
Curcumin as a Stabilizer of Macrophage Polarization during Plasmodium Infection
 ,
,  , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Animals
2.2. Cell Culture and Polarization Treatments
2.3. Curcumin Concentrations
2.4. Cell Infection with Plasmodium berghei NK65 and Treatment with Curcumin
2.5. Phagocytosis and Microbicidal Activity
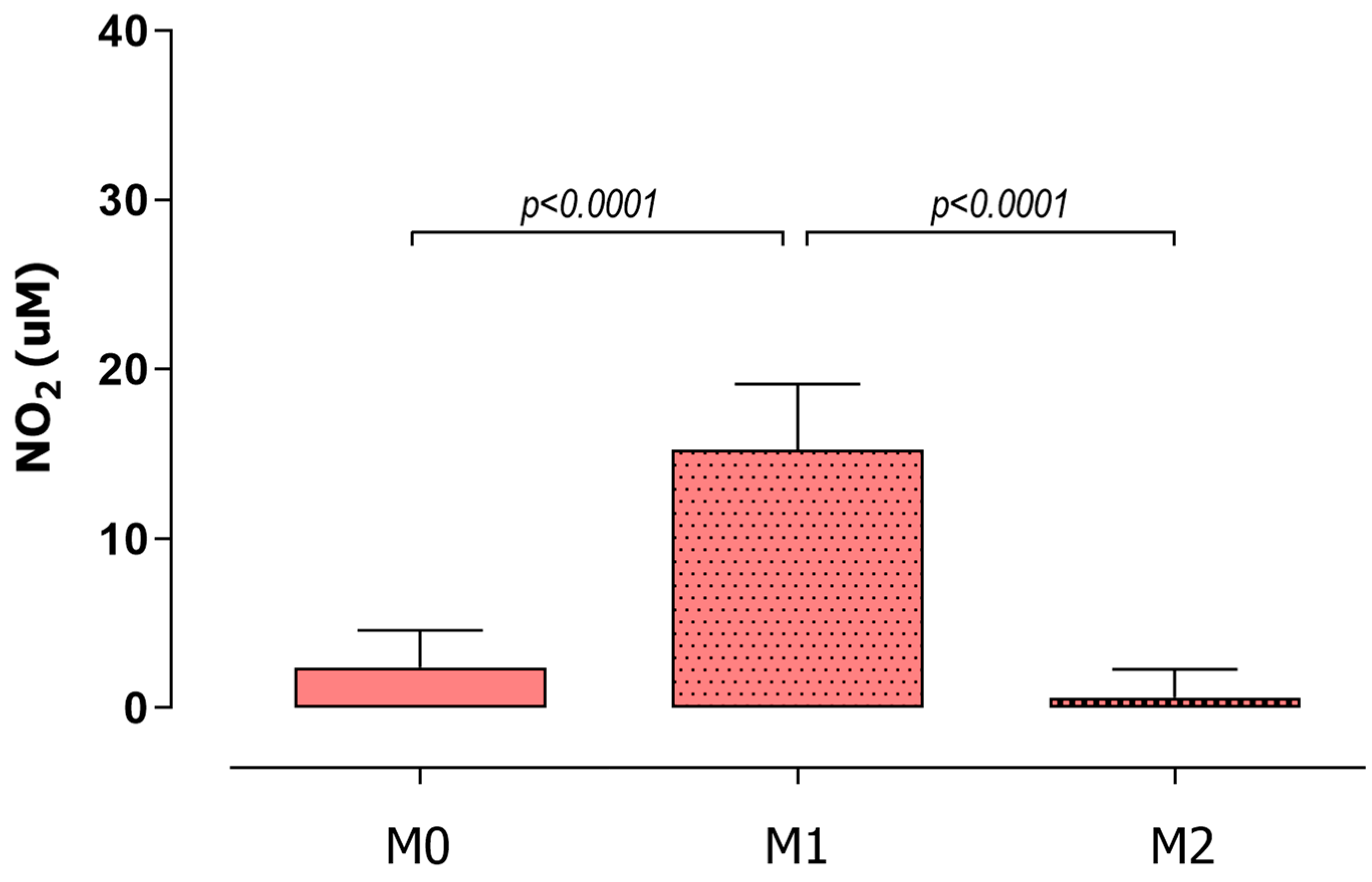
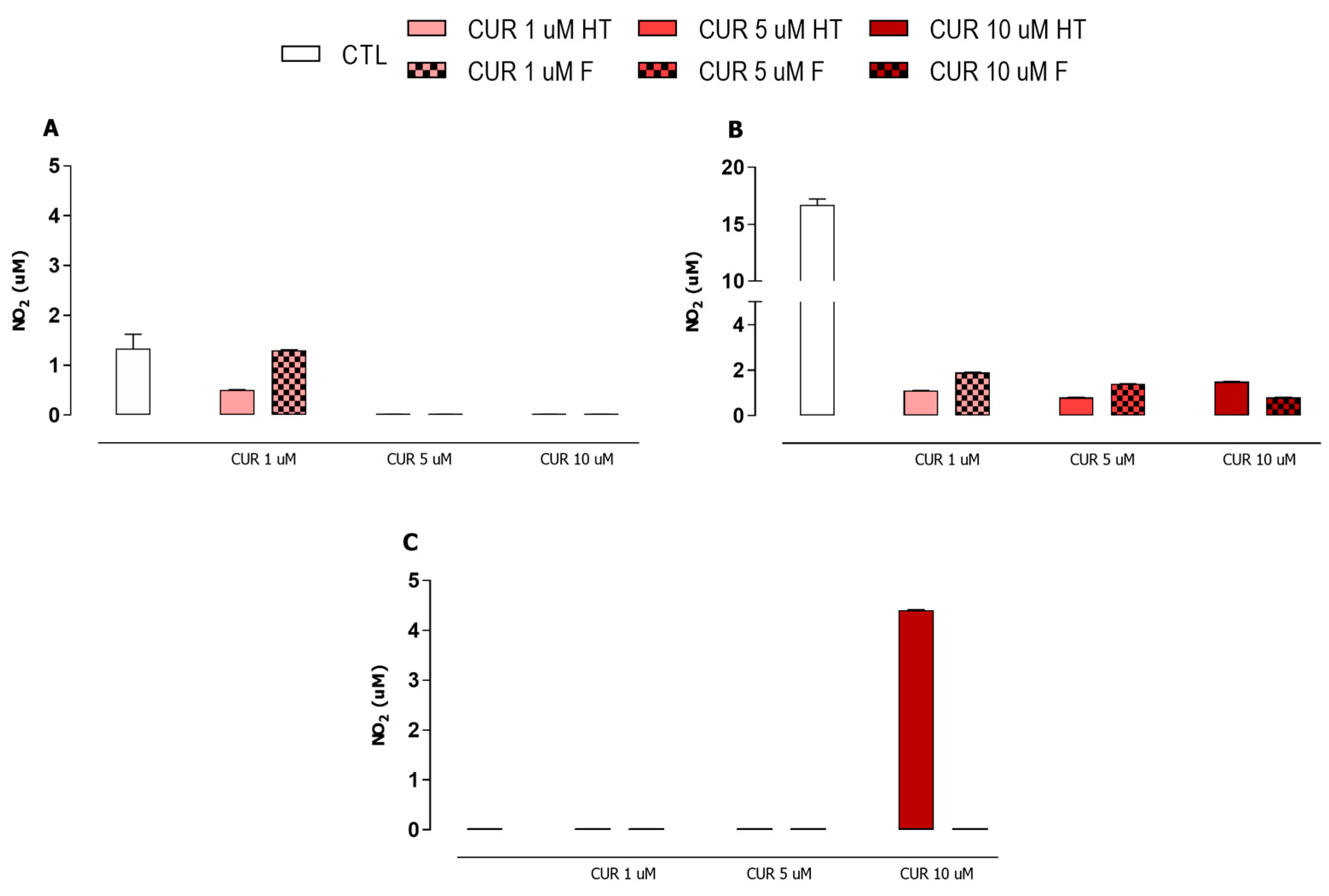
2.6. NO Production Assay
