
Effect of Sex and Cross-Sex Hormone Treatment on Renal Monocarboxylate-Transporter Expression in Rats

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals, Hormone Treatment and Tissue Collection
2.3. Quantification of Estrogen, Progesterone and Testosterone in Plasma
2.4. qPCR
2.5. Western Blot Analysis
2.6. Data Analysis
3. Results
3.1. Sex Hormone Concentrations
3.2. MCT1 Expression
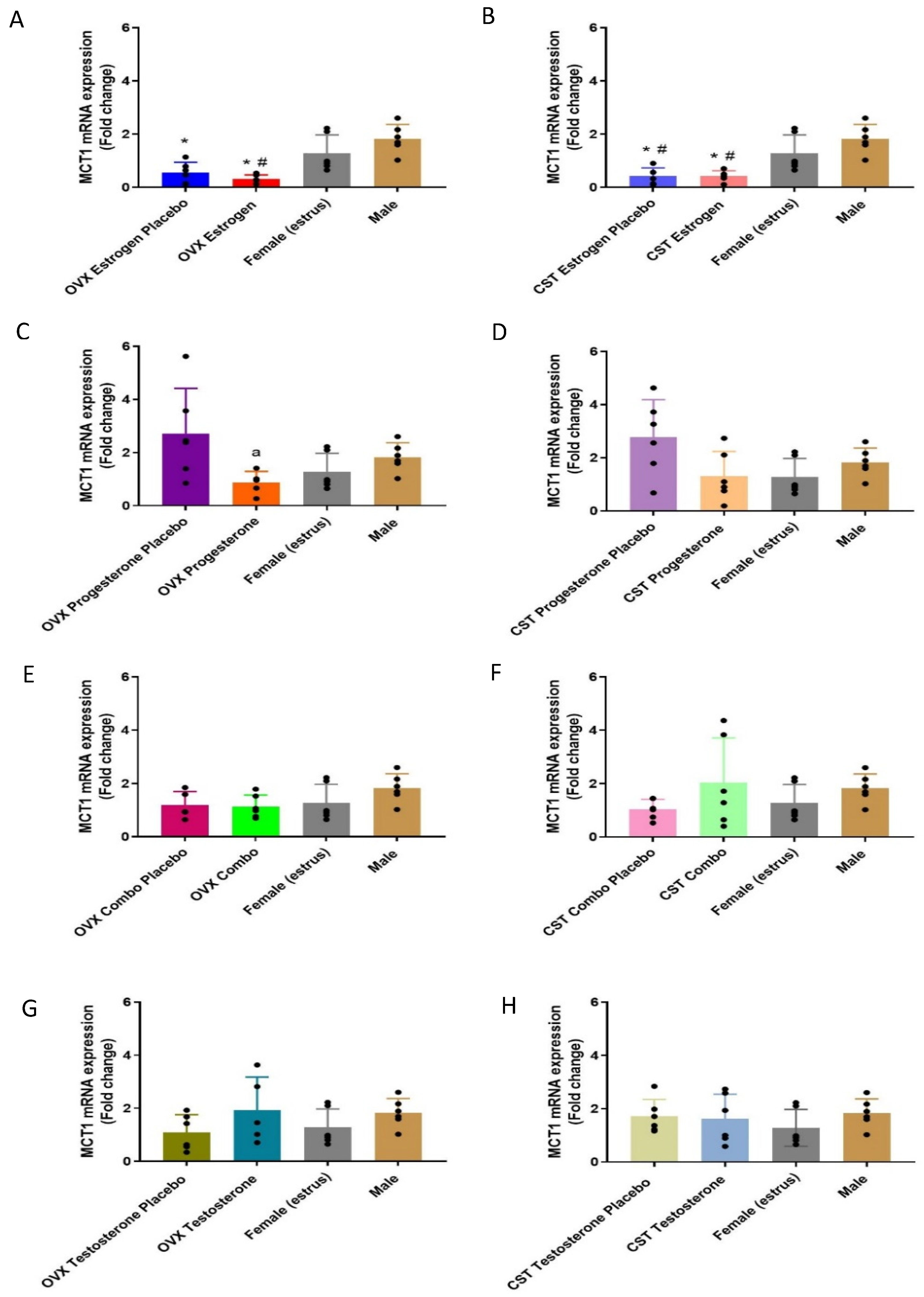
3.2.1. mRNA Expression
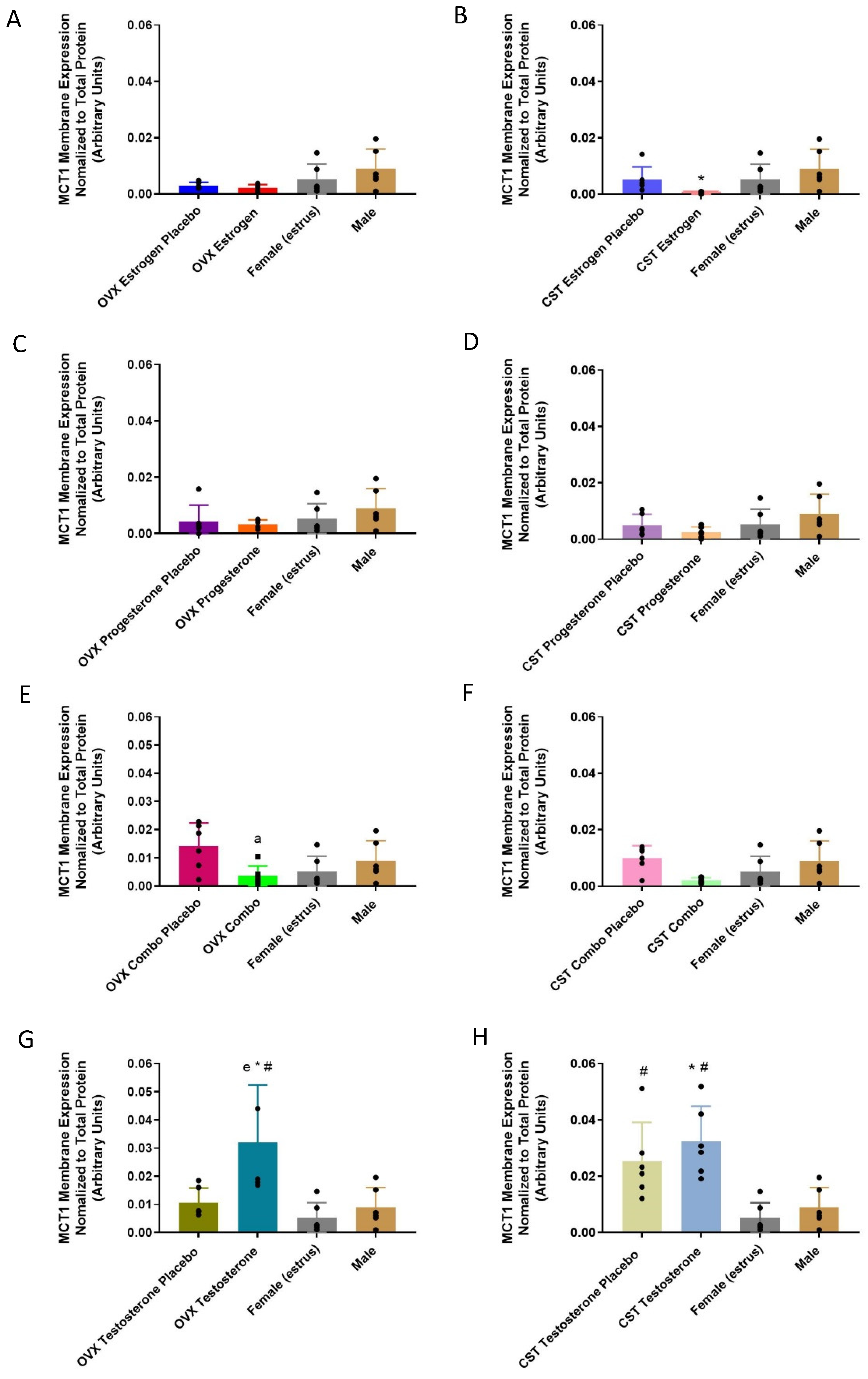
3.2.2. Membrane-Bound Protein Expression
3.3. MCT4 Expression
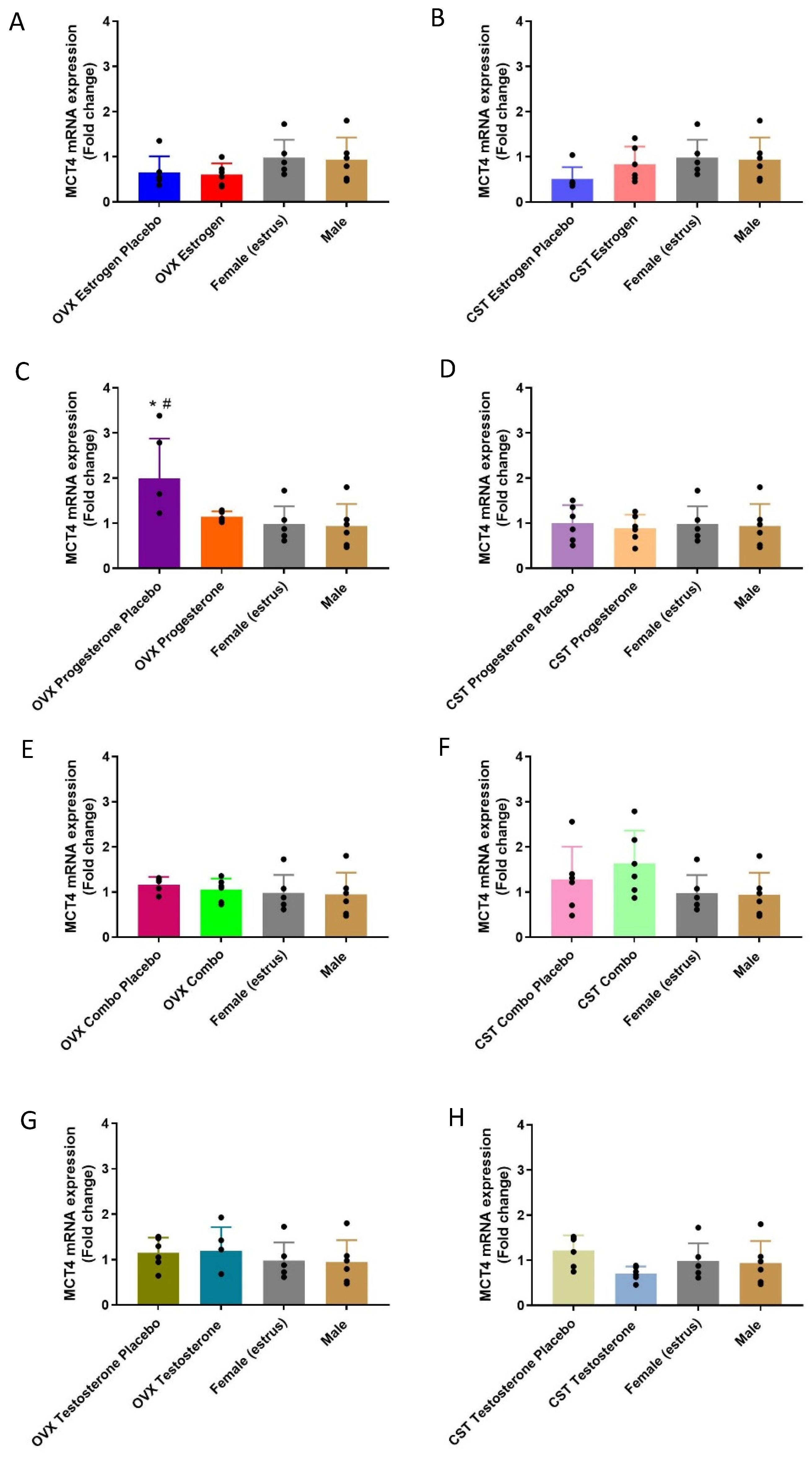
3.3.1. mRNA Expression
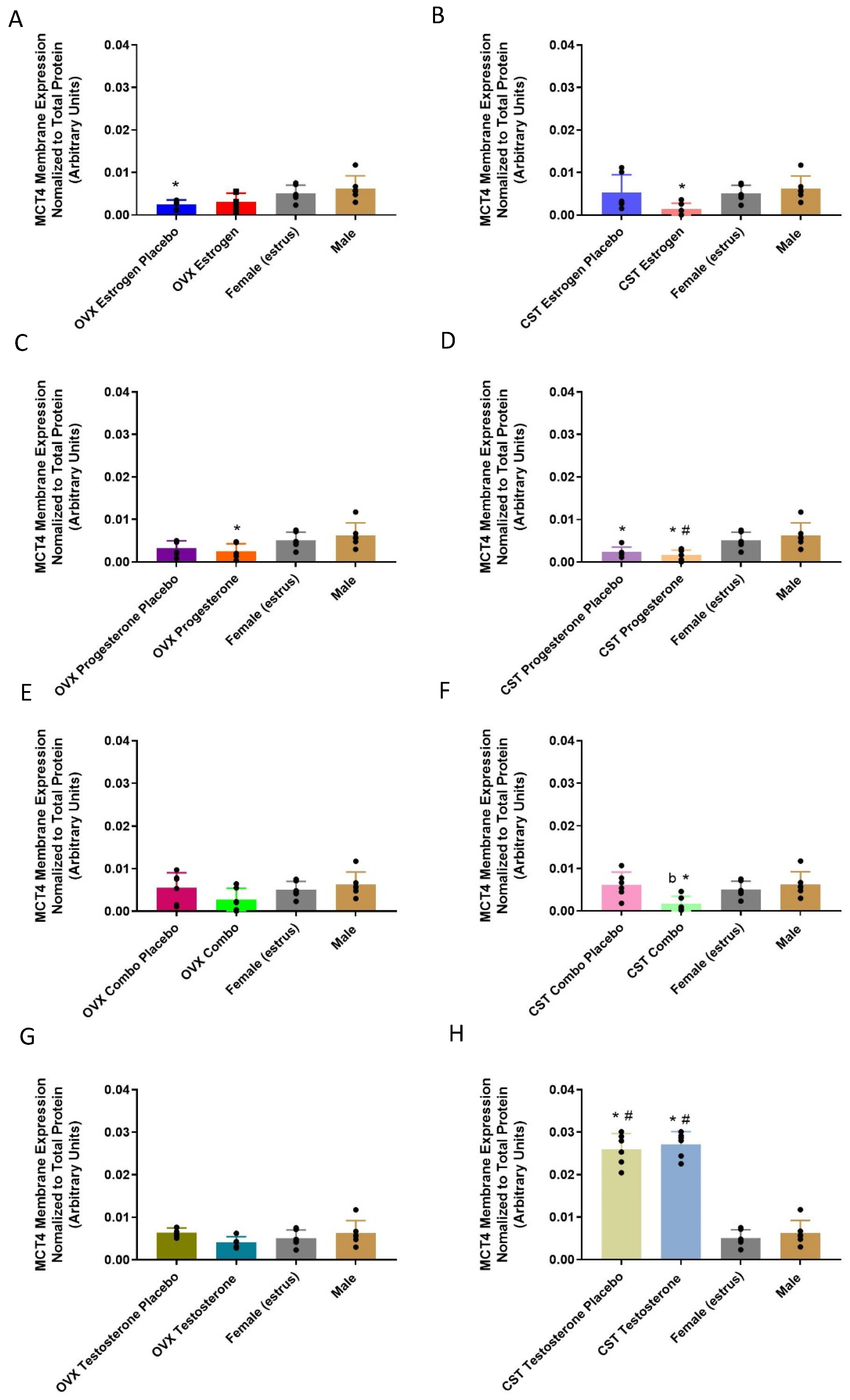
3.3.2. Membrane-Bound Protein Expression
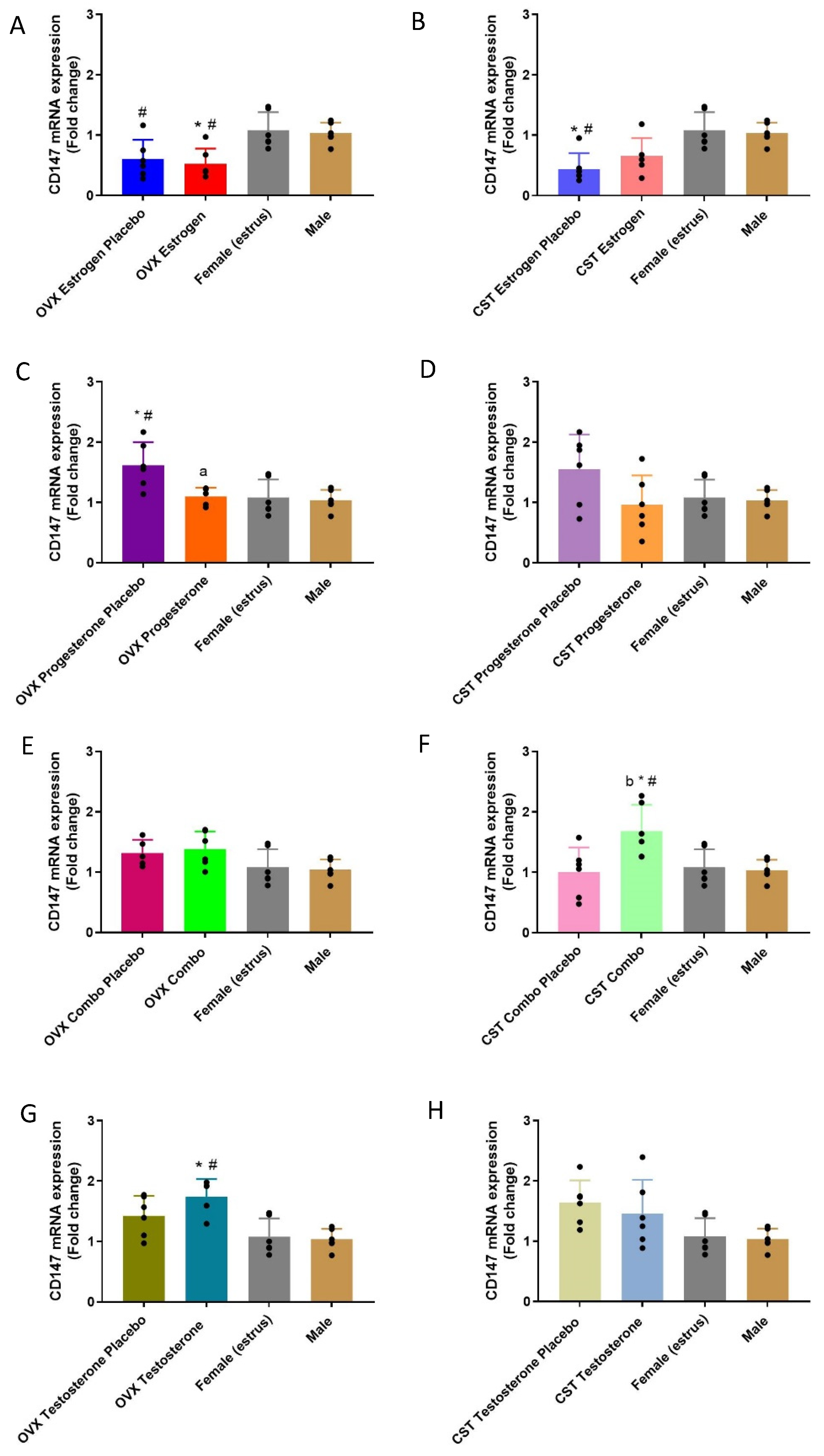
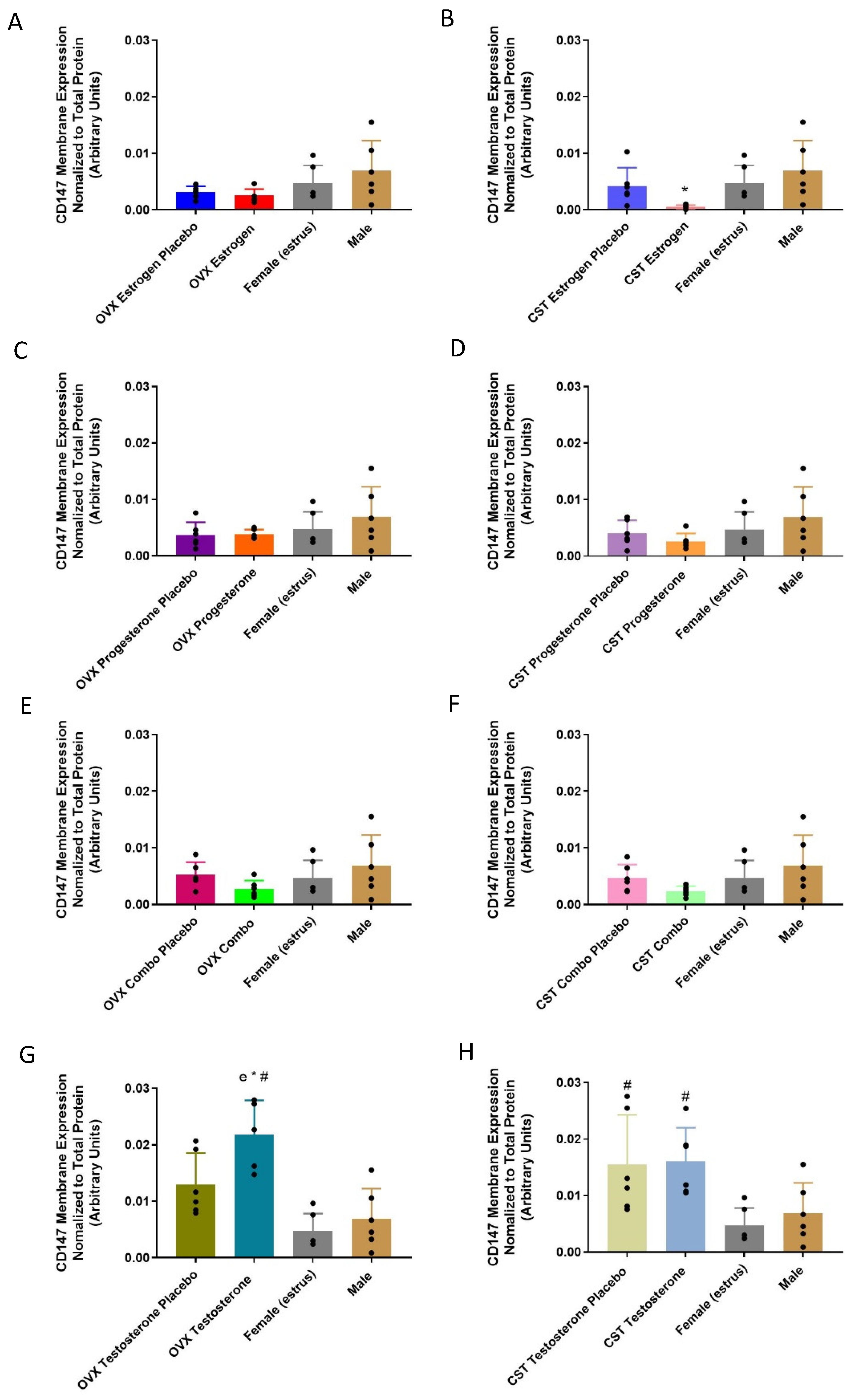
3.4. CD147 Expression
3.4.1. mRNA Expression
3.4.2. Membrane-Bound Protein Expression
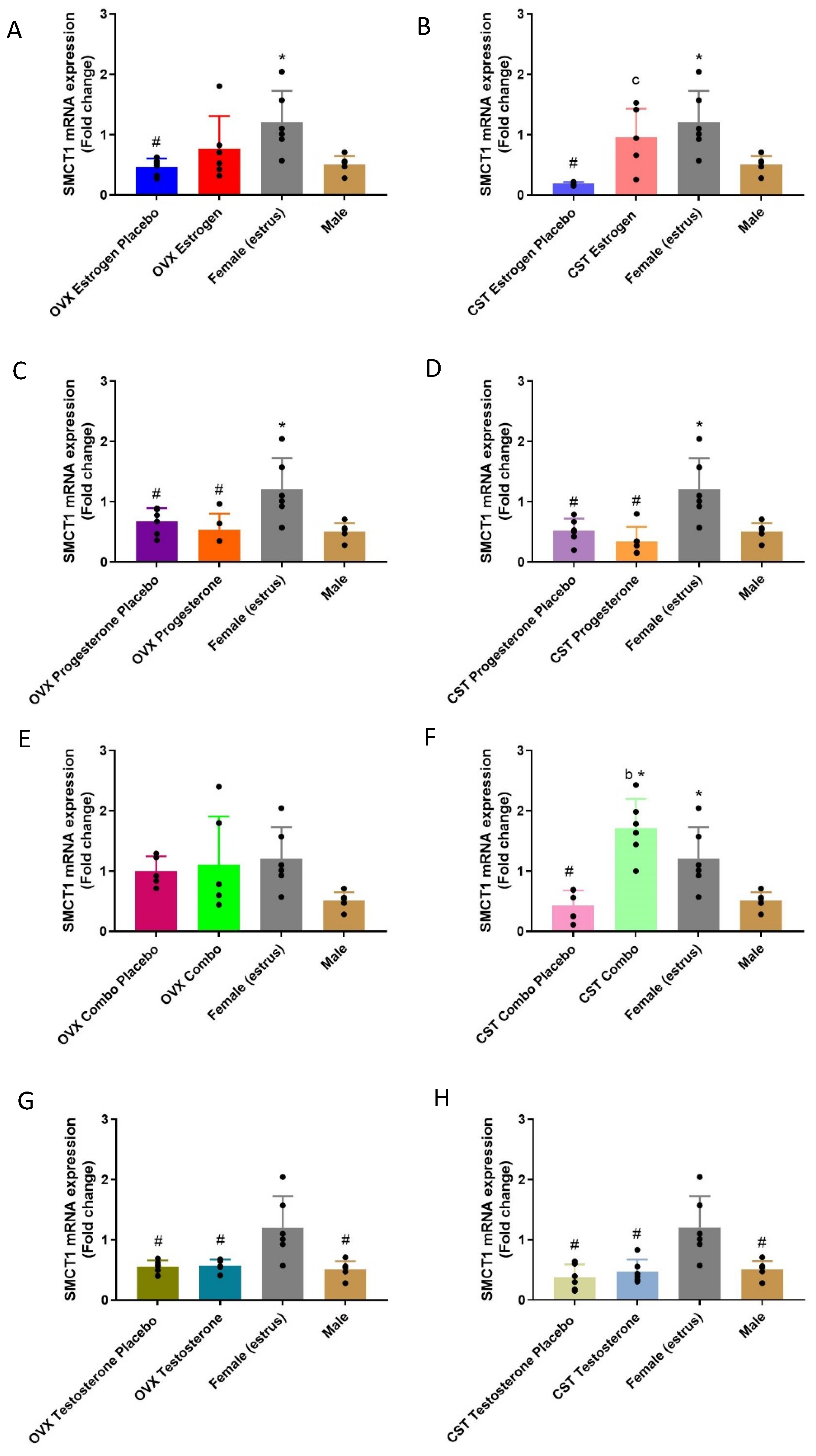
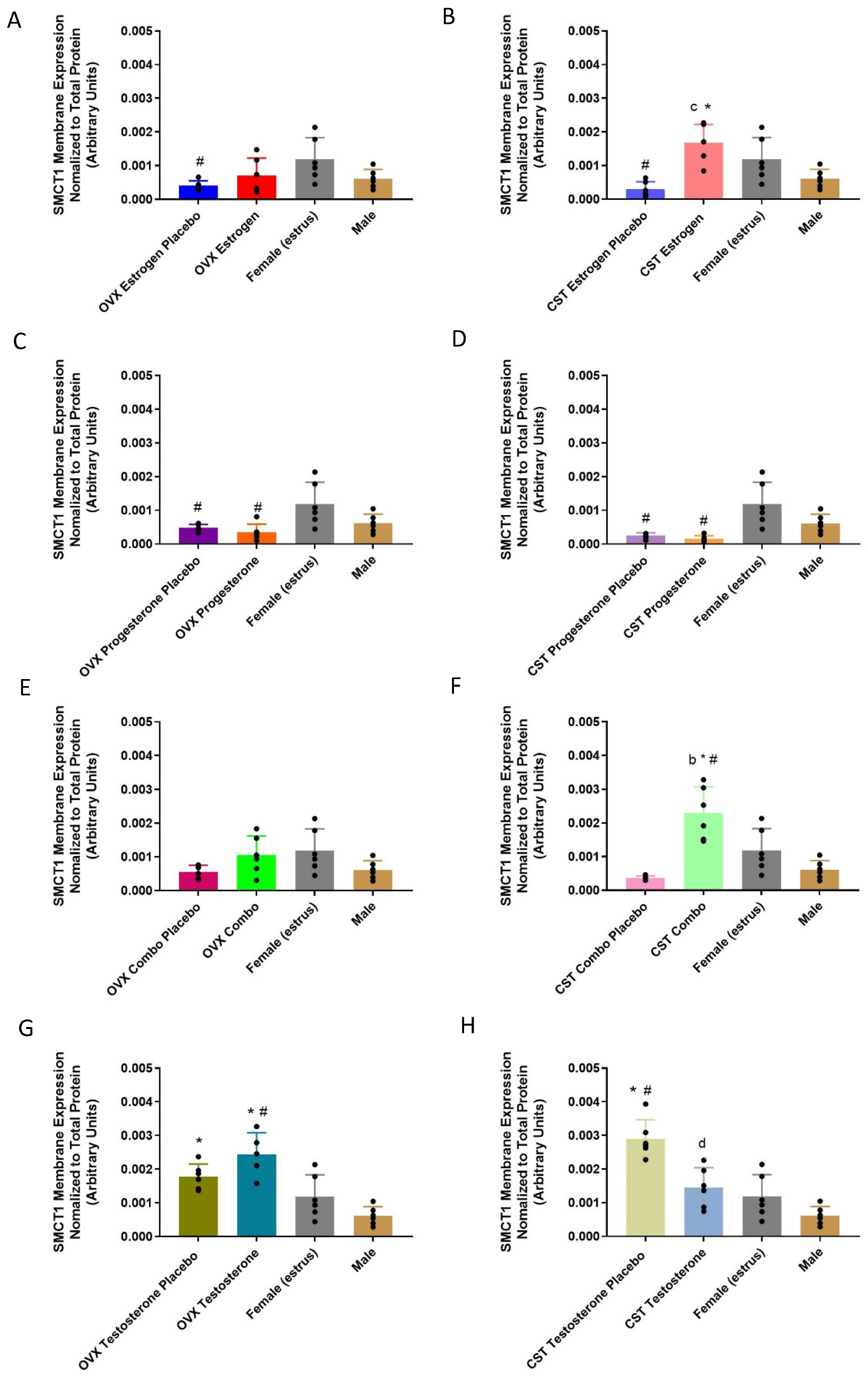
3.5. SMCT1 Expression
3.5.1. mRNA Expression
3.5.2. Membrane-Bound Protein Expression
