
- Latvanlehto, A.; Snellman, A.; Tu, H.; Pihlajaniemi, T. Type XIII Collagen and Some Other Transmembrane Collagens Contain Two Separate Coiled-Coil Motifs, Which May Function as Independent Oligomerization Domains. J. Biol. Chem. 2003, 278, 37590–37599. [Google Scholar] [CrossRef]
- Tian, J.; Li, Z.; Jiang, Y.; Gu, W.; Kong, E.; Hao, Q.; Kong, B.; Sun, L. Downregulation of SOX21-AS1 Alleviated Cisplatin Resistance in Cervical Cancer Through Epithelial–Mesenchymal Transition Inhibition. Rejuvenation Res. 2022, 25, 243–252. [Google Scholar] [CrossRef]
- Yu, D.; Zhao, Z.; Wang, L.; Qiao, S.; Yang, Z.; Wen, Q.; Zhu, G. SOX21-AS1 Activated by STAT6 Promotes Pancreatic Cancer Progression via up-Regulation of SOX21. J. Transl. Med. 2022, 20, 511. [Google Scholar] [CrossRef]
- Hogarty, M.D.; Norris, M.D.; Davis, K.; Liu, X.; Evageliou, N.F.; Hayes, C.S.; Pawel, B.; Guo, R.; Zhao, H.; Sekyere, E.; et al. ODC1 Is a Critical Determinant of MYCN Oncogenesis and a Therapeutic Target in Neuroblastoma. Cancer Res. 2008, 68, 9735–9745. [Google Scholar] [CrossRef]
- Raymundo, D.P.; Doultsinos, D.; Guillory, X.; Carlesso, A.; Eriksson, L.A.; Chevet, E. Pharmacological Targeting of IRE1 in Cancer. Trends Cancer 2020, 6, 1018–1030. [Google Scholar] [CrossRef]
- Fu, L.; Fu, H.; Wu, Q.; Pang, Y.; Xu, K.; Zhou, L.; Qiao, J.; Ke, X.; Xu, K.; Shi, J. High Expression of ETS2 Predicts Poor Prognosis in Acute Myeloid Leukemia and May Guide Treatment Decisions. J. Transl. Med. 2017, 15, 159. [Google Scholar] [CrossRef] [PubMed]
- Sarmiento-Salinas, F.L.; Perez-Gonzalez, A.; Acosta-Casique, A.; Ix-Ballote, A.; Diaz, A.; Treviño, S.; Rosas-Murrieta, N.H.; Millán-Perez-Peña, L.; Maycotte, P. Reactive Oxygen Species: Role in Carcinogenesis, Cancer Cell Signaling and Tumor Progression. Life Sci. 2021, 284, 119942. [Google Scholar] [CrossRef] [PubMed]
- Nové, M.; Kincses, A.; Szalontai, B.; Rácz, B.; Blair, J.M.A.; González-Prádena, A.; Benito-Lama, M.; Domínguez-Álvarez, E.; Spengler, G. Biofilm Eradication by Symmetrical Selenoesters for Food-Borne Pathogens. Microorganisms 2020, 8, 566. [Google Scholar] [CrossRef] [Green Version]
- Dobiasová, S.; Szemerédi, N.; Kučerová, D.; Koucká, K.; Václavíková, R.; Gbelcová, H.; Ruml, T.; Domínguez-Álvarez, E.; Spengler, G.; Viktorová, J. Ketone-Selenoesters as Potential Anticancer and Multidrug Resistance Modulation Agents in 2D and 3D Ovarian and Breast Cancer in Vitro Models. Sci. Rep. 2022, 12, 6548. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Ratio: EDAG:Doxorubicin | CI | Interaction | SD |
|---|---|---|---|---|
| Se-K1 | 0.96:1 | 1.60 | Antagonism | 0.59 |
| Se-K2 | 0.24:1 | 0.37 | Synergism | 0.10 |
| Se-K3 | 0.1:1 | 3.41 | Strong antagonism | 1.47 |
