
A Multifunctional Hybrid Nanocarrier for Non-Invasive siRNA Delivery to the Retina
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Cells
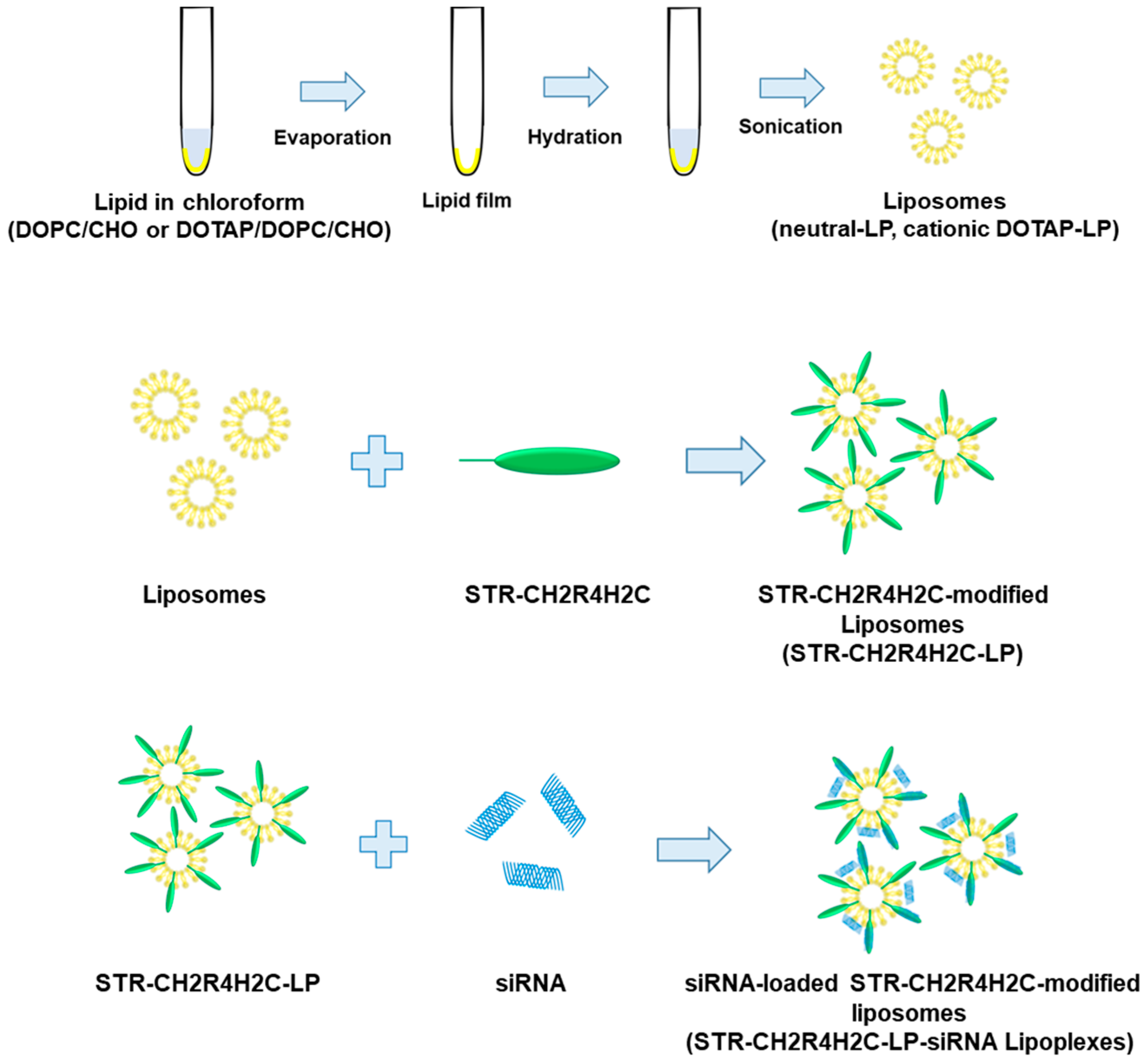
2.2. Preparation of STR-CH2R4H2C-LP and STR-CH2R4H2C-LP-siRNA Lipoplexes
2.3. Physical Properties of the STR-CH2R4H2C-LP-siRNA Lipoplexes
2.4. Cellular Uptake of STR-CH2R4H2C-LP-siRNA Lipoplexes in RPE Cells
2.5. Evaluation of siRNA Intracellular Dynamics
2.6. Suppression of VEGF Expression in RPE Cells with STR-CH2R4H2C-LP-siVEGF Lipoplexes
2.7. Intraocular Migration of STR-CH2R4H2C-LP Following Instillation into the Rat Eye
2.8. Statistical Analysis
3. Results
3.1. Physical Properties of the STR-CH2R4H2C-LP-siRNA Lipoplexes
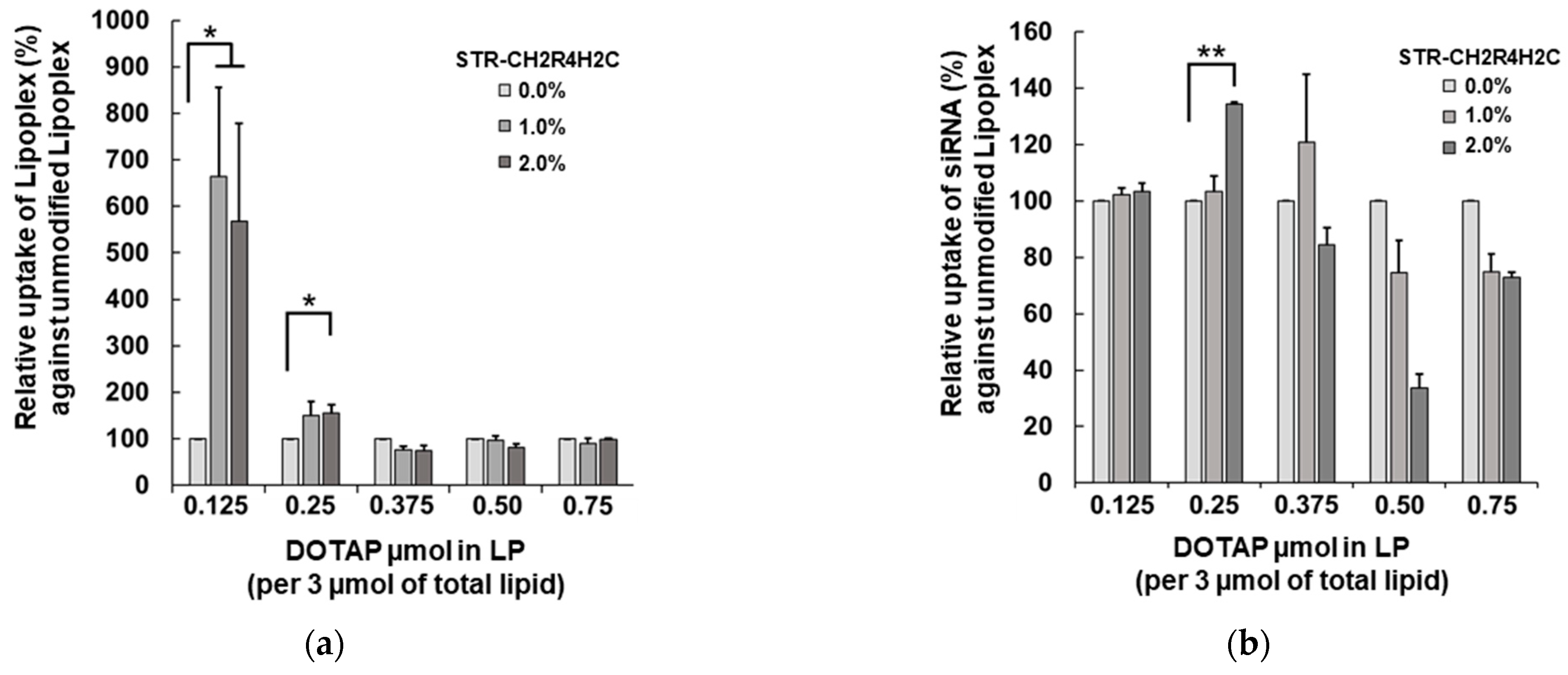
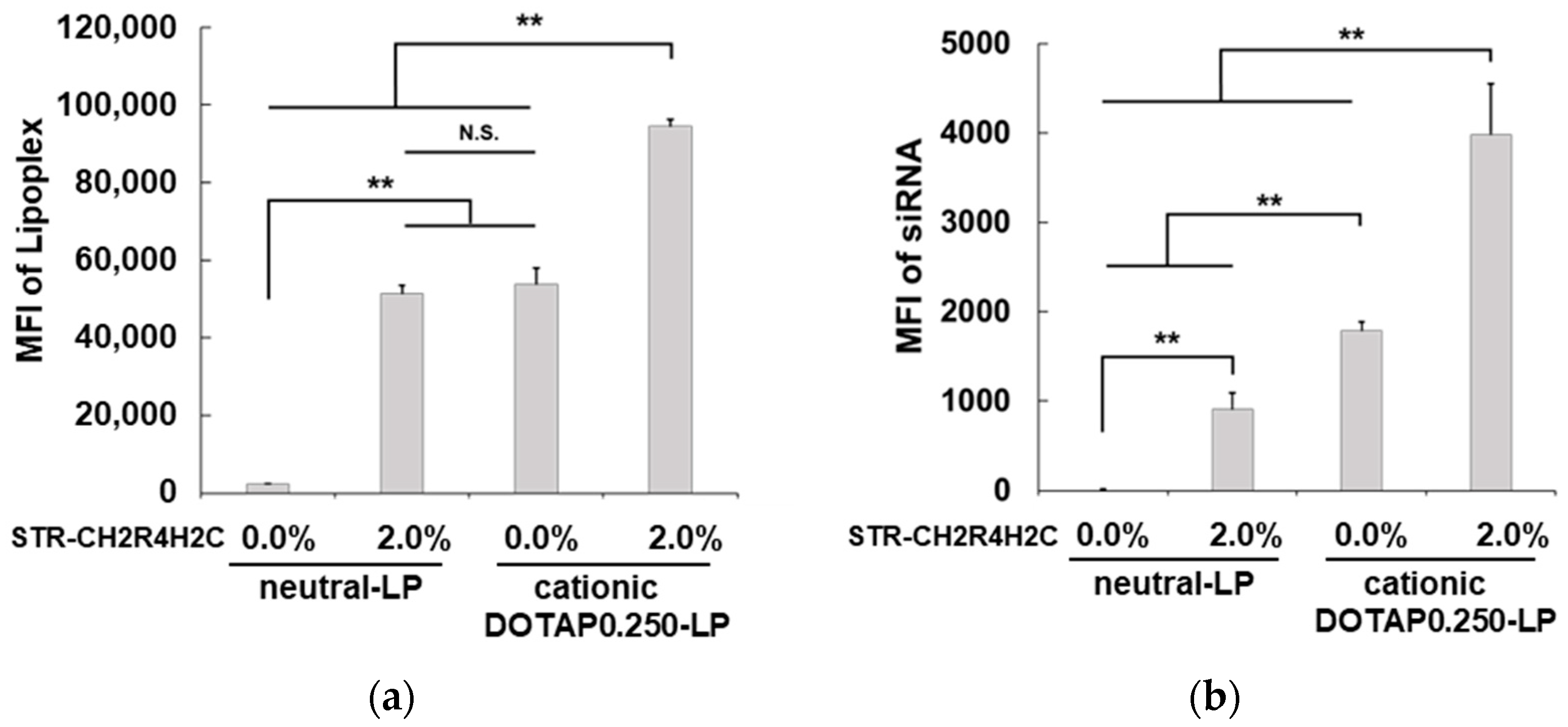
3.2. Cellular Uptake of STR-CH2R4H2C-LP-siRNA Lipoplexes and siRNA Transfection in RPE Cells
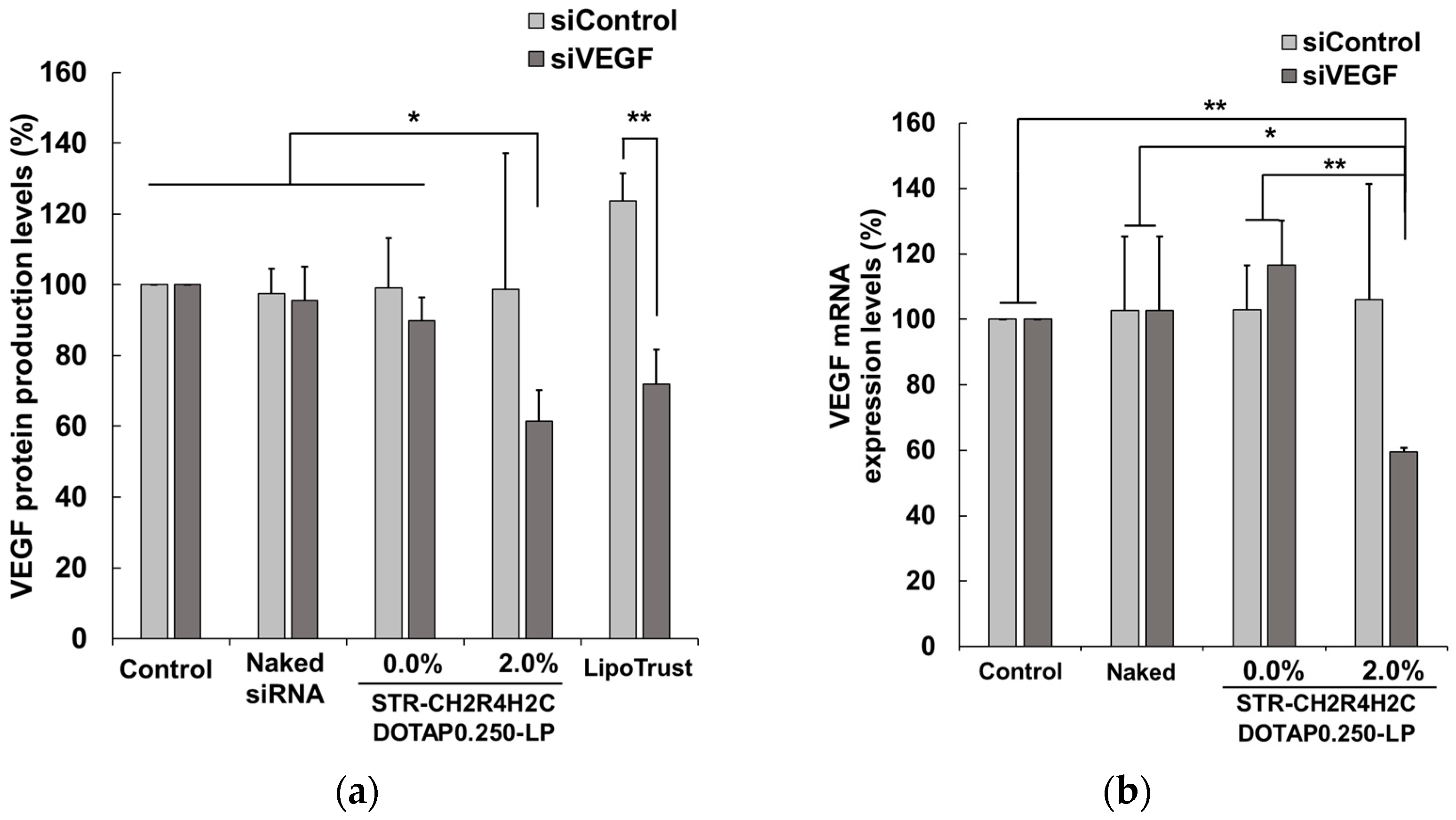
3.3. Advantages of STR-CH2R4H2C-LP-siRNA Lipoplexes for the Intracellular Dynamics of siRNA and siRNA Activity in RPE Cells
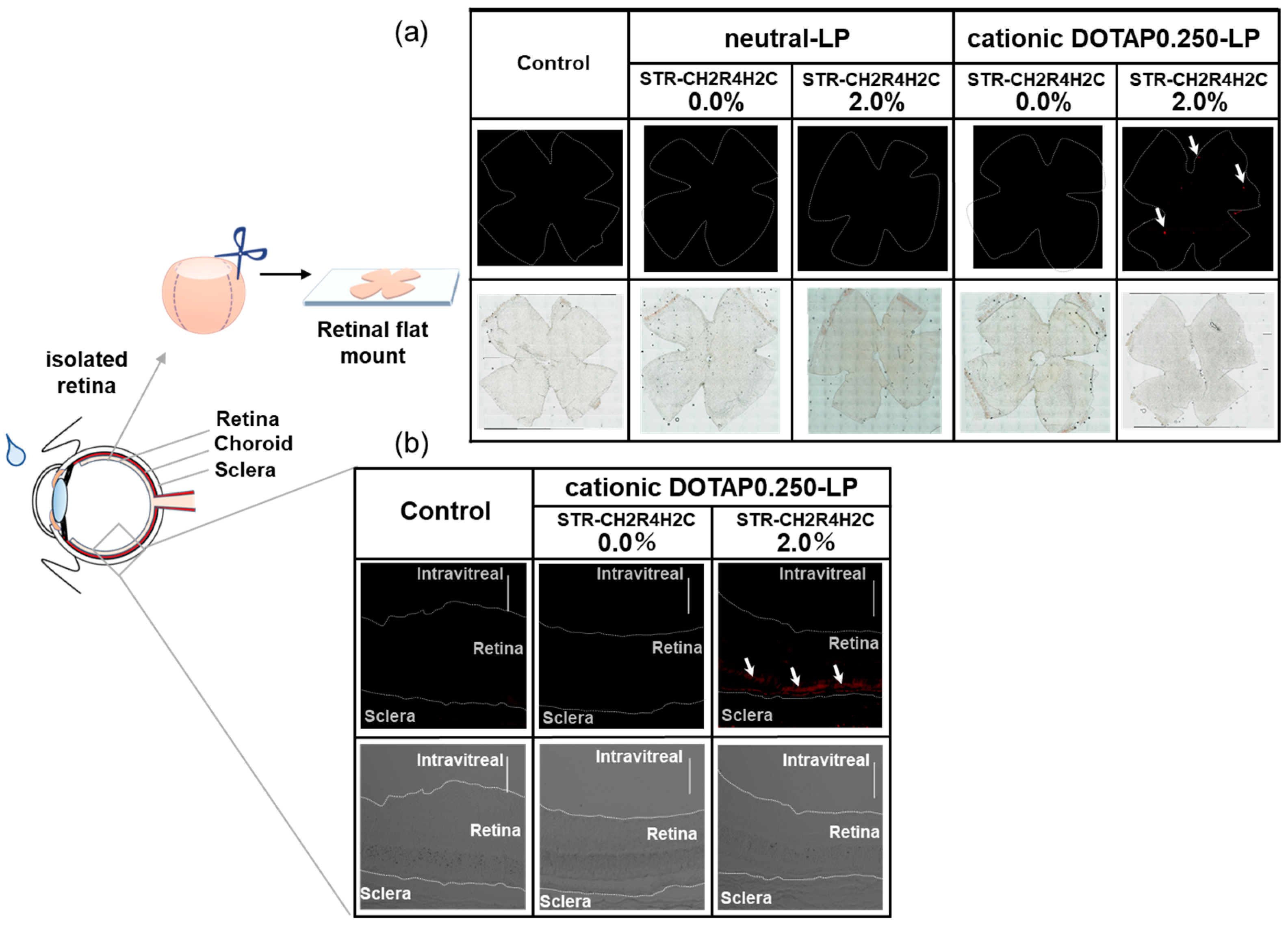
3.4. Intraocular Migration of STR-CH2R4H2C-LP Following Instillation into Rat Eyes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wan, L.W.; Xinyi, S.; Xiang, L.; Chui, M.C.; Ronald, K.; Ching-Yu, C.; Tien, Y.W. Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: A systematic review and meta-analysis. Lancet Glob. Health 2014, 2, e106–e116. [Google Scholar] [CrossRef]
- Cheung, N.; Mitchell, P.; Wong, T.Y. Diabetic retinopathy. Lancet 2010, 376, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Ian, J.M.; Trevor, G.R. Obesity and post-prandial lipid metabolism. Feast or famine? J. Nutr. Biochem. 2004, 15, 130–141. [Google Scholar] [CrossRef]
- Chakravarthy, U.; Bailey, C.C.; Scanlon, P.H.; McKibbin, M.; Khan, R.S.; Mahmood, S.; Downey, L.; Dhingra, N.; Brand, C.; Brittain, C.; et al. Progression from Early/Intermediate to Advanced Forms of Age-Related Macular Degeneration in a Large UK Cohort: Rates and Risk Factors. Ophthalmol. Retina 2020, 4, 662–672. [Google Scholar] [CrossRef]
- Rudraraju, M.; Narayanan, S.P.; Somanath, P.R. Regulation of blood-retinal barrier cell-junctions in diabetic retinopathy. Pharmacol. Res. 2020, 161, 105115. [Google Scholar] [CrossRef]
