
Immunotherapy with Cleavage-Specific 12A12mAb Reduces the Tau Cleavage in Visual Cortex and Improves Visuo-Spatial Recognition Memory in Tg2576 AD Mouse Model
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Ethical Approval
2.2. Immunization Scheme
2.3. Tissue Collection, Harvesting and Preparation
2.4. Western Blot Analysis and Semi-Quantitative Densitometry
2.5. Golgi–Cox Staining and Dendritic Spine Analysis
2.6. Visual Acuity Test (VAT)
- 10 points = direct correct answer (mouse went directly to P1);
- 7.5 points = in between correct answer (borderline behaviour: mouse went to the middle of the P1-arm, but did not run the whole arm);
- 5 points = indirect correct answer (mouse went to P0, right after on P1);
- 0 points = wrong arm (mouse did not explore P1).
- gray pattern (P0) vs. 2 cm standard striped pattern (P1) card in trials 1–10;
- gray pattern (P0) vs. 1.5 cm striped pattern (P2) card in trials 11–20;
- gray pattern (P0) vs. 1 cm striped pattern (P3) card in trials 21–30;
- gray pattern (P0) vs. 0.5 cm striped pattern (P4) card in trials 31–40.
- 0 point = 0–1 correct answer;
- 2.5 points = 2–3 correct answers;
- 5 points = 4–5 correct answers;
- 7.5 points = 6–8 correct answers;
- 10 points = 9–10 correct answers.
2.7. Immunofluorescence on Visual Cortex Cryosections
2.8. Histopathological Analysis
2.9. Data Management and Statistical Analysis
3. Results
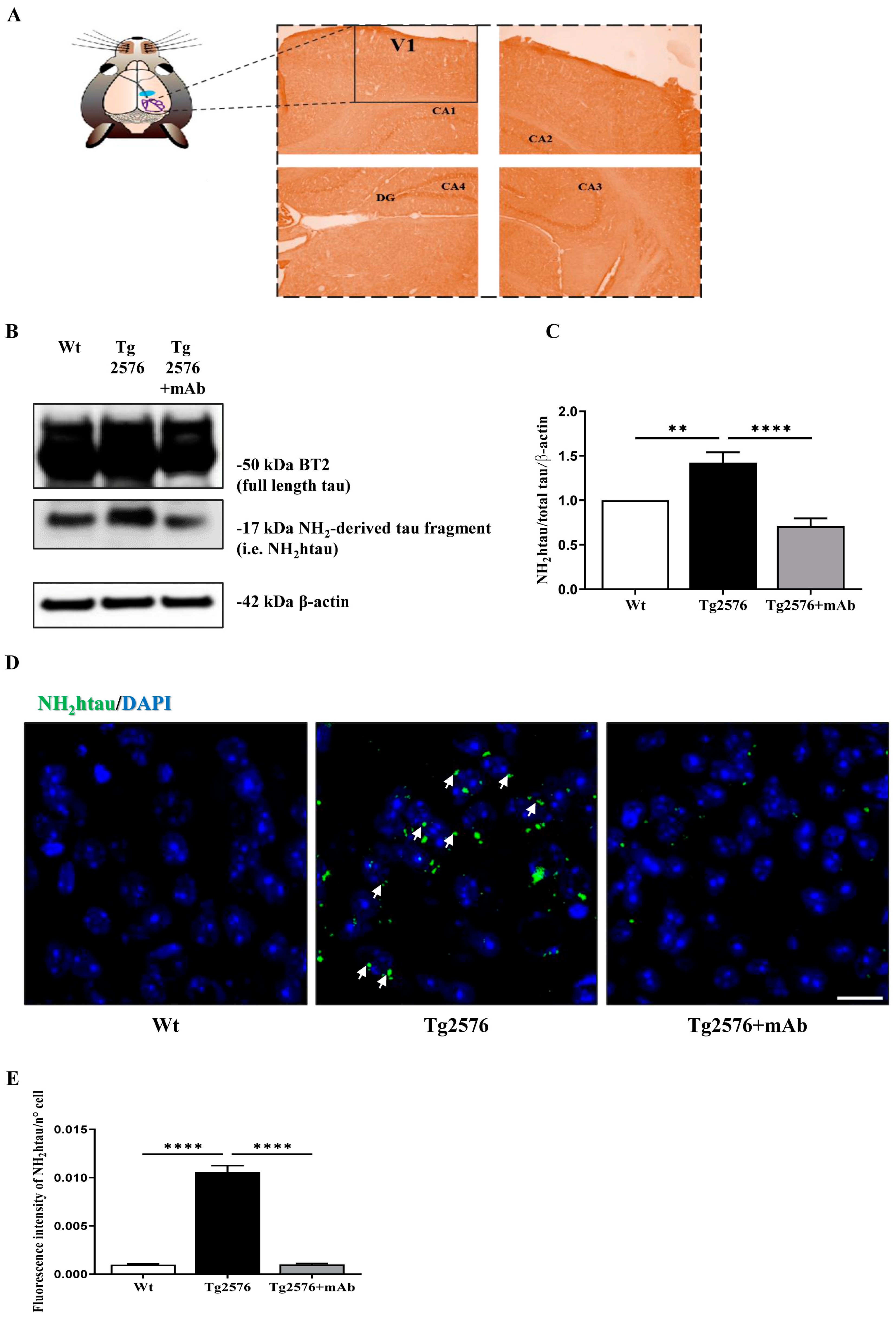
3.1. Abnormal N-Terminal Cleavage of Tau in Primary Visual Cortex (V1 Area) from Tg2576 AD Mice Is Successfully Antagonized by In Vivo Immunization with 12A12mAb
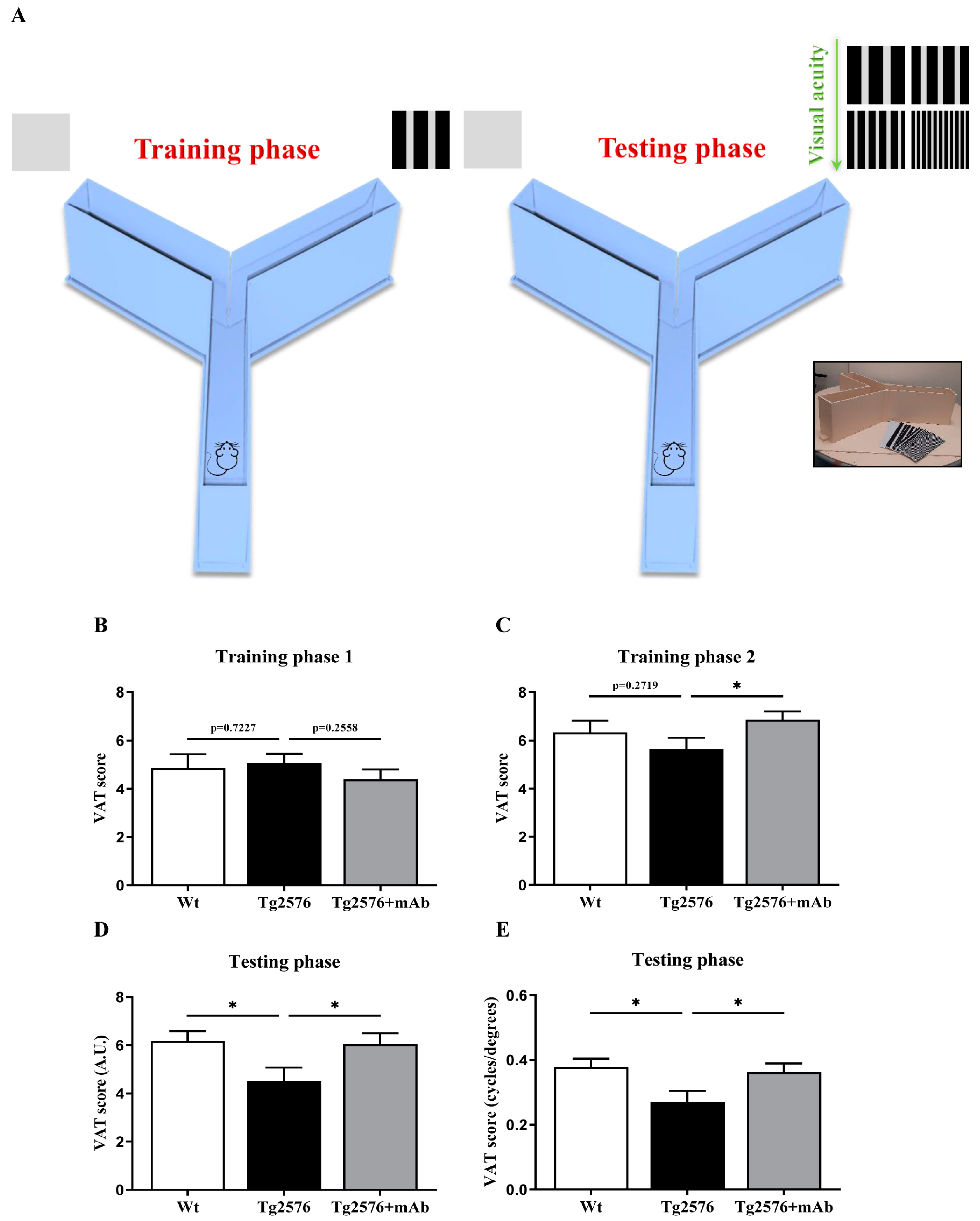
3.2. In Vivo Functional Alterations of Visuo-Spatial Skills Are Recovered by 12A12mAb Immunization in Tg2576 AD Mice
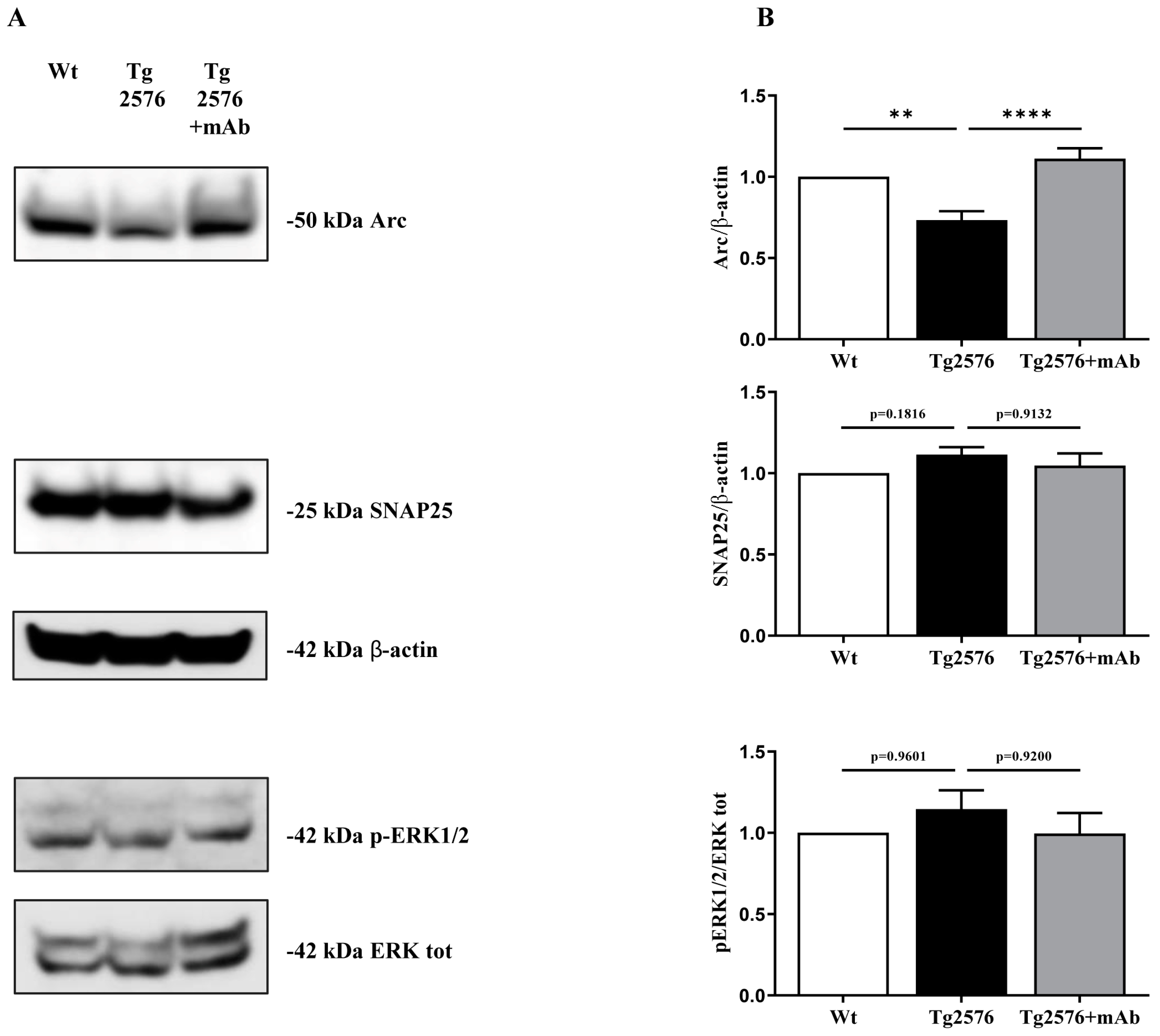
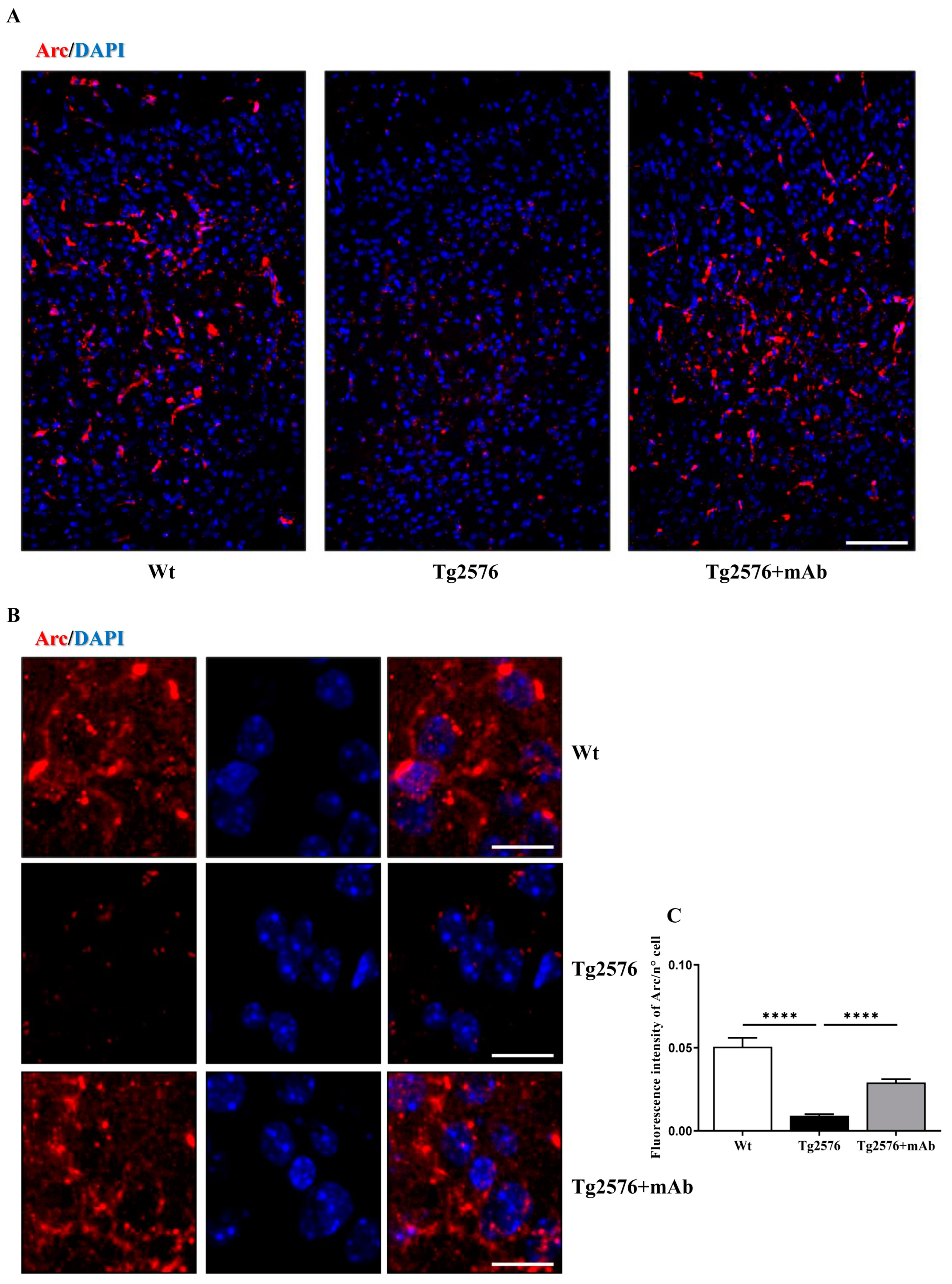
3.3. Treatment of Tg2576 AD Mice with 12A12mAb Normalizes the Changes in the Neural Expression Pattern of Experience-Dependent Markers Closely Coupled to the Synaptic Plasticity of the V1 Area
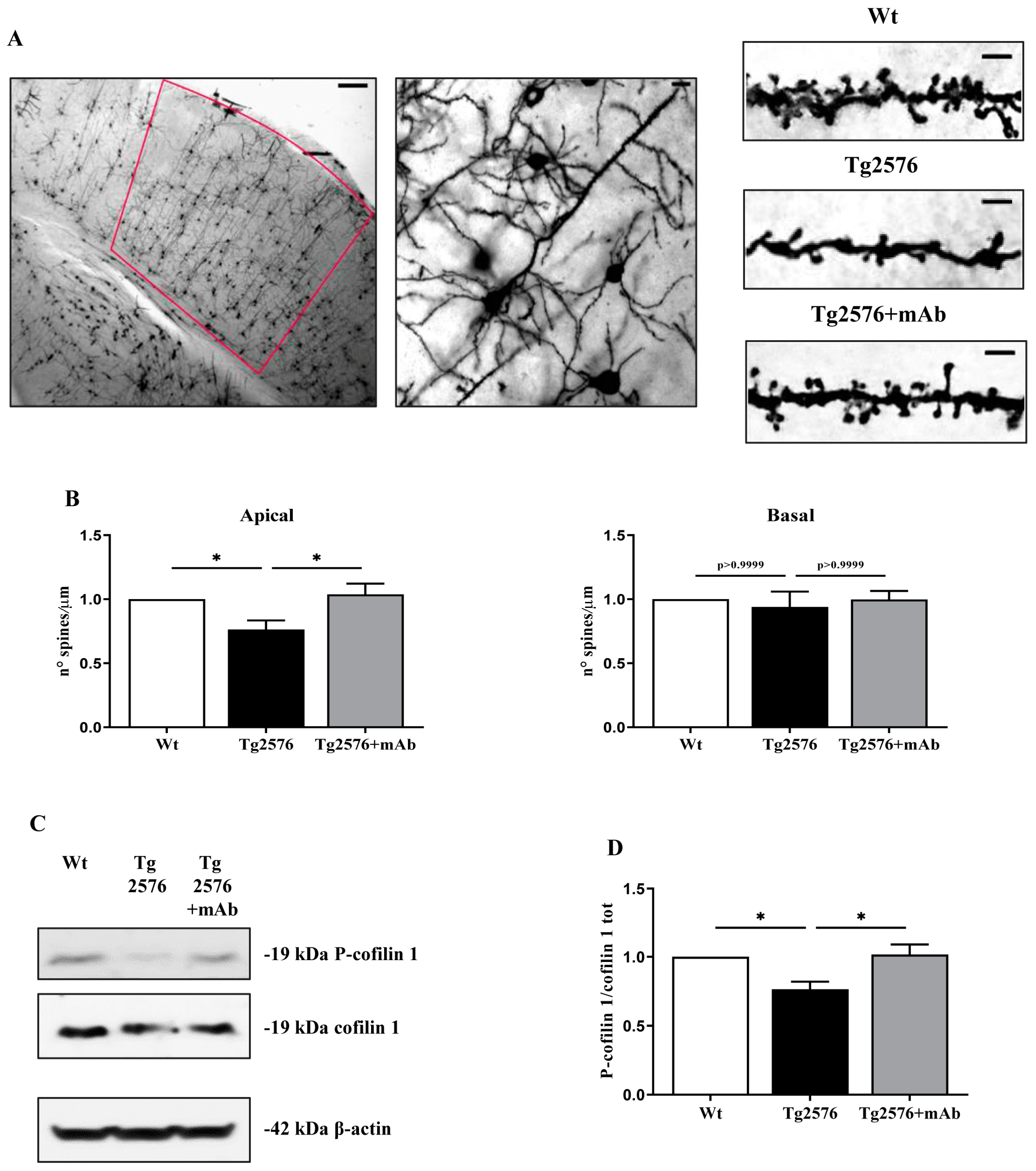
3.4. 12A12mAb Treatment Attenuates the Changes in Dendritic Spine Density and Cofilin Phosphorylation Occurring in the Primary Visual Cortex of Tg2576 AD Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murphy, C. Olfactory and other sensory impairments in Alzheimer disease. Nat. Rev. Neurol. 2019, 15, 11–24. [Google Scholar] [CrossRef]
- Gupta, V.B.; Chitranshi, N.; den Haan, J.; Mirzaei, M.; You, Y.; Lim, J.K.; Basavarajappa, D.; Godinez, A.; Di Angelantonio, S.; Sachdev, P.; et al. Retinal changes in Alzheimer’s disease- integrated prospects of imaging, functional and molecular advances. Prog. Retin. Eye Res. 2021, 82, 100899. [Google Scholar] [CrossRef]
- Mirzaei, N.; Shi, H.; Oviatt, M.; Doustar, J.; Rentsendorj, A.; Fuchs, D.T.; Sheyn, J.; Black, K.L.; Koronyo, Y.; Koronyo-Hamaoui, M. Alzheimer’s Retinopathy: Seeing Disease in the Eyes. Front. Neurosci. 2020, 14, 921. [Google Scholar] [CrossRef]
- Zhang, J.; Shi, L.; Shen, Y. The retina: A window in which to view the pathogenesis of Alzheimer’s disease. Ageing Res. Rev. 2022, 77, 101590. [Google Scholar] [CrossRef]
- Shah, T.M.; Gupta, S.M.; Chatterjee, P.; Campbell, M.; Martins, R.N. Beta-amyloid sequelae in the eye: A critical review on its diagnostic significance and clinical relevance in Alzheimer’s disease. Mol. Psychiatry 2017, 22, 353–363. [Google Scholar] [CrossRef]