2.7. Arginase Activity Assay
2.8. Statistical Analyses
3. Results
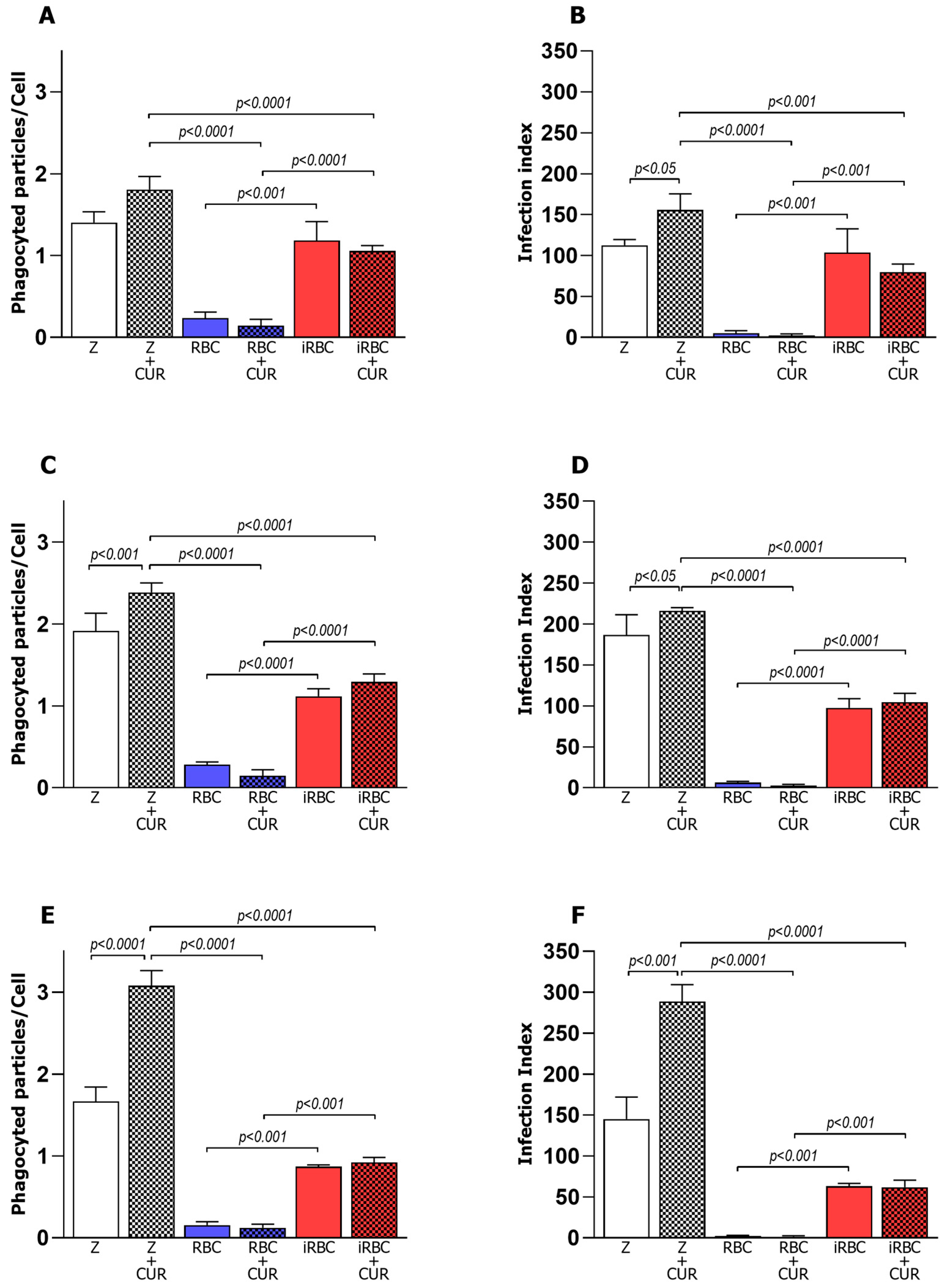
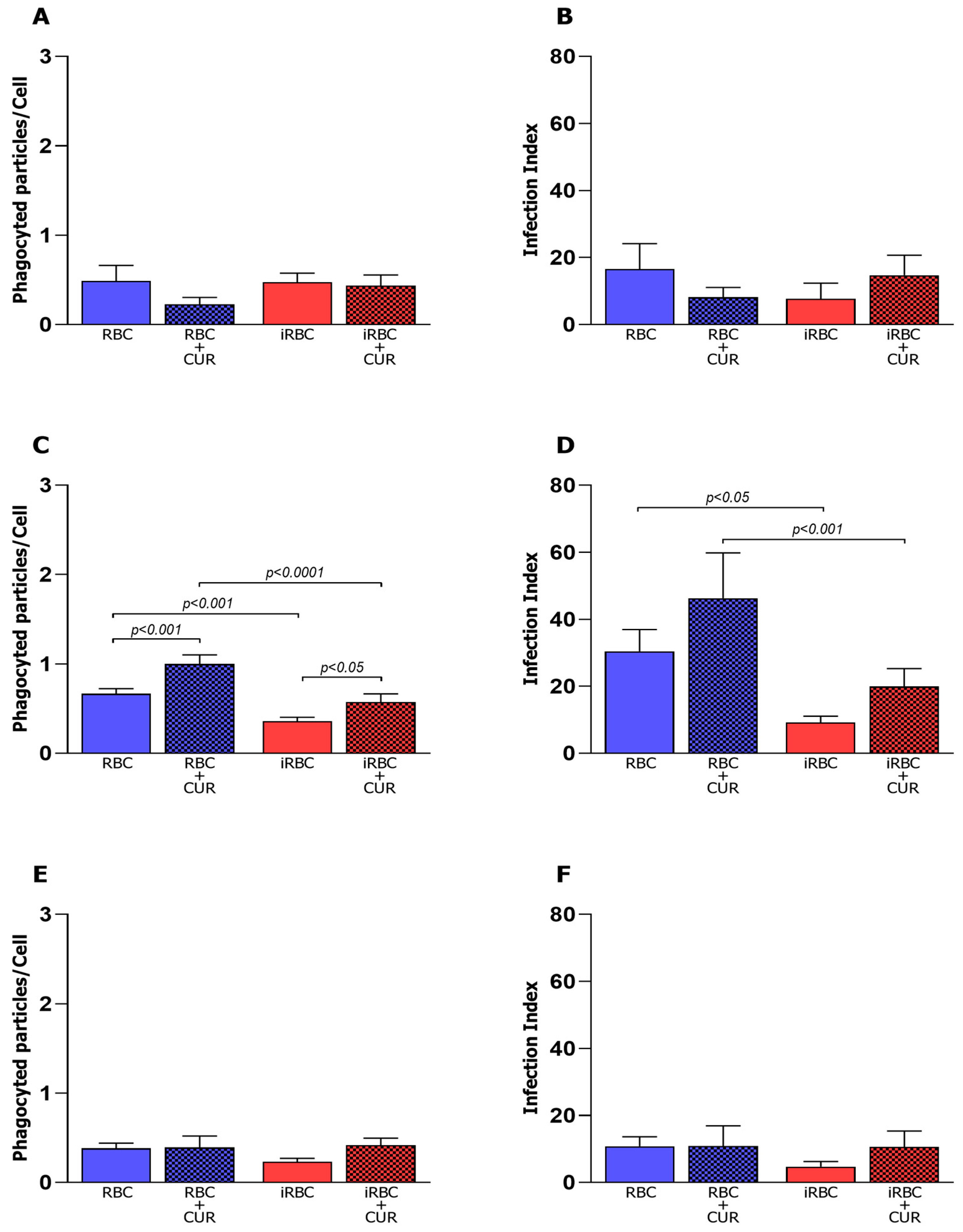
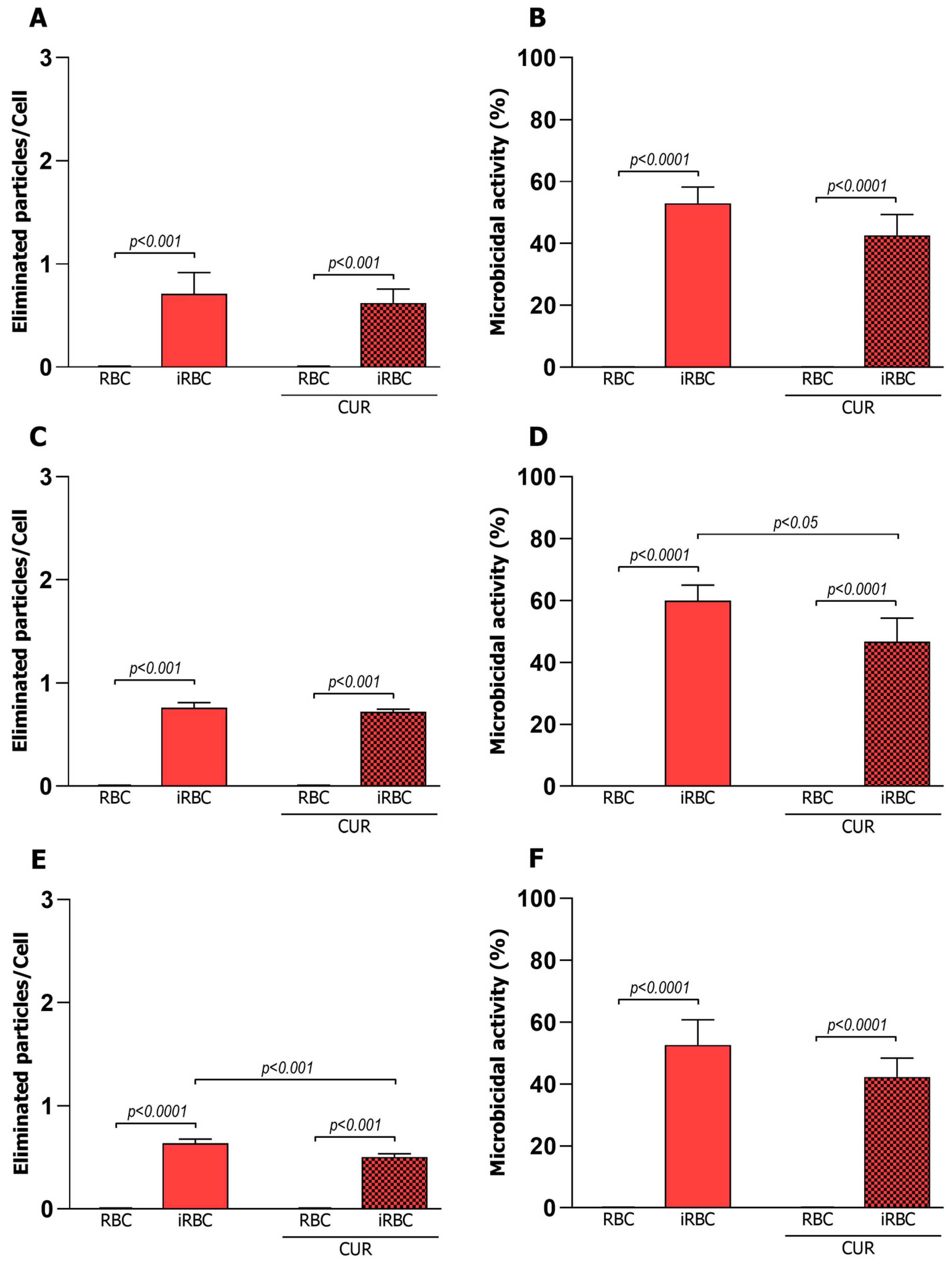
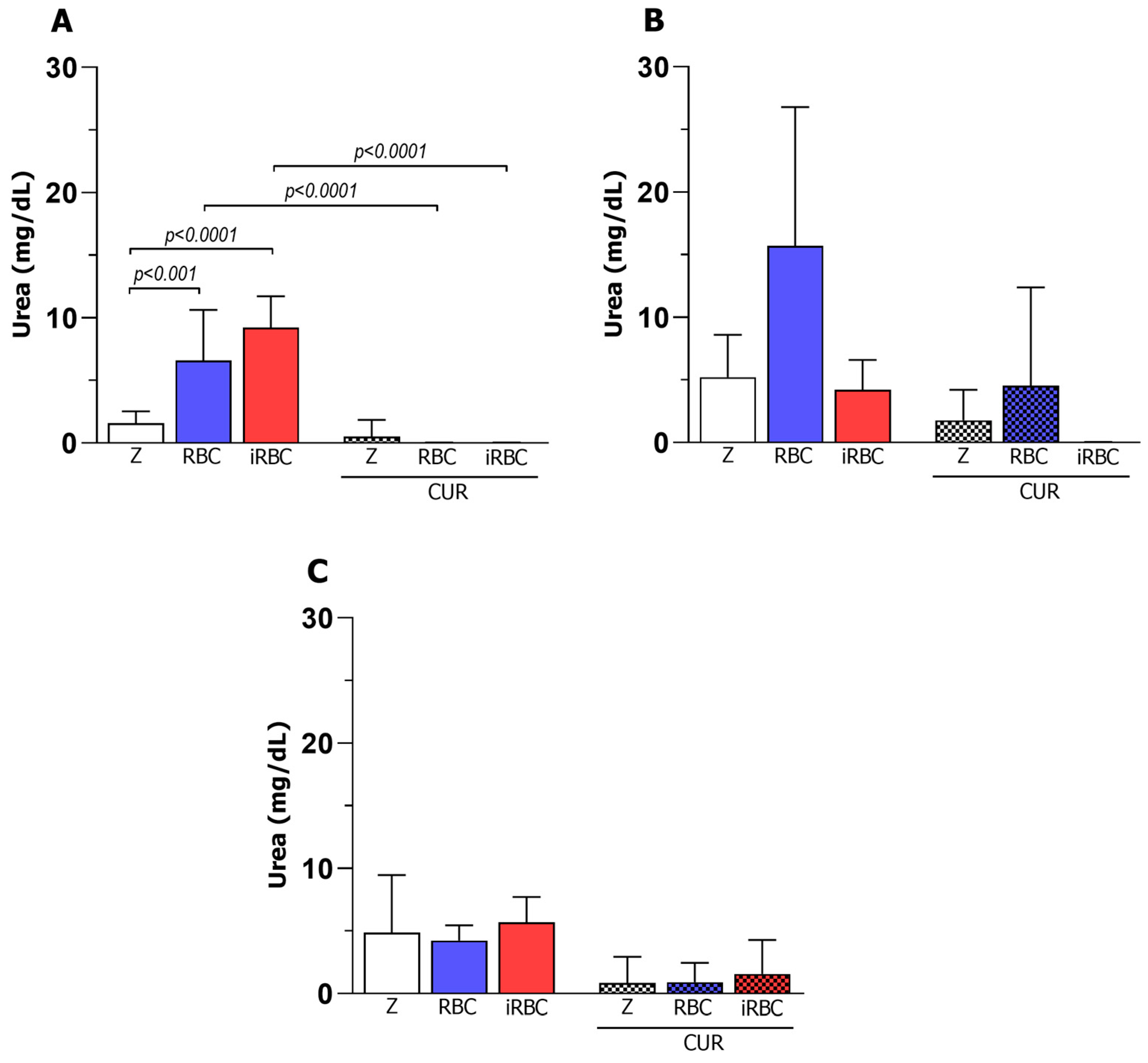
3.1. Pro-Inflammatory Macrophages Increased Phagocytosis and Microbicidal Activity of iRBCs
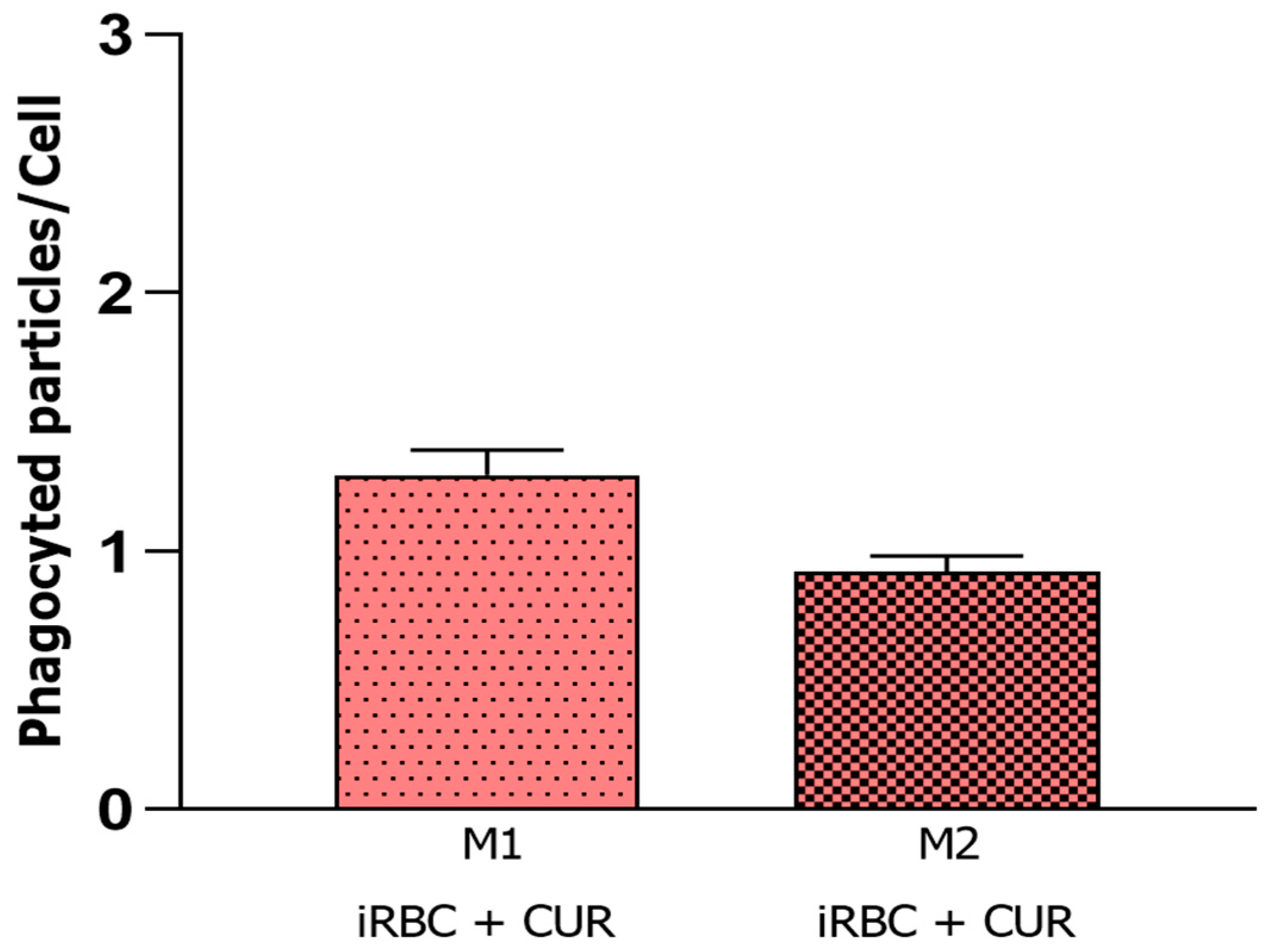
3.2. Curcumin Increased Phagocytosis in Non-Infected and Polarized Macrophages