- Ma, W.; Lee, S.E.; Guo, J.; Qu, W.; Hudson, B.I.; Schmidt, A.M.; Barile, G.R. RAGE Ligand Upregulation of VEGF Secretion in ARPE-19 Cells. Investig. Ophthalmol. Vis. Sci. 2007, 48, 1355–1361. [Google Scholar] [CrossRef]
- Yamada, Y.; Ishibashi, K.; Ishibashi, K.; Bhutto, I.A.; Tian, J.; Lutty, G.A.; Handa, J.T. The expression of advanced glycation endproduct receptors in rpe cells associated with basal deposits in human maculas. Exp. Eye Res. 2006, 82, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Lakkaraju, A.; Umapathy, A.; Tan, L.X.; Daniele, L.; Philp, N.J.; Boesze-Battaglia, K.; Williams, D.S. The cell biology of the retinal pigment epithelium. Prog. Retin. Eye Res. 2020, 78, 100846. [Google Scholar] [CrossRef]
- Varela-Fernández, R.; Díaz-Tomé, V.; Luaces-Rodríguez, A.; Conde-Penedo, A.; García-Otero, X.; Luzardo-Álvarez, A.; Fernández-Ferreiro, A.; Otero-Espinar, F.J. Drug Delivery to the Posterior Segment of the Eye: Biopharmaceutic and Pharmacokinetic Considerations. Pharmaceutics 2020, 12, 269. [Google Scholar] [CrossRef]
- Ehlers, J.P.; Kim, S.J.; Yeh, S.; Thorne, J.E.; Mruthyunjaya, P.; Schoenberger, S.D.; Bakri, S.J. Therapies for Macular Edema Associated with Branch Retinal Vein Occlusion: A Report by the American Academy of Ophthalmology. Ophthalmology 2017, 124, 1412–1423. [Google Scholar] [CrossRef]
- de Vries, V.A.; Bassil, F.L.; Ramdas, W.D. The effects of intravitreal injections on intraocular pressure and retinal nerve fiber layer: A systematic review and meta-analysis. Sci. Rep. 2020, 10, 13248. [Google Scholar] [CrossRef] [PubMed]
- Mulfaul, K.; Giacalone, J.C.; Voigt, A.P.; Riker, M.J.; Ochoa, D.; Han, I.C.; Stone, E.M.; Mullins, R.F.; Tucker, B.A. Stepwise differentiation and functional characterization of human induced pluripotent stem cell-derived choroidal endothelial cells. Stem Cell Res. Ther. 2020, 11, 409. [Google Scholar] [CrossRef] [PubMed]
- Hayreh, S.S. Posterior Ciliary Artery Circulation in Health and Disease. Investig. Ophthalmol. Vis. Sci. 2004, 45, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Guymer, R.H.; Bird, A.C.; Hageman, G.S. Cytoarchitecture of Choroidal Capillary Endothelial Cells. Investig. Ophthalmol. Vis. Sci. 2004, 45, 1660–1666. [Google Scholar] [CrossRef]
- Gorantla, S.; Rapalli, V.K.; Waghule, T.; Singh, P.P.; Dubey, S.K.; Saha, R.N.; Singhvi, G. Nanocarriers for ocular drug delivery: Current status and translational opportunity. RSC Adv. 2020, 10, 27835–27855. [Google Scholar] [CrossRef] [PubMed]
