
Investigation of the Impact of Saccharides on the Relative Activity of Trypsin and Catalase after Droplet and Spray Drying
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Miniaturized Drying (MD)
Evaporation Rate and Aspect Ratio
2.3. Spray Drying (SD)
Determination of Moisture Content
2.4. Molecular Docking and Molecular Dynamics Simulations
2.5. Characterization of the Enzymatic Activity of the MD and SD Samples
2.5.1. Enzymatic Activity of Catalase Using UV-Vis Spectroscopic Analysis
2.5.2. Enzymatic Activity of Trypsin Using UV-Vis Spectroscopy Analysis
2.5.3. Statistical Analysis and Generation of Graphs
3. Results
3.1. Drying of Catalase Formulations
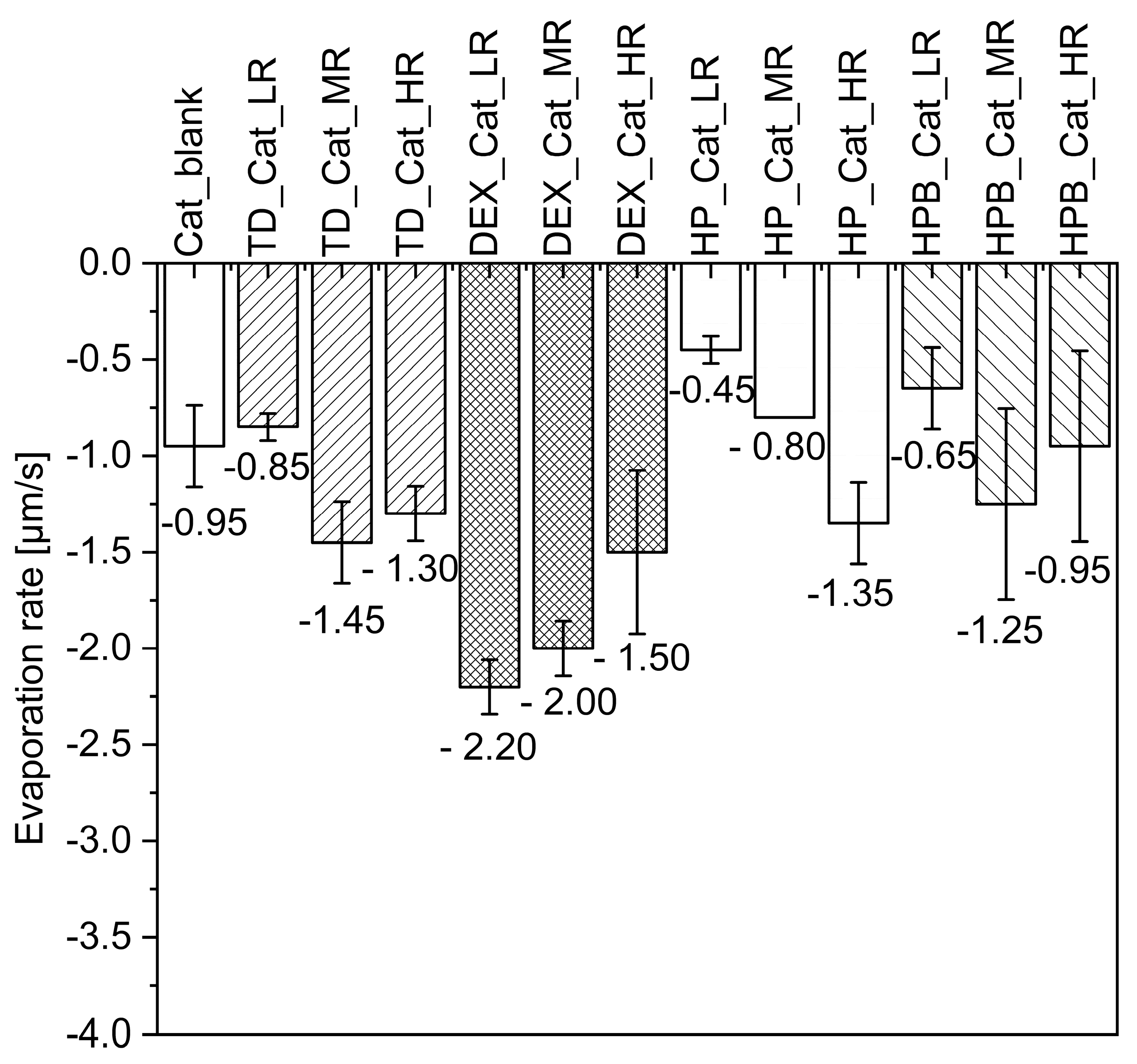
3.1.1. Formation of Catalase Particles during MD
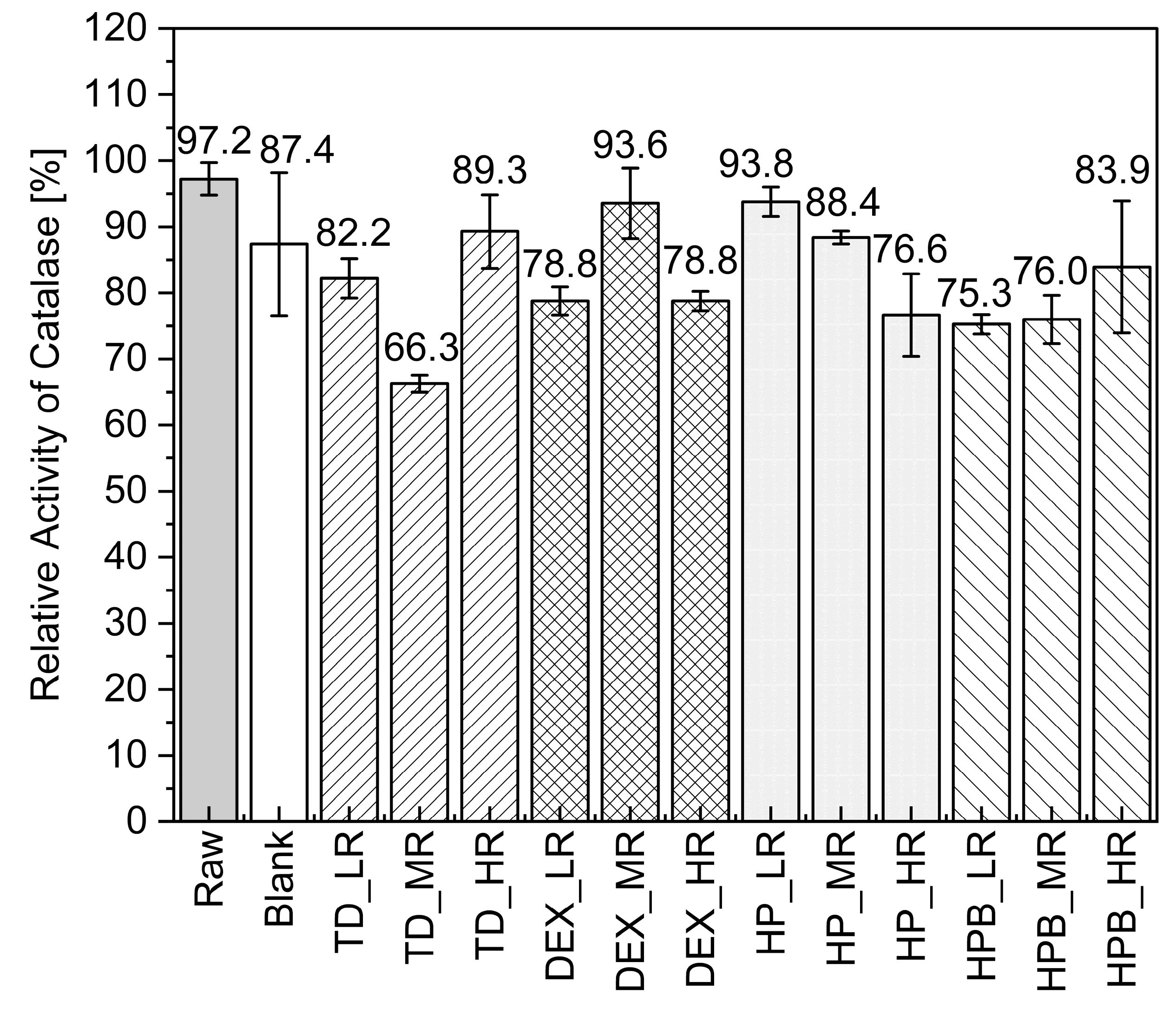
3.1.2. Catalase in-Process Activity during MD
3.1.3. Formation of Catalase Particles and Activity during SD
3.2. Drying of Trypsin Formulations
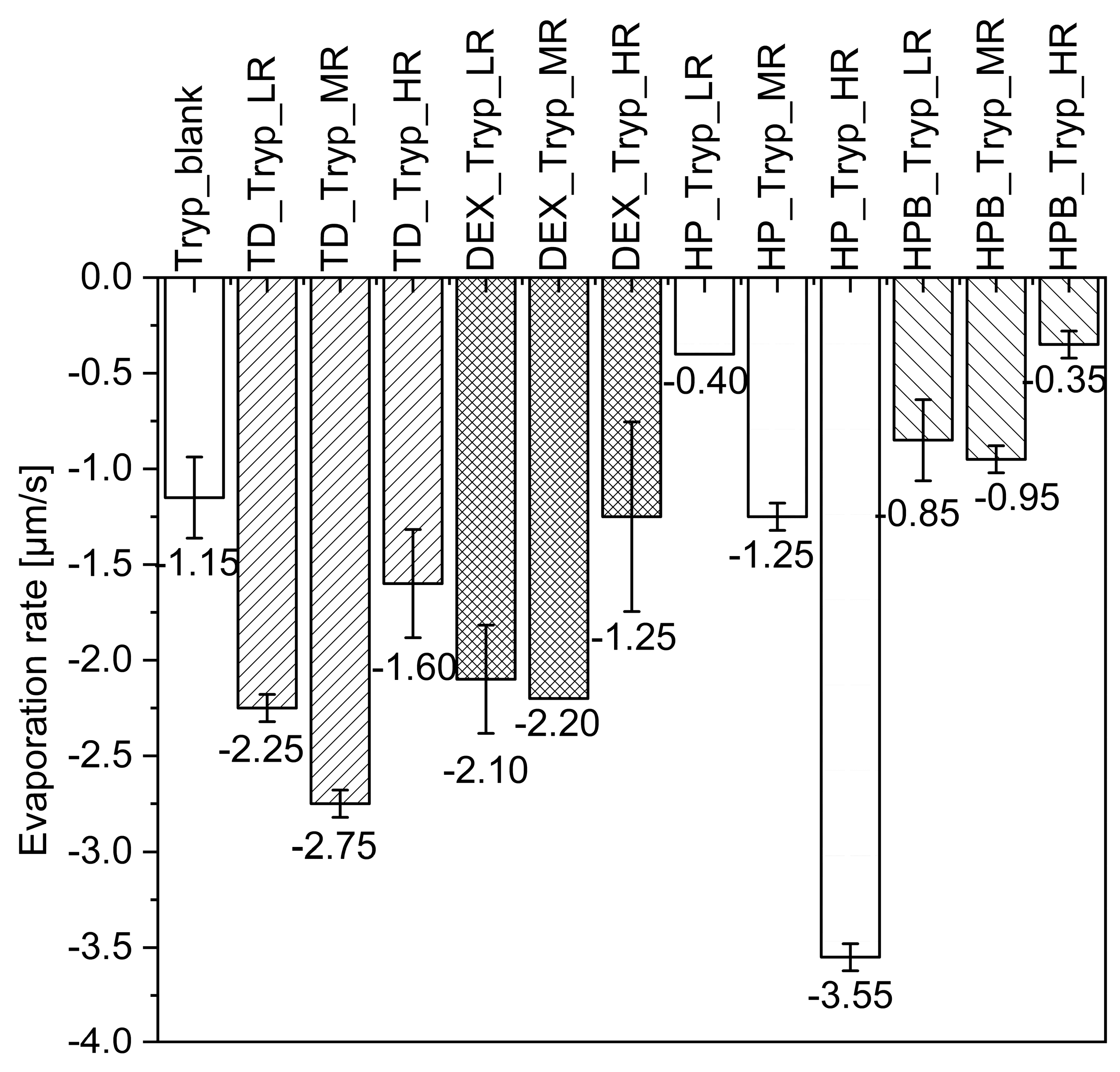
3.2.1. Formation of Trypsin Particles during MD
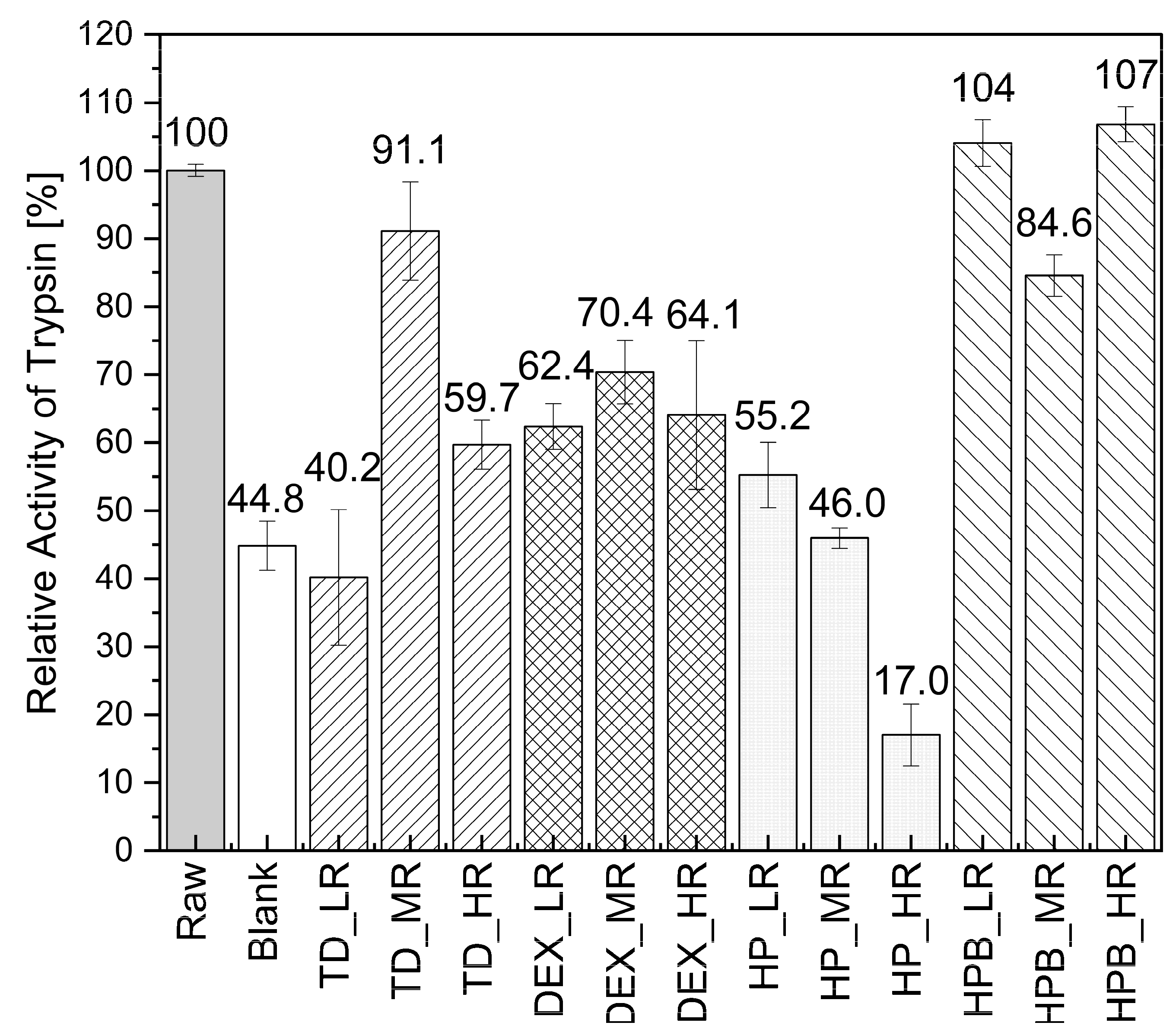
3.2.2. Trypsin Enzymatic Activity during MD
3.2.3. Formation of Trypsin Particles and Activity during SD
3.3. Modeling with Trypsin and HP or HPB
3.3.1. Protein-Protein and Protein-Cyclodextrin Docking with Trypsin
3.3.2. Molecular Dynamics of Trypsin-Trypsin and Trypsin-Cyclodextrin Interactions
4. Discussion
4.1. Drying of Catalase and Trypsin without the Addition of Saccharides
4.2. Drying of Catalase and Trypsin in the Presence of TD
4.3. Drying of Catalase and Trypsin in the Presence of DEX
4.4. Drying of Catalase and Trypsin in the Presence of HP and HPB
4.5. Molecular Modeling of Trypsin and Cyclodextrin Systems
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- InsightAce Analytic Pvt. Ltd. Lyophilization Services for Biopharmaceuticals Market Size, Share & Trends Analysis Report by Type of Biologic Lyophilized (Antibodies, Cell and Gene Therapies, Vaccines, Other Biologics), by Type of Primary Packaging System, by Region, and by Segment Forecasts, 2023–2031. Report ID:1350. 2023. Available online: https://www.insightaceanalytic.com/report/global-lyophilization-services-for-biopharmaceuticals-market/1350 (accessed on 10 October 2023).
