
Nanoparticles for Drug and Gene Delivery in Pediatric Brain Tumors’ Cancer Stem Cells: Current Knowledge and Future Perspectives
 , , ,
, , ,  and
and
Abstract
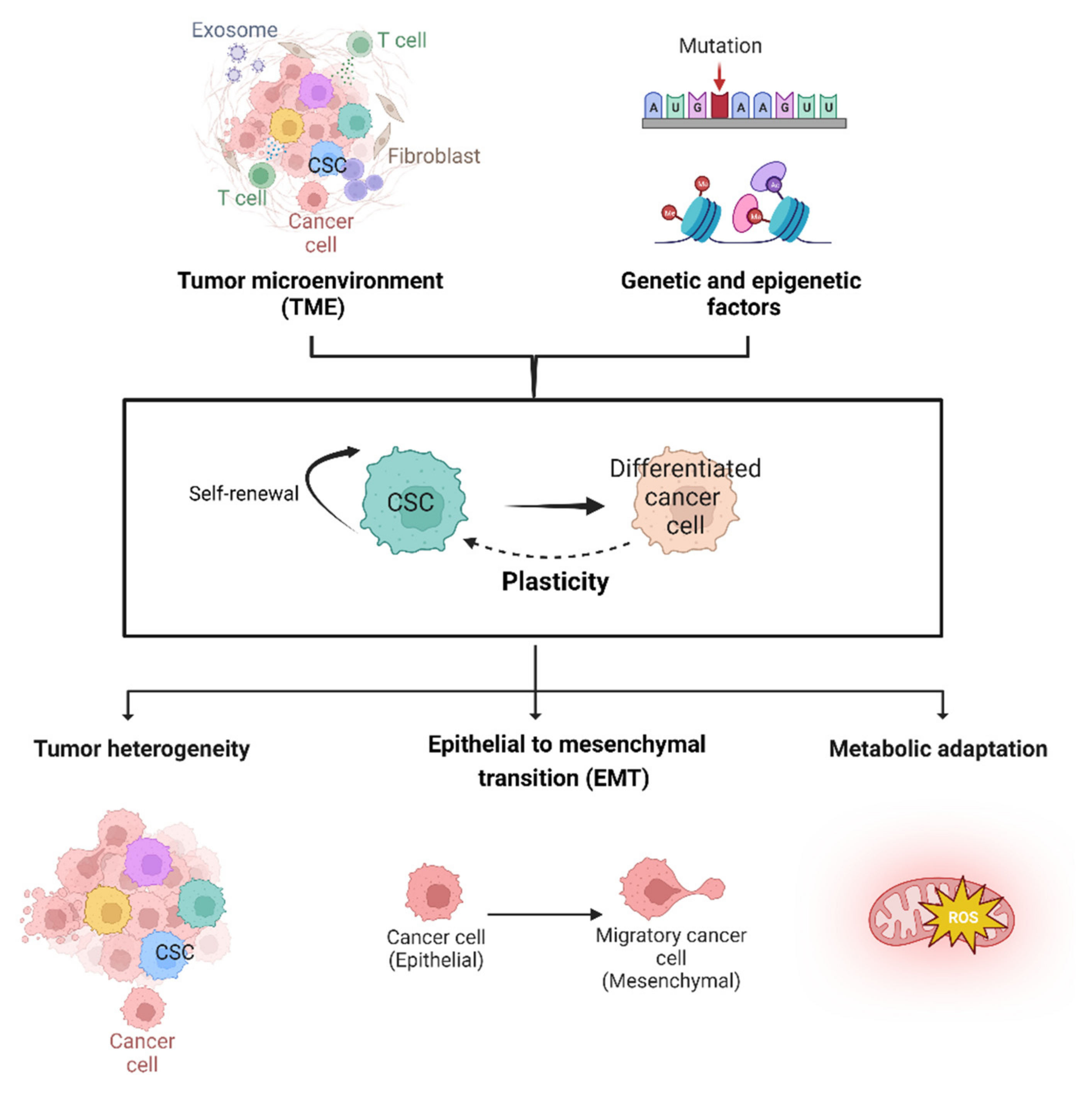
:1. Introduction
2. Nanoparticles and Target Therapy
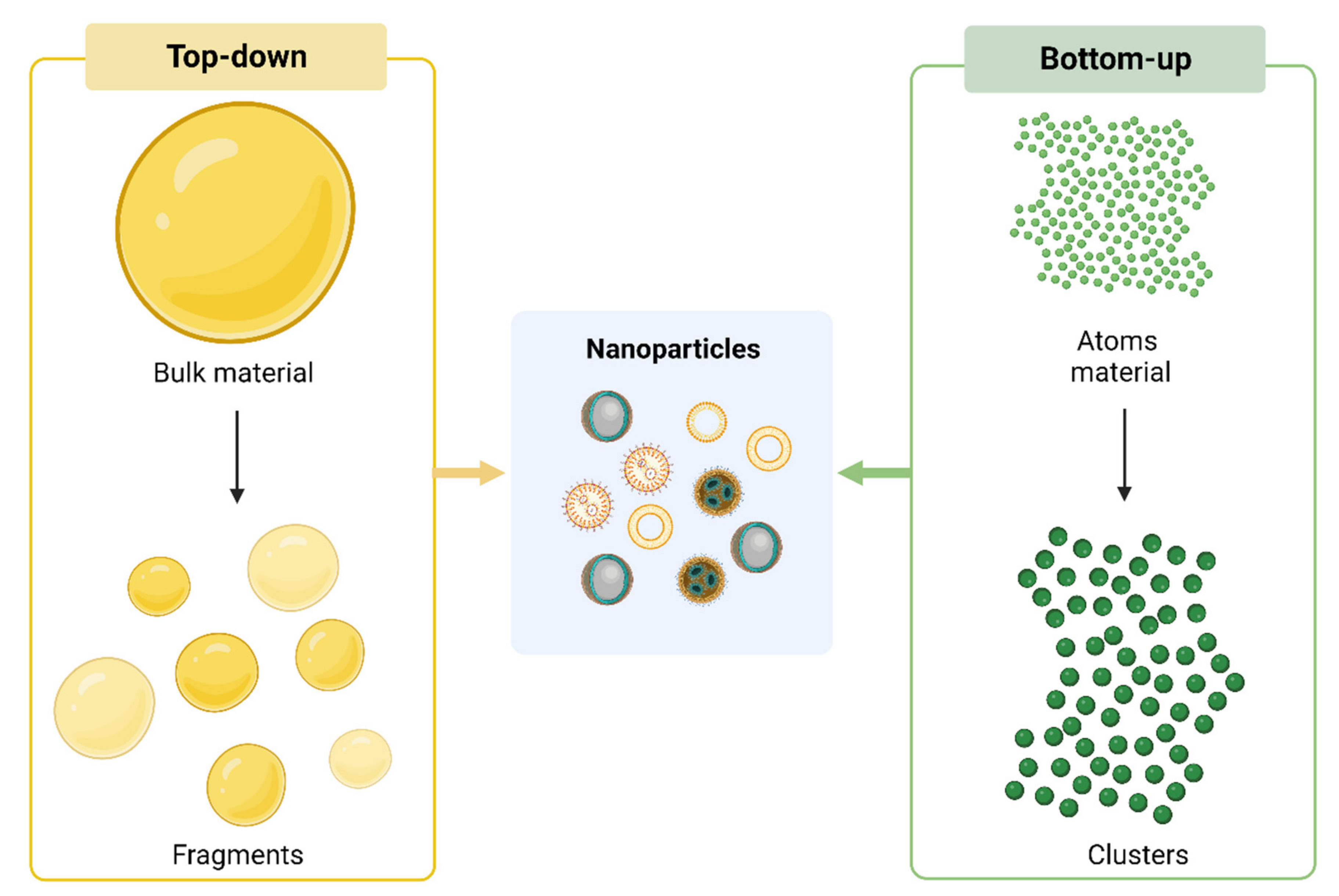
2.1. Synthesis of NPs
2.2. Mechanisms of Action
2.3. Passive Targeting
2.4. Active Targeting
2.4.1. Epidermal Growth Factor Receptor
2.4.2. Transferrin Receptor
2.4.3. Insulin Receptor
2.4.4. Lipoprotein
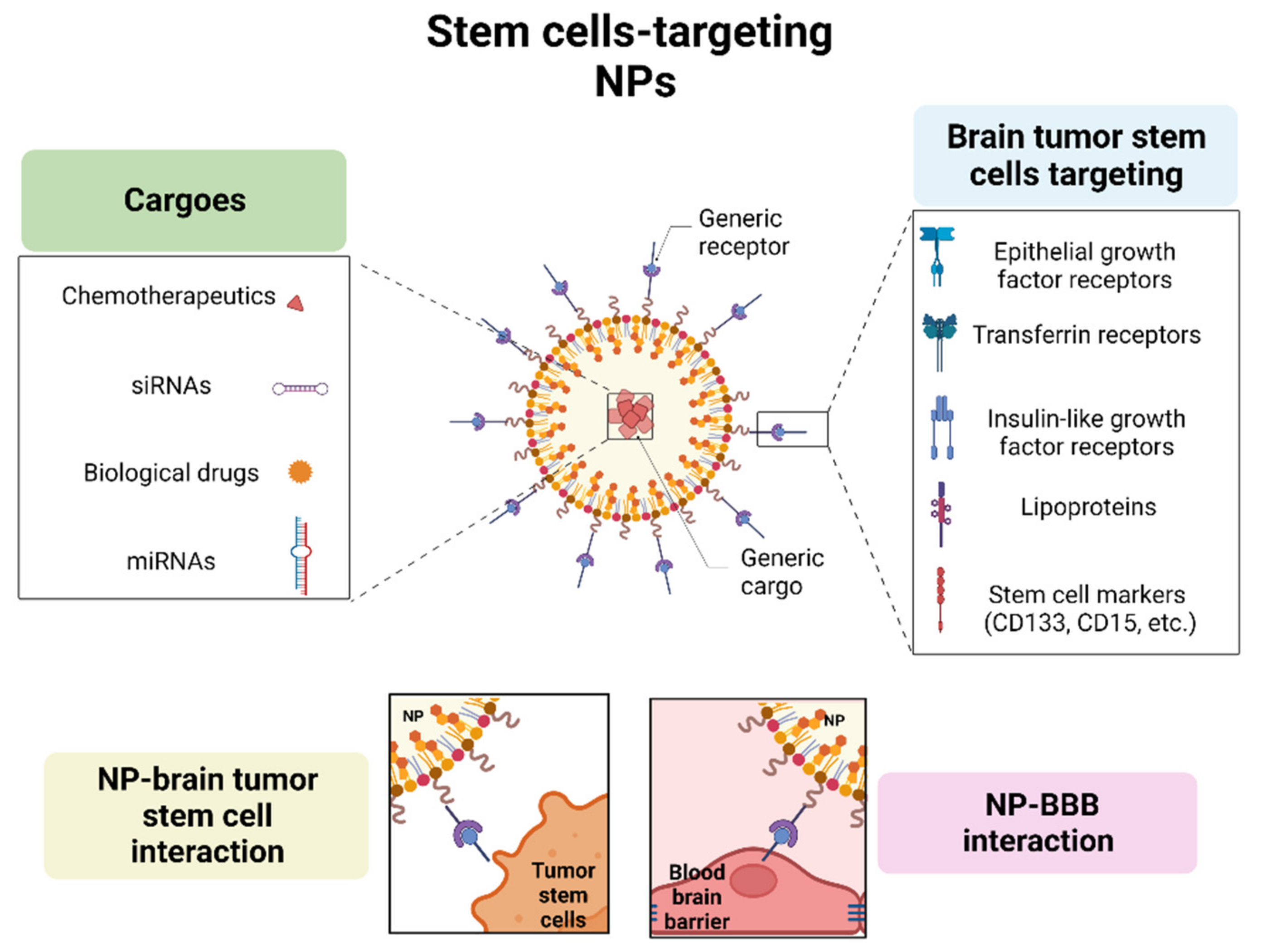
3. Nanoparticles and Brain CSCs Compartment
4. Relevant Cargoes for Brain CSCs
4.1. Drugs
4.2. Small Interfering RNA

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nanoparticle | Cargo | Application | Type of Cancer | Mechanism of Action | In Vitro/In Vivo Models | References |
|---|---|---|---|---|---|---|
| Self-assembling amphiphilic polymer forming micelles, called mPEG5kDa-cholane | Glabrescione B (Hedgehog inhibitor) | Drug delivery | SHH MB | Inhibition of tumor growth |
| [67] |
| HDL-mimetic nanoparticles (eHNPs) composed of apolipoprotein A1 and CD15 | LDE-225 (Smo inhibitor, sonidegib) | Drug delivery | SHH MB | Intracellular cholesterol depletion and cytotoxicity |
| [68] |
| Amphiphilic polymeric nanoparticles modified with a protease resistant peptide | SN-38 (Topoisomerase I inhibitor) | Drug delivery | DIPG | Apoptosis |
| [69] |
| pH-sensitive core-shell nanoparticles | Doxorubicin hydrochloride and curcumin | Drug delivery | Glioma | Cytotoxicity | Rat model of glioma | [70] |
| Immunoliposome using angiopep-2 and anti-CD133 monoclonal antibody | Temozolomide | Drug delivery | GBM | Cytotoxicity and reduction of CD133+-positive cells | Glioblastoma orthotopic mouse model | [71] |
| CD133-Functionalized Gold Nanoparticles | GLS1 inhibitor Telaglenastat (CB-839) | Drug delivery | GBM | Inhibition of glutaminolysis |
| [74] |
| Hyaluronan (HA)-grafted lipid-based nanoparticles | RNAi polo-like kinase 1 (PLK1) | Small interfering (siRNA) delivery | GBM | Inhibition of glutaminolysis |
| [78] |
| Lipopolymeric nanoparticle | Multiple siRNAs (SOX2, OLIG2, SALL2 and POU3F2) | Small interfering (siRNA) delivery | GBM | Inhibition of self-renewal and tumorigenicity |
| [79] |
| Bioreducible poly(beta-amino ester) nanoparticles | miR-148a and miR-296-5p | microRNAs (miRNAs) delivery | GBM | Inhibition of tumorigenicity |
| [92] |
| Polyfunctional gold–iron oxide nanoparticles (polyGION) | miR-100, anti-miR-21 and temozolomide | microRNAs (miRNAs) and drug delivery | GBM | Increased in vivo survival |
| [93] |
| Serum albumin coated passion fruit-like nanoarchitectures (NAs-HSA) | Doxorubicin | Drug delivery | HGG | Apoptosis in vitro but not in vivo |
| [65] |
4.3. microRNAs
5. Immunotherapy and NPs in Pediatric Brain Tumors
6. Advanced Pre-Clinical Models to Study NPs in Pediatric Brain Tumors
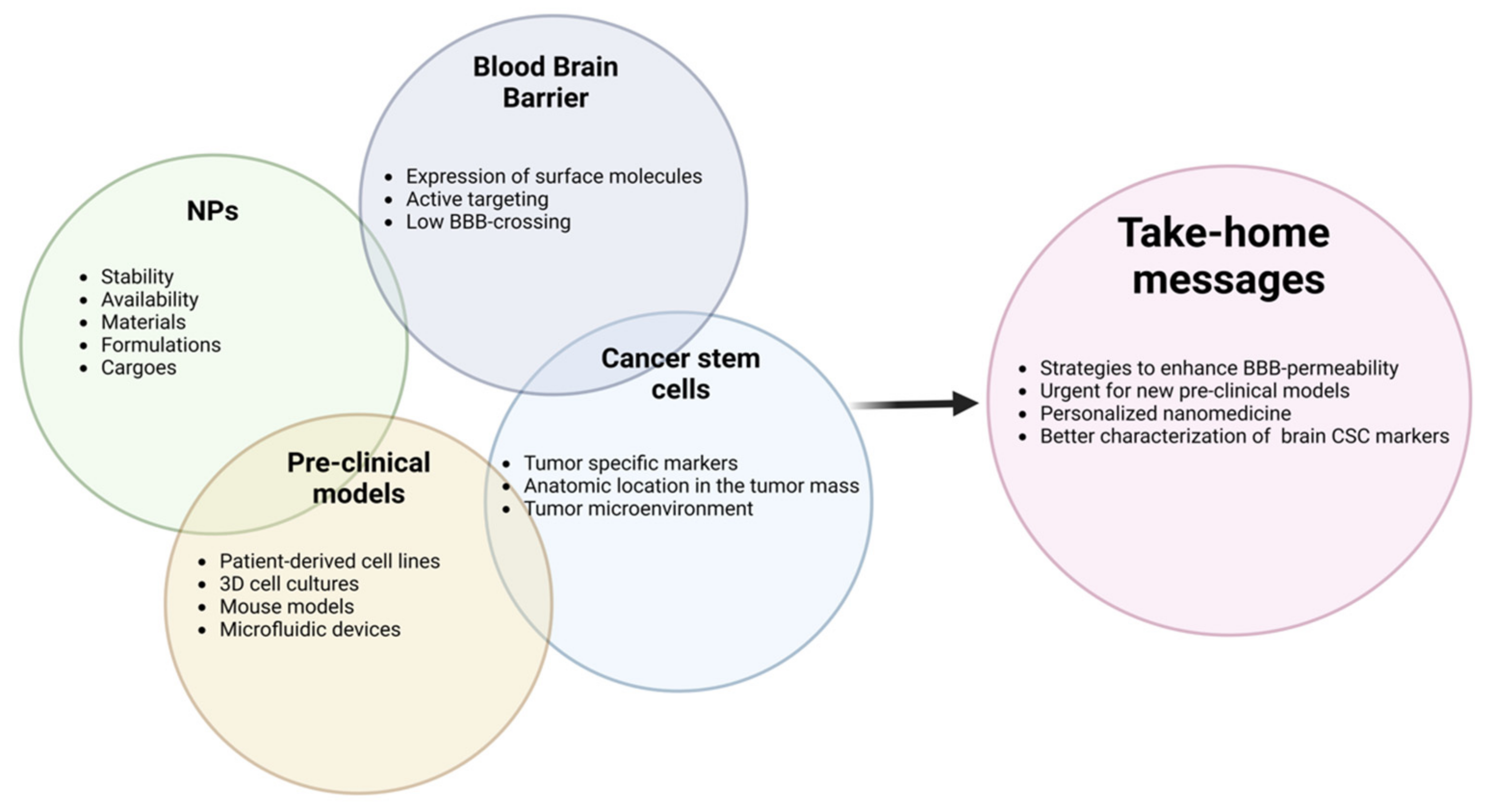
7. Future Perspectives
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cohen, A.R. Brain Tumors in Children. N. Engl. J. Med. 2022, 386, 1922–1931. [Google Scholar] [CrossRef] [PubMed]
- Pollack, I.F.; Agnihotri, S.; Broniscer, A. Childhood Brain Tumors: Current Management, Biological Insights, and Future Directions. J. Neurosurg. Pediatr. 2019, 23, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Claude, F.; Ubertini, G.; Szinnai, G. Endocrine Disorders in Children with Brain Tumors: At Diagnosis, after Surgery, Radiotherapy and Chemotherapy. Children 2022, 9, 1617. [Google Scholar] [CrossRef]
- Otth, M.; Wyss, J.; Scheinemann, K. Long-Term Follow-Up of Pediatric CNS Tumor Survivors-A Selection of Relevant Long-Term Issues. Children 2022, 9, 447. [Google Scholar] [CrossRef] [PubMed]
- El Demerdash, N.; Kedda, J.; Ram, N.; Brem, H.; Tyler, B. Novel Therapeutics for Brain Tumors: Current Practice and Future Prospects. Expert Opin. Drug Deliv. 2020, 17, 9–21. [Google Scholar] [CrossRef]
- Haydar, D.; Ibañez-Vega, J.; Krenciute, G. T-Cell Immunotherapy for Pediatric High-Grade Gliomas: New Insights to Overcoming Therapeutic Challenges. Front. Oncol. 2021, 11, 718030. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Conniot, J.; Amorim, J.; Jin, Y.; Prasad, R.; Yan, X.; Fan, K.; Conde, J. Nucleic Acid-Based Therapy for Brain Cancer: Challenges and Strategies. J. Control. Release Off. J. Control. Release Soc. 2022, 350, 80–92. [Google Scholar] [CrossRef]
- Dobson, T.H.W.; Gopalakrishnan, V. Preclinical Models of Pediatric Brain Tumors-Forging Ahead. Bioengineering 2018, 5, 81. [Google Scholar] [CrossRef]
- Löscher, W.; Potschka, H. Drug Resistance in Brain Diseases and the Role of Drug Efflux Transporters. Nat. Rev. Neurosci. 2005, 6, 591–602. [Google Scholar] [CrossRef]
- Haumann, R.; Videira, J.C.; Kaspers, G.J.L.; van Vuurden, D.G.; Hulleman, E. Overview of Current Drug Delivery Methods Across the Blood-Brain Barrier for the Treatment of Primary Brain Tumors. CNS Drugs 2020, 34, 1121–1131. [Google Scholar] [CrossRef]