3.6. Correlation Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Halestrap, A.P. The SLC16 gene family—Structure, role and regulation in health and disease. Mol. Asp. Med. 2013, 34, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Felmlee, M.A.; Jones, R.S.; Rodriguez-Cruz, V.; Follman, K.E.; Morris, M.E. Monocarboxylate Transporters (SLC16): Function, Regulation, and Role in Health and Disease. Pharmacol. Rev. 2020, 72, 466–485. [Google Scholar] [CrossRef] [PubMed]
- Ganapathy, V.; Thangaraju, M.; Gopal, E.; Martin, P.M.; Itagaki, S.; Miyauchi, S.; Prasad, P.D. Sodium-coupled monocarboxylate transporters in normal tissues and in cancer. Aaps. J. 2008, 10, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Halestrap, A.P.; Meredith, D. The SLC16 gene family-from monocarboxylate transporters (MCTs) to aromatic amino acid transporters and beyond. Pflugers Arch. 2004, 447, 619–628. [Google Scholar] [CrossRef]
- Dave, R.A.; Morris, M.E. Semi-mechanistic kidney model incorporating physiologically-relevant fluid reabsorption and transporter-mediated renal reabsorption: Pharmacokinetics of γ-hydroxybutyric acid and L-lactate in rats. J. Pharmacokinet. Pharmacodyn. 2015, 42, 497–513. [Google Scholar] [CrossRef]
