
Enhancing Immunogenicity of a Thermostable, Efficacious SARS-CoV-2 Vaccine Formulation through Oligomerization and Adjuvant Choice

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
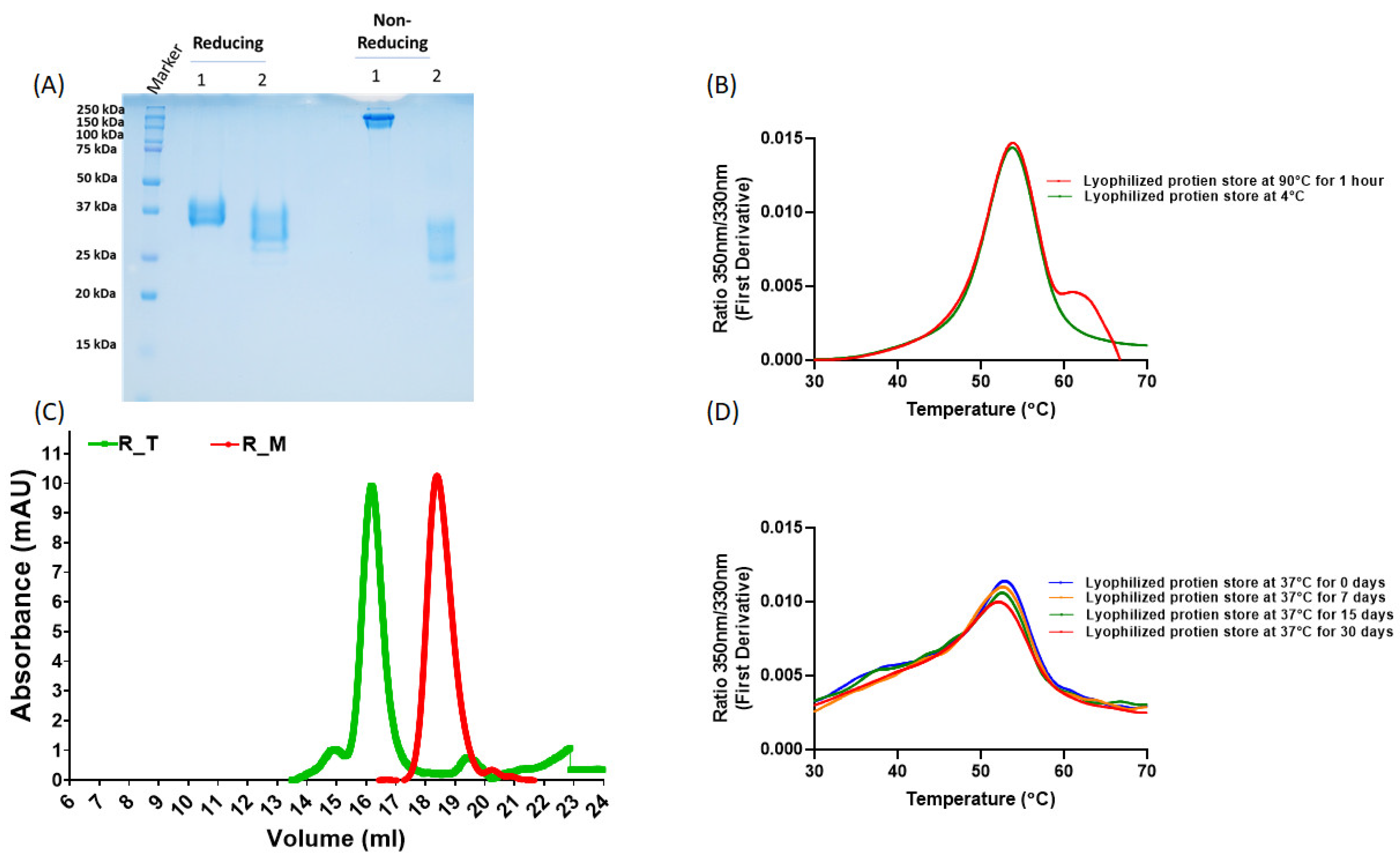
2.1. Protein Expression and Purification
2.2. NanoDSF Thermal Melt Studies
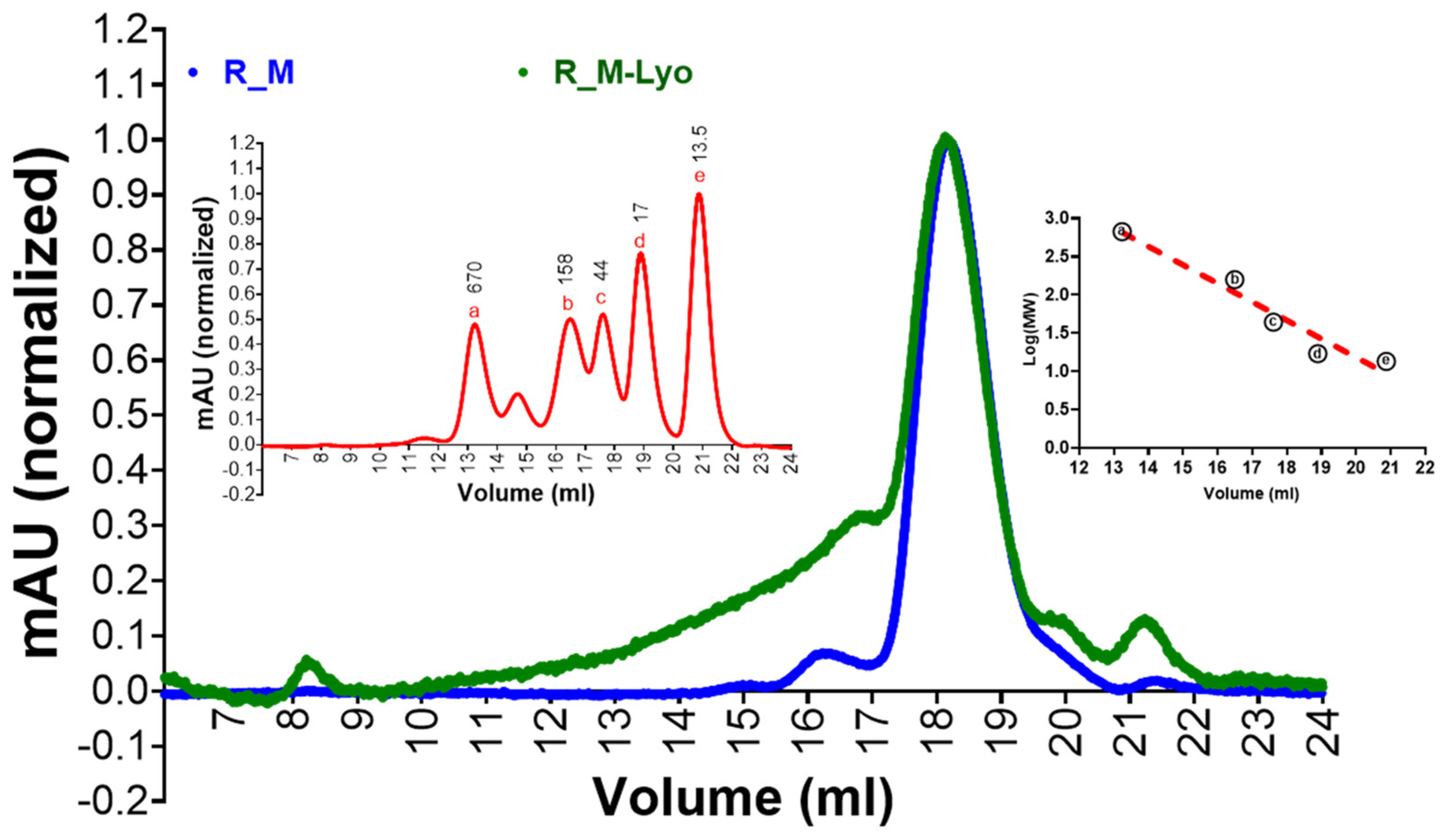
2.3. Size Exclusion Chromatography (SEC)
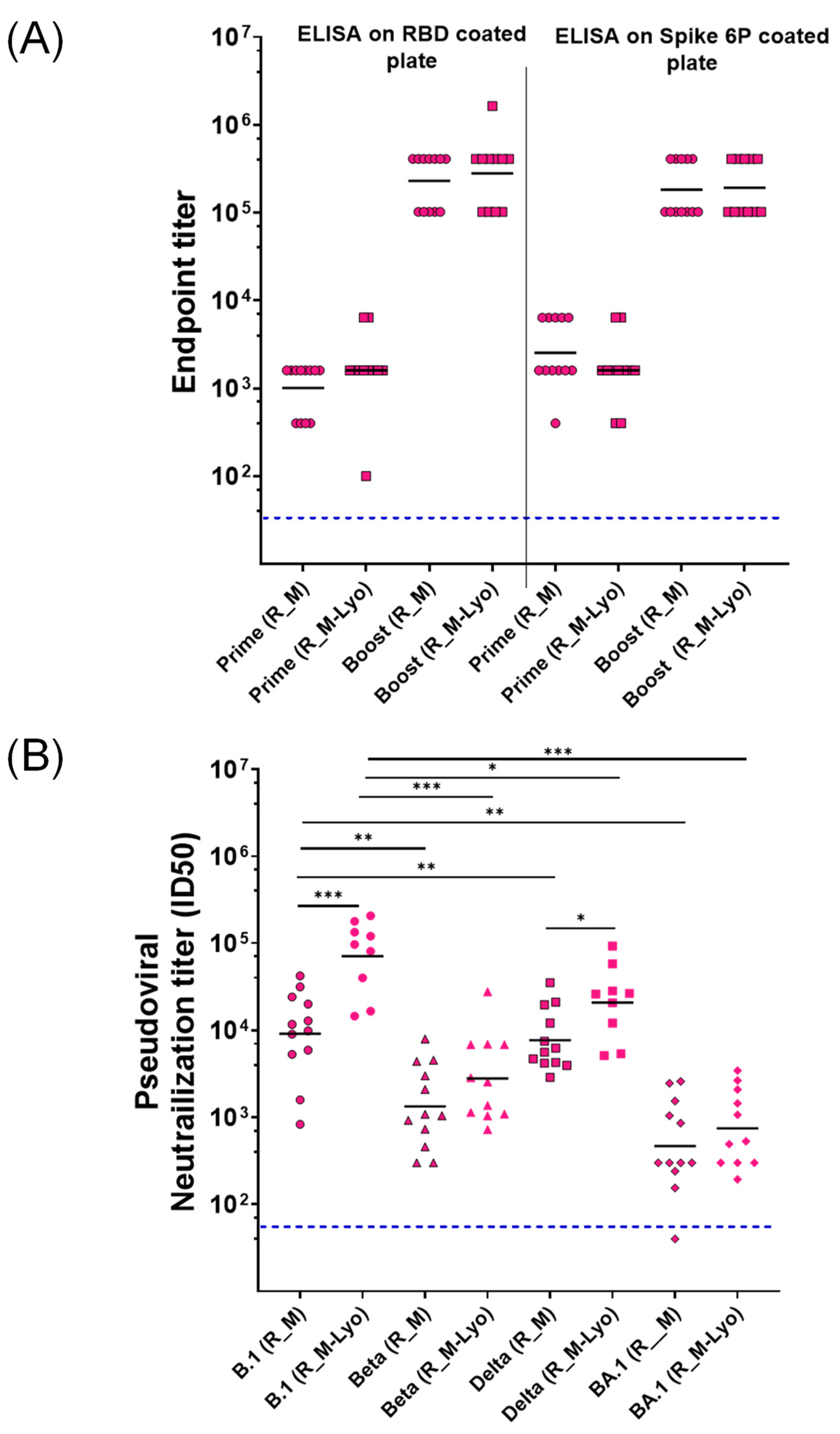
2.4. ELISA for Measurement of Serum Binding Antibody Endpoint Titers in hAce2 Expressing Transgenic Mice
2.5. SARS-CoV-2 Pseudovirus Preparation and Neutralization Assay
2.6. Adjuvant Preparation and Formulation Characterization
2.7. Mouse Immunization for Characterization of B- and T-Cell Responses
2.8. ICS Method for T-Cell Responses (IL-2, IFNγ and Th2 Cytokines IL-4, IL-5 and IL-13)
2.9. ELISA for Quantitating Antibody Responses in C57Bl/6 Mice
2.10. IFNγ Cytokine Quantification by ELISA
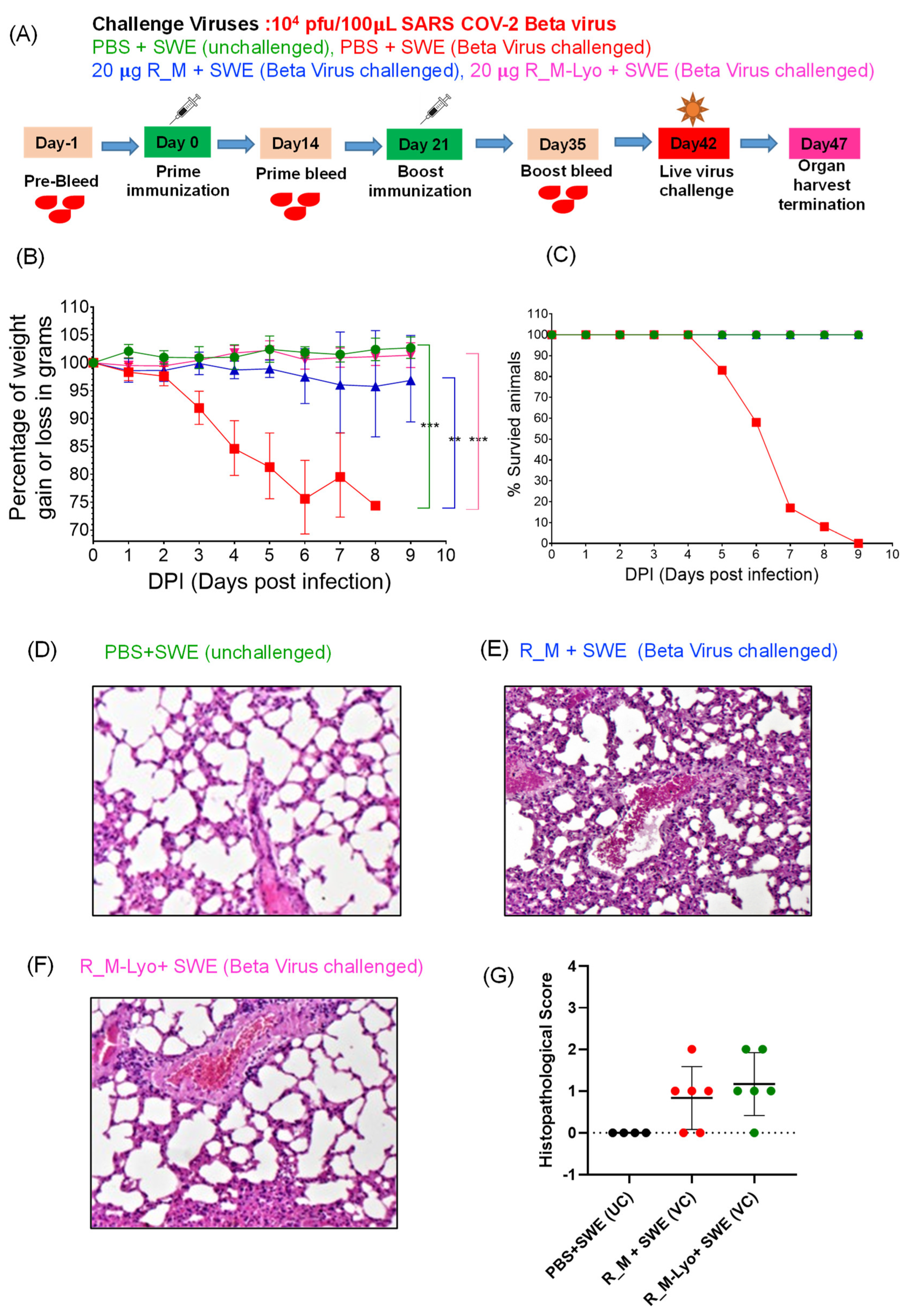
2.11. Challenge Studies
3. Results
3.1. Protein Expression and Characterization
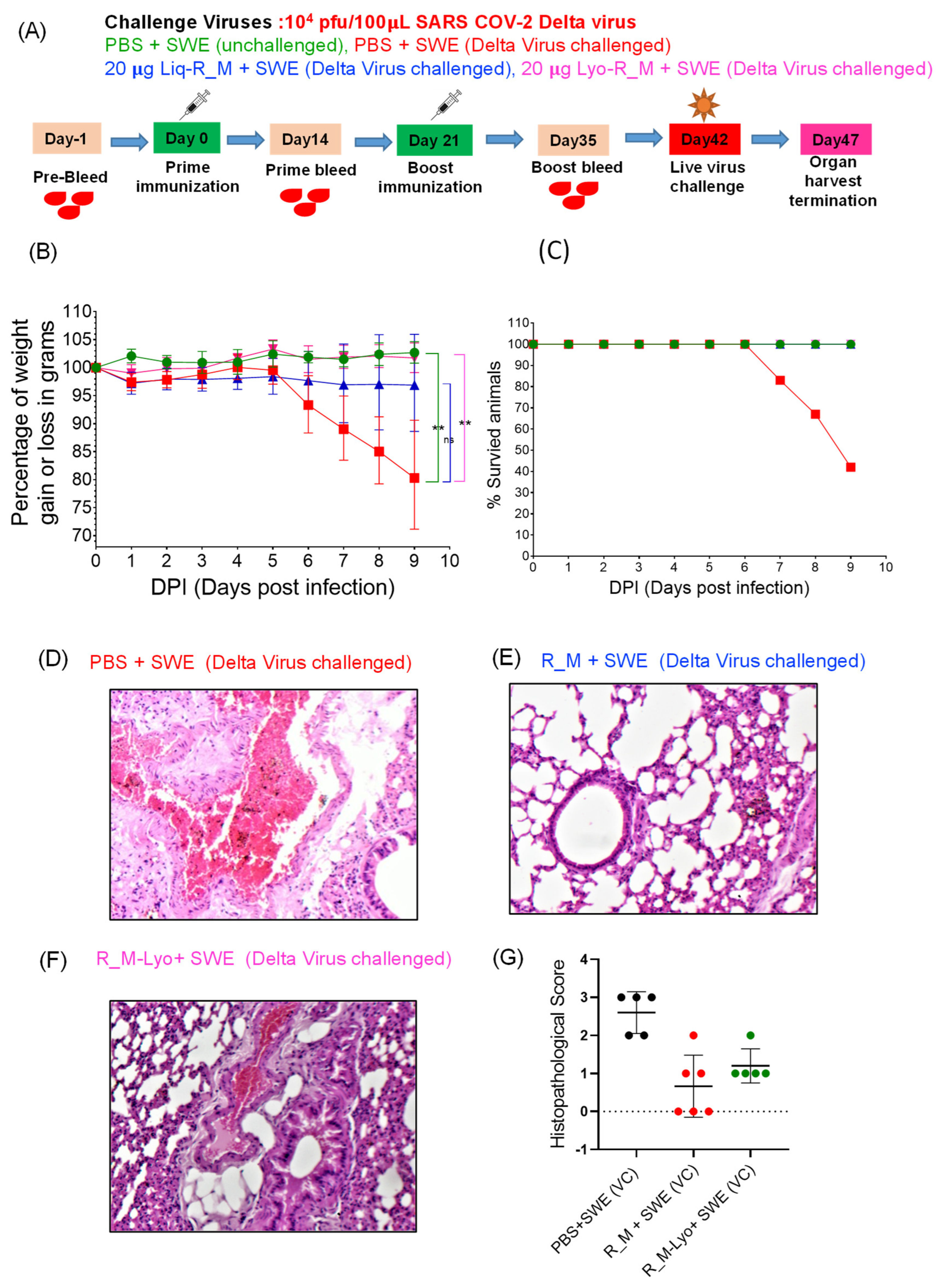
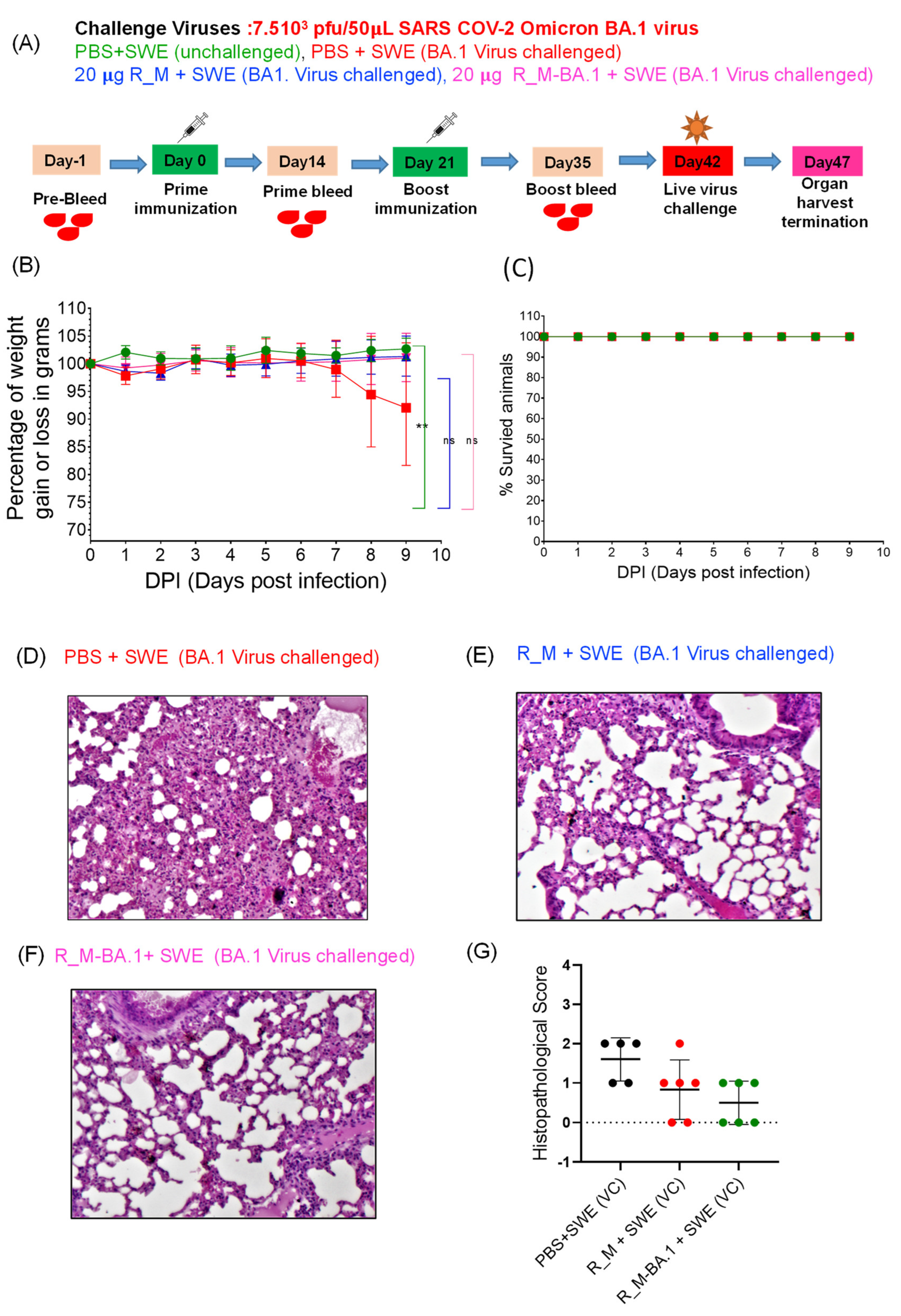
3.2. Immunization and Challenge Studies with R_M in hACE2 Expressing Transgenic Mice