- Ibaraki, H.; Kanazawa, T.; Oogi, C.; Takashima, Y.; Seta, Y. Effects of surface charge and flexibility of liposomes on dermal drug delivery. JDDST 2019, 50, 155–162. [Google Scholar] [CrossRef]
- Liu, Y.; Ran, R.; Chen, J.; Kuang, Q.; Tang, J.; Mei, L.; Zhang, Q.; Gao, H.; Zhang, Z.; He, Q. Paclitaxel loaded liposomes decorated with a multifunctional tandem peptide for glioma targeting. Biomaterials 2014, 35, 4835–4847. [Google Scholar] [CrossRef]
- Liu, C.; Jiang, K.; Tai, L.; Liu, Y.; Wei, G.; Lu, W.; Pan, W. Facile Noninvasive Retinal Gene Delivery Enabled by Penetratin. ACS Appl. Mater. Interfaces 2016, 8, 19256–19267. [Google Scholar] [CrossRef]
- Jiang, K.; Chen, J.; Tai, L.; Liu, C.; Chen, X.; Wei, G.; Lu, W.; Pan, W. Inhibition of post-trabeculectomy fibrosis via topically instilled antisense oligonucleotide complexes co-loaded with fluorouracil. Acta Pharm. Sin. B 2020, 10, 1754–1768. [Google Scholar] [CrossRef]
- Suda, K.; Murakami, T.; Gotoh, N.; Fukuda, R.; Hashida, Y.; Hashida, M.; Tsujikawa, A.; Yoshimura, N. High-density lipoprotein mutant eye drops for the treatment of posterior eye Diseases. J. Control. Release 2017, 266, 301–309. [Google Scholar] [CrossRef] [Green Version]
- Kanazawa, T.; Taki, H.; Okada, H. Nose-to-brain drug delivery system with ligand/cell-penetrating peptide-modified polymeric nano-micelles for intracerebral gliomas. Eur. J. Pharm. Biopharm. 2020, 152, 85–94. [Google Scholar] [CrossRef]
- Kamei, N.; Takeda-Morishita, M. Brain delivery of insulin boosted by intranasal coadministration with cell-penetrating peptides. J. Control. Release 2015, 197, 105–110. [Google Scholar] [CrossRef]
- Zorko, M.; Langel, Ü. Cell-penetrating Peptides. Methods Mol. Biol. 2022, 2383, 3–32. [Google Scholar] [CrossRef] [PubMed]
- Hirose, H.; Takeuchi, T.; Osakada, H.; Pujals, S.; Katayama, S.; Nakase, I.; Kobayashi, S.; Haraguchi, T.; Futaki, S. Transient Focal Membrane Deformation Induced by Arginine-rich Peptides Leads to Their Direct Penetration into Cells. Mol. Ther. 2012, 20, 984–993. [Google Scholar] [CrossRef] [PubMed]
- Duchardt, F.; Fotin-Mleczek, M.; Schwarz, H.; Fischer, R.; Brock, R. A Comprehensive Model for the Cellular Uptake of Cationic Cell-penetrating Peptides. Traffic 2007, 8, 848–866. [Google Scholar] [CrossRef]