- Adijanto, J.; Philp, N.J. The SLC16A family of monocarboxylate transporters (MCTs)--physiology and function in cellular metabolism, pH homeostasis, and fluid transport. Curr. Top. Membr. 2012, 70, 275–311. [Google Scholar] [CrossRef]
- Becker, H.M.; Mohebbi, N.; Perna, A.; Ganapathy, V.; Capasso, G.; Wagner, C.A. Localization of members of MCT monocarboxylate transporter family Slc16 in the kidney and regulation during metabolic acidosis. Am. J. Physiol. Renal. Physiol. 2010, 299, F141–F154. [Google Scholar] [CrossRef]
- Felmlee, M.A.; Morse, B.L.; Morris, M.E. gamma-Hydroxybutyric Acid: Pharmacokinetics, Pharmacodynamics, and Toxicology. AAPS J. 2021, 23, 22. [Google Scholar] [CrossRef]
- Garcia, C.K.; Brown, M.S.; Pathak, R.K.; Goldstein, J.L. cDNA cloning of MCT2, a second monocarboxylate transporter expressed in different cells than MCT1. J. Biol. Chem. 1995, 270, 1843–1849. [Google Scholar] [CrossRef]
- Deora, A.A.; Philp, N.; Hu, J.; Bok, D.; Rodriguez-Boulan, E. Mechanisms regulating tissue-specific polarity of monocarboxylate transporters and their chaperone CD147 in kidney and retinal epithelia. Proc. Natl. Acad. Sci. USA 2005, 102, 16245–16250. [Google Scholar] [CrossRef]
- Gopal, E.; Umapathy, N.S.; Martin, P.M.; Ananth, S.; Gnana-Prakasam, J.P.; Becker, H.; Wagner, C.A.; Ganapathy, V.; Prasad, P.D. Cloning and functional characterization of human SMCT2 (SLC5A12) and expression pattern of the transporter in kidney. Biochim. Biophys. Acta 2007, 1768, 2690–2697. [Google Scholar] [CrossRef]
- Kirk, P.; Wilson, M.C.; Heddle, C.; Brown, M.H.; Barclay, A.N.; Halestrap, A.P. CD147 is tightly associated with lactate transporters MCT1 and MCT4 and facilitates their cell surface expression. Embo J. 2000, 19, 3896–3904. [Google Scholar] [CrossRef]
- Le Floch, R.; Chiche, J.; Marchiq, I.; Naiken, T.; Ilc, K.; Murray, C.M.; Critchlow, S.E.; Roux, D.; Simon, M.P.; Pouysségur, J. CD147 subunit of lactate/H+ symporters MCT1 and hypoxia-inducible MCT4 is critical for energetics and growth of glycolytic tumors. Proc. Natl. Acad. Sci. USA 2011, 108, 16663–16668. [Google Scholar] [CrossRef]