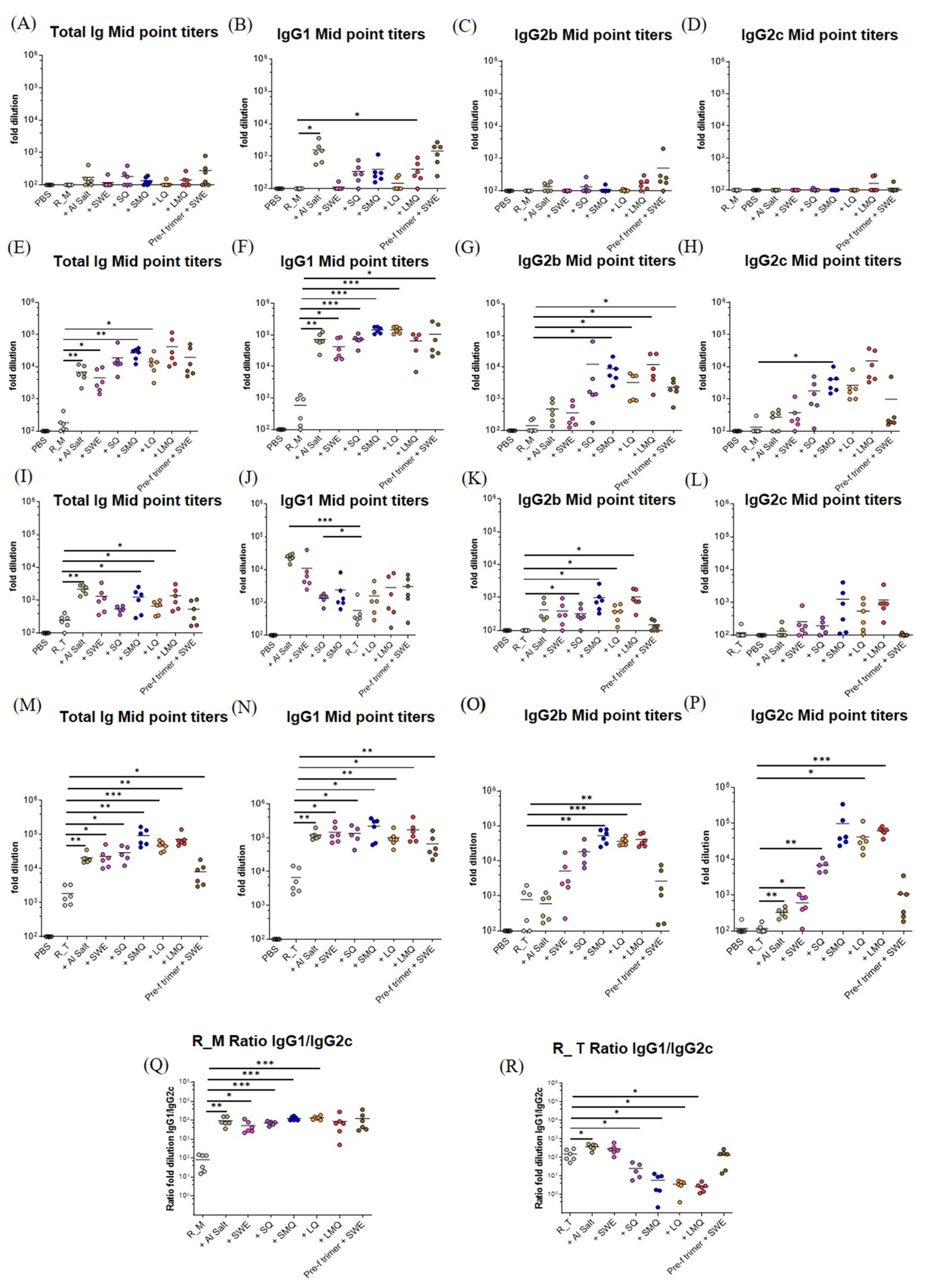
3.3. Potency and T Helper Bias of Adjuvanted, Formulated Material
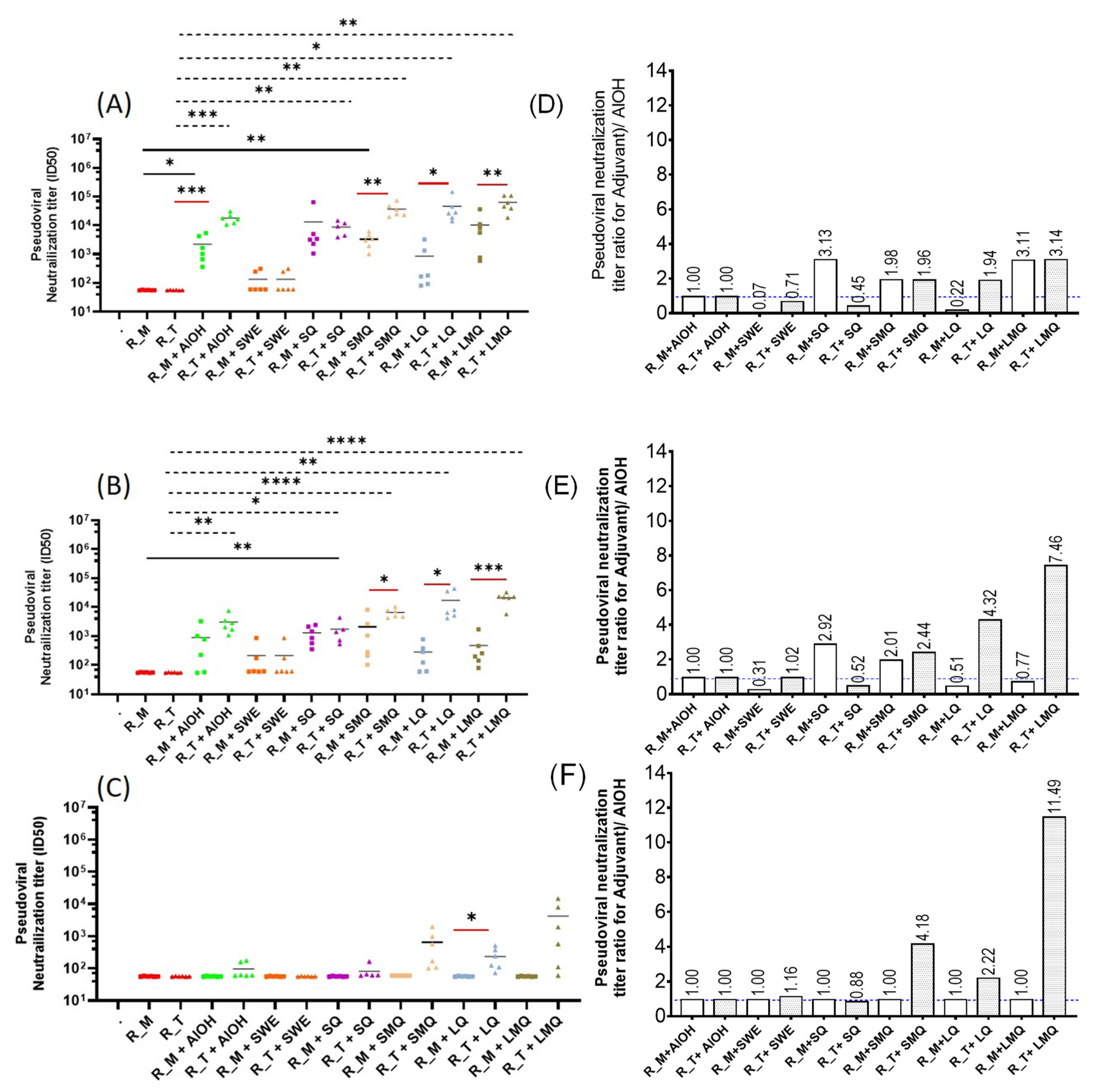
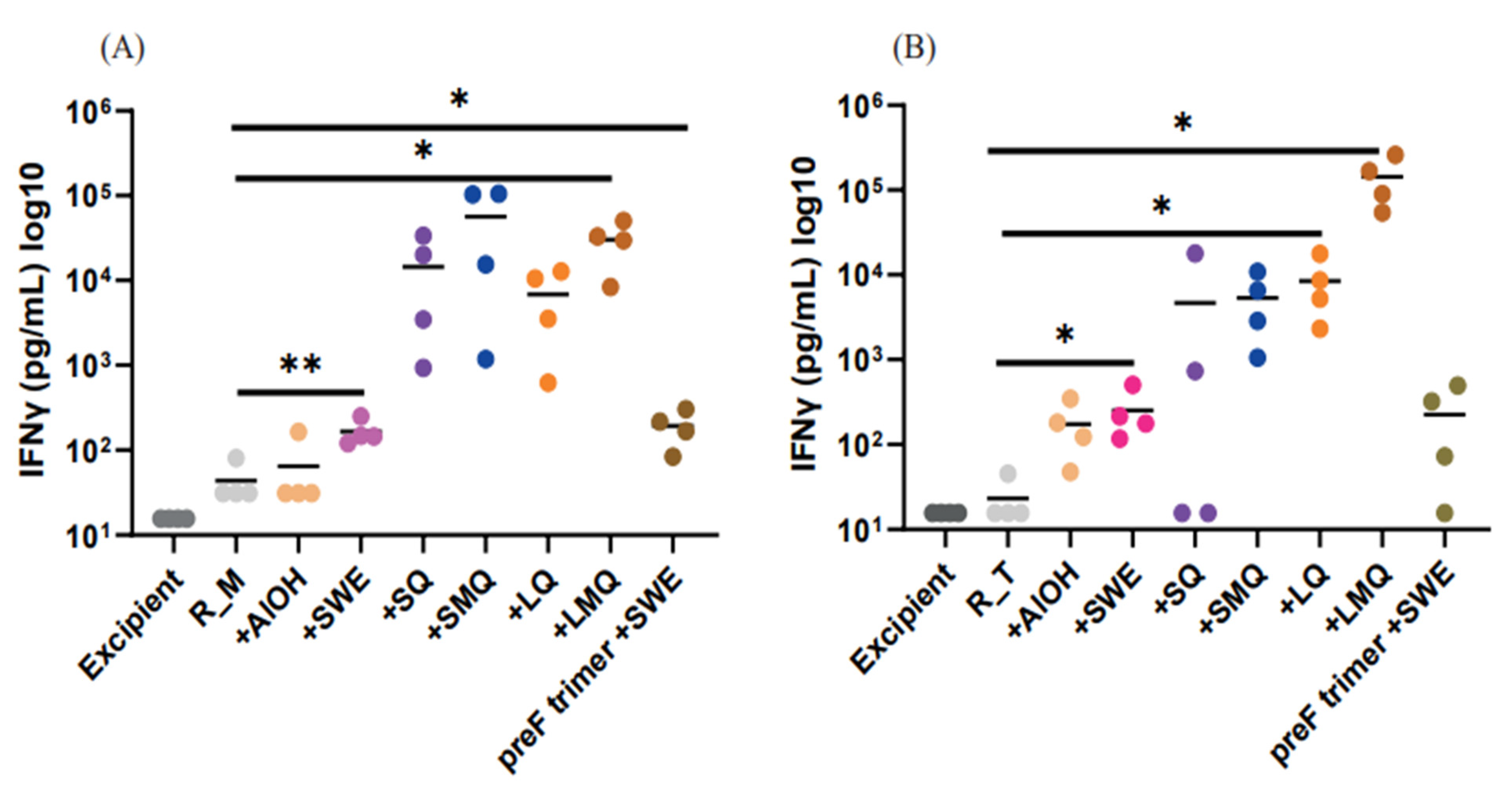
3.4. pVNT Results for R_M and R_T Immunogens with Sera in from C57BL/6 Mice
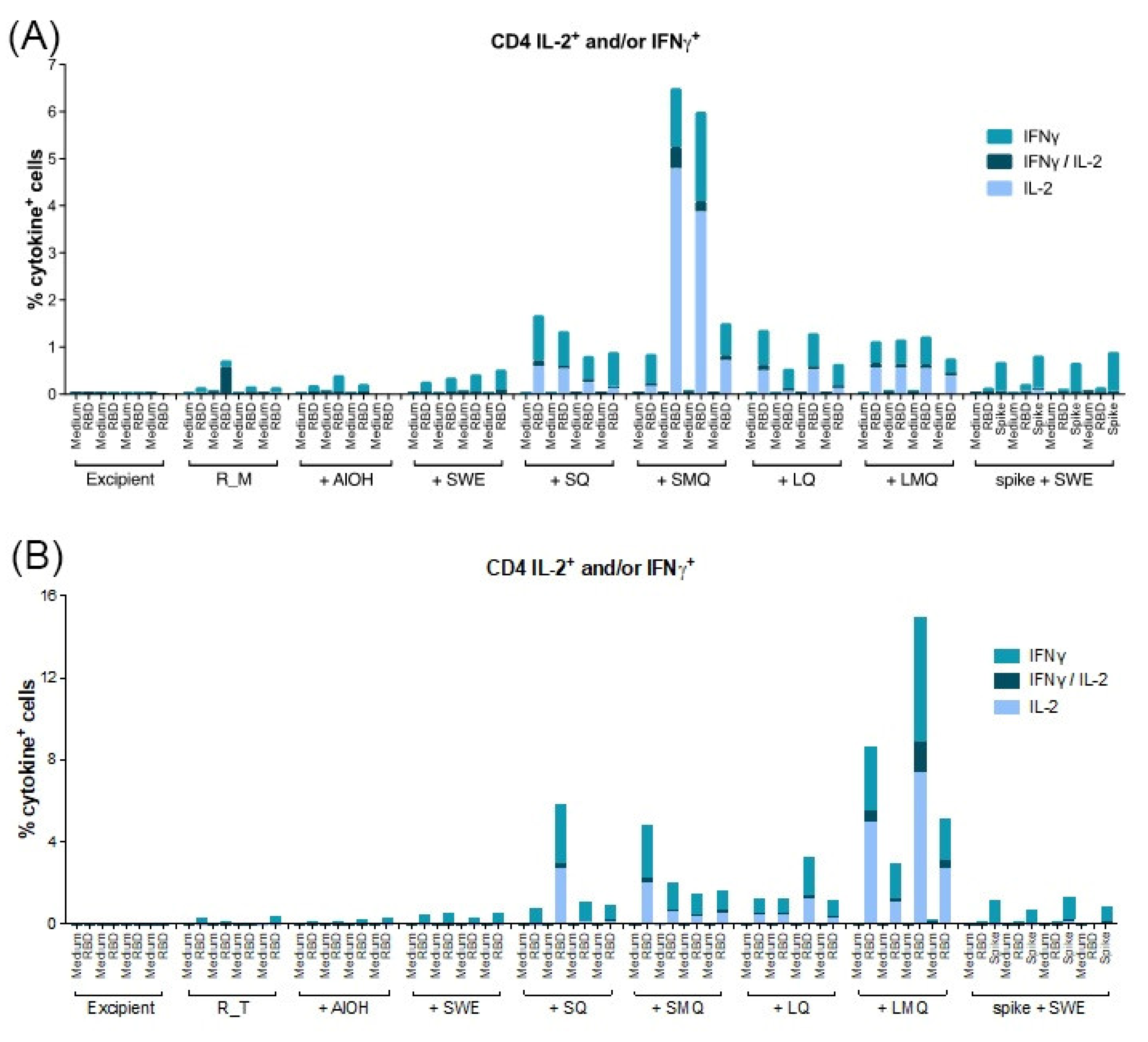
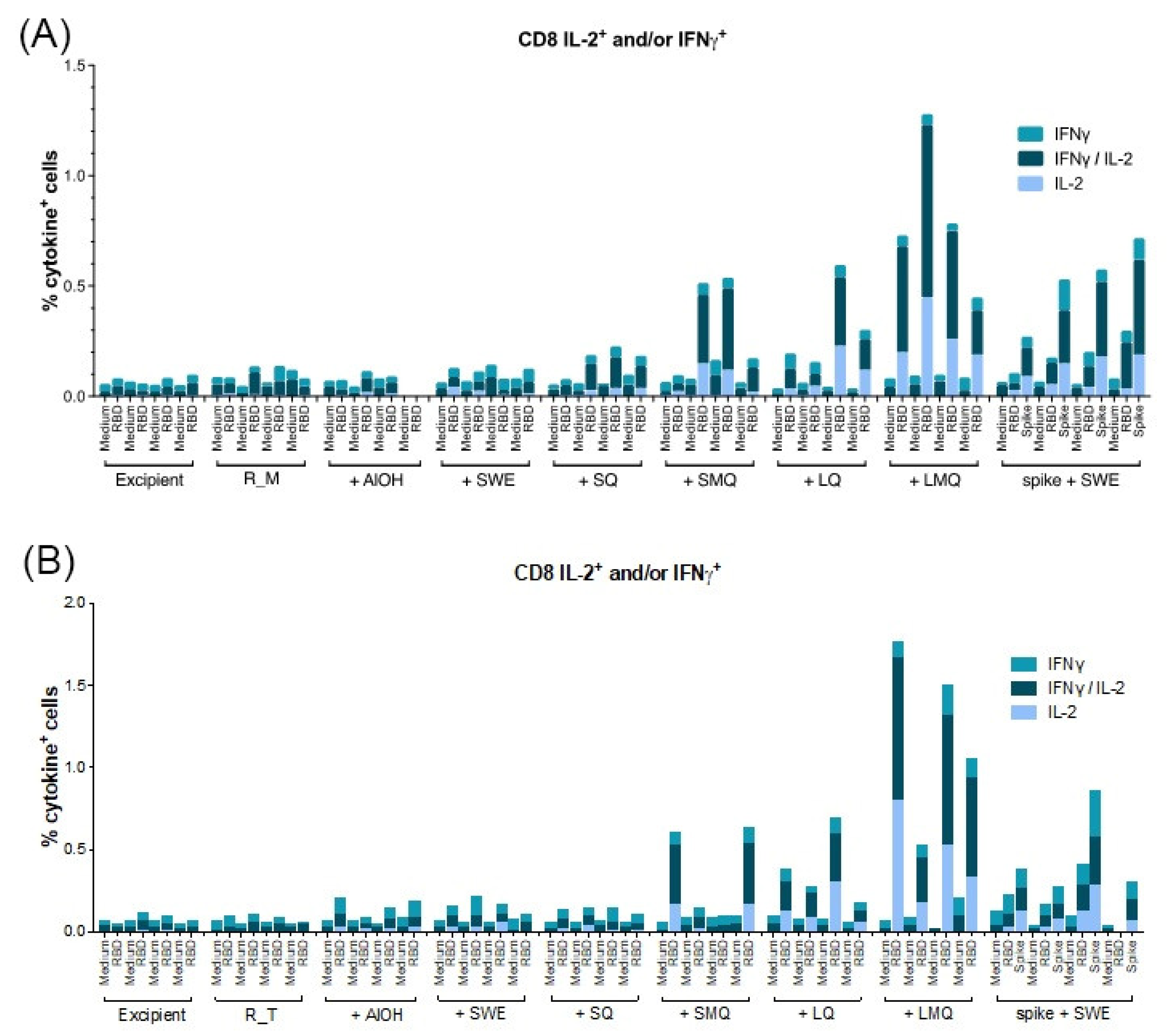
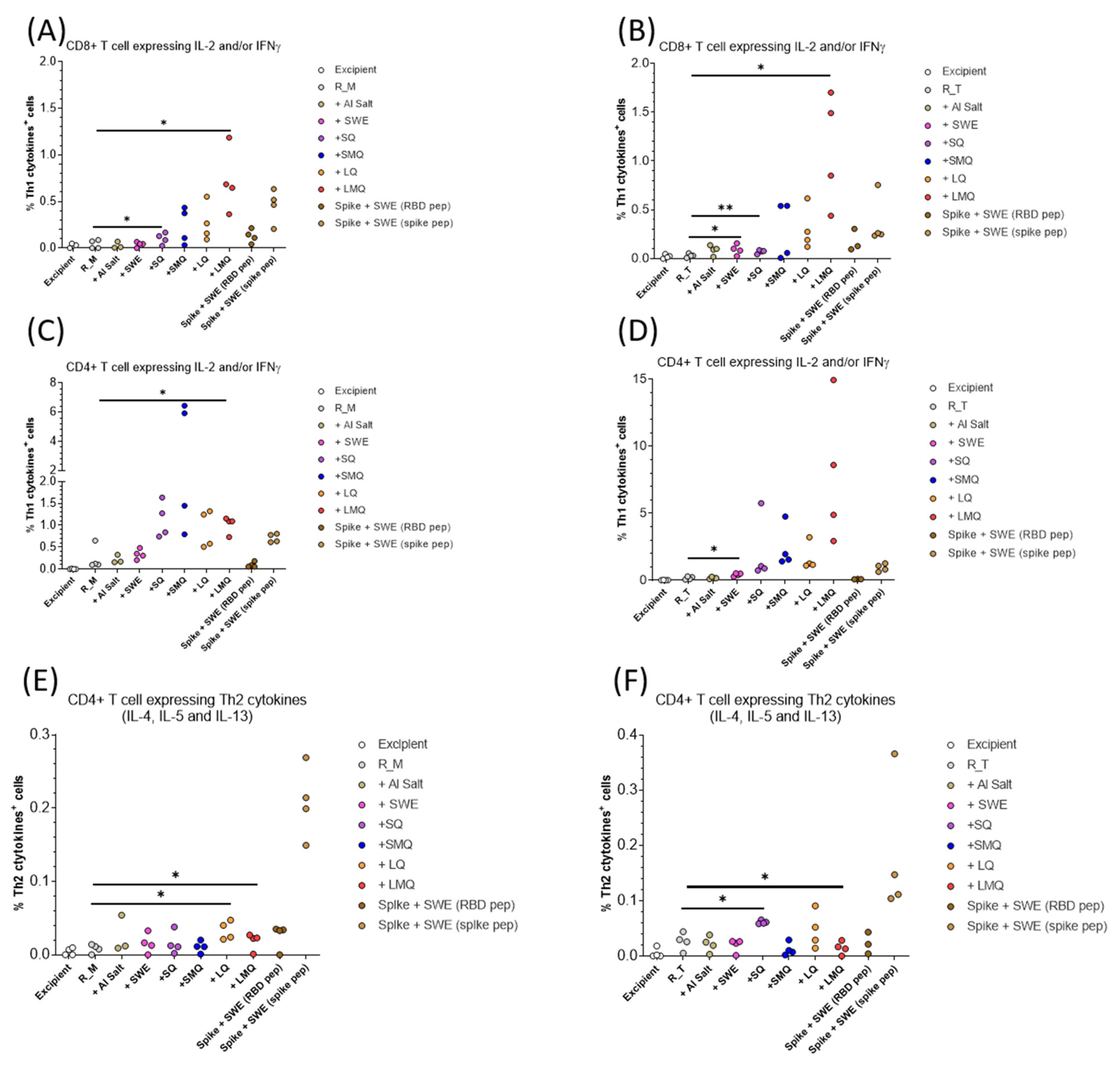
3.5. T-Cell Responses Elicited by R_M and R_T Formulations
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barnes, C.O.; Jette, C.A.; Abernathy, M.E.; Dam, K.-M.A.; Esswein, S.R.; Gristick, H.B.; Malyutin, A.G.; Sharaf, N.G.; Huey-Tubman, K.E.; Lee, Y.E.; et al. SARS-CoV-2 Neutralizing Antibody Structures Inform Therapeutic Strategies. Nature 2020, 588, 682–687. [Google Scholar] [CrossRef]
- Barnes, C.O.; West, A.P.; Huey-Tubman, K.E.; Hoffmann, M.A.G.; Sharaf, N.G.; Hoffman, P.R.; Koranda, N.; Gristick, H.B.; Gaebler, C.; Muecksch, F.; et al. Structures of Human Antibodies Bound to SARS-CoV-2 Spike Reveal Common Epitopes and Recurrent Features of Antibodies. Cell 2020, 182, 828–842.e16. [Google Scholar] [CrossRef]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.-L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM Structure of the 2019-NCoV Spike in the Prefusion Conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef] [PubMed]
