

Phage Therapy against Staphylococcus aureus: Selection and Optimization of Production Protocols of Novel Broad-Spectrum Silviavirus Phages
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain Collection
2.2. Molecular Characterization of Bacterial Strains
2.3. Phage Isolation and Propagation
2.4. Ultracentrifugation
2.5. Phage Genome Sequencing and Bio-Informatic Analysis
2.6. Host Range Assessment
2.7. Transmission Electron Microscopy (TEM)
2.8. Optimization of Phage Production Protocols
2.9. Statistical Analysis
3. Results
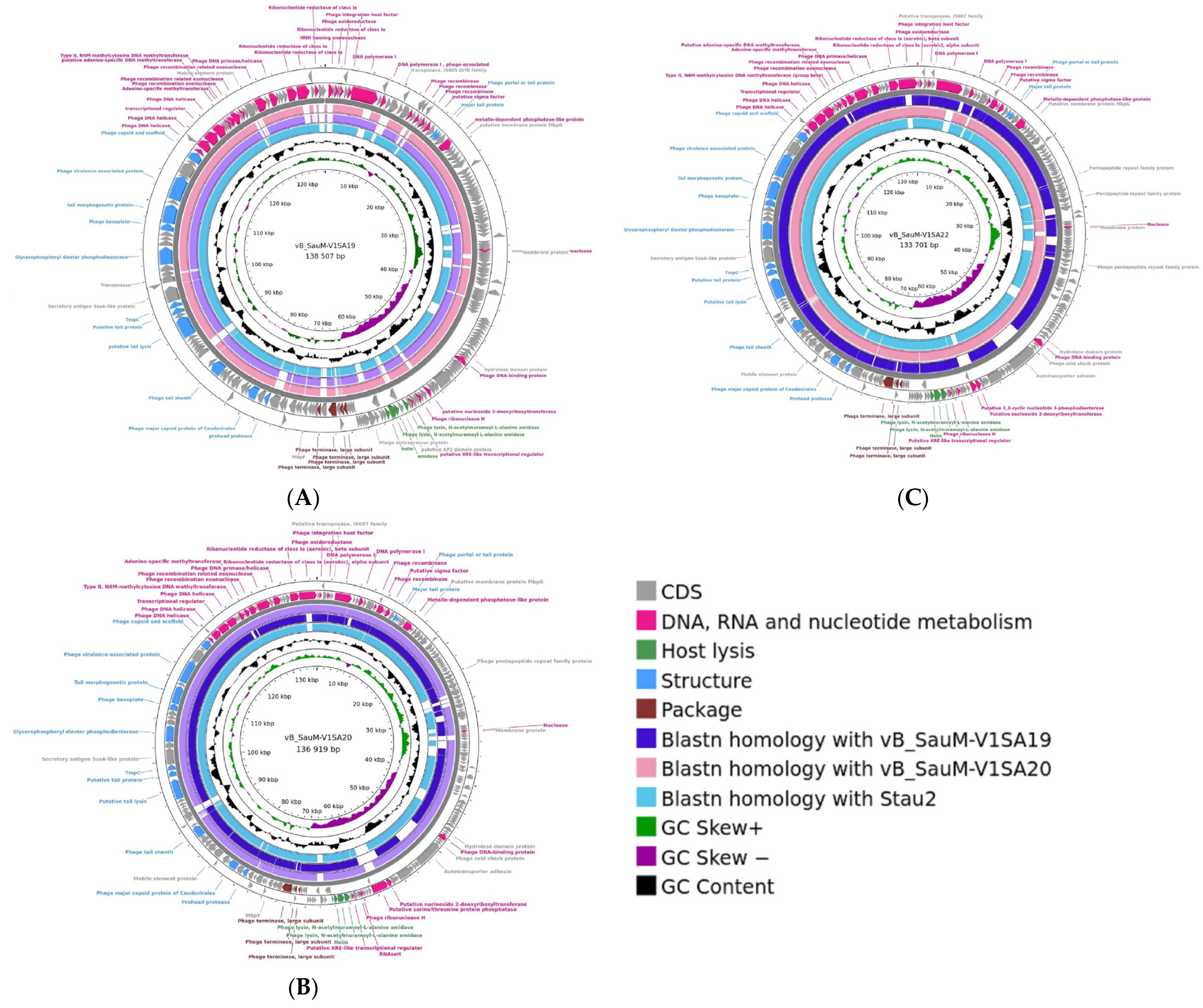
3.1. Phage Description
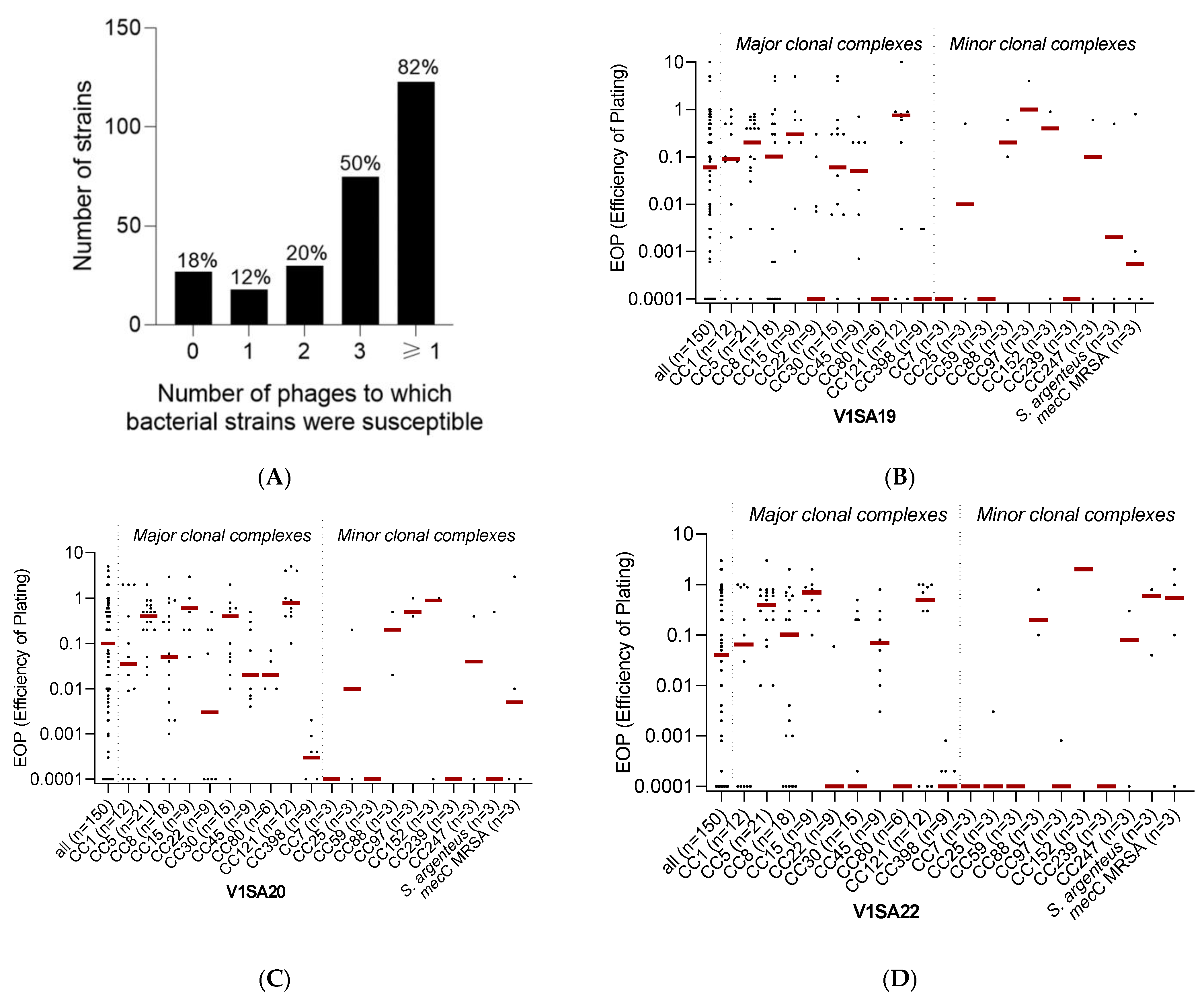
3.2. Host Range of Phages
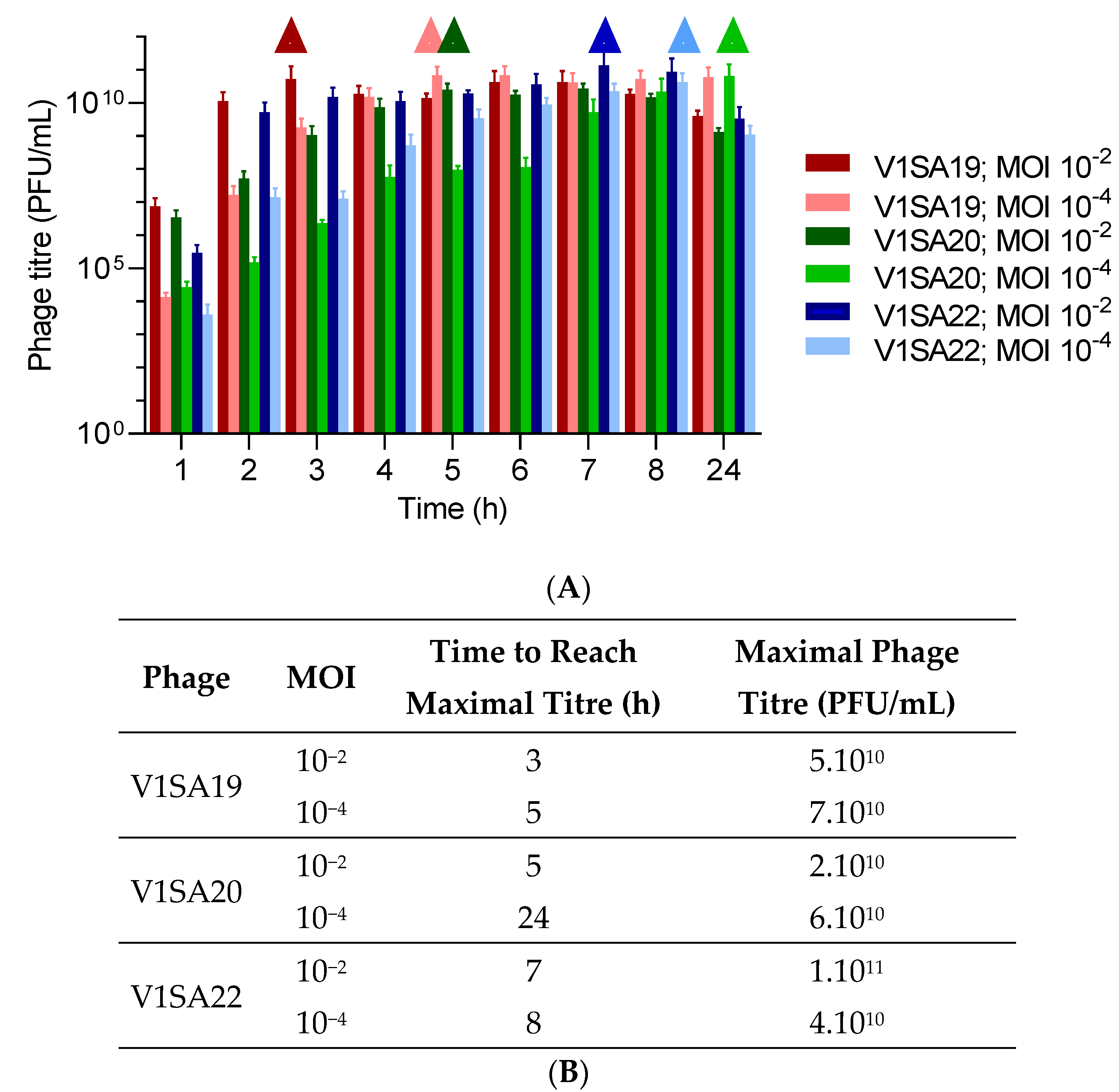
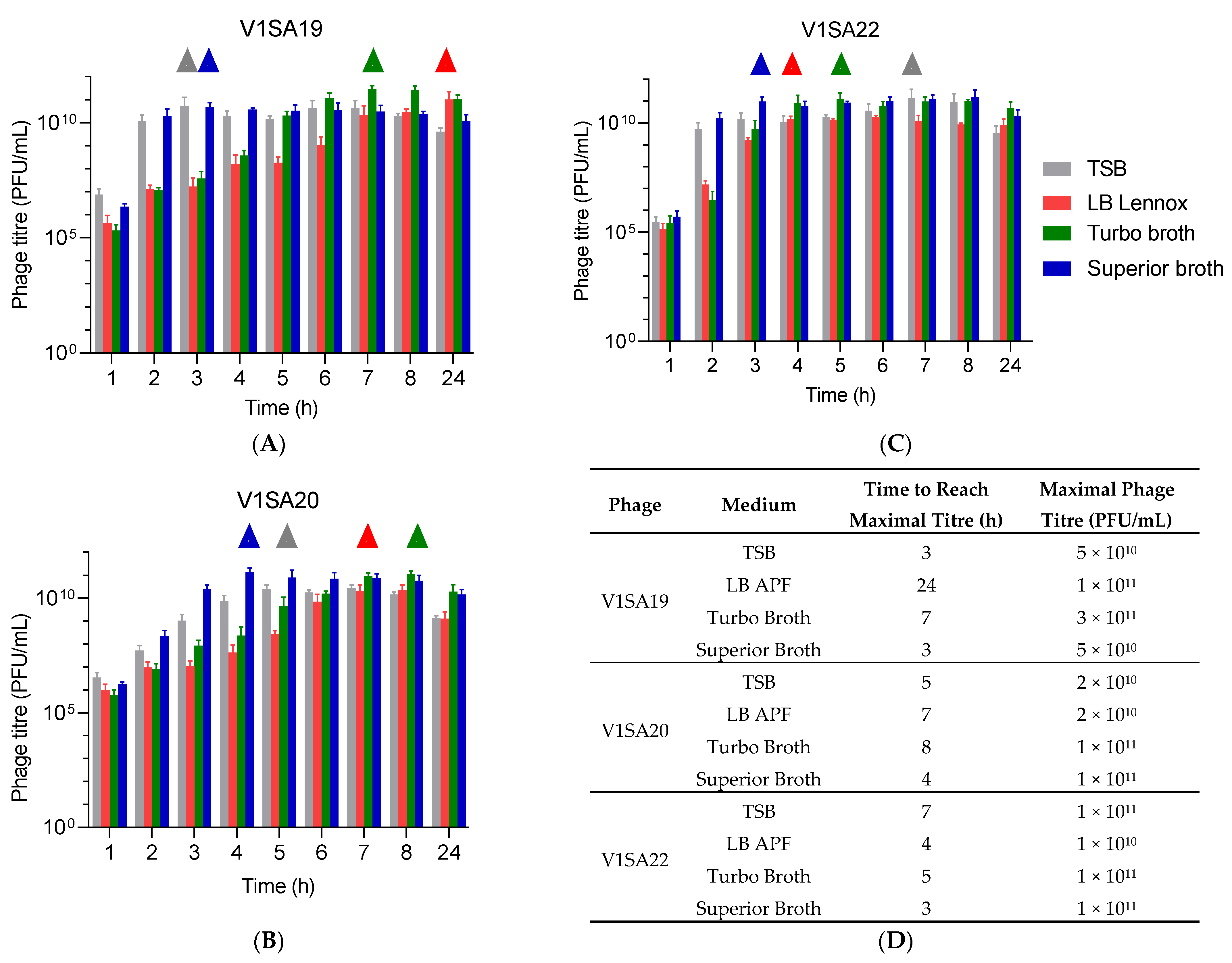
3.3. Optimization of Production Protocols
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus Aureus Infections: Epidemiology, Pathophysiology, Clinical Manifestations, and Management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, Research, and Development of New Antibiotics: The WHO Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Sanchez, C.J.; Mende, K.; Beckius, M.L.; Akers, K.S.; Romano, D.R.; Wenke, J.C.; Murray, C.K. Biofilm Formation by Clinical Isolates and the Implications in Chronic Infections. BMC Infect. Dis. 2013, 13, 47. [Google Scholar] [CrossRef] [PubMed]
- Nikolich, M.P.; Filippov, A.A. Bacteriophage Therapy: Developments and Directions. Antibiotics 2020, 9, 135. [Google Scholar] [CrossRef]