- Marchesi, N.; Fahmideh, F.; Boschi, F.; Pascale, A.; Barbieri, A. Ocular Neurodegenerative Diseases: Interconnection between Retina and Cortical Areas. Cells 2021, 10, 2394. [Google Scholar] [CrossRef]
- Guo, L.; Ravindran, N.; Shamsher, E.; Hill, D.; Cordeiro, M.F. Retinal Changes in Transgenic Mouse Models of Alzheimer’s Disease. Curr. Alzheimer Res. 2021, 18, 89–102. [Google Scholar] [CrossRef]
- Chiquita, S.; Rodrigues-Neves, A.C.; Baptista, F.I.; Carecho, R.; Moreira, P.I.; Castelo-Branco, M.; Ambrósio, A.F. The Retina as a Window or Mirror of the Brain Changes Detected in Alzheimer’s Disease: Critical Aspects to Unravel. Mol. Neurobiol. 2019, 56, 5416–5435. [Google Scholar] [CrossRef]
- London, A.; Benhar, I.; Schwartz, M. The retina as a window to the brain: From eye research to CNS disorders. Nat. Rev. Neurol. 2013, 9, 44–53. [Google Scholar] [CrossRef]
- Chiasseu, M.; Alarcon-Martinez, L.; Belforte, N.; Quintero, H.; Dotigny, F.; Destroismaisons, L.; Velde, C.V.; Panayi, F.; Louis, C.; Di Polo, A. Tau accumulation in the retina promotes early neuronal dysfunction and precedes brain pathology in a mouse model of Alzheimer’s disease. Mol. Neurodegener. 2017, 12, 58. [Google Scholar] [CrossRef] [Green Version]
- Criscuolo, C.; Cerri, E.; Fabiani, C.; Capsoni, S.; Cattaneo, A.; Domenici, L. The retina as a window to early dysfunctions of Alzheimer’s disease following studies with a 5xFAD mouse model. Neurobiol. Aging 2018, 67, 181–188. [Google Scholar] [CrossRef]
- Latina, V.; Giacovazzo, G.; Cordella, F.; Balzamino, B.O.; Micera, A.; Varano, M.; Marchetti, C.; Malerba, F.; Florio, R.; Ercole, B.B.; et al. Systemic delivery of a specific antibody targeting the pathological N-terminal truncated tau peptide reduces retinal degeneration in a mouse model of Alzheimer’s Disease. Acta Neuropathol. Commun. 2021, 9, 38. [Google Scholar] [CrossRef]
- Cormack, F.K.; Tovee, M.; Ballard, C. Contrast sensitivity and visual acuity in patients with Alzheimer’s disease. Int. J. Geriatr. Psychiatry 2000, 15, 614–620. [Google Scholar] [CrossRef]
- Albers, M.W.; Gilmore, G.C.; Kaye, J.; Murphy, C.; Wingfield, A.; Bennett, D.A.; Boxer, A.L.; Buchman, A.S.; Cruickshanks, K.J.; Devanand, D.P.; et al. At the interface of sensory and motor dysfunctions and Alzheimer’s disease. Alzheimer’s Dement. 2015, 11, 70–98. [Google Scholar] [CrossRef]
- Armstrong, R.; Kergoat, H. Oculo-visual changes and clinical considerations affecting older patients with dementia. Ophthalmic Physiol. Opt. 2015, 35, 352–376. [Google Scholar] [CrossRef]
- Kusne, Y.; Wolf, A.B.; Townley, K.; Conway, M.; Peyman, G.A. Visual system manifestations of Alzheimer’s disease. Acta Ophthalmol. 2017, 95, e668–e676. [Google Scholar] [CrossRef]
- Glosser, G.; Gallo, J.; Duda, N.; de Vries, J.J.; Clark, C.M.; Grossman, M. Visual perceptual functions predict instrumental activities of daily living in patients with dementia. Neuropsychiatry Neuropsychol. Behav. Neurol. 2002, 15, 198–206. [Google Scholar]
- Rizzo, M.; Anderson, S.W.; Dawson, J.; Nawrot, M. Vision and cognition in Alzheimer’s disease. Neuropsychologia 2000, 38, 1157–1169. [Google Scholar] [CrossRef]