- Ben Djemaa, S.; Hervé-Aubert, K.; Lajoie, L.; Falanga, A.; Galdiero, S.; Nedellec, S.; Soucé, M.; Munnier, E.; Chourpa, I.; David, S.; et al. gH625 Cell-Penetrating Peptide Promotes the Endosomal Escape of Nanovectorized siRNA in a Triple-Negative Breast Cancer. Cell Line Biomacromol. 2019, 20, 3076–3086. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, P.; El-Andaloussi, S.; Sütlü, T.; Johansson, H.; Langel, U. Delivery of short interfering RNA using endosomolytic cell-penetrating peptides. FASEB J. 2007, 21, 2664–2671. [Google Scholar] [CrossRef]
- Tai, W.; Gao, X. Functional peptides for siRNA delivery. Adv. Drug Deliv. Rev. 2017, 110–111, 157–168. [Google Scholar] [CrossRef]
- Tanaka, K.; Kanazawa, T.; Ogawa, T.; Takashima, Y.; Fukuda, T.; Okada, H. Disulfide crosslinked stearoyl carrier peptides containing arginine and histidine enhance siRNA uptake and gene silencing. Int. J. Pharm. 2010, 398, 219–224. [Google Scholar] [CrossRef]
- Tanaka, K.; Kanazawa, T.; Ogawa, T.; Suda, Y.; Takashima, Y.; Fukuda, T.; Okada, H. A novel, bio-reducible gene vector containing arginine and histidine enhances gene transfection and expression of plasmid DNA. Chem. Pharm. Bull. 2011, 59, 202–207. [Google Scholar] [CrossRef] [Green Version]
- Ibaraki, H.; Kanazawa, T.; Takashima, Y.; Okada, H.; Seta, Y. Development of an Innovative Intradermal siRNA Delivery System Using a Combination of a Functional Stearylated Cytoplasm-Responsive Peptide and a Tight Junction-Opening Peptide. Molecules. 2016, 21, 1279. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, T.; Endo, T.; Arima, N.; Ibaraki, H.; Takashima, Y.; Seta, Y. Systemic delivery of small interfering RNA targeting nuclear factor κB in mice with collagen-induced arthritis using arginine-histidine-cysteine based oligopeptide-modified polymer nanomicelles. Int. J. Pharm. 2016, 515, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, T.; Hamasaki, T.; Endo, T.; Tamano, K.; Sogabe, K.; Seta, Y.; Ohgi, T.; Okada, H. Functional peptide nanocarriers for delivery of novel anti-RelA RNA interference agents as a topical treatment of atopic dermatitis. Int. J. Pharm. 2015, 489, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Okada, H.; Ogawa, T.; Tanaka, K.; Kanazawa, T.; Takashima, Y. Cytoplasm-Responsive Delivery Systems for siRNA Using Cell-Penetrating Peptide. J. Drug Deliv. Sci. Technol. 2014, 24, 3–11. [Google Scholar] [CrossRef]