- Kolenda, C.; Josse, J.; Medina, M.; Fevre, C.; Lustig, S.; Ferry, T.; Laurent, F. Evaluation of the Activity of a Combination of Three Bacteriophages Alone or in Association with Antibiotics on Staphylococcus Aureus Embedded in Biofilm or Internalized in Osteoblasts. Antimicrob. Agents Chemother. 2020, 64, e02231-19. [Google Scholar] [CrossRef]
- Hsieh, S.-E.; Lo, H.-H.; Chen, S.-T.; Lee, M.-C.; Tseng, Y.-H. Wide Host Range and Strong Lytic Activity of Staphylococcus aureus Lytic Phage Stau2. Appl. Environ. Microbiol. 2011, 77, 756–761. [Google Scholar] [CrossRef]
- Botka, T.; Pantůček, R.; Mašlaňová, I.; Benešík, M.; Petráš, P.; Růžičková, V.; Havlíčková, P.; Varga, M.; Žemličková, H.; Koláčková, I.; et al. Lytic and Genomic Properties of Spontaneous Host-Range Kayvirus Mutants Prove Their Suitability for Upgrading Phage Therapeutics against Staphylococci. Sci. Rep. 2019, 9, 5475. [Google Scholar] [CrossRef]
- Vandersteegen, K.; Mattheus, W.; Ceyssens, P.-J.; Bilocq, F.; De Vos, D.; Pirnay, J.-P.; Noben, J.-P.; Merabishvili, M.; Lipinska, U.; Hermans, K.; et al. Microbiological and Molecular Assessment of Bacteriophage ISP for the Control of Staphylococcus Aureus. PLoS ONE 2011, 6, e24418. [Google Scholar] [CrossRef]
- Sáez Moreno, D.; Visram, Z.; Mutti, M.; Restrepo-Córdoba, M.; Hartmann, S.; Kremers, A.I.; Tišáková, L.; Schertler, S.; Wittmann, J.; Kalali, B.; et al. Ε2-Phages Are Naturally Bred and Have a Vastly Improved Host Range in Staphylococcus Aureus over Wild Type Phages. Pharmaceuticals 2021, 14, 325. [Google Scholar] [CrossRef]
- Gutiérrez, D.; Vandenheuvel, D.; Martínez, B.; Rodríguez, A.; Lavigne, R.; García, P. Two Phages, PhiIPLA-RODI and PhiIPLA-C1C, Lyse Mono- and Dual-Species Staphylococcal Biofilms. Appl. Environ. Microbiol. 2015, 81, 3336–3348. [Google Scholar] [CrossRef] [PubMed]