- Zost, S.J.; Gilchuk, P.; Case, J.B.; Binshtein, E.; Chen, R.E.; Nkolola, J.P.; Schäfer, A.; Reidy, J.X.; Trivette, A.; Nargi, R.S.; et al. Potently Neutralizing and Protective Human Antibodies against SARS-CoV-2. Nature 2020, 584, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Zost, S.J.; Gilchuk, P.; Chen, R.E.; Case, J.B.; Reidy, J.X.; Trivette, A.; Nargi, R.S.; Sutton, R.E.; Suryadevara, N.; Chen, E.C.; et al. Rapid Isolation and Profiling of a Diverse Panel of Human Monoclonal Antibodies Targeting the SARS-CoV-2 Spike Protein. Nat. Med. 2020, 26, 1422–1427. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 Spike Receptor-Binding Domain Bound to the ACE2 Receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical Features of Patients Infected with 2019 Novel Coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Premkumar, L.; Segovia-Chumbez, B.; Jadi, R.; Martinez, D.R.; Raut, R.; Markmann, A.; Cornaby, C.; Bartelt, L.; Weiss, S.; Park, Y.; et al. The Receptor Binding Domain of the Viral Spike Protein is an Immunodominant and Highly Specific Target of Antibodies in SARS-CoV-2 Patients. Sci. Immunol. 2020, 5, eabc8413. [Google Scholar] [CrossRef]