- Tippett, L.J.; Blackwood, K.; Farah, M.J. Visual object and face processing in mild-to-moderate Alzheimer’s disease: From segmentation to imagination. Neuropsychologia 2003, 41, 453–468. [Google Scholar] [CrossRef]
- Ngolab, J.; Honma, P.; Rissman, R.A. Reflections on the Utility of the Retina as a Biomarker for Alzheimer’s Disease: A Literature Review. Neurol. Ther. 2019, 8, 57–72. [Google Scholar] [CrossRef]
- den Haan, J.; Morrema, T.H.J.; Verbraak, F.D.; de Boer, J.F.; Scheltens, P.; Rozemuller, A.J.; Bergen, A.A.B.; Bouwman, F.H.; Hoozemans, J.J. Amyloid-beta and phosphorylated tau in post-mortem Alzheimer’s disease retinas. Acta Neuropathol. Commun. 2018, 6, 147. [Google Scholar] [CrossRef]
- Blazes, M.; Lee, C.S. Understanding the Brain through Aging Eyes. Adv. Geriatr. Med. Res. 2021, 3, e210008. [Google Scholar] [CrossRef]
- Hart, N.J.; Koronyo, Y.; Black, K.L.; Koronyo-Hamaoui, M. Ocular indicators of Alzheimer’s: Exploring disease in the retina. Acta Neuropathol. 2016, 132, 767–787. [Google Scholar] [CrossRef]
- Majeed, A.; Marwick, B.; Yu, H.; Fadavi, H.; Tavakoli, M. Ophthalmic Biomarkers for Alzheimer’s Disease: A Review. Front. Aging Neurosci. 2021, 13, 720167. [Google Scholar] [CrossRef]
- Ge, Y.J.; Xu, W.; Ou, Y.N.; Qu, Y.; Ma, Y.H.; Huang, Y.Y.; Shen, X.N.; Chen, S.D.; Tan, L.; Zhao, Q.H.; et al. Retinal biomarkers in Alzheimer’s disease and mild cognitive impairment: A systematic review and meta-analysis. Ageing Res. Rev. 2021, 69, 101361. [Google Scholar] [CrossRef]
- Liao, C.; Xu, J.; Chen, Y.; Ip, N.Y. Retinal Dysfunction in Alzheimer’s Disease and Implications for Biomarkers. Biomolecules 2021, 11, 1215. [Google Scholar] [CrossRef]
- Binetti, G.; Cappa, S.F.; Magni, E.; Padovani, A.; Bianchetti, A.; Trabucchi, M. Visual and spatial perception in the early phase of Alzheimer’s disease. Neuropsychology 1998, 12, 29–33. [Google Scholar] [CrossRef]
- Alescio-Lautier, B.; Michel, B.F.; Herrera, C.; Elahmadi, A.; Chambon, C.; Touzet, C.; Paban, V. Visual and visuospatial short-term memory in mild cognitive impairment and Alzheimer disease: Role of attention. Neuropsychologia 2007, 45, 1948–1960. [Google Scholar] [CrossRef]
- Tales, A.; Haworth, J.; Nelson, S.; Snowden, R.J.; Wilcock, G. Abnormal visual search in mild cognitive impairment and Alzheimer’s disease. Neurocase 2005, 11, 80–84. [Google Scholar] [CrossRef]
- Lemos, R.; Santana, I.; Caetano, G.; Bernardino, I.; Morais, R.; Farivar, R.; Castelo-Branco, M. Three-Dimensional Face Recognition in Mild Cognitive Impairment: A Psychophysical and Structural MR Study. J. Int. Neuropsychol. Soc. 2016, 22, 744–754. [Google Scholar] [CrossRef]
- Graewe, B.; Lemos, R.; Ferreira, C.; Santana, I.; Farivar, R.; De Weerd, P.; Castelo-Branco, M. Impaired processing of 3D motion-defined faces in mild cognitive impairment and healthy aging: An fMRI study. Cereb. Cortex 2013, 23, 2489–2499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marquié, M.; Castilla-Martí, M.; Valero, S.; Martínez, J.; Sánchez, D.; Hernández, I.; Rosende-Roca, M.; Vargas, L.; Mauleón, A.; Rodríguez-Gómez, O.; et al. Visual impairment in aging and cognitive decline: Experience in a Memory Clinic. Sci. Rep. 2019, 9, 8698. [Google Scholar] [CrossRef] [PubMed]
- Tong, F. Primary visual cortex and visual awareness. Nat. Rev. Neurosci. 2003, 4, 219–229. [Google Scholar] [CrossRef]