- Aldape, K.; Brindle, K.M.; Chesler, L.; Chopra, R.; Gajjar, A.; Gilbert, M.R.; Gottardo, N.; Gutmann, D.H.; Hargrave, D.; Holland, E.C.; et al. Challenges to Curing Primary Brain Tumours. Nat. Rev. Clin. Oncol. 2019, 16, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Hawkins, C.; Clarke, I.D.; Squire, J.A.; Bayani, J.; Hide, T.; Henkelman, R.M.; Cusimano, M.D.; Dirks, P.B. Identification of Human Brain Tumour Initiating Cells. Nature 2004, 432, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Hemmati, H.D.; Nakano, I.; Lazareff, J.A.; Masterman-Smith, M.; Geschwind, D.H.; Bronner-Fraser, M.; Kornblum, H.I. Cancerous Stem Cells Can Arise from Pediatric Brain Tumors. Proc. Natl. Acad. Sci. USA 2003, 100, 15178–15183. [Google Scholar] [CrossRef] [PubMed]
- Valent, P.; Bonnet, D.; De Maria, R.; Lapidot, T.; Copland, M.; Melo, J.V.; Chomienne, C.; Ishikawa, F.; Schuringa, J.J.; Stassi, G.; et al. Cancer Stem Cell Definitions and Terminology: The Devil Is in the Details. Nat. Rev. Cancer 2012, 12, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.; Wu, Q.; McLendon, R.E.; Hao, Y.; Shi, Q.; Hjelmeland, A.B.; Dewhirst, M.W.; Bigner, D.D.; Rich, J.N. Glioma Stem Cells Promote Radioresistance by Preferential Activation of the DNA Damage Response. Nature 2006, 444, 756–760. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, Y.; Yu, T.-S.; McKay, R.M.; Burns, D.K.; Kernie, S.G.; Parada, L.F. A Restricted Cell Population Propagates Glioblastoma Growth after Chemotherapy. Nature 2012, 488, 522–526. [Google Scholar] [CrossRef]
- Lauko, A.; Lo, A.; Ahluwalia, M.S.; Lathia, J.D. Cancer Cell Heterogeneity & Plasticity in Glioblastoma and Brain Tumors. Semin. Cancer Biol. 2022, 82, 162–175. [Google Scholar] [CrossRef]
- Bocci, F.; Gearhart-Serna, L.; Boareto, M.; Ribeiro, M.; Ben-Jacob, E.; Devi, G.R.; Levine, H.; Onuchic, J.N.; Jolly, M.K. Toward Understanding Cancer Stem Cell Heterogeneity in the Tumor Microenvironment. Proc. Natl. Acad. Sci. USA 2019, 116, 148–157. [Google Scholar] [CrossRef]
- Dando, I.; Cordani, M.; Dalla Pozza, E.; Biondani, G.; Donadelli, M.; Palmieri, M. Antioxidant Mechanisms and ROS-Related MicroRNAs in Cancer Stem Cells. Oxid. Med. Cell. Longev. 2015, 2015, 425708. [Google Scholar] [CrossRef]
- Gimple, R.C.; Yang, K.; Halbert, M.E.; Agnihotri, S.; Rich, J.N. Brain Cancer Stem Cells: Resilience through Adaptive Plasticity and Hierarchical Heterogeneity. Nat. Rev. Cancer 2022, 22, 497–514. [Google Scholar] [CrossRef]
- Gulaia, V.; Kumeiko, V.; Shved, N.; Cicinskas, E.; Rybtsov, S.; Ruzov, A.; Kagansky, A. Molecular Mechanisms Governing the Stem Cell’s Fate in Brain Cancer: Factors of Stemness and Quiescence. Front. Cell. Neurosci. 2018, 12, 388. [Google Scholar] [CrossRef] [PubMed]
- Abballe, L.; Miele, E. Epigenetic Modulators for Brain Cancer Stem Cells: Implications for Anticancer Treatment. World J. Stem Cells 2021, 13, 670–684. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering Precision Nanoparticles for Drug Delivery. Nat. Rev. Drug Discov. 2021, 20, 101–124. [Google Scholar] [CrossRef] [PubMed]
- Farokhzad, O.C.; Langer, R. Impact of Nanotechnology on Drug Delivery. ACS Nano 2009, 3, 16–20. [Google Scholar] [CrossRef]
- Cheng, Y.; Morshed, R.A.; Auffinger, B.; Tobias, A.L.; Lesniak, M.S. Multifunctional Nanoparticles for Brain Tumor Imaging and Therapy. Adv. Drug Deliv. Rev. 2014, 66, 42–57. [Google Scholar] [CrossRef]
- Dang, Y.; Guan, J. Nanoparticle-Based Drug Delivery Systems for Cancer Therapy. Smart Mater. Med. 2020, 1, 10–19. [Google Scholar] [CrossRef]
- Mitusova, K.; Peltek, O.O.; Karpov, T.E.; Muslimov, A.R.; Zyuzin, M.V.; Timin, A.S. Overcoming the Blood–Brain Barrier for the Therapy of Malignant Brain Tumor: Current Status and Prospects of Drug Delivery Approaches. J. Nanobiotechnol. 2022, 20, 412. [Google Scholar] [CrossRef]
- Sonali; Viswanadh, M.K.; Singh, R.P.; Agrawal, P.; Mehata, A.K.; Pawde, D.M.; Narendra; Sonkar, R.; Muthu, M.S. Nanotheranostics: Emerging Strategies for Early Diagnosis and Therapy of Brain Cancer. Nanotheranostics 2018, 2, 70–86. [Google Scholar] [CrossRef]
- Ahmad, F.; Varghese, R.; Panda, S.; Ramamoorthy, S.; Areeshi, M.Y.; Fagoonee, S.; Haque, S. Smart Nanoformulations for Brain Cancer Theranostics: Challenges and Promises. Cancers 2022, 14, 5389. [Google Scholar] [CrossRef]
- Ertas, Y.N.; Abedi Dorcheh, K.; Akbari, A.; Jabbari, E. Nanoparticles for Targeted Drug Delivery to Cancer Stem Cells: A Review of Recent Advances. Nanomaterials 2021, 11, 1755. [Google Scholar] [CrossRef]
- Baig, N.; Kammakakam, I.; Falath, W. Nanomaterials: A Review of Synthesis Methods, Properties, Recent Progress, and Challenges. Mater. Adv. 2021, 2, 1821–1871. [Google Scholar] [CrossRef]
- Shafey, A.M.E. Green Synthesis of Metal and Metal Oxide Nanoparticles from Plant Leaf Extracts and Their Applications: A Review. Green Process. Synth. 2020, 9, 304–339. [Google Scholar] [CrossRef]