- Yang, J.; Wang, W.; Chen, Z.; Lu, S.; Yang, F.; Bi, Z.; Bao, L.; Mo, F.; Li, X.; Huang, Y.; et al. A Vaccine Targeting the RBD of the S Protein of SARS-CoV-2 Induces Protective Immunity. Nature 2020, 586, 572–577. [Google Scholar] [CrossRef]
- Ahmed, S.; Khan, M.S.; Gayathri, S.; Singh, R.; Kumar, S.; Patel, U.R.; Malladi, S.K.; Rajmani, R.S.; van Vuren, P.J.; Riddell, S.; et al. A Stabilized, Monomeric, Receptor Binding Domain Elicits High-Titer Neutralizing Antibodies against All SARS-CoV-2 Variants of Concern. Front. Immunol. 2021, 12, 765211. [Google Scholar] [CrossRef]
- Jansen van Vuren, P.; McAuley, A.J.; Kuiper, M.J.; Singanallur, N.B.; Bruce, M.P.; Riddell, S.; Goldie, S.; Mangalaganesh, S.; Chahal, S.; Drew, T.W.; et al. Highly Thermotolerant SARS-CoV-2 Vaccine Elicits Neutralising Antibodies against Delta and Omicron in Mice. Viruses 2022, 14, 800. [Google Scholar] [CrossRef]
- Malladi, S.K.; Singh, R.; Pandey, S.; Gayathri, S.; Kanjo, K.; Ahmed, S.; Khan, M.S.; Kalita, P.; Girish, N.; Upadhyaya, A.; et al. Design of a Highly Thermotolerant, Immunogenic SARS-CoV-2 Spike Fragment. J. Biol. Chem. 2021, 296, 100025. [Google Scholar] [CrossRef]