- Enoki, T.; Yoshida, Y.; Lally, J.; Hatta, H.; Bonen, A. Testosterone increases lactate transport, monocarboxylate transporter (MCT) 1 and MCT4 in rat skeletal muscle. J. Physiol. 2006, 577, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Manente, A.G.; Pinton, G.; Zonca, S.; Cilli, M.; Rinaldi, M.; Daga, A.; Nilsson, S.; Moro, L. Intracellular lactate-mediated induction of estrogen receptor beta (ERbeta) in biphasic malignant pleural mesothelioma cells. Oncotarget 2015, 6, 25121–25134. [Google Scholar] [CrossRef] [PubMed]
- Rato, L.; Alves, M.G.; Socorro, S.; Carvalho, R.A.; Cavaco, J.E.; Oliveira, P.F. Metabolic modulation induced by oestradiol and DHT in immature rat Sertoli cells cultured in vitro. Biosci. Rep. 2012, 32, 61–69. [Google Scholar] [CrossRef]
- Boussouar, F.; Mauduit, C.; Tabone, E.; Pellerin, L.; Magistretti, P.J.; Benahmed, M. Developmental and hormonal regulation of the monocarboxylate transporter 2 (MCT2) expression in the mouse germ cells. Biol. Reprod. 2003, 69, 1069–1078. [Google Scholar] [CrossRef]