- Pinto, J.T.; Faulhammer, E.; Dieplinger, J.; Dekner, M.; Makert, C.; Nieder, M.; Paudel, A. Progress in Spray-Drying of Protein Pharmaceuticals: Literature Analysis of Trends in Formulation and Process Attributes. Dry. Technol. 2021, 39, 1415–1446. [Google Scholar] [CrossRef]
- Ziaee, A.; Albadarin, A.B.; Padrela, L.; Femmer, T.; O’Reilly, E.; Walker, G. Spray Drying of Pharmaceuticals and Biopharmaceuticals: Critical Parameters and Experimental Process Optimization Approaches. Eur. J. Pharm. Sci. 2019, 127, 300–318. [Google Scholar] [CrossRef] [PubMed]
- Ziaee, A.; Albadarin, A.B.; Padrela, L.; Ung, M.T.; Femmer, T.; Walker, G.; O’Reilly, E. A Rational Approach towards Spray Drying of Biopharmaceuticals: The Case of Lysozyme. Powder Technol. 2020, 366, 206–215. [Google Scholar] [CrossRef]
- Ferreira, M.A.; Martins, J.P.; Hirvonen, J.; Santos, H.A. Spray-Drying for the Formulation of Oral Drug Delivery Systems, 1st ed.; Academic Press: London, UK, 2020. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, H.; Ghosh, D. The Stabilizing Excipients in Dry State Therapeutic Phage Formulations. AAPS PharmSciTech 2020, 21, 133. [Google Scholar] [CrossRef] [PubMed]
- Baynes, B.M.; Wang, D.I.C.; Trout, B.L. Role of Arginine in the Stabilization of Proteins against Aggregation. Biochemistry 2005, 44, 4919–4925. [Google Scholar] [CrossRef] [PubMed]
- Labrude, P.; Rasolomanana, M.; Vigneron, C.; Thirion, C.; Chaillot, B. Protective Effect of Sucrose on Spray Drying of Oxyhemoglobin. J. Pharm. Sci. 1989, 78, 223–229. [Google Scholar] [CrossRef]
- Cleland, J.L.; Lam, X.; Kendrick, B.; Yang, J.; Yang, T.H.; Overcashier, D.; Brooks, D.; Hsu, C.; Carpenter, J.F. A Specific Molar Ratio of Stabilizer to Protein Is Required for Storage Stability of a Lyophilized Monoclonal Antibody. J. Pharm. Sci. 2001, 90, 310–321. [Google Scholar] [CrossRef]
- Grasmeijer, N.; Tiraboschi, V.; Woerdenbag, H.J.; Frijlink, H.W.; Hinrichs, W.L.J. Identifying Critical Process Steps to Protein Stability during Spray Drying Using a Vibrating Mesh or a Two-Fluid Nozzle. Eur. J. Pharm. Sci. 2019, 128, 152–157. [Google Scholar] [CrossRef]
- Grasmeijer, N. Improving Protein Stabilization by Spray Drying: Formulation and Process Development. Ph.D. Thesis, University of Groningen, Groningen, The Netherlands, 2016. Available online: https://pure.rug.nl/ws/portalfiles/portal/26868430/Complete_thesis.pdf (accessed on 13 February 2023).
- Carpenter, J.F.; Crowe, J.H. An Infrared Spectroscopic Study of the Interactions of Carbohydrates with Dried Proteins. Biochemistry 1989, 28, 3916–3922. [Google Scholar] [CrossRef]
- Boel, E.; Koekoekx, R.; Dedroog, S.; Babkin, I.; Vetrano, M.R.; Clasen, C.; Van den Mooter, G. Unraveling Particle Formation: From Single Droplet Drying to Spray Drying and Electrospraying. Pharmaceutics 2020, 12, 625. [Google Scholar] [CrossRef]
- de Souza Lima, R.; Ré, M.I.; Arlabosse, P. Drying Droplet as a Template for Solid Formation: A Review. Powder Technol. 2020, 359, 161–171. [Google Scholar] [CrossRef]
- Both, E.M.; Tersteeg, S.M.B.; Boom, R.M.; Schutyser, M.A.I. Drying Kinetics and Viscoelastic Properties of Concentrated Thin Films as a Model System for Spray Drying. Colloids Surf. A Physicochem. Eng. Asp. 2020, 585, 124075. [Google Scholar] [CrossRef]
- Dieplinger, J.; Pinto, J.T.; Dekner, M.; Brachtl, G.; Paudel, A. Impact of Different Saccharides on the In-Process Stability of a Protein Drug During Evaporative Drying: From Sessile Droplet Drying to Lab-Scale Spray Drying. Pharm. Res. 2023, 40, 1283–1298. [Google Scholar] [CrossRef]
- Product Information on Trypsin from Bovine Pancreas. Available online: https://www.sigmaaldrich.com/AT/en/product/sigma/t7309 (accessed on 9 March 2022).
- Product Information on Catalase from Bovine Liver. Available online: https://www.sigmaaldrich.com/AT/en/product/sigma/c9322 (accessed on 9 March 2022).