- Ikonomovic, M.D.; Mufson, E.J.; Wuu, J.; Bennett, D.A.; DeKosky, S.T. Reduction of choline acetyltransferase activity in primary visual cortex in mild to moderate Alzheimer’s disease. Arch. Neurol. 2005, 62, 425–430. [Google Scholar] [CrossRef]
- Javaid, F.Z.; Brenton, J.; Guo, L.; Cordeiro, M.F. Visual and Ocular Manifestations of Alzheimer’s Disease and Their Use as Biomarkers for Diagnosis and Progression. Front. Neurol. 2016, 7, 55. [Google Scholar] [CrossRef]
- Gămănuţ, R.; Shimaoka, D. Anatomical and functional connectomes underlying hierarchical visual processing in mouse visual system. Brain Struct. Funct. 2021, 227, 1297–1315. [Google Scholar] [CrossRef]
- Miller, M.W.; Vogt, B.A. Direct connections of rat visual cortex with sensory, motor, and association cortices. J. Comp. Neurol. 1984, 226, 184–202. [Google Scholar] [CrossRef]
- Vaudano, E.; Legg, C.R.; Glickstein, M. Afferent and Efferent Connections of Temporal Association Cortex in the Rat: A Horseradish Peroxidase Study. Eur. J. Neurosci. 1991, 3, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Lavenex, P.; Amaral, D.G. Hippocampal-neocortical interaction: A hierarchy of associativity. Hippocampus 2000, 10, 420–430. [Google Scholar] [CrossRef]
- Furtak, S.C.; Wei, S.M.; Agster, K.L.; Burwell, R.D. Functional neuroanatomy of the parahippocampal region in the rat: The perirhinal and postrhinal cortices. Hippocampus 2007, 17, 709–722. [Google Scholar] [CrossRef]
- Haggerty, D.C.; Ji, D. Activities of visual cortical and hippocampal neurons co-fluctuate in freely moving rats during spatial behavior. Elife 2015, 4, e08902. [Google Scholar] [CrossRef] [PubMed]
- Corsetti, V.; Borreca, A.; Latina, V.; Giacovazzo, G.; Pignataro, A.; Krashia, P.; Natale, F.; Cocco, S.; Rinaudo, M.; Malerba, F.; et al. Passive immunotherapy for N-truncated tau ameliorates the cognitive deficits in two mouse Alzheimer’s disease models. Brain Commun. 2020, 2, fcaa039. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, K.; Chapman, P.; Nilsen, S.; Eckman, C.; Harigaya, Y.; Younkin, S.; Yang, F.; Cole, G. Correlative memory deficits, Abeta elevation, and amyloid plaques in transgenic mice. Science 1996, 274, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Sasaguri, H.; Nilsson, P.; Hashimoto, S.; Nagata, K.; Saito, T.; De Strooper, B.; Hardy, J.; Vassar, R.; Winblad, B.; Saido, T.C. APP mouse models for Alzheimer’s disease preclinical studies. EMBO J. 2017, 36, 2473–2487. [Google Scholar] [CrossRef]
- Yassine, N.; Lazaris, A.; Dorner-Ciossek, C.; Després, O.; Meyer, L.; Maitre, M.; Mensah-Nyagan, A.G.; Cassel, J.C.; Mathis, C. Detecting spatial memory deficits beyond blindness in tg2576 Alzheimer mice. Neurobiol. Aging 2013, 34, 716–730. [Google Scholar] [CrossRef]
- Castillo-Carranza, D.L.; Guerrero-Muñoz, M.J.; Sengupta, U.; Hernandez, C.; Barrett, A.D.; Dineley, K.; Kayed, R. Tau immunotherapy modulates both pathological tau and upstream amyloid pathology in an Alzheimer’s disease mouse model. J. Neurosci. 2015, 35, 4857–4868. [Google Scholar] [CrossRef]
- Schmolesky, M.; Kolb, H.; Fernandez, E.; Nelson, R. The Primary Visual Cortex. In Webvision: The Organization of the Retina and Visual System [Internet]; University of Utah Health Sciences Center: Salt Lake City, UT, USA, 1995; Bookshelf ID: NBK11524. Available online: https://www.ncbi.nlm.nih.gov/books/NBK11524 (accessed on 1 May 2005).