- Takiue, Y.; Hosoyamada, M.; Kimura, M.; Saito, H. The effect of female hormones upon urate transport systems in the mouse kidney. Nucleosides Nucleotides Nucleic Acids 2011, 30, 113–119. [Google Scholar] [CrossRef]
- Cao, J. The Role of Sex Hormones on Monocarboxylate Transporter expression in Tissues Related to Drug Disposition. Ph.D. Thesis, Pharmaceutics and Medicinal Chemistry, University of the Pacific, Stockton, CA, USA, 2019. [Google Scholar]
- Hosoyamada, M.; Takiue, Y.; Shibasaki, T.; Saito, H. The effect of testosterone upon the urate reabsorptive transport system in mouse kidney. Nucleosides Nucleotides Nucleic Acids 2010, 29, 574–579. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Ng, M.; Felmlee, M.A. Sex Hormones Regulate Rat Hepatic Monocarboxylate Transporter Expression and Membrane Trafficking. J. Pharm. Pharm. Sci. 2017, 20, 435–444. [Google Scholar] [CrossRef]
- Juck, M.; Long, W. Determination of Hormones in Serum by LC/MS/MS Using Agilent Bond Elut Plexa SPE. 2017. Available online: https://www.agilent.com/cs/library/applications/5991-8042EN.pdf (accessed on 6 September 2022).