- Bhardwaj, B.; Singh, P.; Kumar, A.; Kumar, S.; Budhwar, V. Eco-Friendly Greener Synthesis of Nanoparticles. Adv. Pharm. Bull. 2020, 10, 566–576. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y.; Maeda, H. A New Concept for Macromolecular Therapeutics in Cancer Chemotherapy: Mechanism of Tumoritropic Accumulation of Proteins and the Antitumor Agent Smancs. Cancer Res. 1986, 46, 6387–6392. [Google Scholar]
- Benjamin, L.E.; Golijanin, D.; Itin, A.; Pode, D.; Keshet, E. Selective Ablation of Immature Blood Vessels in Established Human Tumors Follows Vascular Endothelial Growth Factor Withdrawal. J. Clin. Investig. 1999, 103, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Wu, J. The Enhanced Permeability and Retention (EPR) Effect: The Significance of the Concept and Methods to Enhance Its Application. J. Pers. Med. 2021, 11, 771. [Google Scholar] [CrossRef] [PubMed]
- Bazak, R.; Houri, M.; El Achy, S.; Kamel, S.; Refaat, T. Cancer Active Targeting by Nanoparticles: A Comprehensive Review of Literature. J. Cancer Res. Clin. Oncol. 2015, 141, 769–784. [Google Scholar] [CrossRef]
- Salahpour Anarjan, F. Active Targeting Drug Delivery Nanocarriers: Ligands. Nano-Struct. Nano Objects 2019, 19, 100370. [Google Scholar] [CrossRef]
- Bayda, S.; Adeel, M.; Tuccinardi, T.; Cordani, M.; Rizzolio, F. The History of Nanoscience and Nanotechnology: From Chemical-Physical Applications to Nanomedicine. Molecules 2019, 25, 112. [Google Scholar] [CrossRef]
- Yao, Y.; Zhou, Y.; Liu, L.; Xu, Y.; Chen, Q.; Wang, Y.; Wu, S.; Deng, Y.; Zhang, J.; Shao, A. Nanoparticle-Based Drug Delivery in Cancer Therapy and Its Role in Overcoming Drug Resistance. Front. Mol. Biosci. 2020, 7, 193. [Google Scholar] [CrossRef]
- Sigismund, S.; Avanzato, D.; Lanzetti, L. Emerging Functions of the EGFR in Cancer. Mol. Oncol. 2018, 12, 3–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Zhao, Z.; Gao, H.; Rostami, I.; You, Q.; Jia, X.; Wang, C.; Zhu, L.; Yang, Y. Enhanced Blood-Brain-Barrier Penetrability and Tumor-Targeting Efficiency by Peptide-Functionalized Poly(Amidoamine) Dendrimer for the Therapy of Gliomas. Nanotheranostics 2019, 3, 311–330. [Google Scholar] [CrossRef]
- Cheng, Y.; Meyers, J.D.; Agnes, R.S.; Doane, T.L.; Kenney, M.E.; Broome, A.-M.; Burda, C.; Basilion, J.P. Addressing Brain Tumors with Targeted Gold Nanoparticles: A New Gold Standard for Hydrophobic Drug Delivery? Small Weinh. Bergstr. Ger. 2011, 7, 2301–2306. [Google Scholar] [CrossRef] [PubMed]
- Whittle, J.R.; Lickliter, J.D.; Gan, H.K.; Scott, A.M.; Simes, J.; Solomon, B.J.; MacDiarmid, J.A.; Brahmbhatt, H.; Rosenthal, M.A. First in Human Nanotechnology Doxorubicin Delivery System to Target Epidermal Growth Factor Receptors in Recurrent Glioblastoma. J. Clin. Neurosci. Off. J. Neurosurg. Soc. Australas. 2015, 22, 1889–1894. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, R.R.; Mahajan, S.D.; Pliss, A.; Prasad, P.N. Small Molecule Based EGFR Targeting of Biodegradable Nanoparticles Containing Temozolomide and Cy5 Dye for Greatly Enhanced Image-Guided Glioblastoma Therapy. Nanomed. Nanotechnol. Biol. Med. 2022, 41, 102513. [Google Scholar] [CrossRef] [PubMed]
- Meola, A.; Rao, J.; Chaudhary, N.; Sharma, M.; Chang, S.D. Gold Nanoparticles for Brain Tumor Imaging: A Systematic Review. Front. Neurol. 2018, 9, 328. [Google Scholar] [CrossRef] [PubMed]
- Hadjipanayis, C.G.; Machaidze, R.; Kaluzova, M.; Wang, L.; Schuette, A.J.; Chen, H.; Wu, X.; Mao, H. EGFRvIII Antibody-Conjugated Iron Oxide Nanoparticles for Magnetic Resonance Imaging-Guided Convection-Enhanced Delivery and Targeted Therapy of Glioblastoma. Cancer Res. 2010, 70, 6303–6312. [Google Scholar] [CrossRef]
- Shen, Y.; Li, X.; Dong, D.; Zhang, B.; Xue, Y.; Shang, P. Transferrin Receptor 1 in Cancer: A New Sight for Cancer Therapy. Am. J. Cancer Res. 2018, 8, 916–931. [Google Scholar] [PubMed]
- Rosager, A.M.; Sørensen, M.D.; Dahlrot, R.H.; Hansen, S.; Schonberg, D.L.; Rich, J.N.; Lathia, J.D.; Kristensen, B.W. Transferrin Receptor-1 and Ferritin Heavy and Light Chains in Astrocytic Brain Tumors: Expression and Prognostic Value. PLoS ONE 2017, 12, e0182954. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M.; Chou, T. Mathematical Models of Blood-Brain Barrier Transport of Monoclonal Antibodies Targeting the Transferrin Receptor and the Insulin Receptor. Pharmaceuticals 2021, 14, 535. [Google Scholar] [CrossRef]