- Vehring, R. Pharmaceutical Particle Engineering via Spray Drying. Pharm. Res. 2008, 25, 999–1022. [Google Scholar] [CrossRef]
- Chemical Computing Group ULC. Molecular Operating Environment (MOE). 2022.02; Chemical Computing Group ULC: Montreal, QC, Canada, 2023. [Google Scholar]
- Maeki, M.; Ito, S.; Takeda, R.; Ueno, G.; Ishida, A.; Tani, H.; Yamamoto, M.; Tokeshi, M. Room-Temperature Crystallography Using a Microfluidic Protein Crystal Array Device and Its Application to Protein-Ligand Complex Structure Analysis. Chem. Sci. 2020, 11, 9072–9087. [Google Scholar] [CrossRef]
- Case, D.A.; Aktulga, H.M.; Belfon, K.; Ben-Shalom, I.Y.; Berryman, J.T.; Brozell, S.R.; Cerutti, D.S.; Cheatham III, T.E.; Cisneros, G.A.; Cruzeiro, V.W.D.; et al. AMBER22; University of California, San Francisco: San Francisco, CA, USA, 2022. [Google Scholar]
- Best, R.B.; Zhu, X.; Shim, J.; Lopes, P.E.M.; Mittal, J.; Feig, M.; MacKerell, A.D. Optimization of the Additive CHARMM All-Atom Protein Force Field Targeting Improved Sampling of the Backbone φ, ψ and Side-Chain Χ1 and Χ2 Dihedral Angles. J. Chem. Theory Comput. 2012, 8, 3257–3273. [Google Scholar] [CrossRef]
- Vanommeslaeghe, K.; Hatcher, E.; Acharya, C.; Kundu, S.; Zhong, S.; Shim, J.; Darian, E.; Guvench, O.; Lopes, P.; Vorobyov, I.; et al. CHARMM General Force Field (CGenFF): A Force Field for Drug-like Molecules Compatible with the CHARMM All-Atom Additive Biological Force Fields. J. Comput. Chem. 2010, 31, 671–690. [Google Scholar] [CrossRef]
- Lee, J.; Cheng, X.; Swails, J.M.; Yeom, M.S.; Eastman, P.K.; Lemkul, J.A.; Wei, S.; Buckner, J.; Jeong, J.C.; Qi, Y.; et al. CHARMM-GUI Input Generator for NAMD, GROMACS, AMBER, OpenMM, and CHARMM/OpenMM Simulations Using the CHARMM36 Additive Force Field. J. Chem. Theory Comput. 2016, 12, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Sunhwan, J.; Taehoon, K.; Vidyashankara, G.I.; Wonpil, I. Software News and Updates CHARMM-GUI: A Web-Based Graphical User Interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of Simple Potential Functions for Simulating Liquid Water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Merck KGaA. Procedure for Enzymatic Assay of Trypsin (EC 3.4.21.4). Available online: https://www.sigmaaldrich.com/AT/en/technical-documents/protocol/protein-biology/enzyme-activity-assays/enzymatic-assay-of-trypsin (accessed on 27 October 2021).
- Merck KGaA. Enzymatic Assay of Catalase (EC 1.11.1.6). Available online: https://www.sigmaaldrich.com/AT/en/technical-documents/protocol/protein-biology/enzyme-activity-assays/enzymatic-assay-of-catalase (accessed on 27 October 2021).
- Larson, R.G. Transport and Deposition Patterns in Drying Sessile Droplets. AIChE J. 2014, 60, 1538–1571. [Google Scholar] [CrossRef]
- Kerdpol, K.; Kicuntod, J.; Wolschann, P.; Mori, S.; Rungnim, C.; Kunaseth, M.; Okumura, H.; Kungwan, N.; Rungrotmongkol, T. Cavity Closure of 2-Hydroxypropyl-β-Cyclodextrin: Replica Exchange Molecular Dynamics Simulations. Polymers 2019, 11, 145. [Google Scholar] [CrossRef] [PubMed]
- Tarasevich, Y.Y.; Vodolazskaya, I.V.; Isakova, O.P. Desiccating Colloidal Sessile Drop: Dynamics of Shape and Concentration. Colloid Polym. Sci. 2011, 289, 1015–1023. [Google Scholar] [CrossRef]
- Anton Paar GmbH. The Viscosity of Protein Therapeutics; Anton Paar GmbH: Graz, Austria; Available online: https://wiki.anton-paar.com/en/viscosity-of-protein-therapeutics/ (accessed on 26 June 2023).