- Paxinos, G.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates, 2nd ed.; Academic Press: New York, NY, USA, 2001; ISBN 0-12-547637-X. [Google Scholar]
- Gao, M.; Sossa, K.; Song, L.; Errington, L.; Cummings, L.; Hwang, H.; Kuhl, D.; Worley, P.; Lee, H.K. A specific requirement of Arc/Arg3.1 for visual experience-induced homeostatic synaptic plasticity in mouse primary visual cortex. J. Neurosci. 2010, 30, 7168–7178. [Google Scholar] [CrossRef]
- El-Boustani, S.; Ip, J.P.K.; Breton-Provencher, V.; Knott, G.W.; Okuno, H.; Bito, H.; Sur, M. Locally coordinated synaptic plasticity of visual cortex neurons in vivo. Science 2018, 360, 1349–1354. [Google Scholar] [CrossRef]
- D’Amelio, M.; Cavallucci, V.; Middei, S.; Marchetti, C.; Pacioni, S.; Ferri, A.; Diamantini, A.; De Zio, D.; Carrara, P.; Battistini, L.; et al. Caspase-3 triggers early synaptic dysfunction in a mouse model of Alzheimer’s disease. Nat. Neurosci. 2011, 14, 69–76. [Google Scholar] [CrossRef]
- Schägger, H.; von Jagow, G. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal. Biochem. 1987, 166, 368–379. [Google Scholar] [CrossRef]
- Amadoro, G.; Corsetti, V.; Atlante, A.; Florenzano, F.; Capsoni, S.; Bussani, R.; Mercanti, D.; Calissano, P. Interaction between NH(2)-tau fragment and Aβ in Alzheimer’s disease mitochondria contributes to the synaptic deterioration. Neurobiol. Aging 2012, 33, 833.e1–833.e25. [Google Scholar] [CrossRef] [PubMed]
- Corsetti, V.; Amadoro, G.; Gentile, A.; Capsoni, S.; Ciotti, M.T.; Cencioni, M.T.; Atlante, A.; Canu, N.; Rohn, T.T.; Cattaneo, A.; et al. Identification of a caspase-derived N-terminal tau fragment in cellular and animal Alzheimer’s disease models. Mol. Cell. Neurosci. 2008, 38, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Rohn, T.T.; Rissman, R.A.; Davis, M.C.; Kim, Y.E.; Cotman, C.W.; Head, E. Caspase-9 activation and caspase cleavage of tau in the Alzheimer’s disease brain. Neurobiol. Dis. 2002, 11, 341–354. [Google Scholar] [CrossRef]
- Pignataro, A.; Meli, G.; Pagano, R.; Fontebasso, V.; Battistella, R.; Conforto, G.; Ammassari-Teule, M.; Middei, S. Activity-Induced Amyloid-β Oligomers Drive Compensatory Synaptic Rearrangements in Brain Circuits Controlling Memory of Presymptomatic Alzheimer’s Disease Mice. Biol. Psychiatry 2019, 86, 185–195. [Google Scholar] [CrossRef]
- Rosoklija, G.B.; Petrushevski, V.M.; Stankov, A.; Dika, A.; Jakovski, Z.; Pavlovski, G.; Davcheva, N.; Lipkin, R.; Schnieder, T.; Scobie, K.; et al. Reliable and durable Golgi staining of brain tissue from human autopsies and experimental animals. J. Neurosci. Methods 2014, 230, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Risher, W.C.; Ustunkaya, T.; Alvarado, J.S.; Eroglu, C. Rapid Golgi analysis method for efficient and unbiased classification of dendritic spines. PLoS ONE 2014, 9, e107591. [Google Scholar] [CrossRef]
- Gibb, R.; Kolb, B. A method for vibratome sectioning of Golgi-Cox stained whole rat brain. J. Neurosci. Methods 1998, 79, 1–4. [Google Scholar] [CrossRef]
- Pignataro, A.; Borreca, A.; Ammassari-Teule, M.; Middei, S. CREB Regulates Experience-Dependent Spine Formation and Enlargement in Mouse Barrel Cortex. Neural Plast. 2015, 2015, 651469. [Google Scholar] [CrossRef]
- Prusky, G.T.; West, P.W.; Douglas, R.M. Behavioral assessment of visual acuity in mice and rats. Vis. Res. 2000, 40, 2201–2209. [Google Scholar] [CrossRef]
- Robinson, L.; Bridge, H.; Riedel, G. Visual discrimination learning in the water maze: A novel test for visual acuity. Behav. Brain Res. 2001, 119, 77–84. [Google Scholar] [CrossRef]
- Robinson, L.; Harbaran, D.; Riedel, G. Visual acuity in the water maze: Sensitivity to muscarinic receptor blockade in rats and mice. Behav. Brain Res. 2004, 151, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Marrocco, E.; Indrieri, A.; Esposito, F.; Tarallo, V.; Carboncino, A.; Alvino, F.G.; De Falco, S.; Franco, B.; De Risi, M.; De Leonibus, E. α-synuclein overexpression in the retina leads to vision impairment and degeneration of dopaminergic amacrine cells. Sci. Rep. 2020, 10, 9619. [Google Scholar] [CrossRef] [PubMed]
- Crijns, E.; Op de Beeck, H. The Visual Acuity of Rats in Touchscreen Setups. Vision 2019, 4, 4. [Google Scholar] [CrossRef] [PubMed]
- Himmelhan, D.K.; Rawashdeh, O.; Oelschläger, H.H.A. Early postnatal development of the visual cortex in mice with retinal degeneration. Mech. Dev. 2018, 151, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Palomero-Gallagher, N.; Zilles, K.; Paxinos, I.G. (Eds.) The Rat Nervous System; Elsevier-Academic Press: Amsterdam, The Netherlands, 2004; pp. 729–757, Chapter 23. [Google Scholar]
- Paxinos, G.; Franklin, K.B.J. Paxinos and Franklin’s the Mouse Brain in Stereotaxic Coordinates, 4th ed.; Academic Press: Cambridge, MA, USA, 2012. [Google Scholar]
- Allen Institute for Brain Science. Allen’s Mouse Brain Atlas—The Brain Explorer® 2; Allen Institute for Brain Science: Seattle, WA, USA, 2014; Online Program Version. [Google Scholar]
- Shihan, M.H.; Novo, S.G.; Le Marchand, S.J.; Wang, Y.; Duncan, M.K. A simple method for quantitating confocal fluorescent images. Biochem. Biophys. Rep. 2021, 25, 100916. [Google Scholar] [CrossRef]
- Morrissette, D.A.; Parachikova, A.; Green, K.N.; LaFerla, F.M. Relevance of transgenic mouse models to human Alzheimer disease. J. Biol. Chem. 2009, 284, 6033–6037. [Google Scholar] [CrossRef]
- Priebe, N.J.; McGee, A.W. Mouse vision as a gateway for understanding how experience shapes neural circuits. Front. Neural Circuits 2014, 8, 123. [Google Scholar] [CrossRef]
- Niell, C.M.; Stryker, M.P. Highly selective receptive fields in mouse visual cortex. J. Neurosci. 2008, 28, 7520–7536. [Google Scholar] [CrossRef]
- Iacaruso, M.F.; Gasler, I.T.; Hofer, S.B. Synaptic organization of visual space in primary visual cortex. Nature 2017, 547, 449–452. [Google Scholar] [CrossRef] [PubMed]
- Asavapanumas, N.; Brawek, B.; Martus, P.; Garaschuk, O. Role of intracellular Ca2+ stores for an impairment of visual processing in a mouse model of Alzheimer’s disease. Neurobiol. Dis. 2019, 121, 315–326. [Google Scholar] [CrossRef]
- Jorge, L.; Canário, N.; Martins, R.; Santiago, B.; Santana, I.; Quental, H.; Ambrósio, F.; Bernardes, R.; Castelo-Branco, M. The Retinal Inner Plexiform Synaptic Layer Mirrors Grey Matter Thickness of Primary Visual Cortex with Increased Amyloid β Load in Early Alzheimer’s Disease. Neural Plast. 2020, 2020, 8826087. [Google Scholar] [CrossRef]
- Jorge, L.; Canário, N.; Quental, H.; Bernardes, R.; Castelo-Branco, M. Is the Retina a Mirror of the Aging Brain? Aging of Neural Retina Layers and Primary Visual Cortex Across the Lifespan. Front. Aging Neurosci. 2020, 11, 360. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, L.; Joly, S.; Zine-Eddine, F.; Mdzomba, J.B.; Pernet, V. Tau modulates visual plasticity in adult and old mice. Neurobiol. Aging 2020, 95, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Leuba, G.; Kraftsik, R. Visual cortex in Alzheimer’s disease: Occurrence of neuronal death and glial proliferation, and correlation with pathological hallmarks. Neurobiol. Aging 1994, 15, 29–43. [Google Scholar] [CrossRef] [PubMed]
- Ho, W.; Leung, Y.; Tsang, A.W.; So, K.F.; Chiu, K.; Chang, R.C. Review: Tauopathy in the retina and optic nerve: Does it shadow pathological changes in the brain? Mol. Vis. 2012, 18, 2700–2710. [Google Scholar] [PubMed]
- Rahimi, J.; Milenkovic, I.; Kovacs, G.G. Patterns of Tau and α-Synuclein Pathology in the Visual System. J. Park. Dis. 2015, 5, 333–340. [Google Scholar] [CrossRef]
- Arouche-Delaperche, L.; Cadoni, S.; Joffrois, C.; Labernede, G.; Valet, M.; César, Q.; Dégardin, J.; Girardon, S.; Gabriel, C.; Krantic, S.; et al. Dysfunction of the glutamatergic photoreceptor synapse in the P301S mouse model of tauopathy. Acta Neuropathol Commun. 2023, 11, 5. [Google Scholar] [CrossRef]
- Cerquera-Jaramillo, M.A.; Nava-Mesa, M.O.; González-Reyes, R.E.; Tellez-Conti, C.; de-la-Torre, A. Visual Features in Alzheimer’s Disease: From Basic Mechanisms to Clinical Overview. Neural Plast. 2018, 14, 2941783. [Google Scholar] [CrossRef]