- Pardridge, W.M. A Historical Review of Brain Drug Delivery. Pharmaceutics 2022, 14, 1283. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Lakkadwala, S.; Modgil, A.; Singh, J. The Role of Cell-Penetrating Peptide and Transferrin on Enhanced Delivery of Drug to Brain. Int. J. Mol. Sci. 2016, 17, 806. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Mao, J.; Jiang, Z.; Sun, T.; Hu, Y.; Jiang, Z.; Zhang, C.; Dong, J.; Huang, Q.; Lan, Q. Transferrin-Modified Doxorubicin-Loaded Biodegradable Nanoparticles Exhibit Enhanced Efficacy in Treating Brain Glioma-Bearing Rats. Cancer Biother. Radiopharm. 2013, 28, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Chang, J.; Yan, C.; Qian, X.; Long, L.; He, B.; Yuan, X.; Kang, C.; Betbeder, D.; Sheng, J.; et al. Development of Transferrin Functionalized Poly(Ethylene Glycol)/Poly(Lactic Acid) Amphiphilic Block Copolymeric Micelles as a Potential Delivery System Targeting Brain Glioma. J. Mater. Sci. Mater. Med. 2010, 21, 2673–2681. [Google Scholar] [CrossRef]
- Xiao, W.; Wang, Y.; Zhang, H.; Liu, Y.; Xie, R.; He, X.; Zhou, Y.; Liang, L.; Gao, H. The Protein Corona Hampers the Transcytosis of Transferrin-Modified Nanoparticles through Blood-Brain Barrier and Attenuates Their Targeting Ability to Brain Tumor. Biomaterials 2021, 274, 120888. [Google Scholar] [CrossRef]
- Sheykhzadeh, S.; Luo, M.; Peng, B.; White, J.; Abdalla, Y.; Tang, T.; Mäkilä, E.; Voelcker, N.H.; Tong, W.Y. Transferrin-Targeted Porous Silicon Nanoparticles Reduce Glioblastoma Cell Migration across Tight Extracellular Space. Sci. Rep. 2020, 10, 2320. [Google Scholar] [CrossRef]
- Luo, M.; Lewik, G.; Ratcliffe, J.C.; Choi, C.H.J.; Mäkilä, E.; Tong, W.Y.; Voelcker, N.H. Systematic Evaluation of Transferrin-Modified Porous Silicon Nanoparticles for Targeted Delivery of Doxorubicin to Glioblastoma. ACS Appl. Mater. Interfaces 2019, 11, 33637–33649. [Google Scholar] [CrossRef]
- Liu, D.-Z.; Cheng, Y.; Cai, R.-Q.; Wang Bd, W.-W.; Cui, H.; Liu, M.; Zhang, B.; Mei, Q.-B.; Zhou, S.-Y. The Enhancement of SiPLK1 Penetration across BBB and Its Anti Glioblastoma Activity in Vivo by Magnet and Transferrin Co-Modified Nanoparticle. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 991–1003. [Google Scholar] [CrossRef]
- Sun, T.; Wu, H.; Li, Y.; Huang, Y.; Yao, L.; Chen, X.; Han, X.; Zhou, Y.; Du, Z. Targeting Transferrin Receptor Delivery of Temozolomide for a Potential Glioma Stem Cell-Mediated Therapy. Oncotarget 2017, 8, 74451–74465. [Google Scholar] [CrossRef]
- Kim, S.-S.; Rait, A.; Rubab, F.; Rao, A.K.; Kiritsy, M.C.; Pirollo, K.F.; Wang, S.; Weiner, L.M.; Chang, E.H. The Clinical Potential of Targeted Nanomedicine: Delivering to Cancer Stem-like Cells. Mol. Ther. J. Am. Soc. Gene Ther. 2014, 22, 278–291. [Google Scholar] [CrossRef]
- Shilo, M.; Motiei, M.; Hana, P.; Popovtzer, R. Transport of Nanoparticles through the Blood-Brain Barrier for Imaging and Therapeutic Applications. Nanoscale 2014, 6, 2146–2152. [Google Scholar] [CrossRef]
- Wu, D.; Yang, J.; Pardridge, W.M. Drug Targeting of a Peptide Radiopharmaceutical through the Primate Blood-Brain Barrier in Vivo with a Monoclonal Antibody to the Human Insulin Receptor. J. Clin. Investig. 1997, 100, 1804–1812. [Google Scholar] [CrossRef] [PubMed]
- Dieu, L.-H.; Wu, D.; Palivan, C.G.; Balasubramanian, V.; Huwyler, J. Polymersomes Conjugated to 83-14 Monoclonal Antibodies: In Vitro Targeting of Brain Capillary Endothelial Cells. Eur. J. Pharm. Biopharm. Off. J. Arb. Pharm. Verfahr. EV 2014, 88, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Ulbrich, K.; Knobloch, T.; Kreuter, J. Targeting the Insulin Receptor: Nanoparticles for Drug Delivery across the Blood-Brain Barrier (BBB). J. Drug Target. 2011, 19, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Pawar, S.; Koneru, T.; McCord, E.; Tatiparti, K.; Sau, S.; Iyer, A.K. LDL Receptors and Their Role in Targeted Therapy for Glioma: A Review. Drug Discov. Today 2021, 26, 1212–1225. [Google Scholar] [CrossRef]
- Hayavi, S.; Halbert, G.W. Synthetic Low-Density Lipoprotein, a Novel Biomimetic Lipid Supplement for Serum-Free Tissue Culture. Biotechnol. Prog. 2005, 21, 1262–1268. [Google Scholar] [CrossRef] [PubMed]
- Grafals-Ruiz, N.; Rios-Vicil, C.I.; Lozada-Delgado, E.L.; Quiñones-Díaz, B.I.; Noriega-Rivera, R.A.; Martínez-Zayas, G.; Santana-Rivera, Y.; Santiago-Sánchez, G.S.; Valiyeva, F.; Vivas-Mejía, P.E. Brain Targeted Gold Liposomes Improve RNAi Delivery for Glioblastoma. Int. J. Nanomed. 2020, 15, 2809–2828. [Google Scholar] [CrossRef] [PubMed]
- Nikanjam, M.; Blakely, E.A.; Bjornstad, K.A.; Shu, X.; Budinger, T.F.; Forte, T.M. Synthetic Nano-Low Density Lipoprotein as Targeted Drug Delivery Vehicle for Glioblastoma Multiforme. Int. J. Pharm. 2007, 328, 86–94. [Google Scholar] [CrossRef]
- Nikanjam, M.; Gibbs, A.R.; Hunt, C.A.; Budinger, T.F.; Forte, T.M. Synthetic Nano-LDL with Paclitaxel Oleate as a Targeted Drug Delivery Vehicle for Glioblastoma Multiforme. J. Control. Release Off. J. Control. Release Soc. 2007, 124, 163–171. [Google Scholar] [CrossRef]
- Kadari, A.; Pooja, D.; Gora, R.H.; Gudem, S.; Kolapalli, V.R.M.; Kulhari, H.; Sistla, R. Design of Multifunctional Peptide Collaborated and Docetaxel Loaded Lipid Nanoparticles for Antiglioma Therapy. Eur. J. Pharm. Biopharm. Off. J. Arb. Pharm. Verfahr. EV 2018, 132, 168–179. [Google Scholar] [CrossRef]