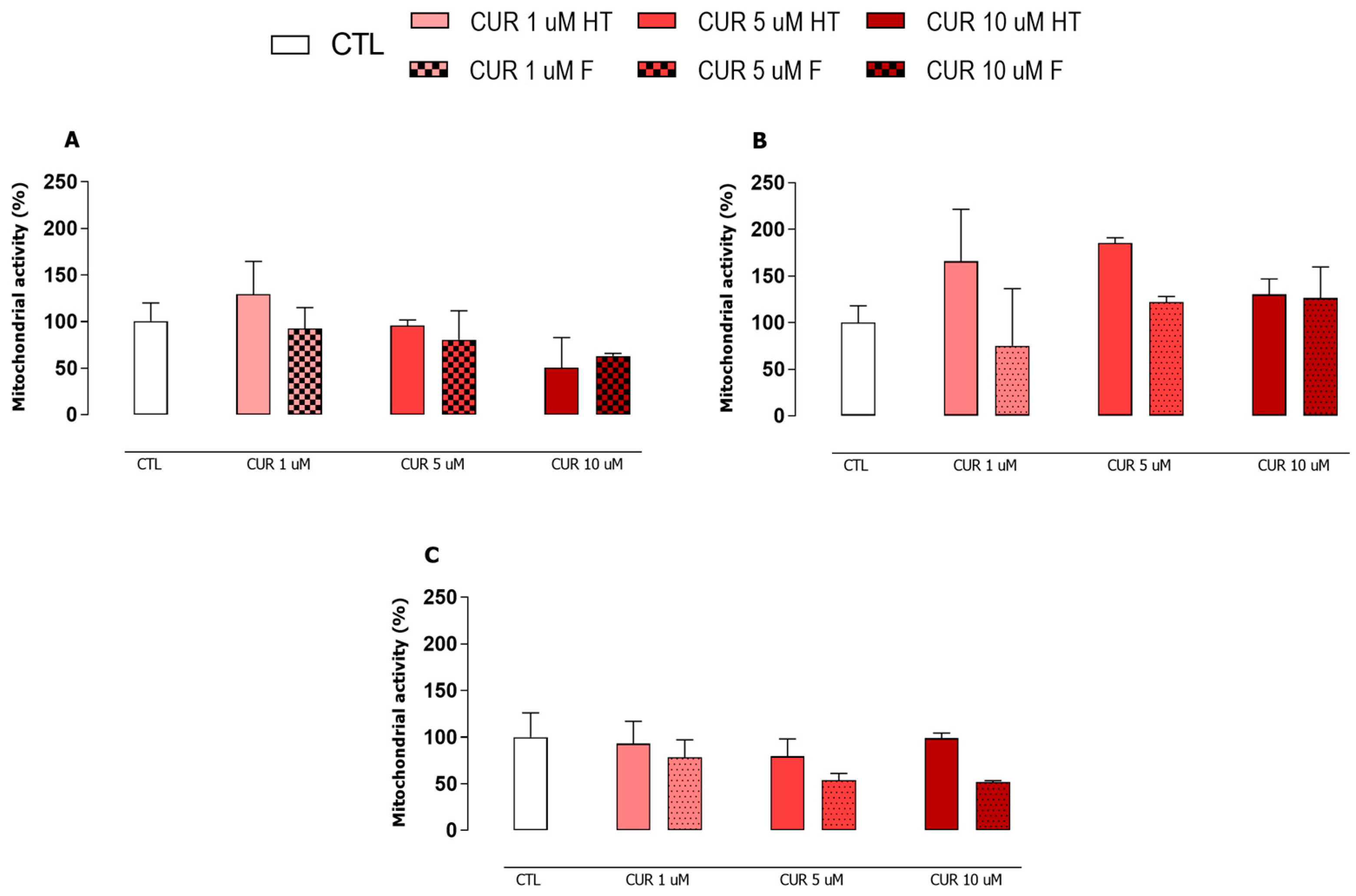
3.3. Curcumin Reduced the Microbicidal Activity of the Pro-Inflammatory Macrophages
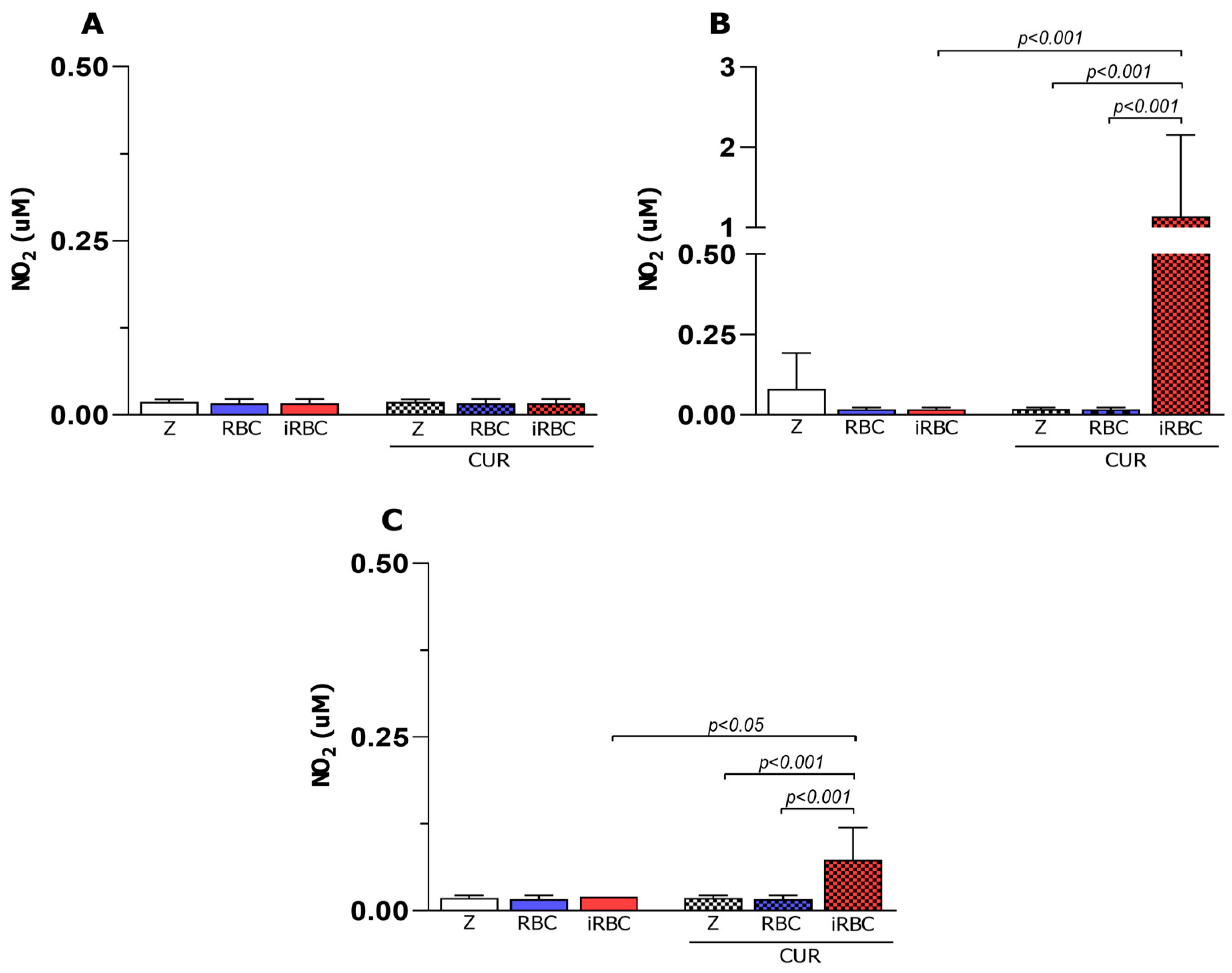
3.4. iNOS Enzyme Had No Participation in Macrophage Microbicidal Activity after Plasmodium Infection and Curcumin Treatment
3.5. Curcumin Decreased Arginase Activity in Infected and Non-Infected Macrophages
3.6. Curcumin Stabilized Macrophage Polarization during Plasmodium Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A