- Nagel, T.; Musila, L.; Muthoni, M.; Nikolich, M.; Nakavuma, J.L.; Clokie, M.R. Phage Banks as Potential Tools to Rapidly and Cost-Effectively Manage Antimicrobial Resistance in the Developing World. Curr. Opin. Virol. 2022, 53, 101208. [Google Scholar] [CrossRef] [PubMed]
- Mutti, M.; Corsini, L. Robust Approaches for the Production of Active Ingredient and Drug Product for Human Phage Therapy. Front. Microbiol. 2019, 10, 2289. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.G.; Kwon, J.; Giri, S.S.; Yun, S.; Kim, H.J.; Kim, S.W.; Kang, J.W.; Lee, S.B.; Jung, W.J.; Park, S.C. Strategy for Mass Production of Lytic Staphylococcus aureus Bacteriophage PSa-3: Contribution of Multiplicity of Infection and Response Surface Methodology. Microb. Cell Fact. 2021, 20, 56. [Google Scholar] [CrossRef] [PubMed]
- Rohde, C.; Resch, G.; Pirnay, J.-P.; Blasdel, B.; Debarbieux, L.; Gelman, D.; Górski, A.; Hazan, R.; Huys, I.; Kakabadze, E.; et al. Expert Opinion on Three Phage Therapy Related Topics: Bacterial Phage Resistance, Phage Training and Prophages in Bacterial Production Strains. Viruses 2018, 10, 178. [Google Scholar] [CrossRef]
- Laumay, F.; Corvaglia, A.-R.; Diene, S.M.; Girard, M.; Oechslin, F.; van der Mee-Marquet, N.; Entenza, J.M.; François, P. Temperate Prophages Increase Bacterial Adhesin Expression and Virulence in an Experimental Model of Endocarditis Due to Staphylococcus Aureus From the CC398 Lineage. Front. Microbiol. 2019, 10, 742. [Google Scholar] [CrossRef]