- Ibaraki, H.; Kanazawa, T.; Takashima, Y.; Okada, H.; Seta, Y. Transdermal anti-nuclear kappaB siRNA therapy for atopic dermatitis using a combination of two kinds of functional oligopeptide. Int. J. Pharm. 2018, 542, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Ibaraki, H.; Takeda, A.; Arima, N.; Hatakeyama, N.; Takashima, Y.; Seta, Y.; Kanazawa, T. In Vivo Fluorescence Imaging of Passive Inflammation Site Accumulation of Liposomes via Intravenous Administration Focused on Their Surface Charge and PEG Modification. Pharmaceutics 2021, 13, 104. [Google Scholar] [CrossRef]
- Nakase, I.; Noguchi, K.; Aoki, A.; Takatani-Nakase, T.; Fujii, I.; Futaki, S. Arginine-rich cell-penetrating peptide-modified extracellular vesicles for active macropinocytosis induction and efficient intracellular delivery. Sci. Rep. 2017, 7, 1991. [Google Scholar] [CrossRef]
- Kobayashi, S.; Nakase, I.; Kawabata, N.; Yu, H.H.; Pujals, S.; Imanishi, M.; Giralt, E.; Futaki, S. Cytosolic Targeting of Macromolecules Using a pH-Dependent Fusogenic Peptide in Combination with Cationic Liposomes. Bioconj. Chem. 2009, 20, 953–959. [Google Scholar] [CrossRef]
- Ichimizu, S.; Watanabe, H.; Maeda, H.; Hamasaki, K.; Nakamura, Y.; Chuang, V.T.G.; Kinoshita, R.; Nishida, K.; Tanaka, R.; Enoki, Y.; et al. Design and tuning of a cell-penetrating albumin derivative as a versatile nanovehicle for intracellular drug delivery. J. Control. Release 2018, 277, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Sreekumar, P.G.; Kannan, R.; de Silva, A.T.; Burton, R.; Ryan, S.J.; Hinton, D.R. Thiol regulation of vascular endothelial growth factor-A and its receptors in human retinal pigment epithelial cells. Biochem. Biophys. Res. Commun. 2006, 346, 1200–1206. [Google Scholar] [CrossRef]
- Miranda-Ortiz, H.; Fernández-Figueroa, E.A.; Ruíz-García, E.B.; Muñoz-Rivas, A.; Méndez-Pérez, A.; Méndez-Galván, J.; Astudillo-de la Vega, H.; Gabiño-López, B.; Nava-Monroy, R.; López-Flores A la Torre, A.; et al. Development of an alternative saliva test for diagnosis of SARS-CoV-2 using TRIzol: Adapting to countries with lower incomes looking for a large-scale detection program. PLoS ONE 2021, 16, e0255807. [Google Scholar] [CrossRef]