- Bell, J.B.; Rink, J.S.; Eckerdt, F.; Clymer, J.; Goldman, S.; Thaxton, C.S.; Platanias, L.C. HDL Nanoparticles Targeting Sonic Hedgehog Subtype Medulloblastoma. Sci. Rep. 2018, 8, 1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Ding, C.; Tan, W.; Zhang, A. Medulloblastoma: Molecular Understanding, Treatment Evolution, and New Developments. Pharmacol. Ther. 2020, 210, 107516. [Google Scholar] [CrossRef]
- Jones, D.T.W.; Jäger, N.; Kool, M.; Zichner, T.; Hutter, B.; Sultan, M.; Cho, Y.-J.; Pugh, T.J.; Hovestadt, V.; Stütz, A.M.; et al. Dissecting the Genomic Complexity Underlying Medulloblastoma. Nature 2012, 488, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Garcia, H.; Alvarado-Estrada, K.; Krishnan, S.; Quinones-Hinojosa, A.; Trifiletti, D.M. Nanoparticles for Stem Cell Therapy Bioengineering in Glioma. Front. Bioeng. Biotechnol. 2020, 8, 558375. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.J.; Bisht, S.; Bar, E.E.; Maitra, A.; Eberhart, C.G. A Polymeric Nanoparticle Formulation of Curcumin Inhibits Growth, Clonogenicity and Stem-like Fraction in Malignant Brain Tumors. Cancer Biol. Ther. 2011, 11, 464–473. [Google Scholar] [CrossRef]
- Ryskalin, L.; Biagioni, F.; Busceti, C.L.; Lazzeri, G.; Frati, A.; Fornai, F. The Multi-Faceted Effect of Curcumin in Glioblastoma from Rescuing Cell Clearance to Autophagy-Independent Effects. Molecules 2020, 25, 4839. [Google Scholar] [CrossRef]
- Ignatova, T.N.; Kukekov, V.G.; Laywell, E.D.; Suslov, O.N.; Vrionis, F.D.; Steindler, D.A. Human Cortical Glial Tumors Contain Neural Stem-like Cells Expressing Astroglial and Neuronal Markers in Vitro. Glia 2002, 39, 193–206. [Google Scholar] [CrossRef]
- Ung, C.; Tsoli, M.; Liu, J.; Cassano, D.; Pocoví-Martínez, S.; Upton, D.H.; Ehteda, A.; Mansfeld, F.M.; Failes, T.W.; Farfalla, A.; et al. Doxorubicin-Loaded Gold Nanoarchitectures as a Therapeutic Strategy against Diffuse Intrinsic Pontine Glioma. Cancers 2021, 13, 1278. [Google Scholar] [CrossRef]
- Heravi Shargh, V.; Luckett, J.; Bouzinab, K.; Paisey, S.; Turyanska, L.; Singleton, W.G.B.; Lowis, S.; Gershkovich, P.; Bradshaw, T.D.; Stevens, M.F.G.; et al. Chemosensitization of Temozolomide-Resistant Pediatric Diffuse Midline Glioma Using Potent Nanoencapsulated Forms of a N(3)-Propargyl Analogue. ACS Appl. Mater. Interfaces 2021, 13, 35266–35280. [Google Scholar] [CrossRef]
- Infante, P.; Malfanti, A.; Quaglio, D.; Balducci, S.; De Martin, S.; Bufalieri, F.; Mastrotto, F.; Basili, I.; Garofalo, M.; Lospinoso Severini, L.; et al. Glabrescione B Delivery by Self-Assembling Micelles Efficiently Inhibits Tumor Growth in Preclinical Models of Hedgehog-Dependent Medulloblastoma. Cancer Lett. 2021, 499, 220–231. [Google Scholar] [CrossRef]
- Kim, J.; Dey, A.; Malhotra, A.; Liu, J.; Ahn, S.I.; Sei, Y.J.; Kenney, A.M.; MacDonald, T.J.; Kim, Y. Engineered Biomimetic Nanoparticle for Dual Targeting of the Cancer Stem-like Cell Population in Sonic Hedgehog Medulloblastoma. Proc. Natl. Acad. Sci. USA 2020, 117, 24205–24212. [Google Scholar] [CrossRef]
- Bukchin, A.; Sanchez-Navarro, M.; Carrera, A.; Resa-Pares, C.; Castillo-Ecija, H.; Balaguer-Lluna, L.; Teixidó, M.; Olaciregui, N.G.; Giralt, E.; Carcaboso, A.M.; et al. Amphiphilic Polymeric Nanoparticles Modified with a Protease-Resistant Peptide Shuttle for the Delivery of SN-38 in Diffuse Intrinsic Pontine Glioma. ACS Appl. Nano Mater. 2021, 4, 1314–1329. [Google Scholar] [CrossRef]
- Xu, H.-L.; Fan, Z.-L.; ZhuGe, D.-L.; Tong, M.-Q.; Shen, B.-X.; Lin, M.-T.; Zhu, Q.-Y.; Jin, B.-H.; Sohawon, Y.; Yao, Q.; et al. Ratiometric Delivery of Two Therapeutic Candidates with Inherently Dissimilar Physicochemical Property through PH-Sensitive Core-Shell Nanoparticles Targeting the Heterogeneous Tumor Cells of Glioma. Drug Deliv. 2018, 25, 1302–1318. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Shin, D.H.; Kim, J.-S. Dual-Targeting Immunoliposomes Using Angiopep-2 and CD133 Antibody for Glioblastoma Stem Cells. J. Control. Release Off. J. Control. Release Soc. 2018, 269, 245–257. [Google Scholar] [CrossRef]
- Glumac, P.M.; LeBeau, A.M. The Role of CD133 in Cancer: A Concise Review. Clin. Transl. Med. 2018, 7, 18. [Google Scholar] [CrossRef]
- Affinito, A.; Quintavalle, C.; Esposito, C.L.; Roscigno, G.; Vilardo, C.; Nuzzo, S.; Ricci-Vitiani, L.; De Luca, G.; Pallini, R.; Kichkailo, A.S.; et al. The Discovery of RNA Aptamers That Selectively Bind Glioblastoma Stem Cells. Mol. Ther. Nucleic Acids 2019, 18, 99–109. [Google Scholar] [CrossRef]
- Poonaki, E.; Nickel, A.-C.; Shafiee Ardestani, M.; Rademacher, L.; Kaul, M.; Apartsin, E.; Meuth, S.G.; Gorji, A.; Janiak, C.; Kahlert, U.D. CD133-Functionalized Gold Nanoparticles as a Carrier Platform for Telaglenastat (CB-839) against Tumor Stem Cells. Int. J. Mol. Sci. 2022, 23, 5479. [Google Scholar] [CrossRef] [PubMed]
- Lundy, D.J.; Nguyễn, H.; Hsieh, P.C.H. Emerging Nano-Carrier Strategies for Brain Tumor Drug Delivery and Considerations for Clinical Translation. Pharmaceutics 2021, 13, 1193. [Google Scholar] [CrossRef]