- Taylor, V.L.; Fitzpatrick, A.D.; Islam, Z.; Maxwell, K.L. The Diverse Impacts of Phage Morons on Bacterial Fitness and Virulence. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2019; Volume 103, pp. 1–31. ISBN 978-0-12-817722-8. [Google Scholar]
- Sweere, J.M.; Van Belleghem, J.D.; Ishak, H.; Bach, M.S.; Popescu, M.; Sunkari, V.; Kaber, G.; Manasherob, R.; Suh, G.A.; Cao, X.; et al. Bacteriophage Trigger Antiviral Immunity and Prevent Clearance of Bacterial Infection. Science 2019, 363, eaat9691. [Google Scholar] [CrossRef]
- Green, S.I.; Kaelber, J.T.; Ma, L.; Trautner, B.W.; Ramig, R.F.; Maresso, A.W. Bacteriophages from ExPEC Reservoirs Kill Pandemic Multidrug-Resistant Strains of Clonal Group ST131 in Animal Models of Bacteremia. Sci. Rep. 2017, 7, 46151. [Google Scholar] [CrossRef]
- van der Mee-Marquet, N.; Hernandez, D.; Bertrand, X.; Quentin, R.; Corvaglia, A.-R.; François, P. Whole-Genome Sequence of the Ancestral Animal-Borne ST398 Staphylococcus Aureus Strain S123. Genome Announc. 2013, 1, e00692-13. [Google Scholar] [CrossRef]
- Diene, S.M.; Corvaglia, A.R.; François, P.; van der Mee-Marquet, N. Prophages and Adaptation of Staphylococcus Aureus ST398 to the Human Clinic. BMC Genom. 2017, 18, 133. [Google Scholar] [CrossRef] [Green Version]
- Gillet, Y.; Tristan, A.; Rasigade, J.-P.; Saadatian-Elahi, M.; Bouchiat, C.; Bes, M.; Dumitrescu, O.; Leloire, M.; Dupieux, C.; Laurent, F.; et al. Prognostic Factors of Severe Community-Acquired Staphylococcal Pneumonia in France. Eur. Respir. J. 2021, 58, 2004445. [Google Scholar] [CrossRef] [PubMed]