- Malladi, S.K.; Patel, U.R.; Rajmani, R.S.; Singh, R.; Pandey, S.; Kumar, S.; Khaleeq, S.; van Vuren, P.J.; Riddell, S.; Goldie, S.; et al. Immunogenicity and Protective Efficacy of a Highly Thermotolerant, Trimeric SARS-CoV-2 Receptor Binding Domain Derivative. ACS Infect. Dis. 2021, 7, 2546–2564. [Google Scholar] [CrossRef]
- Grigoryan, L.; Lee, A.; Walls, A.C.; Lai, L.; Franco, B.; Arunachalam, P.S.; Feng, Y.; Luo, W.; Vanderheiden, A.; Floyd, K.; et al. Adjuvanting a Subunit SARS-CoV-2 Vaccine with Clinically Relevant Adjuvants Induces Durable Protection in Mice. NPJ Vaccines 2022, 7, 55. [Google Scholar] [CrossRef]
- Pulendran, B.; Arunachalam, P.S.; O’Hagan, D.T. Emerging Concepts in the Science of Vaccine Adjuvants. Nat. Rev. Drug Discov. 2021, 20, 454–475. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, J.S.; Isaacs, A.; Jakob, V.; Lebas, C.; Barnes, J.B.; Reading, P.C.; Young, P.R.; Watterson, D.; Dubois, P.M.; Collin, N.; et al. Characterization and Comparison of Novel Adjuvants for a Prefusion Clamped MERS Vaccine. Front. Immunol. 2022, 13, 976968. [Google Scholar] [CrossRef] [PubMed]
- Rapaka, R.R.; Cross, A.S.; Mcarthur, M.A. Using Adjuvants to Drive T Cell Responses for Next-Generation Infectious Disease Vaccines. Vaccines 2021, 9, 820. [Google Scholar] [CrossRef] [PubMed]