- Paik, J.S.; Ha, M.; Jung, Y.H.; Kim, G.H.; Han, K.D.; Kim, H.S.; Lim, D.H.; Na, K.S. Low vision and the risk of dementia: A nationwide population-based cohort study. Sci. Rep. 2020, 10, 9109. [Google Scholar] [CrossRef]
- Elyashiv, S.M.; Shabtai, E.L.; Belkin, M. Correlation between visual acuity and cognitive functions. Br. J. Ophthalmol. 2014, 98, 129–132. [Google Scholar] [CrossRef]
- Liebscher, S.; Keller, G.B.; Goltstein, P.M.; Bonhoeffer, T.; Hübener, M. Selective Persistence of Sensorimotor Mismatch Signals in Visual Cortex of Behaving Alzheimer’s Disease mice. Curr. Biol. 2016, 26, 956–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grienberger, C.; Rochefort, N.L.; Adelsberger, H.; Henning, H.A.; Hill, D.N.; Reichwald, J.; Staufenbiel, M.; Konnerth, A. Staged decline of neuronal function in vivo in an animal model of Alzheimer’s disease. Nat. Commun. 2012, 3, 774. [Google Scholar] [CrossRef]
- Papanikolaou, A.; Rodrigues, F.R.; Holeniewska, J.; Phillips, K.G.; Saleem, A.B.; Solomon, S.G. Plasticity in visual cortex is disrupted in a mouse model of tauopathy. Commun. Biol. 2022, 5, 77. [Google Scholar] [CrossRef]
- Stover, K.R.; Brown, R.E. Age-related changes in visual acuity, learning and memory in the APPswe/PS1dE9 mouse model of Alzheimer’s disease. Behav. Brain Res. 2012, 231, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Vit, J.P.; Fuchs, D.T.; Angel, A.; Levy, A.; Lamensdorf, I.; Black, K.L.; Koronyo, Y.; Koronyo-Hamaoui, M. Color and contrast vision in mouse models of aging and Alzheimer’s disease using a novel visual-stimuli four-arm maze. Sci. Rep. 2021, 11, 1255. [Google Scholar] [CrossRef] [PubMed]
- Turner, K.M.; Simpson, C.G.; Burne, T.H.J. Touchscreen-based Visual Discrimination and Reversal Tasks for Mice to Test Cognitive Flexibility. Bio-Protocol 2017, 7, e2583. [Google Scholar] [CrossRef]
- Storchi, R.; Rodgers, J.; Gracey, M.; Martial, F.P.; Wynne, J.; Ryan, S.; Twining, C.J.; Cootes, T.F.; Killick, R.; Lucas, R.J. Measuring vision using innate behaviours in mice with intact and impaired retina function. Sci. Rep. 2019, 9, 10396. [Google Scholar] [CrossRef]
- Nikolaienko, O.; Patil, S.; Eriksen, M.S.; Bramham, C.R. Arc protein: A flexible hub for synaptic plasticity and cognition. Semin. Cell Dev. Biol. 2018, 77, 33–42. [Google Scholar] [CrossRef]
- Tagawa, Y.; Kanold, P.O.; Majdan, M.; Shatz, C.J. Multiple periods of functional ocular dominance plasticity in mouse visual cortex. Nat. Neurosci. 2005, 8, 380–388. [Google Scholar] [CrossRef]
- Jenks, K.R.; Kim, T.; Pastuzyn, E.D.; Okuno, H.; Taibi, A.V.; Bito, H.; Bear, M.F.; Shepherd, J.D. Arc restores juvenile plasticity in adult mouse visual cortex. Proc. Natl. Acad. Sci. USA 2017, 114, 9182–9187. [Google Scholar] [CrossRef]
- William, C.M.; Stern, M.A.; Pei, X.; Saqran, L.; Ramani, M.; Frosch, M.P.; Hyman, B.T. Impairment of visual cortical plasticity by amyloid-beta species. Neurobiol. Dis. 2021, 154, 105344. [Google Scholar] [CrossRef] [PubMed]
- Rudinskiy, N.; Hawkes, J.M.; Wegmann, S.; Kuchibhotla, K.V.; Muzikansky, A.; Betensky, R.A.; Spires-Jones, T.L.; Hyman, B.T. Tau pathology does not affect experience-driven single-neuron and network-wide Arc/Arg3.1 responses. Acta Neuropathol. Commun. 2014, 2, 63. [Google Scholar] [CrossRef]
- Rudinskiy, N.; Hawkes, J.M.; Betensky, R.A.; Eguchi, M.; Yamaguchi, S.; Spires-Jones, T.L.; Hyman, B.T. Orchestrated experience-driven Arc responses are disrupted in a mouse model of Alzheimer’s disease. Nat. Neurosci. 2012, 15, 1422–1429. [Google Scholar] [CrossRef] [PubMed]
- Dickey, C.A.; Loring, J.F.; Montgomery, J.; Gordon, M.N.; Eastman, P.S.; Morgan, D. Selectively reduced expression of synaptic plasticity-related genes in amyloid precursor protein + presenilin-1 transgenic mice. J. Neurosci. 2003, 23, 5219–5226. [Google Scholar] [CrossRef] [PubMed]
- Dickey, C.A.; Gordon, M.N.; Mason, J.E.; Wilson, N.J.; Diamond, D.M.; Guzowski, J.F.; Morgan, D. Amyloid suppresses induction of genes critical for memory consolidation in APP + PS1 transgenic mice. J. Neurochem. 2004, 88, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Di Cristo, G.; Berardi, N.; Cancedda, L.; Pizzorusso, T.; Putignano, E.; Ratto, G.M.; Maffei, L. Requirement of ERK activation for visual cortical plasticity. Science 2001, 292, 2337–2340. [Google Scholar] [CrossRef]
- Caleo, M.; Restani, L.; Gianfranceschi, L.; Costantin, L.; Rossi, C.; Rossetto, O.; Montecucco, C.; Maffei, L. Transient synaptic silencing of developing striate cortex has persistent effects on visual function and plasticity. J. Neurosci. 2007, 27, 4530–4540. [Google Scholar] [CrossRef] [PubMed]
- Fox, L.M.; William, C.M.; Adamowicz, D.H.; Pitstick, R.; Carlson, G.A.; Spires-Jones, T.L.; Hyman, B.T. Soluble tau species, not neurofibrillary aggregates, disrupt neural system integration in a tau transgenic model. J. Neuropathol. Exp. Neurol. 2011, 70, 588–595. [Google Scholar] [CrossRef]
- Morin, J.P.; Cerón-Solano, G.; Velázquez-Campos, G.; Pacheco-López, G.; Bermúdez-Rattoni, F.; Díaz-Cintra, S. Spatial Memory Impairment is Associated with Intraneural Amyloid-β Immunoreactivity and Dysfunctional Arc Expression in the Hippocampal-CA3 Region of a Transgenic Mouse Model of Alzheimer’s Disease. J. Alzheimer’s Dis. 2016, 51, 69–79. [Google Scholar] [CrossRef]
- Wang, K.H.; Majewska, A.; Schummers, J.; Farley, B.; Hu, C.; Sur, M.; Tonegawa, S. In vivo two-photon imaging reveals a role of arc in enhancing orientation specificity in visual cortex. Cell 2006, 126, 389–402. [Google Scholar] [CrossRef]
- Ramirez-Amaya, V.; Vazdarjanova, A.; Mikhael, D.; Rosi, S.; Worley, P.F.; Barnes, C.A. Spatial exploration-induced Arc mRNA and protein expression: Evidence for selective, network-specific reactivation. J. Neurosci. 2005, 25, 1761–1768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lind, D.; Franken, S.; Kappler, J.; Jankowski, J.; Schilling, K. Characterization of the neuronal marker NeuN as a multiply phosphorylated antigen with discrete subcellular localization. J. Neurosci. Res. 2005, 79, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Wallace, W.; Bear, M.F. A morphological correlate of synaptic scaling in visual cortex. J. Neurosci. 2004, 24, 6928–6938. [Google Scholar] [CrossRef] [PubMed]
- Tropea, D.; Majewska, A.K.; Garcia, R.; Sur, M. Structural dynamics of synapses in vivo correlate with functional changes during experience-dependent plasticity in visual cortex. J. Neurosci. 2010, 30, 11086–11095. [Google Scholar] [CrossRef]
- Chen, J.L.; Villa, K.L.; Cha, J.W.; So, P.T.C.; Kubota, Y.; Nedivi, E. Clustered Dynamics of Inhibitory Synapses and Dendritic Spines in the Adult Neocortex. Neuron 2012, 74, 361–373. [Google Scholar] [CrossRef]
- Yu, H.; Majewska, A.K.; Sur, M. Rapid experience-dependent plasticity of synapse function and structure in ferret visual cortex in vivo. Proc. Natl. Acad. Sci. USA 2011, 108, 21235–21240. [Google Scholar] [CrossRef]
- Zablah, B.Y.; Merovitch, N.; Jia, Z. The Role of ADF/Cofilin in Synaptic Physiology and Alzheimer’s Disease. Front. Cell Dev. Biol. 2020, 8, 594998. [Google Scholar] [CrossRef]
- Renouard, L.; Bridi, M.C.D.; Coleman, T.; Arckens, L.; Frank, M.G. Anatomical correlates of rapid eye movement sleep-dependent plasticity in the developing cortex. Sleep 2018, 41, zsy124. [Google Scholar] [CrossRef]
- Dahlhaus, M.; Li, K.W.; Van Der Schors, R.C.; Saiepour, M.H.; Van Nierop, P.; Heimel, J.A.; Hermans, J.M.; Loos, M.; Smit, A.B.; Levelt, C.N. The synaptic proteome during development and plasticity of the mouse visual cortex. Mol. Cell. Proteom. 2011, 10, M110.005413. [Google Scholar] [CrossRef]
- Bornia, N.; Taboada, A.; Dapueto, A.; Rossi, F.M. Identification of cofilin 1 as a candidate protein associated to mouse visual cortex plasticity. Neurosci. Lett. 2020, 731, 135056. [Google Scholar] [CrossRef]