- Leitner, L.; Ujmajuridze, A.; Chanishvili, N.; Goderdzishvili, M.; Chkonia, I.; Rigvava, S.; Chkhotua, A.; Changashvili, G.; McCallin, S.; Schneider, M.P.; et al. Intravesical Bacteriophages for Treating Urinary Tract Infections in Patients Undergoing Transurethral Resection of the Prostate: A Randomised, Placebo-Controlled, Double-Blind Clinical Trial. Lancet Infect. Dis. 2021, 21, 427–436. [Google Scholar] [CrossRef]
- Ferry, T.; Kolenda, C.; Briot, T.; Lustig, S.; Leboucher, G.; Laurent, F.; Ferry, T.; Valour, F.; Perpoint, T.; Ader, F.; et al. Implementation of a Complex Bone and Joint Infection Phage Therapy Centre in France: Lessons to Be Learned after 4 Years’ Experience. Clin. Microbiol. Infect. 2022, 28, 145–146. [Google Scholar] [CrossRef]
- McCallin; Sacher; Zheng; Chan Current State of Compassionate Phage Therapy. Viruses 2019, 11, 343. [CrossRef]
- Lehman, S.; Mearns, G.; Rankin, D.; Cole, R.; Smrekar, F.; Branston, S.; Morales, S. Design and Preclinical Development of a Phage Product for the Treatment of Antibiotic-Resistant Staphylococcus Aureus Infections. Viruses 2019, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Moller, A.G.; Winston, K.; Ji, S.; Wang, J.; Hargita Davis, M.N.; Solís-Lemus, C.R.; Read, T.D. Genes Influencing Phage Host Range in Staphylococcus Aureus on a Species-Wide Scale. mSphere 2021, 6, e01263-20. [Google Scholar] [CrossRef]
- Göller, P.C.; Elsener, T.; Lorgé, D.; Radulovic, N.; Bernardi, V.; Naumann, A.; Amri, N.; Khatchatourova, E.; Coutinho, F.H.; Loessner, M.J.; et al. Multi-Species Host Range of Staphylococcal Phages Isolated from Wastewater. Nat. Commun. 2021, 12, 6965. [Google Scholar] [CrossRef]