- Qi, X.; Liu, H.; Ren, Y.; Zhu, Y.; Wang, Q.; Zhang, Y.; Wu, Y.; Yuan, L.; Yan, H.; Liu, M. Effects of Combined Binding of Chlorogenic Acid/Caffeic Acid and Gallic Acid to Trypsin on Their Synergistic Antioxidant Activity, Enzyme Activity and Stability. Food Chem. X 2023, 18, 100664. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Zou, T.; Li, H.; Cheng, H. The Interaction of Sodium Dodecyl Sulfate with Trypsin: Multi-Spectroscopic Analysis, Molecular Docking, and Molecular Dynamics Simulation. Int. J. Biol. Macromol. 2020, 162, 1546–1554. [Google Scholar] [CrossRef]
- Zhang, H.M.; Cao, J.; Tang, B.P.; Wang, Y.Q. Effect of TiO2 Nanoparticles on the Structure and Activity of Catalase. Chem. Biol. Interact. 2014, 219, 168–174. [Google Scholar] [CrossRef]
- Haj-Ahmad, R.R.; Elkordy, A.A.; Chaw, C.S.; Moore, A. Compare and Contrast the Effects of Surfactants (Pluronic ® F-127 and Cremophor ® EL) and Sugars (β-Cyclodextrin and Inulin) on Properties of Spray Dried and Crystallised Lysozyme. Eur. J. Pharm. Sci. 2013, 49, 519–534. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Z.; Yang, J.; Yue, Y.; Zhang, H. Evaporation Characteristics of Viscous Droplets on Stainless Steel Superhydrophobic Surface. Int. J. Therm. Sci. 2022, 183, 107843. [Google Scholar] [CrossRef]
- Giuffrida, S.; Cordone, L.; Cottone, G. Bioprotection Can Be Tuned with a Proper Protein/Saccharide Ratio: The Case of Solid Amorphous Matrices. J. Phys. Chem. B 2018, 122, 8642–8653. [Google Scholar] [CrossRef]
- Ohtake, S.; Kita, Y.; Arakawa, T. Interactions of Formulation Excipients with Proteins in Solution and in the Dried State. Adv. Drug Deliv. Rev. 2011, 63, 1053–1073. [Google Scholar] [CrossRef]
- Samra, H.S.; He, F.; Bhambhani, A.; Pipkin, J.D.; Zimmerer, R.; Joshi, S.B.; Middaugh, C.R. The Effects of Substituted Cyclodextrins on the Colloidal and Conformational Stability of Selected Proteins. J. Pharm. Sci. 2010, 99, 2800–2818. [Google Scholar] [CrossRef]
- Shao, X.; Hou, Y.; Zhong, X. Modulation of Evaporation-Affected Crystal Motion in a Drying Droplet by Saline and Surfactant Concentrations. Colloids Surf. A Physicochem. Eng. Asp. 2021, 623, 126701. [Google Scholar] [CrossRef]
- Park, J.K.; Khan, T. Other Microbial Polysaccharides: Pullulan, Scleroglucan, Elsinan, Levan, Alternant, Dextran; Woodhead Publishing Limited: Sawston, UK, 2009. [Google Scholar] [CrossRef]
- Sadek, C.; Schuck, P.; Fallourd, Y.; Pradeau, N.; Le Floch-Fouéré, C.; Jeantet, R. Drying of a Single Droplet to Investigate Process–Structure–Function Relationships: A Review. Dairy Sci. Technol. 2015, 95, 771–794. [Google Scholar] [CrossRef]
- Larsen, B.S.; Skytte, J.; Svagan, A.J.; Meng-Lund, H.; Grohganz, H.; Löbmann, K. Using Dextran of Different Molecular Weights to Achieve Faster Freeze-Drying and Improved Storage Stability of Lactate Dehydrogenase. Pharm. Dev. Technol. 2019, 24, 323–328. [Google Scholar] [CrossRef]
- Milani, S.; Faghihi, H.; Roulholamini Najafabadi, A.; Amini, M.; Montazeri, H.; Vatanara, A. Hydroxypropyl Beta Cyclodextrin: A Water-Replacement Agent or a Surfactant upon Spray Freeze-Drying of IgG with Enhanced Stability and Aerosolization. Drug Dev. Ind. Pharm. 2020, 46, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, K.; Teng, M.; Li, M.; Sui, X.; Liu, B.; Tian, B.; Fu, Q. Functionality-Related Characteristics of Hydroxypropyl-β-Cyclodextrin for the Complexation. J. Mol. Liq. 2022, 365, 120105. [Google Scholar] [CrossRef]
- Sohajda, T. The Effect of Different Degrees of Substitution in the Case of (2-Hydroxypropyl) Beta-Cyclodextrin; Carbohyde: Budapest, Hungary, 2022; Available online: https://www.carbohyde.com/the-effect-of-different-degrees-of-substitution-in-the-case-of-2-hydroxypropyl-beta-cyclodextrin/ (accessed on 15 June 2023).