- Levine, N.D.; Rademacher, D.J.; Collier, T.J.; O’Malley, J.A.; Kells, A.P.; San Sebastian, W.; Bankiewicz, K.S.; Steece-Collier, K. Advances in Thin Tissue Golgi-Cox Impregnation: Fast, Reliable Methods for Multi-Assay Analyses in Rodent and Non-human Primate Brain. J. Neurosci. Methods 2013, 213, 214–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanz, T.A.; Carter, D.B.; Merchant, K.M. Dendritic spine loss in the hippocampus of young PDAPP and Tg2576 mice and its prevention by the ApoE2 genotype. Neurobiol. Dis. 2003, 13, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, J.S.; Wu, C.C.; Redwine, J.M.; Comery, T.A.; Arias, R.; Bowlby, M.; Martone, R.; Morrison, J.H.; Pangalos, M.N.; Reinhart, P.H.; et al. Early-onset behavioral and synaptic deficits in a mouse model of Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2006, 103, 5161–5166. [Google Scholar] [CrossRef] [PubMed]
- Rahman, T.; Davies, D.S.; Tannenberg, R.K.; Fok, S.; Shepherd, C.; Dodd, P.R.; Cullen, K.M.; Goldsbury, C. Cofilin rods and aggregates concur with tau pathology and the development of Alzheimer’s disease. J. Alzheimer’s Dis. 2014, 42, 1443–1460. [Google Scholar] [CrossRef] [PubMed]
- Bamburg, J.R.; Bernstein, B.W. Actin dynamics and cofilin-actin rods in alzheimer disease. Cytoskeleton 2016, 73, 477–497. [Google Scholar] [CrossRef]
- Kang, D.E.; Woo, J.A. Cofilin, a Master Node Regulating Cytoskeletal Pathogenesis in Alzheimer’s Disease. J. Alzheimer’s Dis. 2019, 72, S131–S144. [Google Scholar] [CrossRef]
- Davis, R.C.; Marsden, I.T.; Maloney, M.T.; Minamide, L.S.; Podlisny, M.; Selkoe, D.J.; Bamburg, J.R. Amyloid beta dimers/trimers potently induce cofilin-actin rods that are inhibited by maintaining cofilin-phosphorylation. Mol. Neurodegener. 2011, 6, 10. [Google Scholar] [CrossRef]
- Mendoza-Naranjo, A.; Contreras-Vallejos, E.; Henriquez, D.R.; Otth, C.; Bamburg, J.R.; Maccioni, R.B.; Gonzalez-Billault, C. Fibrillar amyloid-β1–42 modifies actin organization affecting the cofilin phosphorylation state: A role for Rac1/cdc42 effector proteins and the slingshot phosphatase. J. Alzheimer’s Dis. 2012, 29, 63–77. [Google Scholar] [CrossRef]
- Barone, E.; Mosser, S.; Fraering, P.C. Inactivation of brain Cofilin-1 by age, Alzheimer’s disease and γ-secretase. Biochim. Biophys. Acta 2014, 1842, 2500–2509. [Google Scholar] [CrossRef]
- Norton, D.J.; Parra, M.A.; Sperling, R.A.; Baena, A.; Guzman-Velez, E.; Jin, D.S.; Andrea, N.; Khang, J.; Schultz, A.; Rentz, D.M.; et al. Visual short-term memory relates to tau and amyloid burdens in preclinical autosomal dominant Alzheimer’s disease. Alzheimer’s Res. Ther. 2020, 12, 99. [Google Scholar] [CrossRef]
- Bocanegra, Y.; Fox-Fuller, J.T.; Baena, A.; Guzmán-Vélez, E.; Vila-Castelar, C.; Martínez, J.; Torrico-Teave, H.; Lopera, F.; Quiroz, Y.T. Association Between Visual Memory and In Vivo Amyloid and Tau Pathology in Preclinical Autosomal Dominant Alzheimer’s Disease. J. Int. Neuropsychol. Soc. 2021, 27, 47–55. [Google Scholar] [CrossRef]
- Putcha, D.; Brickhouse, M.; Touroutoglou, A.; Collins, J.A.; Quimby, M.; Wong, B.; Eldaief, M.; Schultz, A.; El Fakhri, G.; Johnson, K.; et al. Visual cognition in non-amnestic Alzheimer’s disease: Relations to tau, amyloid, and cortical atrophy. Neuroimage Clin. 2019, 23, 101889. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, G.C.; Cronin-Golomb, A.; Neargarder, S.A.; Morrison, S.R. Enhanced stimulus contrast normalizes visual processing of rapidly presented letters in Alzheimer’s disease. Vis. Res. 2005, 45, 1013–1020. [Google Scholar] [CrossRef] [PubMed]
- Kavcic, V.; Duffy, C.J. Attentional dynamics and visual perception: Mechanisms of spatial disorientation in Alzheimer’s disease. Brain 2003, 126, 1173–1181. [Google Scholar] [CrossRef]
- Schlotterer, G.; Moscovitch, M.; Crapper-McLachlan, D. Visual processing deficits as assessed by spatial frequency contrast sensitivity and backward masking in normal ageing and Alzheimer’s disease. Brain 1984, 107, 309–325. [Google Scholar] [CrossRef] [PubMed]
- Bonney, K.R.; Almeida, O.P.; Flicker, L.; Davies, S.; Clarnette, R.; Anderson, M.; Lautenschlager, N.T. Inspection time in non-demented older adults with mild cognitive impairment. Neuropsychologia 2006, 44, 1452–1456. [Google Scholar] [CrossRef] [PubMed]
- Perry, R.J.; Hodges, J.R. Dissociation between top-down attentional control and the time course of visual attention as measured by attentional dwell time in patients with mild cognitive impairment. Eur. J. Neurosci. 2003, 18, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Cunha, J.P.; Moura-Coelho, N.; Proença, R.P.; Dias-Santos, A.; Ferreira, J.; Louro, C.; Castanheira-Dinis, A. Alzheimer’s disease: A review of its visual system neuropathology. Optical coherence tomography—A potential role as a study tool in vivo. Graefes Arch. Clin. Exp. Ophthalmol. 2016, 254, 2079–2092. [Google Scholar] [CrossRef] [PubMed]
- Kirby, E.; Bandelow, S.; Hogervorst, E. Visual impairment in Alzheimer’s disease: A critical review. J. Alzheimer’s Dis. 2010, 21, 15–34. [Google Scholar] [CrossRef]
- Chang, L.Y.L.; Lowe, J.; Ardiles, A.; Lim, J.; Grey, A.C.; Robertson, K.; Danesh-Meyer, H.; Palacios, A.G.; Acosta, M.L. Alzheimer’s disease in the human eye. Clinical tests that identify ocular and visual information processing deficit as biomarkers. Alzheimers Dement. 2014, 10, 251–261. [Google Scholar] [CrossRef]
- Huang, J.; Beach, P.; Bozoki, A.; Zhu, D.C. Alzheimer’s Disease Progressively Reduces Visual Functional Network Connectivity. J. Alzheimer’s Dis. Rep. 2021, 5, 549–562. [Google Scholar] [CrossRef]
- Huang, J.; Beach, P.; Bozoki, A.; Zhu, D.C. Alzheimer’s Disease Progressively Alters the Face-Evoked Visual-Processing Network. J. Alzheimer’s Dis. 2020, 77, 1025–1042. [Google Scholar] [CrossRef] [PubMed]
- Rehan, S.; Giroud, N.; Al-Yawer, F.; Wittich, W.; Phillips, N. Visual Performance and Cortical Atrophy in Vision-Related Brain Regions Differ Between Older Adults with (or at Risk for) Alzheimer’s Disease. J. Alzheimer’s Dis. 2021, 83, 1125–1148. [Google Scholar] [CrossRef]
- Chen, C.; Ma, X.; Wei, J.; Shakir, N.; Zhang, J.K.; Zhang, L.; Nehme, A.; Cui, Y.; Ferguson, D.; Bai, F.; et al. Early impairment of cortical circuit plasticity and connectivity in the 5XFAD Alzheimer’s disease mouse model. Transl. Psychiatry 2022, 12, 371. [Google Scholar] [CrossRef]
- O’Leary, T.P.; Brown, R.E. Visuo-spatial learning and memory deficits on the Barnes maze in the 16-month-old APPswe/PS1dE9 mouse model of Alzheimer’s disease. Behav. Brain Res. 2009, 201, 120–127. [Google Scholar] [CrossRef]
- Wong, A.A.; Brown, R.E. Visual detection, pattern discrimination and visual acuity in 14 strains of mice. Genes Brain Behav. 2006, 5, 389–403. [Google Scholar] [CrossRef]
- Brown, R.E.; Wong, A.A. The influence of visual ability on learning and memory performance in 13 strains of mice. Learn. Mem. 2007, 14, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Buscher, N.; van Dorsselaer, P.; Steckler, T.; Talpos, J.C. Evaluating aged mice in three touchscreen tests that differ in visual demands: Impaired cognitive function and impaired visual abilities. Behav. Brain Res. 2017, 333, 142–149. [Google Scholar] [CrossRef]
- Poort, J.; Khan, A.G.; Pachitariu, M.; Nemri, A.; Orsolic, I.; Krupic, J.; Bauza, M.; Sahani, M.; Keller, G.B.; Mrsic-Flogel, T.D.; et al. Learning Enhances Sensory and Multiple Non-sensory Representations in Primary Visual Cortex. Neuron 2015, 86, 1478–1490. [Google Scholar] [CrossRef] [PubMed]
- Goard, M.J.; Pho, G.N.; Woodson, J.; Sur, M. Distinct roles of visual, parietal, and frontal motor cortices in memory-guided sensorimotor decisions. Elife 2016, 5, e13764. [Google Scholar] [CrossRef] [PubMed]
- Resulaj, A.; Ruediger, S.; Olsen, S.R.; Scanziani, M. First spikes in visual cortex enable perceptual discrimination. Elife 2018, 7, e34044. [Google Scholar] [CrossRef] [PubMed]
- Wolf, A.; Bauer, B.; Abner, E.L.; Ashkenazy-Frolinger, T.; Hartz, A.M. A Comprehensive Behavioral Test Battery to Assess Learning and Memory in 129S6/Tg2576 Mice. PLoS ONE 2016, 11, e0147733. [Google Scholar] [CrossRef] [PubMed]
- Ognibene, E.; Middei, S.; Daniele, S.; Adriani, W.; Ghirardi, O.; Caprioli, A.; Laviola, G. Aspects of spatial memory and behavioral disinhibition in Tg2576 transgenic mice as a model of Alzheimer’s disease. Behav. Brain Res. 2005, 156, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, R.; Lews, T.L.; Strazielle, C.; Kim, H.; Fukuchi, K. Transgenic mice expressing the betaAPP695SWE mutation: Effects on exploratory activity, anxiety, and motor coordination. Brain Res. 2003, 977, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Gil-Bea, F.J.; Aisa, B.; Schliebs, R.; Ramírez, M.J. Increase of locomotor activity underlying the behavioral disinhibition in tg2576 mice. Behav. Neurosci. 2007, 121, 340–344. [Google Scholar] [CrossRef]