- Pirnay, J.-P.; Blasdel, B.G.; Bretaudeau, L.; Buckling, A.; Chanishvili, N.; Clark, J.R.; Corte-Real, S.; Debarbieux, L.; Dublanchet, A.; De Vos, D.; et al. Quality and Safety Requirements for Sustainable Phage Therapy Products. Pharm. Res. 2015, 32, 2173–2179. [Google Scholar] [CrossRef]
- Malik, D.J. Approaches for Manufacture, Formulation, Targeted Delivery and Controlled Release of Phage-Based Therapeutics. Curr. Opin. Biotechnol. 2021, 68, 262–271. [Google Scholar] [CrossRef]
- El Haddad, L.; Ben Abdallah, N.; Plante, P.-L.; Dumaresq, J.; Katsarava, R.; Labrie, S.; Corbeil, J.; St-Gelais, D.; Moineau, S. Improving the Safety of Staphylococcus Aureus Polyvalent Phages by Their Production on a Staphylococcus Xylosus Strain. PLoS ONE 2014, 9, e102600. [Google Scholar] [CrossRef] [Green Version]
- Doub, J.B.; Ng, V.Y.; Johnson, A.; Amoroso, A.; Kottilil, S.; Wilson, E. Potential Use of Adjuvant Bacteriophage Therapy with Debridement, Antibiotics, and Implant Retention Surgery to Treat Chronic Prosthetic Joint Infections. Open Forum Infect. Dis. 2021, 8, ofab277. [Google Scholar] [CrossRef] [PubMed]
- Jérôme, G. Prophage in Phage Manufacturing: Is the Risk Overrated Compared to Other Therapies or Food? Antibiotics 2020, 9, 435. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, J.A. Genomic Variation and Evolution of Staphylococcus aureus. Int. J. Med. Microbiol. 2010, 300, 98–103. [Google Scholar] [CrossRef]