- Mirzaei, S.; Mahabady, M.K.; Zabolian, A.; Abbaspour, A.; Fallahzadeh, P.; Noori, M.; Hashemi, F.; Hushmandi, K.; Daneshi, S.; Kumar, A.P.; et al. Small Interfering RNA (SiRNA) to Target Genes and Molecular Pathways in Glioblastoma Therapy: Current Status with an Emphasis on Delivery Systems. Life Sci. 2021, 275, 119368. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Tao, W.; Zou, Y.; Farokhzad, O.C.; Shi, B. Nanotechnology-Based Strategies for SiRNA Brain Delivery for Disease Therapy. Trends Biotechnol. 2018, 36, 562–575. [Google Scholar] [CrossRef]
- Liu, Z.; Yan, H.; Li, H. Silencing of DNA Repair Sensitizes Pediatric Brain Tumor Cells to γ-Irradiation Using Gold Nanoparticles. Environ. Toxicol. Pharmacol. 2017, 53, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Cohen, Z.R.; Ramishetti, S.; Peshes-Yaloz, N.; Goldsmith, M.; Wohl, A.; Zibly, Z.; Peer, D. Localized RNAi Therapeutics of Chemoresistant Grade IV Glioma Using Hyaluronan-Grafted Lipid-Based Nanoparticles. ACS Nano 2015, 9, 1581–1591. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Khan, O.F.; Suvà, M.L.; Dong, B.; Panek, W.K.; Xiao, T.; Wu, M.; Han, Y.; Ahmed, A.U.; Balyasnikova, I.V.; et al. Multiplexed RNAi Therapy against Brain Tumor-Initiating Cells via Lipopolymeric Nanoparticle Infusion Delays Glioblastoma Progression. Proc. Natl. Acad. Sci. USA 2017, 114, E6147–E6156. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Deng, X.; Sheng, H.; Rong, Y.; Zheng, Y.; Zhang, Y.; Lin, J. Noncoding RNAs in Pediatric Brain Tumors: Molecular Functions and Pathological Implications. Mol. Ther. Nucleic Acids 2021, 26, 417–431. [Google Scholar] [CrossRef] [PubMed]
- Braoudaki, M.; Lambrou, G.I.; Giannikou, K.; Papadodima, S.A.; Lykoudi, A.; Stefanaki, K.; Sfakianos, G.; Kolialexi, A.; Tzortzatou-Stathopoulou, F.; Tzetis, M.; et al. MiR-15a and MiR-24-1 as Putative Prognostic MicroRNA Signatures for Pediatric Pilocytic Astrocytomas and Ependymomas. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2016, 37, 9887–9897. [Google Scholar] [CrossRef]
- Tantawy, M.; Elzayat, M.G.; Yehia, D.; Taha, H. Identification of MicroRNA Signature in Different Pediatric Brain Tumors. Genet. Mol. Biol. 2018, 41, 27–34. [Google Scholar] [CrossRef]
- Catanzaro, G.; Besharat, Z.M.; Carai, A.; Jäger, N.; Splendiani, E.; Colin, C.; Po, A.; Chiacchiarini, M.; Citarella, A.; Gianno, F.; et al. MiR-1248: A New Prognostic Biomarker Able to Identify Supratentorial Hemispheric Pediatric Low-Grade Gliomas Patients Associated with Progression. Biomark. Res. 2022, 10, 44. [Google Scholar] [CrossRef]
- Sanchez-Diaz, P.C.; Hsiao, T.-H.; Chang, J.C.; Yue, D.; Tan, M.C.; Chen, H.-I.H.; Tomlinson, G.E.; Huang, Y.; Chen, Y.; Hung, J.Y. De-Regulated MicroRNAs in Pediatric Cancer Stem Cells Target Pathways Involved in Cell Proliferation, Cell Cycle and Development. PLoS ONE 2013, 8, e61622. [Google Scholar] [CrossRef]
- Abdelfattah, N.; Rajamanickam, S.; Panneerdoss, S.; Timilsina, S.; Yadav, P.; Onyeagucha, B.C.; Garcia, M.; Vadlamudi, R.; Chen, Y.; Brenner, A.; et al. MiR-584-5p Potentiates Vincristine and Radiation Response by Inducing Spindle Defects and DNA Damage in Medulloblastoma. Nat. Commun. 2018, 9, 4541. [Google Scholar] [CrossRef]
- Ferretti, E.; De Smaele, E.; Miele, E.; Laneve, P.; Po, A.; Pelloni, M.; Paganelli, A.; Di Marcotullio, L.; Caffarelli, E.; Screpanti, I.; et al. Concerted MicroRNA Control of Hedgehog Signalling in Cerebellar Neuronal Progenitor and Tumour Cells. EMBO J. 2008, 27, 2616–2627. [Google Scholar] [CrossRef]
- Miele, E.; Po, A.; Mastronuzzi, A.; Carai, A.; Besharat, Z.M.; Pediconi, N.; Abballe, L.; Catanzaro, G.; Sabato, C.; De Smaele, E.; et al. Downregulation of MiR-326 and Its Host Gene β-Arrestin1 Induces pro-Survival Activity of E2F1 and Promotes Medulloblastoma Growth. Mol. Oncol. 2021, 15, 523–542. [Google Scholar] [CrossRef]
- Andolfo, I.; Liguori, L.; De Antonellis, P.; Cusanelli, E.; Marinaro, F.; Pistollato, F.; Garzia, L.; De Vita, G.; Petrosino, G.; Accordi, B.; et al. The Micro-RNA 199b-5p Regulatory Circuit Involves Hes1, CD15, and Epigenetic Modifications in Medulloblastoma. Neuro Oncology 2012, 14, 596–612. [Google Scholar] [CrossRef] [PubMed]
- Catanzaro, G.; Sabato, C.; Russo, M.; Rosa, A.; Abballe, L.; Besharat, Z.M.; Po, A.; Miele, E.; Bellavia, D.; Chiacchiarini, M.; et al. Loss of MiR-107, MiR-181c and MiR-29a-3p Promote Activation of Notch2 Signaling in Pediatric High-Grade Gliomas (PHGGs). Int. J. Mol. Sci. 2017, 18, 2742. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.-L.; Hsieh, T.-H.; Ng, K.-H.; Tsai, Y.-N.; Tsai, C.-F.; Chao, M.-E.; Liu, D.-J.; Chu, S.-S.; Chen, W.; Liu, Y.-R.; et al. Downregulation of MiR-137 and MiR-6500-3p Promotes Cell Proliferation in Pediatric High-Grade Gliomas. Oncotarget 2016, 7, 19723–19737. [Google Scholar] [CrossRef]