- Chang, M.M.; Li, A.; Feissner, R.; Ahmad, T. RT-qPCR demonstrates light-dependent AtRBCS1A and AtRBCS3B mRNA expressions in Arabidopsis thaliana leaves. Biochem. Mol. Biol Educ. 2016, 44, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.K., 3rd; West, P.C.; Mattern-Schain, S.I.; Best, M.D.; Kirkpatrick, S.S.; Dieter, R.A., 3rd; Arnold, J.D.; Buckley, M.R.; McNally, M.M.; Freeman, M.B.; et al. Advances in the Formulation and Assembly of Non-Cationic Lipid Nanoparticles for the Medical Application of Gene Therapeutics. Nanomaterials 2021, 11, 825. [Google Scholar] [CrossRef]
- Hwang, S.Y.; Cho, D.Y.; Kim, H.K.; Cho, S.H.; Choo, J.; Yoon, W.J.; Lee, E.K. Preparation of Targeting Proteoliposome by Postinsertion of a Linker Molecule Conjugated with Recombinant Human Epidermal Growth Factor. Bioconj. Chem. 2010, 21, 345–351. [Google Scholar] [CrossRef]
- Kato, N.; Sato, T.; Fuchigami, Y.; Suga, T.; Geng, L.; Tsurumaru, M.; Hagimori, M.; Mukai, H.; Kawakami, S. Synthesis and evaluation of a novel adapter lipid derivative for preparation of cyclic peptide-modified PEGylated liposomes: Application of cyclic RGD peptide. Eur. J. Pharm. Sci. 2022, 176, 106239. [Google Scholar] [CrossRef]
- Carvalheiro, M.; Ferreira-Silva, M.; Holovanchuk, D.; Marinho, H.S.; Moreira, J.N.; Soares, H.; Corvo, M.L.; Cruz, M.E.M. Antagonist G-targeted liposomes for improved delivery of anticancer drugs in small cell lung carcinoma. Int. J. Pharm. 2022, 612, 12138. [Google Scholar] [CrossRef] [PubMed]
- Kolašinac, R.; Bier, D.; Schmitt, L.; Yabluchanskiy, A.; Neumaier, B.; Merkel, R.; Csiszár, A. Delivery of the Radionuclide 131I Using Cationic Fusogenic Liposomes as Nanocarriers. Int. J. Mol. Sci. 2021, 22, 457. [Google Scholar] [CrossRef]
- Collins, E.; Birchall, J.C.; Williams, J.L.; Gumbleton, M. Nuclear localisation and pDNA condensation in non-viral gene delivery. J. Gene Med. 2007, 9, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Rothbard, J.B.; Jessop, T.C.; Lewis, R.S.; Murray, B.A.; Wender, P.A. Role of Membrane Potential and Hydrogen Bonding in the Mechanism of Translocation of Guanidinium-Rich Peptides into Cells. J. Am. Chem. Soc. 2004, 126, 9506–9507. [Google Scholar] [CrossRef] [PubMed]
- Potocky, T.B.; Silvius, J.; Menon, A.K.; Gellman, S.H. HeLa Cell Entry by Guanidinium-Rich β-Peptides: Importance of Specific Cation–Cell Surface Interactions. ChemBioChem 2007, 8, 917–926. [Google Scholar] [CrossRef]