- Ingmer, H.; Gerlach, D.; Wolz, C. Temperate Phages of Staphylococcus aureus. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Euler, C.W.; Juncosa, B.; Ryan, P.A.; Deutsch, D.R.; McShan, W.M.; Fischetti, V.A. Targeted Curing of All Lysogenic Bacteriophage from Streptococcus Pyogenes Using a Novel Counter-Selection Technique. PLoS ONE 2016, 11, e0146408. [Google Scholar] [CrossRef]
- Ali, J.; Rafiq, Q.; Ratcliffe, E. A Scaled-down Model for the Translation of Bacteriophage Culture to Manufacturing Scale. Biotechnol. Bioeng. 2019, 116, 972–984. [Google Scholar] [CrossRef]
- Dalpke, A.; Frank, J.; Peter, M.; Heeg, K. Activation of Toll-Like Receptor 9 by DNA from Different Bacterial Species. Infect. Immun. 2006, 74, 940–946. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Species | ST | Description (Accession Numbers) | EOP | Production Yields (PFU/mL) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| V1SA19 | V1SA20 | V1SA22 | V1SA19 | V1SA20 | V1SA22 | |||||||
| MOI 1 | MOI 10−3 | MOI 1 | MOI 10−3 | MOI 1 | MOI 10−3 | |||||||
| P2SA39-ST20191845 | S. aureus | 398 | S123 [20] | 0 | 0 | 0 | NT | NT | NT | NT | NT | NT |
| P2SA40-ST20191846 | S. aureus | 398 | S124 [21] | 0 | 0 | 0 | NT | NT | NT | NT | NT | NT |
| P2SA225-ST20170423 | S. aureus | 6 | This study a | 0.8 | 1.3 | 0.4 | 8 × 109 | 4 × 1010 | 2 × 109 | 2 × 1010 | 0 | 0 |
| P2SA236-ST20111368 | S. aureus | 15 | [22] b | 0.9 | 1.1 | 4.3 | NT | NT | NT | NT | NT | NT |
| P2SA237-ST20111713 | S. aureus | 15 | [22] c | 0.2 | 1.2 | 15.0 | 5 × 109 | 4 × 1010 | 2 × 1010 | 3 × 1010 | 2 × 1010 | 5 × 1010 |
| P2SA238-ST20140414 | S. aureus | 15 | [22] d | 0.9 | 3.5 | 11.7 | NT | NT | NT | NT | NT | NT |
| P2SA239-ST20150287 | S. aureus | 582 | This study e | 0.4 | 0.5 | 3.8 | 1 × 109 | 3 × 108 | 4 × 109 | 2 × 106 | 4 × 109 | 4 × 109 |