- Catanzaro, G.; Besharat, Z.M.; Miele, E.; Chiacchiarini, M.; Po, A.; Carai, A.; Marras, C.E.; Antonelli, M.; Badiali, M.; Raso, A.; et al. The MiR-139-5p Regulates Proliferation of Supratentorial Paediatric Low-Grade Gliomas by Targeting the PI3K/AKT/MTORC1 Signalling. Neuropathol. Appl. Neurobiol. 2018, 44, 687–706. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Bertoni, H.; Kozielski, K.L.; Rui, Y.; Lal, B.; Vaughan, H.; Wilson, D.R.; Mihelson, N.; Eberhart, C.G.; Laterra, J.; Green, J.J. Bioreducible Polymeric Nanoparticles Containing Multiplexed Cancer Stem Cell Regulating MiRNAs Inhibit Glioblastoma Growth and Prolong Survival. Nano Lett. 2018, 18, 4086–4094. [Google Scholar] [CrossRef] [PubMed]
- Sukumar, U.K.; Bose, R.J.C.; Malhotra, M.; Babikir, H.A.; Afjei, R.; Robinson, E.; Zeng, Y.; Chang, E.; Habte, F.; Sinclair, R.; et al. Intranasal Delivery of Targeted Polyfunctional Gold-Iron Oxide Nanoparticles Loaded with Therapeutic MicroRNAs for Combined Theranostic Multimodality Imaging and Presensitization of Glioblastoma to Temozolomide. Biomaterials 2019, 218, 119342. [Google Scholar] [CrossRef]
- Poot, E.; Maguregui, A.; Brunton, V.G.; Sieger, D.; Hulme, A.N. Targeting Glioblastoma through Nano- and Micro-Particle-Mediated Immune Modulation. Bioorg. Med. Chem. 2022, 72, 116913. [Google Scholar] [CrossRef]
- Guido, C.; Baldari, C.; Maiorano, G.; Mastronuzzi, A.; Carai, A.; Quintarelli, C.; De Angelis, B.; Cortese, B.; Gigli, G.; Palamà, I.E. Nanoparticles for Diagnosis and Target Therapy in Pediatric Brain Cancers. Diagnostics 2022, 12, 173. [Google Scholar] [CrossRef]
- Galstyan, A.; Markman, J.L.; Shatalova, E.S.; Chiechi, A.; Korman, A.J.; Patil, R.; Klymyshyn, D.; Tourtellotte, W.G.; Israel, L.L.; Braubach, O.; et al. Blood-Brain Barrier Permeable Nano Immunoconjugates Induce Local Immune Responses for Glioma Therapy. Nat. Commun. 2019, 10, 3850. [Google Scholar] [CrossRef]
- Meng, L.; Wang, C.; Lu, Y.; Sheng, G.; Yang, L.; Wu, Z.; Xu, H.; Han, C.; Lu, Y.; Han, F. Targeted Regulation of Blood-Brain Barrier for Enhanced Therapeutic Efficiency of Hypoxia-Modifier Nanoparticles and Immune Checkpoint Blockade Antibodies for Glioblastoma. ACS Appl. Mater. Interfaces 2021, 13, 11657–11671. [Google Scholar] [CrossRef] [PubMed]
- Voth, B.L.; Pelargos, P.E.; Barnette, N.E.; Bhatt, N.S.; Chen, C.H.J.; Lagman, C.; Chung, L.K.; Nguyen, T.; Sheppard, J.P.; Romiyo, P.; et al. Intratumor Injection of CCL21-Coupled Vault Nanoparticles Is Associated with Reduction in Tumor Volume in an in Vivo Model of Glioma. J. Neurooncol. 2020, 147, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Parayath, N.N.; Ene, C.I.; Stephan, S.B.; Koehne, A.L.; Coon, M.E.; Holland, E.C.; Stephan, M.T. Genetic Programming of Macrophages to Perform Anti-Tumor Functions Using Targeted MRNA Nanocarriers. Nat. Commun. 2019, 10, 3974. [Google Scholar] [CrossRef] [PubMed]
- Azambuja, J.H.; Schuh, R.S.; Michels, L.R.; Gelsleichter, N.E.; Beckenkamp, L.R.; Iser, I.C.; Lenz, G.S.; de Oliveira, F.H.; Venturin, G.; Greggio, S.; et al. Nasal Administration of Cationic Nanoemulsions as CD73-SiRNA Delivery System for Glioblastoma Treatment: A New Therapeutical Approach. Mol. Neurobiol. 2020, 57, 635–649. [Google Scholar] [CrossRef]
- Kadiyala, P.; Gregory, J.V.; Lowenstein, P.R.; Lahann, J.; Castro, M.G. Targeting Gliomas with STAT3-Silencing Nanoparticles. Mol. Cell. Oncol. 2021, 8, 1870647. [Google Scholar] [CrossRef]
- Lenzen, A.; Cole, L.; Lauing, K.L.; Zhai, L.; Ladomersky, E.; Lulla, R.R.; Hashizume, R.; Stegh, A.; Wainwright, D.A. Immu-24. Immunotherapeutic Nanotechnology Targeting Ido1 for Pediatric Diffuse Intrinsic Pontine Glioma. Neuro Oncol. 2018, 20, i103. [Google Scholar] [CrossRef]
- Sayour, E.; Mendez-Gomez, H.; Grippin, A.; De Leon, G.; Stover, B.; Flores, C.; Pham, C.; Mitchell, D. Mbrs-02. Personalized Immunotherapy with Translatable Rna Nanoparticles Targeting Medulloblastoma. Neuro Oncol. 2018, 20, i128–i129. [Google Scholar] [CrossRef]
- Mendez-Gomez, H.; McGuiness, J.; Grippin, A.; Weidert, F.; Carrera-Justiz, S.; Mitchell, D.; Sayour, E. Immu-13. Customizable Multi-Lamellar Rna-Nanoparticles for Pediatric Glioma. Neuro Oncol. 2021, 23, i29–i30. [Google Scholar] [CrossRef]
- Perini, G.; Giulimondi, F.; Palmieri, V.; Augello, A.; Digiacomo, L.; Quagliarini, E.; Pozzi, D.; Papi, M.; Caracciolo, G. Inhibiting the Growth of 3D Brain Cancer Models with Bio-Coronated Liposomal Temozolomide. Pharmaceutics 2021, 13, 378. [Google Scholar] [CrossRef]
- Straehla, J.P.; Hajal, C.; Safford, H.C.; Offeddu, G.S.; Boehnke, N.; Dacoba, T.G.; Wyckoff, J.; Kamm, R.D.; Hammond, P.T. A Predictive Microfluidic Model of Human Glioblastoma to Assess Trafficking of Blood-Brain Barrier-Penetrant Nanoparticles. Proc. Natl. Acad. Sci. USA 2022, 119, e2118697119. [Google Scholar] [CrossRef]
- Jiang, Y.; Yang, W.; Zhang, J.; Meng, F.; Zhong, Z. Protein Toxin Chaperoned by LRP-1-Targeted Virus-Mimicking Vesicles Induces High-Efficiency Glioblastoma Therapy In Vivo. Adv. Mater. Deerfield Beach Fla 2018, 30, e1800316. [Google Scholar] [CrossRef] [PubMed]