- Mellman, I. Endocytosis and molecular sorting. Annu. Rev. Cell Dev. Biol. 1996, 12, 575–625. [Google Scholar] [CrossRef]
- Jovic, M.; Sharma, M.; Rahajeng, J.; Caplan, S. The early endosome: A busy sorting station for proteins at the crossroads. Histol. Histopathol. 2010, 25, 99–112. [Google Scholar] [CrossRef]
- Lee, D.U.; Park, J.Y.; Kwon, S.; Park, J.Y.; Kim, Y.H.; Khang, D.; Hong, J.H. Apoptotic lysosomal proton sponge effect in tumor tissue by cationic gold nanorods. Nanoscale 2019, 11, 19980–19993. [Google Scholar] [CrossRef]
- ur Rehman, Z.; Hoekstra, D.; Zuhorn, I.S. Mechanism of Polyplex- and Lipoplex-Mediated Delivery of Nucleic Acids: Real-Time Visualization of Transient Membrane Destabilization without Endosomal Lysis. ACS Nano 2013, 7, 3767–3777. [Google Scholar] [CrossRef] [PubMed]
- Lajunen, T.; Hisazumi, K.; Kanazawa, T.; Okada, H.; Seta, Y.; Yliperttula, M.; Urtti, A.; Takashima, Y. Topical drug delivery to retinal pigment epithelium with microfluidizer produced small liposomes. Eur. J. Pharm. Sci. 2014, 62, 23–32. [Google Scholar] [CrossRef]
- Pescina, S.; Ostacolo, C.; Gomez-Monterrey, I.M.; Sala, M.; Bertamino, A.; Sonvico, F.; Padula, C.; Santi, P.; Bianchera, A.; Nicoli, S. Cell penetrating peptides in ocular drug delivery: State of the art. J. Control. Release 2018, 284, 84–102. [Google Scholar] [CrossRef] [PubMed]
- Boote, C.; Sigal, I.A.; Grytz, R.; Hua, Y.; Nguyen, T.D.; Girard, M.J.A. Scleral structure and biomechanics. Prog. Retin. Eye Res. 2020, 74, 100773. [Google Scholar] [CrossRef] [PubMed]
- Pitkänen, L.; Ranta, V.P.; Moilanen, H.; Urtti, A. Permeability of Retinal Pigment Epithelium: Effects of Permeant Molecular Weight and Lipophilicity. Investig. Ophthalmol. Vis. Sci. 2005, 46, 641–646. [Google Scholar] [CrossRef]
- Ambati, J.; Canakis, C.S.; Miller, J.W.; Gragoudas, E.S.; Edwards, A.; Weissgold, D.J.; Kim, I.; Delori, F.C.; Adamis, A.P. Diffusion of High Molecular Weight Compounds through Sclera. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1181–1185. [Google Scholar]
- Cheruvu, N.P.; Kompella, U.B. Bovine and Porcine Transscleral Solute Transport: Influence of Lipophilicity and the Choroid–Bruch’s Layer. Investig. Ophthalmol. Vis. Sci. 2006, 47, 4513–4522. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Estimated ζ-Potential | DOTAP (µmol) | DOPC (µmol) | CHO (µmol) | Total Lipid (µmol) | |