| P2SA240-ST20210147 | S. aureus | 582 | This study f | 0.9 | 0.7 | 2.0 | 4 × 109 | 3 × 107 | 2 × 1010 | 3 × 106 | 1 × 1010 | 2 × 1010 |
| P2SAG2-ST20191229 | S. argenteus | 2250 | This study g | 0 | 0 | 2.2 | NT | NT | NT | NT | NT | NT |
| P2SAG3-ST20191235 | S. argenteus | 2793 | This study h | 0.7 | 0.2 | 2.5 | 5 × 108 | 2 × 106 | 2 × 109 | 2 × 109 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolenda, C.; Medina, M.; Bonhomme, M.; Laumay, F.; Roussel-Gaillard, T.; Martins-Simoes, P.; Tristan, A.; Pirot, F.; Ferry, T.; Laurent, F.; et al. Phage Therapy against Staphylococcus aureus: Selection and Optimization of Production Protocols of Novel Broad-Spectrum Silviavirus Phages. Pharmaceutics 2022, 14, 1885. https://doi.org/10.3390/pharmaceutics14091885
Kolenda C, Medina M, Bonhomme M, Laumay F, Roussel-Gaillard T, Martins-Simoes P, Tristan A, Pirot F, Ferry T, Laurent F, et al. Phage Therapy against Staphylococcus aureus: Selection and Optimization of Production Protocols of Novel Broad-Spectrum Silviavirus Phages. Pharmaceutics. 2022; 14(9):1885. https://doi.org/10.3390/pharmaceutics14091885
Chicago/Turabian StyleKolenda, Camille, Mathieu Medina, Mélanie Bonhomme, Floriane Laumay, Tiphaine Roussel-Gaillard, Patricia Martins-Simoes, Anne Tristan, Fabrice Pirot, Tristan Ferry, Frédéric Laurent, and et al. 2022. "Phage Therapy against Staphylococcus aureus: Selection and Optimization of Production Protocols of Novel Broad-Spectrum Silviavirus Phages" Pharmaceutics 14, no. 9: 1885. https://doi.org/10.3390/pharmaceutics14091885