References
- Fu, Y.; Ding, Y.; Wang, Q.; Zhu, F.; Tan, Y.; Lu, X.; Guo, B.; Zhang, Q.; Cao, Y.; Liu, T.; et al. Blood-stage malaria parasites manipulate host innate immune responses through the induction of sFGL2. Sci. Adv. 2020, 9, eaay9269. [Google Scholar] [CrossRef]
- Shahbodaghi, S.D.; Rathjen, N.A. Malaria: Prevention, Diagnosis, and Treatment. Am. Fam. Physician 2022, 106, 270–278. [Google Scholar]
- Milner, E.M.; Kariger, P.; Pickering, A.J.; Stewart, C.P.; Byrd, K.; Lin, A.; Rao, B.; Dentz, H.N.; Null, C.; Fernald, L.C.H. Association between Malaria Infection and Early Childhood Development Mediated by Anemia in Rural Kenya. Int. J. Environ. Res. Public Health 2020, 17, 902. [Google Scholar] [CrossRef] [PubMed]
- Cutts, J.C.; Agius, P.A.; Lin, Z.; Powell, R.; Moore, K.; Draper, B.; Simpson, J.A.; Fowkes, F.J.I. Pregnancy-specific malarial immunity and risk of malaria in pregnancy and adverse birth outcomes: A systematic review. BMC Med. 2020, 18, 14. [Google Scholar] [CrossRef]
- World Health Organization; Global Malaria Programme. World Malaria Report; Creative Commons; World Health Organization: Mountain View, CA, USA, 2022; p. 293. ISBN 978-92-4-006489-8. Available online: https://www.who.int/publications/i/item/9789240064898 (accessed on 15 October 2023).