- Roberson, E.D.; Scearce-Levie, K.; Palop, J.J.; Yan, F.; Cheng, I.H.; Wu, T.; Gerstein, H.; Yu, G.Q.; Mucke, L. Reducing endogenous tau ameliorates amyloid beta-induced deficits in an Alzheimer’s disease mouse model. Science 2007, 316, 750–754. [Google Scholar] [CrossRef]
- Thompson, S.M.; Berkowitz, L.E.; Clark, B.J. Behavioral and neural subsystems of rodent exploration. Learn. Motiv. 2018, 61, 3–15. [Google Scholar] [CrossRef]
- Kaneko, M.; Fu, Y.; Stryker, M.P. Locomotion induces stimulus-specific response enhancement in adult visual cortex. J. Neurosci. 2017, 37, 3532–3543. [Google Scholar] [CrossRef]
- Goldstein, L.E.; Muffat, J.A.; Cherny, R.A.; Moir, R.D.; Ericsson, M.H.; Huang, X.; Mavros, C.; Coccia, J.A.; Faget, K.Y.; Fitch, K.A.; et al. Cytosolic beta-amyloid deposition and supranuclear cataracts in lenses from people with Alzheimer’s disease. Lancet 2003, 361, 1258–1265. [Google Scholar] [CrossRef]
- Blanks, J.C.; Schmidt, S.Y.; Torigoe, Y.; Porrello, K.V.; Hinton, D.R.; Blanks, R.H. Retinal pathology in Alzheimer’s disease. II. Regional neuron loss and glial changes in GCL. Neurobiol. Aging 1996, 17, 385–395. [Google Scholar] [CrossRef]
- Armstrong, R.A. Visual feld defects in Alzheimer’s disease patients may reflect differential pathology in the primary visual cortex. Optom. Vis. Sci. 1996, 73, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Hof, P.R.; Morrison, J.H. Quantitative analysis of a vulnerable subset of pyramidal neurons in Alzheimer’s disease: II. Primary and secondary visual cortex. J. Comp. Neurol. 1990, 301, 55–64. [Google Scholar] [CrossRef] [PubMed]
- McKee, A.C.; Au, R.; Cabral, H.J.; Kowall, N.W.; Seshadri, S.; Kubilus, C.A.; Drake, J.; Wolf, P.A. Visual association pathology in preclinical Alzheimer disease. J. Neuropathol. Exp. Neurol. 2006, 65, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Berisha, F.; Feke, G.T.; Trempe, C.L.; McMeel, J.W.; Schepens, C.L. Retinal abnormalities in early Alzheimer’s disease. Investig. Ophthalmol. Vis. Sci. 2007, 48, 2285–2289. [Google Scholar] [CrossRef]
- Bublak, P.; Redel, P.; Sorg, C.; Kurz, A.; Förstl, H.; Müller, H.J.; Schneider, W.X.; Finke, K. Staged decline of visual processing capacity in mild cognitive impairment and Alzheimer’s disease. Neurobiol. Aging 2011, 32, 1219–1230. [Google Scholar] [CrossRef]
- Braak, H.; Braak, E.; Bohl, J. Staging of Alzheimer-related cortical destruction. Eur. Neurol. 1993, 33, 403–408. [Google Scholar] [CrossRef]
- Lewis, D.A.; Campbell, M.J.; Terry, R.D.; Morrison, J.H. Laminar and regional distributions of neurofibrillary tangles and neuritic plaques in Alzheimer’s disease: A quantitative study of visual and auditory cortices. J. Neurosci. 1987, 7, 1799–1808. [Google Scholar] [CrossRef]
- Holtmaat, A.; Wilbrecht, L.; Knott, G.W.; Welker, E.; Svoboda, K. Experience-dependent and cell-type-specific spine growth in the neocortex. Nature 2006, 441, 979–983. [Google Scholar] [CrossRef]
- Pelucchi, S.; Stringhi, R.; Marcello, E. Dendritic Spines in Alzheimer’s Disease: How the Actin Cytoskeleton Contributes to Synaptic Failure. Int. J. Mol. Sci. 2020, 21, 908. [Google Scholar] [CrossRef]
- Wang, Q.; Yuan, W.; Yang, X.; Wang, Y.; Li, Y.; Qiao, H. Role of Cofilin in Alzheimer’s Disease. Front. Cell Dev. Biol. 2020, 8, 584898. [Google Scholar] [CrossRef]
- Bamburg, J.R.; Minamide, L.S.; Wiggan, O.; Tahtamouni, L.H.; Kuhn, T.B. Cofilin and Actin Dynamics: Multiple Modes of Regulation and Their Impacts in Neuronal Development and Degeneration. Cells 2021, 10, 2726. [Google Scholar] [CrossRef] [PubMed]
- Maloney, M.T.; Bamburg, J.R. Cofilin-mediated neurodegeneration in Alzheimer’s disease and other amyloidopathies. Mol. Neurobiol. 2007, 35, 21–44. [Google Scholar] [CrossRef]
- Rush, T.; Martinez-Hernandez, J.; Dollmeyer, M.; Frandemiche, M.L.; Borel, E.; Boisseau, S.; Jacquier-Sarlin, M.; Buisson, A. Synaptotoxicity in Alzheimer’s Disease Involved a Dysregulation of Actin Cytoskeleton Dynamics through Cofilin 1 Phosphorylation. J. Neurosci. 2018, 38, 10349–10361. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Vidal, G.S.; Djurisic, M.; William, C.M.; Birnbaum, M.E.; Garcia, K.C.; Hyman, B.T.; Schatz, C.J. Human LilrB2 is a β-amyloid receptor and its murine homolog PirB regulates synaptic plasticity in an Alzheimer’s model. Science 2013, 341, 1399–1404. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.A.; Jung, A.R.; Lakshmana, M.K.; Bedrossian, A.; Lim, Y.; Bu, J.H.; Park, S.A.; Koo, E.H.; Mook-Jung, I.; Kang, D.E. Pivotal role of the RanBP9-cofilin pathway in Aβ-induced apoptosis and neurodegeneration. Cell Death Differ. 2012, 19, 1413–1423. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.A.; Zhao, X.; Khan, H.; Penn, C.; Wang, X.; Joly-Amado, A.; Weeber, E.; Morgan, D.; Kang, D.E. Slingshot-cofilin activation mediates mitochondrial and synaptic dysfunction via Aβ ligation to β1-integrin conformers. Cell Death Differ. 2015, 22, 921–934. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.A.; Bogges, T.; Uhlar, C.; Wang, X.; Khan, H.; Cappos, G.; Joly-Amado, A.; De Narvaez, E.; Majid, S.; Minamide, L.S.; et al. RanBP9 at the intersection between cofilin and Aβ pathologies: Rescue of neurodegenerative changes by RanBP9 reduction. Cell Death Dis. 2015, 6, 1676. [Google Scholar] [CrossRef] [PubMed]
- Kommaddi, R.; Das, D.; Karunakaran, S.; Nanguneri, S.; Bapat, D.; Ray, A.; Shaw, E.; Bennett, D.A.; Nair, D.; Ravindranath, V. Aβ mediates F-actin disassembly in dendritic spines leading to cognitive deficits in Alzheimer’s disease. J. Neurosci. 2018, 38, 1085–1099. [Google Scholar] [CrossRef]
- Deng, Y.; Wei, J.; Cheng, J.; Zhong, P.; Xiong, Z.; Liu, A.; Lin, L.; Chen, S.; Yan, Z. Partial Amelioration of Synaptic and Cognitive Deficits by Inhibiting Cofilin Dephosphorylation in an Animal Model of Alzheimer’s Disease. J. Alzheimer’s Dis. 2016, 53, 1419–1432. [Google Scholar] [CrossRef]
- Williams, P.A.; Thirgood, R.A.; Oliphant, H.; Frizzati, A.; Littlewood, E.; Votruba, M.; Good, M.A.; Williams, J.; Morgan, J.E. Retinal ganglion cell dendritic degeneration in a mouse model of Alzheimer’s disease. Neurobiol. Aging 2013, 34, 1799–1806. [Google Scholar] [CrossRef]
- Bevan, R.J.; Hughes, T.R.; Williams, P.A.; Good, M.A.; Morgan, B.P.; Morgan, J.E. Retinal ganglion cell degeneration correlates with hippocampal spine loss in experimental Alzheimer’s disease. Acta Neuropathol. Commun. 2020, 8, 216. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, D.; Votruba, M. Can the retina be used to diagnose and plot the progression of Alzheimer’s disease? Acta Ophthalmol. 2017, 95, 768–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Latina, V.; De Introna, M.; Caligiuri, C.; Loviglio, A.; Florio, R.; La Regina, F.; Pignataro, A.; Ammassari-Teule, M.; Calissano, P.; Amadoro, G. Immunotherapy with Cleavage-Specific 12A12mAb Reduces the Tau Cleavage in Visual Cortex and Improves Visuo-Spatial Recognition Memory in Tg2576 AD Mouse Model. Pharmaceutics 2023, 15, 509. https://doi.org/10.3390/pharmaceutics15020509
Latina V, De Introna M, Caligiuri C, Loviglio A, Florio R, La Regina F, Pignataro A, Ammassari-Teule M, Calissano P, Amadoro G. Immunotherapy with Cleavage-Specific 12A12mAb Reduces the Tau Cleavage in Visual Cortex and Improves Visuo-Spatial Recognition Memory in Tg2576 AD Mouse Model. Pharmaceutics. 2023; 15(2):509. https://doi.org/10.3390/pharmaceutics15020509
Chicago/Turabian StyleLatina, Valentina, Margherita De Introna, Chiara Caligiuri, Alessia Loviglio, Rita Florio, Federico La Regina, Annabella Pignataro, Martine Ammassari-Teule, Pietro Calissano, and Giuseppina Amadoro. 2023. "Immunotherapy with Cleavage-Specific 12A12mAb Reduces the Tau Cleavage in Visual Cortex and Improves Visuo-Spatial Recognition Memory in Tg2576 AD Mouse Model" Pharmaceutics 15, no. 2: 509. https://doi.org/10.3390/pharmaceutics15020509