- Welsby, I.; Detienne, S.; Kuli, F.N.; Thomas, S.; Wouters, S.; Goriely, S. Lysosome-Dependent Activation of Human Dendritic Cells by the Vaccine Adjuvant QS-21. Front. Immunol. 2017, 7, 663. [Google Scholar] [CrossRef] [PubMed]
- Pifferi, C.; Fuentes, R.; Fernández-Tejada, A. Natural and synthetic carbohydrate-based vaccine adjuvants and their mechanisms of action. Nat. Rev. Chem. 2021, 5, 197–216. [Google Scholar] [CrossRef]
- Martin, R.M.; Brady, J.L.; Lew, A.M. The Need for IgG2c Specific Antiserum When Isotyping Antibodies from C57BL/6 and NOD Mice. J. Immunol. Methods 1998, 212, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Goldschmidt, T.; Salter, H. Possible Allelic Structure of IgG2a and IgG2c in Mice. Mol. Immunol. 2012, 50, 169–171. [Google Scholar] [CrossRef]
- Hashimoto, G.; Wright, P.F.; Karzon, D.T. Antibody-Dependent Cell-Mediated Cytotoxicity against Influenza Virus-Infected Cells. J. Infect. Dis. 1983, 148, 785–794. [Google Scholar] [CrossRef]