- Efstratiou, M.; Christy, J.; Bonn, D.; Sefiane, K. The Effect of Substrate Temperature on the Evaporative Behaviour and Desiccation Patterns of Foetal Bovine Serum Drops. Colloids Interfaces 2021, 5, 43. [Google Scholar] [CrossRef]
- Maa, Y.F.; Costantino, H.R.; Nguyen, P.A.; Hsu, C.C. The Effect of Operating and Formulation Variables on the Morphology of Spray-Dried Protein Particles. Pharm. Dev. Technol. 1997, 2, 213–223. [Google Scholar] [CrossRef]
- Schönbeck, C.; Westh, P.; Madsen, J.C.; Larsen, K.L.; Städe, L.W.; Holm, R. Hydroxypropyl-Substituted β-Cyclodextrins: Influence of Degree of Substitution on the Thermodynamics of Complexation with Tauroconjugated and Glycoconjugated Bile Salts. Langmuir 2010, 26, 17949–17957. [Google Scholar] [CrossRef]
- Kumar, V.; Errington, J.R. Application of the Interface Potential Approach to Calculate the Wetting Properties of a Water Model System. Mol. Simul. 2013, 39, 1143–1152. [Google Scholar] [CrossRef]
- Van Gunsteren, W.F.; Berendsen, H.J.C. Algorithms for Macromolecular Dynamics and Constraint Dynamics. Mol. Phys. 1977, 34, 1311–1327. [Google Scholar] [CrossRef]
- Åqvist, J.; Wennerström, P.; Nervall, M.; Bjelic, S.; Brandsdal, B.O. Molecular Dynamics Simulations of Water and Biomolecules with a Monte Carlo Constant Pressure Algorithm. Chem. Phys. Lett. 2004, 384, 288–294. [Google Scholar] [CrossRef]
- Steinbach, P.J.; Brooks, B.R. New Spherical-cutoff Methods for Long-range Forces in Macromolecular Simulation. J. Comput. Chem. 1994, 15, 667–683. [Google Scholar] [CrossRef]
- Darden, T.; York, D.; Pedersen, L. Particle Mesh Ewald: An N·log(N) Method for Ewald Sums in Large Systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef]
- Roe, D.R.; Cheatham, T.E. PTRAJ and CPPTRAJ: Software for Processing and Analysis of Molecular Dynamics Trajectory Data. J. Chem. Theory Comput. 2013, 9, 3084–3095. [Google Scholar] [CrossRef]
- Roberts, C.J. Therapeutic Protein Aggregation: Mechanisms, Design, and Control. In Trends in Biotechnology; Elsevier Ltd.: Amsterdam, The Netherlands, 2014; pp. 372–380. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition of Formulations | Saccharide Used | Saccharide Concentration [mM] | S/P Ratio | Solid Content [wt.%] |
|---|---|---|---|---|
| Tryp_blank | - | - | - | 9.9 |
| TD_Tryp_HR | TD | 400 | 95.2 | 25.1 |
| TD_Tryp_MR | TD | 200 | 47.6 | 17.6 |
| TD_Tryp_LR | TD | 50 | 11.9 | 11.9 |
| HP_Tryp_HR | HP | 75.6 | 18.0 | 25.0 |
| HP_Tryp_MR | HP | 37.8 | 9.00 | 17.5 |
| HP_Tryp_LR | HP | 1.5 | 0.36 | 13.0 |
| HPB_Tryp_HR | HPB | 75.6 | 18.0 | 21.2 |
| HPB_Tryp_MR | HPB | 37.8 | 9.00 | 15.6 |
| HPB_Tryp_LR | HPB | 1.5 | 0.36 | 10.2 |
| DEX_Tryp_HR | DEX | 3.75 | 0.89 | 20.6 |
| DEX_Tryp_MR | DEX | 1.85 | 0.44 | 15.3 |
| DEX_Tryp_LR | DEX | 0.75 | 0.18 | 10.2 |
| Cat_blank | - | - | - | 10.1 |
| TD_Cat_HR | TD | 400 | 1000 | 25.3 |
| TD_Cat_MR | TD | 200 | 500 | 17.6 |
| TD_Cat_LR | TD | 50 | 125 | 12.1 |
| HP_Cat_HR | HP | 75.6 | 189 | 25.0 |
| HP_Cat_MR | HP | 37.8 | 94.5 | 17.5 |
| HP_Cat_LR | HP | 1.5 | 3.75 | 12.9 |
| HPB_Cat_HR | HPB | 75.6 | 189 | 17.2 |
| HPB _Cat_MR | HPB | 37.8 | 94.5 | 14.9 |
| HPB _Cat_LR | HPB | 1.5 | 3.75 | 10.2 |
| DEX_Cat_HR | DEX | 3.75 | 9.38 | 20.5 |
| DEX_ Cat_MR | DEX | 1.85 | 4.63 | 15.2 |
| DEX_Cat_LR | DEX | 0.75 | 1.88 | 10.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dieplinger, J.; Moser, C.; König, G.; Pinto, J.T.; Paudel, A. Investigation of the Impact of Saccharides on the Relative Activity of Trypsin and Catalase after Droplet and Spray Drying. Pharmaceutics 2023, 15, 2504. https://doi.org/10.3390/pharmaceutics15102504
Dieplinger J, Moser C, König G, Pinto JT, Paudel A. Investigation of the Impact of Saccharides on the Relative Activity of Trypsin and Catalase after Droplet and Spray Drying. Pharmaceutics. 2023; 15(10):2504. https://doi.org/10.3390/pharmaceutics15102504
Chicago/Turabian StyleDieplinger, Johanna, Christina Moser, Gerhard König, Joana T. Pinto, and Amrit Paudel. 2023. "Investigation of the Impact of Saccharides on the Relative Activity of Trypsin and Catalase after Droplet and Spray Drying" Pharmaceutics 15, no. 10: 2504. https://doi.org/10.3390/pharmaceutics15102504