- Hussein, M.I.H.; Albashir, A.A.D.; Elawad, O.A.M.A.; Homeida, A. Malaria and COVID-19: Unmasking their ties. Malar J. 2020, 19, 457. [Google Scholar] [CrossRef] [PubMed]
- Milner, D.A. Malaria Pathogenesis. Cold Spring Harb. Perspect. Med. 2018, 8, a025569. [Google Scholar] [CrossRef]
- Ozarslan, N.; Robinson, J.F.; Gaw, S.L. Circulating Monocytes, Tissue Macrophages, and Malaria. J. Trop. Med. 2019, 2019, 1–9. [Google Scholar] [CrossRef]
- Chua, C.L.L.; Brown, G.; Hamilton, J.A.; Rogerson, S.; Philippe, B. Monocytes and macrophages in malaria: Protection or pathology? Trends Parasitol. 2013, 29, 26–34. [Google Scholar] [CrossRef]
- Drewry, L.L.; Harty, J.T. Balancing in a black box: Potential immunomodulatory roles for TGF-β signaling during blood-stage malaria. Virulence 2020, 11, 159–169. [Google Scholar] [CrossRef]
- Kang, S.; Kumanogoh, A. The spectrum of macrophage activation by immunometabolism. Jpn. Soc. Immunol. 2020, 7, 467–473. [Google Scholar] [CrossRef]
- Atri, C.; Guerfali, F.Z.; Laouini, D. Role of Human Macrophage Polarization in Inflammation during Infectious Diseases. Int. J. Mol. Sci. 2018, 6, 1081. [Google Scholar] [CrossRef] [PubMed]
- Vergadi, E.; Ieronymaki, E.; Lyroni, K.; Vaporidi, K.; Tsatsanis, C. Akt Signaling Pathway in Macrophage Activation and M1/M2 Polarization. J. Immunol. 2017, 198, 1006–1014. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, A.; Blesso, C.N.; Barreto, G.; Banach, M.; Majeed, M.; Sahebkar, A. Macrophage plasticity, polarization and function in response to curcumin, a diet-derived polyphenol, as an immunomodulatory agent. J. Nutr. Biochem. 2019, 66, 1–16. [Google Scholar] [CrossRef]
- Jamil, S.N.H.; Ali, A.H.; Feroz, A.R.; Lam, S.D.; Agustar, H.K.; Razak, M.R.M.A.; Latip, J. Curcumin and Its Derivatives as Potential Antimalarial and Anti-Inflammatory Agents: A Review on Structure–Activity Relationship and Mechanism of Action. Pharmaceutics 2023, 16, 609. [Google Scholar] [CrossRef] [PubMed]
- Generally Recognized as Safe (GRAS). GRAS Notice Inventory No. 822. Available online: https://www.fda.gov/media/132575/download (accessed on 26 September 2023).
- Hosseini-Zare, M.S.; Sarhadi, M.; Zarei, M.; Thilagavathi, R.; Selvam, C. Synergistic effects of curcumin and its analogs with other bioactive compounds: A comprehensive review. Eur. J. Med. Chem. 2021, 15, 113072. [Google Scholar] [CrossRef]
- Dende, C.; Meena, J.; Nagarajan, P.; Nagaraj, V.A.; Panda, A.K.; Padmanaban, G. Nanocurcumin is superior to native curcumin in preventing degenerative changes in Experimental Cerebral Malaria. Sci. Rep. 2017, 30, 10062. [Google Scholar] [CrossRef]
- Manconi, M.; Manca, M.L.; Escribano-Ferrer, E.; Elisabet Martí Coma-Cros, E.M.; Biosca, A.; Lantero, E.; Fernàndez-Busquets, X.; Fadda, A.M.; Caddeo, C. Nanoformulation of curcumin-loaded eudragit-nutriosomes to counteract malaria infection by a dual strategy: Improving antioxidant intestinal activity and systemic efficacy. Int. J. Pharm. 2019, 556, 82–88. [Google Scholar] [CrossRef]
- Tu, Q.D.; Jin, J.; Hu, X.; Ren, Y.; Zhao, L.; He, Q. Curcumin Improves the Renal Autophagy in Rat Experimental Membranous Nephropathy via Regulating the PI3K/AKT/mTOR and Nrf2/HO-1 Signaling Pathways. BioMed Res. Int. 2020, 2020, 7069052. [Google Scholar] [CrossRef]
- Fuentes, A.-L.; Millis, L.; Vapenik, J.; Sigola, L. Lipopolysaccharide-mediated enhancement of zymosan phagocytosis by RAW 264.7 macrophages is independent of opsonins, laminarin, mannan, and complement receptor 3. J. Surg. Res. 2014, 189, 304–312. [Google Scholar] [CrossRef]
- Chan, J.-A.; Boyle, M.J.; Moore, K.A.; Reiling, L.; Lin, Z.; Hasang, W.; Avril, M.; Manning, L.; Mueller, I.; Laman, M.; et al. Antibody targets on the surface of Plasmodium falciparum–infected erythrocytes that are associated with immunity to severe malaria in young children. J. Infect. Dis. 2019, 219, 819–828. [Google Scholar] [CrossRef]
- Shear, H.L.; Nussenzweig, R.S.; Bianco, C. Immune phagocytosis in murine malaria. J. Exp. Med. 1979, 149, 1288–1298. [Google Scholar] [CrossRef] [PubMed]
- Seixas, E.; Moura Nunes, J.F.; Matos, I.; Coutinho, A. The interaction between DC and Plasmodium berghei/chabaudi-infected erythrocytes in mice involves direct cell-to-cell contact, internalization, and TLR. Eur. J. Immunol. 2009, 39, 1850–1863. [Google Scholar] [CrossRef]