| Se-E1 | 18.56:1 | 0.85 | Weak synergism | 0.12 |
| Se-E2 | 9.28:1 | 0.86 | Weak synergism | 0.20 |
| Se-E3 | 6.96:1 | 0.87 | Weak synergism | 0.22 |
| Se-C1 | 0.92:1 | 2.63 | Antagonism | 1.51 |
| Se-C2 | 1.16:1 | 1.55 | Antagonism | 0.43 |
| Se-C3 | 0.23:1 | 1.09 | Weak antagonism | 0.13 |
| Compounds | Concentration (µM) | FSC | SSC | FL-1 | FAR | FAR Quotient (1.00: Verapamil FAR) |
|---|---|---|---|---|---|---|
| Verapamil | 20 | 2398 | 914 | 20.4 | 3.22 | 1.00 |
| Se-K1 | 2 | 2335 | 884 | 69.7 | 11.01 | 3.42 |
| 20 | 2339 | 1459 | 47.2 | 7.46 | 2.32 | |
| Se-K2 | 2 | 2384 | 950 | 66.5 | 10.51 | 3.26 |
| 20 | 2326 | 1414 | 52.9 | 8.36 | 2.60 | |
| Se-K3 | 2 | 2394 | 907 | 67.3 | 10.63 | 3.30 |
| 20 | 2287 | 1532 | 45.1 | 7.12 | 2.21 | |
| Se-E1 | 2 | 2390 | 893 | 5.19 | 0.82 | 0.255 |
| 20 | 2366 | 856 | 6.02 | 0.95 | 0.295 | |
| Se-E2 | 2 | 2436 | 943 | 5.15 | 0.81 | 0.252 |
| 20 | 2350 | 845 | 4.03 | 0.64 | 0.198 | |
| Se-E3 | 2 | 2419 | 896 | 4.45 | 0.70 | 0.217 |
| 20 | 2505 | 894 | 9.66 | 1.53 | 0.475 | |
| Se-C1 | 2 | 2553 | 1012 | 21.5 | 3.40 | 1.06 |
| 20 | 2543 | 987 | 38.8 | 6.13 | 1.90 | |
| Se-C2 | 2 | 2573 | 1055 | 33.2 | 5.24 | 1.63 |
| 20 | 2483 | 939 | 35.2 | 5.56 | 1.73 | |
| Se-C3 | 2 | 2525 | 1064 | 40.1 | 6.33 | 1.97 |
| 20 | 2445 | 973 | 31.9 | 5.04 | 1.57 |
| Compounds | Concentration (µM) | Early Apoptosis % | Late Apoptosis % | Cell Death % |
|---|---|---|---|---|
| Untreated | - | 6.79 | 4.32 | 0.488 |
| M627 | 20 | 58.3 | 34 | 1.48 |
| Se-K1 | 0.25 | 21.7 | 59.1 | 0.967 |
| Se-K2 | 0.25 | 17.1 | 39.9 | 1.89 |
| Se-K3 | 0.25 | 17.8 | 42.3 | 1.49 |
| Se-E1 | 1 | 37 | 30.7 | 1.39 |
| Se-E2 | 1 | 18.1 | 21.6 | 0.945 |
| Se-E3 | 1 | 21.1 | 17 | 0.45 |
| Se-C1 | 0.5 | 13.2 | 12.1 | 0.646 |
| Se-C2 | 0.5 | 10.5 | 8.35 | 0.519 |
| Se-C3 | 0.5 | 11.1 | 11.9 | 0.569 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rácz, B.; Kincses, A.; Laczi, K.; Rákhely, G.; Domínguez-Álvarez, E.; Spengler, G. Reversal of Multidrug Resistance by Symmetrical Selenoesters in Colon Adenocarcinoma Cells. Pharmaceutics 2023, 15, 610. https://doi.org/10.3390/pharmaceutics15020610
Rácz B, Kincses A, Laczi K, Rákhely G, Domínguez-Álvarez E, Spengler G. Reversal of Multidrug Resistance by Symmetrical Selenoesters in Colon Adenocarcinoma Cells. Pharmaceutics. 2023; 15(2):610. https://doi.org/10.3390/pharmaceutics15020610
Chicago/Turabian StyleRácz, Bálint, Annamária Kincses, Krisztián Laczi, Gábor Rákhely, Enrique Domínguez-Álvarez, and Gabriella Spengler. 2023. "Reversal of Multidrug Resistance by Symmetrical Selenoesters in Colon Adenocarcinoma Cells" Pharmaceutics 15, no. 2: 610. https://doi.org/10.3390/pharmaceutics15020610