- Parini, P.; Angelin, B.; Stavréus-Evers, A.; Freyschuss, B.; Eriksson, H.; Rudling, M. Biphasic effects of the natural estrogen 17beta-estradiol on hepatic cholesterol metabolism in intact female rats. Arter. Thromb. Vasc. Biol. 2000, 20, 1817–1823. [Google Scholar] [CrossRef] [PubMed]
- Haim, S.; Shakhar, G.; Rossene, E.; Taylor, A.N.; Ben-Eliyahu, S. Serum levels of sex hormones and corticosterone throughout 4- and 5-day estrous cycles in Fischer 344 rats and their simulation in ovariectomized females. J. Endocrinol. Investig. 2003, 26, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- Lan, H.C.; Lin, J.H.; Chen, C.H.; Chu, P.W.; Cheng, C.P. Estradiol Level of Male Rat is Correlated with Depression and Anxiety after Traumatic Stress. J. Med. Sci. 2018, 38, 176–187. [Google Scholar] [CrossRef]
- Damassa, D.A.; Smith, E.R.; Tennent, B.; Davidson, J.M. The relationship between circulating testosterone levels and male sexual behavior in rats. Horm Behav. 1977, 8, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Shulman, L.M.; Spritzer, M.D. Changes in the sexual behavior and testosterone levels of male rats in response to daily interactions with estrus females. Physiol. Behav. 2014, 133, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Cao, J.; Fallert, T.; Yeo, S.; Felmlee, M. GHB Toxicokinetics and Renal Monocarboxylate Transporter Expression are Influenced by the Estrus Cycle in Rats. BMC Pharmacol. Toxicol. 2023; preprint. [Google Scholar] [CrossRef]
- Ng, M.; Louie, J.; Cao, J.; Felmlee, M.A. Developmental Expression of Monocarboxylate Transporter 1 and 4 in Rat Liver. J. Pharm. Pharm. Sci. 2019, 22, 376–387. [Google Scholar] [CrossRef]
- Moeller, H.; Goecke, B.; Herter, F. Seasonal and diurnal changes of prostatic androgen receptor and circulating testosterone in young mature rats. Res. Exp. Med. 1988, 188, 451–462. [Google Scholar] [CrossRef]
- Kinson, G.A.; Liu, C.C. Diurnal variation in plasma testosterone of the male laboratory rat. Horm Metab. Res. 1973, 5, 233–234. [Google Scholar] [CrossRef]
- Aspatwar, A.; Tolvanen, M.E.E.; Schneider, H.P.; Becker, H.M.; Narkilahti, S.; Parkkila, S.; Deitmer, J.W. Catalytically inactive carbonic anhydrase-related proteins enhance transport of lactate by MCT1. FEBS Open Bio 2019, 9, 1204–1211. [Google Scholar] [CrossRef]
- Wilson, M.C.; Meredith, D.; Halestrap, A.P. Fluorescence resonance energy transfer studies on the interaction between the lactate transporter MCT1 and CD147 provide information on the topology and stoichiometry of the complex in situ. J. Biol. Chem. 2002, 277, 3666–3672. [Google Scholar] [CrossRef]
- Hutson, D.D.; Gurrala, R.; Ogola, B.O.; Zimmerman, M.A.; Mostany, R.; Satou, R.; Lindsey, S.H. Estrogen receptor profiles across tissues from male and female Rattus norvegicus. Biol. Sex. Differ. 2019, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Kruijver, F.P.; Fernández-Guasti, A.; Fodor, M.; Kraan, E.M.; Swaab, D.F. Sex differences in androgen receptors of the human mamillary bodies are related to endocrine status rather than to sexual orientation or transsexuality. J. Clin. Endocrinol. Metab. 2001, 86, 818–827. [Google Scholar] [CrossRef] [PubMed]
- Brundin, P.M.A.; Landgren, B.M.; Fjällström, P.; Shamekh, M.M.; Gustafsson, J.; Johansson, A.F.; Nalvarte, I. Expression of Sex Hormone Receptor and Immune Response Genes in Peripheral Blood Mononuclear Cells During the Menstrual Cycle. Front Endocrinol. 2021, 12, 721813. [Google Scholar] [CrossRef] [PubMed]
- Pullen, T.J.; da Silva Xavier, G.; Kelsey, G.; Rutter, G.A. miR-29a and miR-29b contribute to pancreatic beta-cell-specific silencing of monocarboxylate transporter 1 (Mct1). Mol. Cell. Biol. 2011, 31, 3182–3194. [Google Scholar] [CrossRef]
- Sengupta, P. The Laboratory Rat: Relating Its Age with Human’s. Int. J. Prev. Med. 2013, 4, 624–630. [Google Scholar]
- Picut, C.A.; Remick, A.K.; Asakawa, M.G.; Simons, M.L.; Parker, G.A. Histologic features of prepubertal and pubertal reproductive development in female Sprague-Dawley rats. Toxicol. Pathol. 2014, 42, 403–413. [Google Scholar] [CrossRef]