|---|---|---|---|---|---|
| neutral-LP | ±0 mV | - | 1.800 | 1.2 | 3.0 |
| DOTAP0.125-LP | +20 mV | 0.125 | 1.675 | 1.2 | 3.0 |
| DOTAP0.250-LP | +30 mV | 0.250 | 1.550 | 1.2 | 3.0 |
| DOTAP0.375-LP | +40 mV | 0.375 | 1.425 | 1.2 | 3.0 |
| DOTAP0.500-LP | +50 mV | 0.500 | 1.300 | 1.2 | 3.0 |
| DOTAP0.750-LP | +60 mV | 0.750 | 1.050 | 1.2 | 3.0 |
| STR-CH2R4H2C Modified mol% | Particle Size (nm) | PDI | ζ-Potential (mV) | siRNA Loaded Efficiency (%) | |
|---|---|---|---|---|---|
| neutral-LP | 0.0% | 72.1 ± 33.8 | 0.22 | −18.8 ± 8.1 | 26.9 ± 4.0 |
| 2.0% | 157.2 ± 65.8 | 0.17 | −10.6 ± 4.6 | 60.3 ± 12.3 | |
| DOTAP0.125-LP | 0.0% | 104.1 ± 48.6 | 0.22 | 17.7 ± 5.2 | 90.2 ± 2.1 |
| 1.0% | 108.3 ± 59.4 | 0.30 | 15.9 ± 4.3 | 99.2 ± 4.7 | |
| 2.0% | 142.6 ± 86.3 | 0.37 | 20.3 ± 3.4 | 100.8 ± 5.5 | |
| DOTAP0.250-LP | 0.0% | 126.3 ± 56.6 | 0.20 | 11.1 ± 3.9 | 93.8 ± 11.2 |
| 1.0% | 91.8 ± 50.6 | 0.30 | 32.3 ± 6.5 | 77.4 ± 4.6 | |
| 2.0% | 73.9 ± 29.0 | 0.15 | 38.6 ± 7.4 | 90.9 ± 5.2 | |
| DOTAP0.375-LP | 0.0% | 79.7 ± 39.2 | 0.24 | 44.6 ± 14.8 | 77.9 ± 1.5 |
| 1.0% | 90.3 ± 41.3 | 0.21 | 40.4 ± 8.1 | 74.3 ± 3.1 | |
| 2.0% | 72.8 ± 32.9 | 0.25 | 41.1 ± 12.7 | 70.3 ± 6.9 | |
| DOTAP0.500-LP | 0.0% | 70.3 ± 27.5 | 0.15 | 39.4 ± 33.9 | 86.7 ± 1.8 |
| 1.0% | 72.0 ± 30.8 | 0.18 | 46.4 ± 9.0 | 73.7 ± 2.1 | |
| 2.0% | 65.2 ± 26.0 | 0.16 | 49.0 ± 10.0 | 82.6 ± 12.8 | |
| DOTAP0.750-LP | 0.0% | 66.8 ± 28.5 | 0.19 | 54.3 ± 24.0 | 78.9 ± 12.8 |
| 1.0% | 65.5 ± 33.3 | 0.26 | 47.1 ± 14.3 | 71.6 ± 7.8 | |
| 2.0% | 67.6 ± 36.5 | 0.29 | 49.3 ± 15.1 | 65.1 ± 4.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nishida, S.; Takashima, Y.; Udagawa, R.; Ibaraki, H.; Seta, Y.; Ishihara, H. A Multifunctional Hybrid Nanocarrier for Non-Invasive siRNA Delivery to the Retina. Pharmaceutics 2023, 15, 611. https://doi.org/10.3390/pharmaceutics15020611
Nishida S, Takashima Y, Udagawa R, Ibaraki H, Seta Y, Ishihara H. A Multifunctional Hybrid Nanocarrier for Non-Invasive siRNA Delivery to the Retina. Pharmaceutics. 2023; 15(2):611. https://doi.org/10.3390/pharmaceutics15020611
Chicago/Turabian StyleNishida, Shogo, Yuuki Takashima, Ryotaro Udagawa, Hisako Ibaraki, Yasuo Seta, and Hiroshi Ishihara. 2023. "A Multifunctional Hybrid Nanocarrier for Non-Invasive siRNA Delivery to the Retina" Pharmaceutics 15, no. 2: 611. https://doi.org/10.3390/pharmaceutics15020611