- Zanganeh, S.; Hutter, G.; Spitler, R.; Lenkov, O.; Mahmoudi, M.; Shaw, A.; Pajarinen, J.S.; Nejadnik, H.; Goodman, S.; Moseley, M.; et al. Iron Oxide Nanoparticles Inhibit Tumour Growth by Inducing Pro-Inflammatory Macrophage Polarization in Tumour Tissues. Nat. Nanotechnol. 2016, 11, 986–994. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Liu, T.; Li, Y.; Lau, J.; Yang, Z.; Fan, W.; Zhou, Z.; Shi, C.; Ke, C.; Bregadze, V.I.; et al. Fenton-Reaction-Acceleratable Magnetic Nanoparticles for Ferroptosis Therapy of Orthotopic Brain Tumors. ACS Nano 2018, 12, 11355–11365. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.W.; Tyler, B.M.; Masi, B.C.; Upadhyay, U.M.; Patta, Y.R.; Grossman, R.; Basaldella, L.; Langer, R.S.; Brem, H.; Cima, M.J. Intracranial Microcapsule Drug Delivery Device for the Treatment of an Experimental Gliosarcoma Model. Biomaterials 2011, 32, 2532–2539. [Google Scholar] [CrossRef] [PubMed]
- Baltes, S.; Freund, I.; Lewis, A.L.; Nolte, I.; Brinker, T. Doxorubicin and Irinotecan Drug-Eluting Beads for Treatment of Glioma: A Pilot Study in a Rat Model. J. Mater. Sci. Mater. Med. 2010, 21, 1393–1402. [Google Scholar] [CrossRef]
- Wessel, K.M.; Kaplan, R.N. Targeting Tumor Microenvironment and Metastasis in Children with Solid Tumors. Curr. Opin. Pediatr. 2022, 34, 53–60. [Google Scholar] [CrossRef]
- Van Bree, N.F.H.N.; Wilhelm, M. The Tumor Microenvironment of Medulloblastoma: An Intricate Multicellular Network with Therapeutic Potential. Cancers 2022, 14, 5009. [Google Scholar] [CrossRef]
- Zhou, Z.; Wei, J.; Jiang, W. Characterization of Aging Tumor Microenvironment with Drawing Implications in Predicting the Prognosis and Immunotherapy Response in Low-Grade Gliomas. Sci. Rep. 2022, 12, 5457. [Google Scholar] [CrossRef]
- Duhamel, M.; Drelich, L.; Wisztorski, M.; Aboulouard, S.; Gimeno, J.-P.; Ogrinc, N.; Devos, P.; Cardon, T.; Weller, M.; Escande, F.; et al. Spatial Analysis of the Glioblastoma Proteome Reveals Specific Molecular Signatures and Markers of Survival. Nat. Commun. 2022, 13, 6665. [Google Scholar] [CrossRef]
- Raju, G.S.R.; Pavitra, E.; Varaprasad, G.L.; Bandaru, S.S.; Nagaraju, G.P.; Farran, B.; Huh, Y.S.; Han, Y.-K. Nanoparticles Mediated Tumor Microenvironment Modulation: Current Advances and Applications. J. Nanobiotechnol. 2022, 20, 274. [Google Scholar] [CrossRef]




| Targeting Receptors | Type of NPs | Application | Target Cells | References |
|---|---|---|---|---|
| EGFR | NPs functionalized with Ang2 and EP-1 | Drug delivery | Endothelial cells of BBB (Ang2) and tumor cells (EP-1) | [42] |
| EGF-modified Au NP–Pc 4 | Delivery of photosensitizer silicon phthalocyanine | Tumor cells | [30] | |
| NPs conjugated to an EGFR antibody (Panitumumab/Vectibix) | Drug delivery | Tumor cells | [31] | |
| Magnetic NPs conjugated to an EGFR deletion mutant (EGFRvIII) antibody | Magnetic resonance imaging | Tumor cells | [34] | |
| TfR | Tf-conjugated nanoparticles | Drug delivery | Tumor cells | [53,54,55,56,57,58] |
| Tf-conjugated nanoparticles | Drug delivery | Glioma stem cells and non-stem cells | [47] | |
| Tf-conjugated nanoparticles | Drug delivery | Glioma stem cells and non-stem cells | [53,54,55,56,57,58] | |
| IGFR | NPs functionalized with anti-insulin receptor antibody 83-14 | Drug delivery | BBB | [51] |
| NPs functionalized with anti-insulin receptor monoclonal antibody (29B4) | Drug delivery | BBB | [52] | |
| Lipoproteins | Gold-liposome nanoparticles conjugated with ApoE and RVG | RNAi delivery | Tumor cells (ApoE and RVG) and brain endothelium (RVG) | [67] |
| Nano-LDL particles | Drug delivery | Tumor cells | [67] | |
| NPs conjugated to Angiopep-2 | Drug delivery | BBB and tumor cells | [58] | |
| High-density lipoprotein nanoparticles | Intrinsic activity | MB cells and stem cells | [67] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abballe, L.; Spinello, Z.; Antonacci, C.; Coppola, L.; Miele, E.; Catanzaro, G.; Miele, E. Nanoparticles for Drug and Gene Delivery in Pediatric Brain Tumors’ Cancer Stem Cells: Current Knowledge and Future Perspectives. Pharmaceutics 2023, 15, 505. https://doi.org/10.3390/pharmaceutics15020505
Abballe L, Spinello Z, Antonacci C, Coppola L, Miele E, Catanzaro G, Miele E. Nanoparticles for Drug and Gene Delivery in Pediatric Brain Tumors’ Cancer Stem Cells: Current Knowledge and Future Perspectives. Pharmaceutics. 2023; 15(2):505. https://doi.org/10.3390/pharmaceutics15020505
Chicago/Turabian StyleAbballe, Luana, Zaira Spinello, Celeste Antonacci, Lucia Coppola, Ermanno Miele, Giuseppina Catanzaro, and Evelina Miele. 2023. "Nanoparticles for Drug and Gene Delivery in Pediatric Brain Tumors’ Cancer Stem Cells: Current Knowledge and Future Perspectives" Pharmaceutics 15, no. 2: 505. https://doi.org/10.3390/pharmaceutics15020505