- Fuochi, S.; Galasso, M.E.; Colombo, R.; Giaquinto, D.; De Girolamo, P.; D’Angelo, L. Puberty onset curve in CD (Sprague Dawley) and Long Evans outbred male rats. Lab Anim 2022, 56, 471–475. [Google Scholar] [CrossRef]
- Feltenstein, M.W.; See, R.E. Plasma progesterone levels and cocaine-seeking in freely cycling female rats across the estrous cycle. Drug Alcohol Depend 2007, 89, 183–189. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Progesterone | Progesterone-d9 | Testosterone | Testosterone-d3 |
|---|---|---|---|---|
| Q1/Q3 (+) | 315.2/109.1 | 324.3/113.1 | 289.2/109.1 | 292.3/109.1 |
| Fragmentor (volts) | 100 | 117 | 112 | 107 |
| Collision energy (volts) | 26 | 30 | 26 | 26 |
| Cell acceleration voltage (volts) | 7 | 7 | 7 | 7 |
| Gene | Forward/Reverse Primer | Primer 5′–3′ | Size (bp) |
|---|---|---|---|
| MCT1 | Forward | GCTGTCATGTATGCCGGAG | 204 |
| Reverse | CAATCATAGTCAGAGCTGGG | ||
| CD147 | Forward | GGCACCATCGTAACCTCTGT | 211 |
| Reverse | CAGGCTCAGGAAGGAAGATG | ||
| MCT4 | Forward | GCTGGCTATGCTGTATGGC | 185 |
| Reverse | TTGAGAGCCAGACCCAAGC | ||
| SMCT1 | Forward | GTTGCTGGTGGGGATTCTTA | 200 |
| Reverse | CCACTGTGGTCTGGGAAGTT | ||
| r18s | Forward | GTTGGTTTTCGGAACTGAGGC | 206 |
| Reverse | GTCGGCATCGTTTATGGTCG |
| Plasma Hormone Level | 17β-Estradiol (pg/mL) | 17β-Estradiol Non-Detected | Progesterone (ng/mL) | Progesterone Non-Detected | Testosterone (ng/mL) | Testosterone Non-Detected |
|---|---|---|---|---|---|---|
| OVX Estrogen Placebo | 24.27 ± 6.674 | 0 out of 6 | 1.916 ± 1.120 | 1 out of 6 | 0 | 6 out of 6 |
| OVX Estrogen | 744.9 ± 406.1 | 0 out of 6 | 4.511 ± 3.841 | 0 out of 6 | 0 | 6 out of 6 |
| OVX Progesterone Placebo | 25.46 ± 14.63 | 0 out of 6 | 2.169 ± 2.242 | 2 out of 6 | 0 | 6 out of 6 |
| OVX Progesterone | 25.65 ± 11.32 | 0 out of 6 | 4.000 ± 1.673 | 0 out of 6 | 0 | 6 out of 6 |
| OVX Placebo Combo | 11.62 ± 7.645 | 0 out of 6 | 0.825 ± 0.284 | 3 out of 6 | 0 | 6 out of 6 |
| OVX Combo | 1104 ± 285.5 | 0 out of 6 | 6.241 ± 4.011 | 0 out of 6 | 0 | 6 out of 6 |
| OVX Testosterone placebo | -- | -- | 2.703 ± 2.127 | 2 out of 6 | 0 | 6 out of 6 |
| OVX Testosterone | -- | -- | 2.095 ± 1.063 | 1 out of 6 | 10.29 ± 1.206 | 1 out of 6 |
| Female (Estrus) | 20.97 ± 13.38 | 0 out of 6 | 8.026 ± 2.191 | 0 out of 6 | 0 | 6 out of 6 |
| CST Estrogen Placebo | 12.99 ± 7.311 | 0 out of 6 | 0.711 ± 0.179 | 2 out of 6 | 0 | 6 out of 6 |
| CST Estrogen | 1342 ± 691.2 | 0 out of 6 | 1.829 ± 0.922 | 1 out of 6 | 0 | 6 out of 6 |
| CST Progesterone Placebo | 24.48 ± 3.775 | 1 out of 6 | 0 | 6 out of 6 | 0 | 6 out of 6 |
| CST Progesterone | 12.81 ± 3.662 | 0 out of 6 | 6.815 ± 4.204 | 0 out of 6 | 0 | 6 out of 6 |
| CST Placebo Combo | 10.48 ± 4.896 | 0 out of 6 | 1.377 ± 0.689 | 1 out of 6 | 0 | 6 out of 6 |
| CST Combo | 1385 ± 846.7 | 0 out of 6 | 4.749 ± 2.423 | 0 out of 6 | 0 | 6 out of 6 |
| CST Testosterone placebo | -- | -- | 1.091 ± 1.091 | 3 out of 6 | 0 | 6 out of 6 |
| CST Testosterone | -- | -- | 1.143 ± 1.143 | 4 out of 6 | 7.505 ± 1.348 | 0 out of 6 |
| Male | 17.20 ± 5.161 | 0 out of 6 | 1.791 ± 1.496 | 3 out of 6 | 2.099 ± 0.404 | 1 out of 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, H.; Lee, A.; Zhang, Q.; Felmlee, M.A. Effect of Sex and Cross-Sex Hormone Treatment on Renal Monocarboxylate-Transporter Expression in Rats. Pharmaceutics 2023, 15, 2404. https://doi.org/10.3390/pharmaceutics15102404
Wei H, Lee A, Zhang Q, Felmlee MA. Effect of Sex and Cross-Sex Hormone Treatment on Renal Monocarboxylate-Transporter Expression in Rats. Pharmaceutics. 2023; 15(10):2404. https://doi.org/10.3390/pharmaceutics15102404
Chicago/Turabian StyleWei, Hao, Annie Lee, Qing Zhang, and Melanie A. Felmlee. 2023. "Effect of Sex and Cross-Sex Hormone Treatment on Renal Monocarboxylate-Transporter Expression in Rats" Pharmaceutics 15, no. 10: 2404. https://doi.org/10.3390/pharmaceutics15102404