- Pollara, J.; Hart, L.; Brewer, F.; Pickeral, J.; Packard, B.Z.; Hoxie, J.A.; Komoriya, A.; Ochsenbauer, C.; Kappes, J.C.; Roederer, M.; et al. High-Throughput Quantitative Analysis of HIV-1 and SIV-Specific ADCC-Mediating Antibody Responses. Cytom. Part A 2011, 79, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Yeap, W.H.; Wong, K.L.; Shimasaki, N.; Chi, E.; Teo, Y. CD16 is Indispensable for Antibody- Dependent Cellular Cytotoxicity by Human Monocytes. Sci. Rep. 2016, 6, 34310. [Google Scholar] [CrossRef] [PubMed]
- Krüttgen, A.; Lauen, M.; Klingel, H.; Imöhl, M.; Kleines, M. Two Novel SARS-CoV-2 Surrogate Virus Neutralization Assays are Suitable for Assessing Successful Immunization with MRNA-1273. J. Virol. Methods 2022, 299, 114297. [Google Scholar] [CrossRef] [PubMed]
- Meyer, B.; Reimerink, J.; Torriani, G.; Brouwer, F.; Godeke, G.-J.; Yerly, S.; Hoogerwerf, M.; Vuilleumier, N.; Kaiser, L.; Eckerle, I.; et al. Validation and Clinical Evaluation of a SARS-CoV-2 Surrogate Virus Neutralisation Test (SVNT). Emerg. Microbes Infect. 2020, 9, 2394–2403. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, A.; Saito, T. CD4 CTL, a Cytotoxic Subset of CD4+ T Cells, Their Differentiation and Function. Front. Immunol. 2017, 8, 194. [Google Scholar] [CrossRef]