- Stevenson, M.M.; Urban, B.C. Antigen presentation and dendritic cell biology in malaria. Parasite Immunol. 2006, 28, 5–14. [Google Scholar] [CrossRef]
- Jain, K.; Sood, S.; Gowthamarajan, K. Modulation of cerebral malaria by curcumin as an adjunctive therapy. Braz J. Infect. Dis. 2013, 5, 579–591. [Google Scholar] [CrossRef]
- Ghosh, A.; Banerjee, T.; Bhandary, S.; Surolia, A. Formulation of nanosized curcumin and demonstration of its antimalarial efficacy. Int. J. Nanomed. 2014, 9, 5373–5387. [Google Scholar] [CrossRef] [PubMed]
- Zendehdel, E.; Abdollahi, E.; Momtazi-Borojeni, A.A.; Korani, M.; Alavizadeh, S.H.; Sahebkar, A. The molecular mechanisms of curcumin’s inhibitory effects on cancer stem cells. J. Cell. Biochem. 2018, 4, 4739–4747. [Google Scholar] [CrossRef]
- Kloesch, B.; Gober, L.; Loebsch, S.; Vcelar, B.; Helson, L.; Steiner, G. In Vitro Study of a Liposomal Curcumin Formulation (Lipocurc™): Toxicity and Biological Activity in Synovial Fibroblasts and Macrophages. In Vivo 2016, 4, 413–419. [Google Scholar]
- Brouet, I.; Ohshima, H. Curcumin, an anti-tumor promoter, and anti-inflammatory agent, inhibits induction of nitric oxide synthase in activated macrophages. Biochem. Biophys. Res. Commun. 1995, 206, 533–540. [Google Scholar] [CrossRef]
- Gao, S.; Zhou, J.; Liu, N.; Wang, L.; Gao, Q.; Wu, Y.; Zhao, Q.; Liu, P.; Wang, S.; Liu, Y.; et al. Curcumin induces M2 macrophage polarization by secretion of IL-4 and/or IL-13. J. Mol. Cell. Cardiol. 2015, 85, 131–139. [Google Scholar] [CrossRef]
- Kalantari, P. The Emerging Role of Pattern Recognition Receptors in the Pathogenesis of Malaria. Vaccines 2018, 6, 13. [Google Scholar] [CrossRef]
- Bisht, K.; Choi, W.H.; Park, S.Y.; Chung, M.K.; Koh, W.S. Curcumin enhances the non-inflammatory phagocytic activity of RAW264.7 cells. Biochem. Biophys. Res. Commun. 2009, 379, 632–636. [Google Scholar] [CrossRef] [PubMed]
- Rath, M.; Müller, I.; Kropf, P.; Closs, E.I.; Munder, M. Metabolism via arginase or nitric oxide synthase: Two competing arginine pathways in macrophages. Front. Immunol. 2014, 5, 532. [Google Scholar] [CrossRef] [PubMed]
- Pemmari, A. MKP—1 Promotes Anti -inflammatory M (IL-4/IL-13) Macrophage Phenotype and Mediates the Anti-inflammatory Effects of Glucocorticoids. Basic Clin. Pharmacol. Toxicol. 2018, 124, 404–415. [Google Scholar] [CrossRef]
- Carneiro, P.P.; Conceição, J.; Macedo, M.; Magalhães, V.; Carvalho, E.M.; Bacellar, O. The Role of Nitric Oxide and Reactive Oxygen Species in the Killing of Leishmania braziliensis by Monocytes from Patients with Cutaneous Leishmaniasis. PLoS ONE 2016, 11, e0148084. [Google Scholar] [CrossRef]
- Yang, Z.; Ming, X.-F. Functions of arginase isoforms in macrophage inflammatory responses: Impact on cardiovascular diseases and metabolic disorders. Front. Immunol. 2014, 5, 533. [Google Scholar] [CrossRef]
- Akinyemi, A.J.; Onyebueke, N.; Faboya, O.A.; Onikanni, S.A.; Fadaka, A.; Olayide, I. Curcumin inhibits adenosine deaminase and arginase activities in cadmium-induced renal toxicity in rat kidneys. J. Food Drug Anal. 2017, 25, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Bounaama, A.; Djerdjouri, B.; Laroche-Clary, A.; Le Morvan, V.; Robert, J. Short curcumin treatment modulates oxidative stress, arginase activity, aberrant crypt foci, and TGF-1 and HES-1 transcripts in 1,2-dimethylhydrazine-colon carcinogenesis in mice. Toxicology 2012, 302, 308–317. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cordeiro, M.C.C.; Tomé, F.D.; Arruda, F.S.; da Fonseca, S.G.; Nagib, P.R.A.; Celes, M.R.N. Curcumin as a Stabilizer of Macrophage Polarization during Plasmodium Infection. Pharmaceutics 2023, 15, 2505. https://doi.org/10.3390/pharmaceutics15102505
Cordeiro MCC, Tomé FD, Arruda FS, da Fonseca SG, Nagib PRA, Celes MRN. Curcumin as a Stabilizer of Macrophage Polarization during Plasmodium Infection. Pharmaceutics. 2023; 15(10):2505. https://doi.org/10.3390/pharmaceutics15102505
Chicago/Turabian StyleCordeiro, Maria Clara C., Fernanda D. Tomé, Felipe S. Arruda, Simone Gonçalves da Fonseca, Patrícia R. A. Nagib, and Mara R. N. Celes. 2023. "Curcumin as a Stabilizer of Macrophage Polarization during Plasmodium Infection" Pharmaceutics 15, no. 10: 2505. https://doi.org/10.3390/pharmaceutics15102505