- Bennett, S.R.M.; Carbone, F.R.; Karamalis, F.; Flavell, R.A.; Miller, J.F.A.P. Heath WR: Help for cytotoxic-T-cell responses is mediated by CD40 signalling. Nature 1998, 393, 478–480. [Google Scholar] [CrossRef]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, M.S.; Jakob, V.; Singh, R.; Rajmani, R.S.; Kumar, S.; Lemoine, C.; Kleanthous, H.; Ringe, R.P.; Dubois, P.M.; Varadarajan, R. Enhancing Immunogenicity of a Thermostable, Efficacious SARS-CoV-2 Vaccine Formulation through Oligomerization and Adjuvant Choice. Pharmaceutics 2023, 15, 2759. https://doi.org/10.3390/pharmaceutics15122759
Khan MS, Jakob V, Singh R, Rajmani RS, Kumar S, Lemoine C, Kleanthous H, Ringe RP, Dubois PM, Varadarajan R. Enhancing Immunogenicity of a Thermostable, Efficacious SARS-CoV-2 Vaccine Formulation through Oligomerization and Adjuvant Choice. Pharmaceutics. 2023; 15(12):2759. https://doi.org/10.3390/pharmaceutics15122759
Chicago/Turabian StyleKhan, Mohammad Suhail, Virginie Jakob, Randhir Singh, Raju S. Rajmani, Sahil Kumar, Céline Lemoine, Harry Kleanthous, Rajesh P. Ringe, Patrice M. Dubois, and Raghavan Varadarajan. 2023. "Enhancing Immunogenicity of a Thermostable, Efficacious SARS-CoV-2 Vaccine Formulation through Oligomerization and Adjuvant Choice" Pharmaceutics 15, no. 12: 2759. https://doi.org/10.3390/pharmaceutics15122759