
Synthesis, Formulation and Characterization of Immunotherapeutic Glycosylated Dendrimer/cGAMP Complexes for CD206 Targeted Delivery to M2 Macrophages in Cold Tumors

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
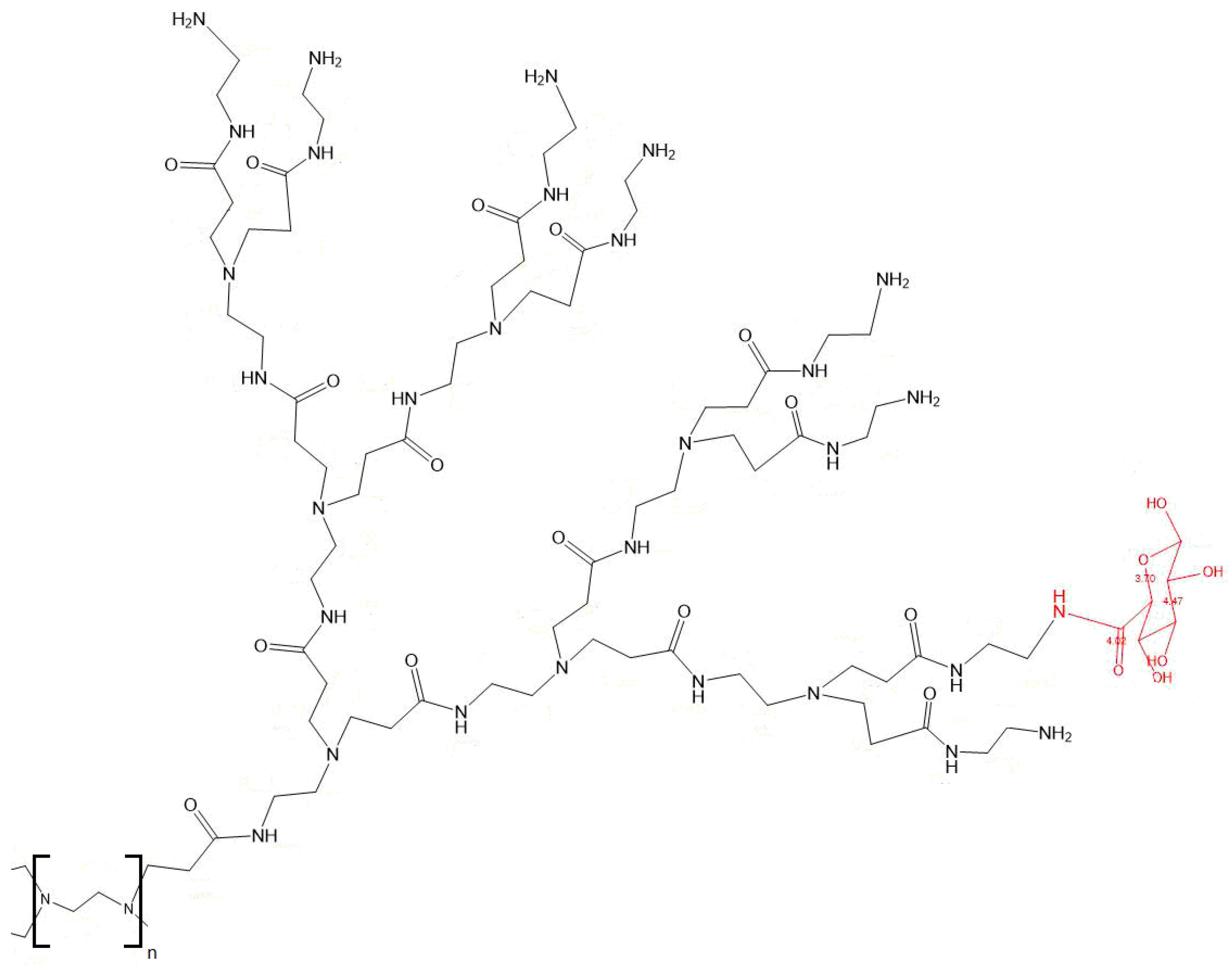
2.2. Synthesis of G3 and G4 PAMAM-D-Glucuronic Acid Derivatives
2.3. Physico-Chemical Characterization of PG3, PG4 and PG/cGAMP
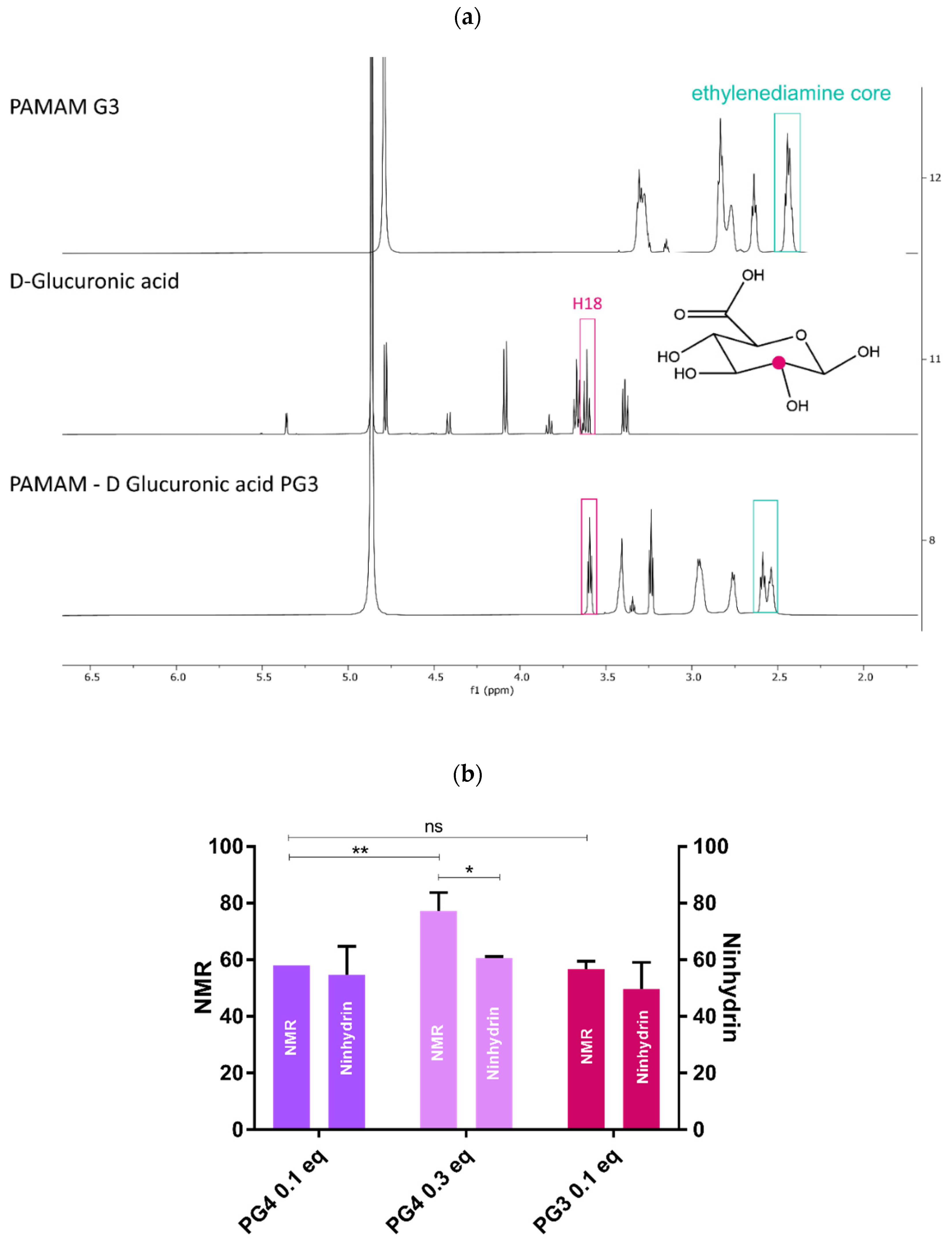
2.3.1. 1H NMR
2.3.2. Ninhydrin Assay
2.3.3. Rheology
2.3.4. Karl-Fischer
2.3.5. Dynamic Light Scattering (DLS)
2.3.6. Nanoparticle Tracking Analysis (NTA)
2.3.7. Asymmetric Flow Field Flow Fractionation (AF4)-RI-UV-MALS-DLS
2.3.8. Gravimetry
2.3.9. Light Microscopy
2.3.10. Scanning Electron Microscopy (SEM)
2.3.11. UPLC-UV for CE% Determination
2.3.12. Fabrication of PG/cGAMP NCs
2.3.13. MST—kD Determination
2.4. In Vitro Experimentation
2.4.1. Interferon β Level
- A 96-well plate was coated with 1/625 diluted 50 µL/well anti-IFN β, RMMB-1 (PBL, 22400-1) and incubated at 4 °C overnight;
- The next day, standard dilutions of mouse 1/840 dilution (from 1000 U/mL–7.8 U/mL) of IFN β (PBL, 12400-1) and samples were added (total 50 µL/well);
- Two hours later, the detection antibody, a 1/12,169 dilution of anti-IFN β (PBL, 32400-1), was added;
- Following this, the secondary antibody, anti-rabbit IgG, 1/1000 dilution HRP-linked (Cell Signaling, 7074S), was added;
- Reaction was induced by the addition of TMB substrate (BD OptEIA, 555214);
- It was stopped by the addition of 25 µL H2SO4;
- Optical absorbance was measured at 450 and 570 nm using a microplate reader (Biotek SynergyMx) and IFN beta levels are expressed as U/mL.
2.4.2. Flow Cytometry Analysis
- CD80-Pe (PE conjugated anti-mouse CD80 Antibody Biolegend);
- CD86-Bv510 (Brilliant Violet 510™ conjugated anti-mouse CD86 Antibody, Biolegend);
- Live/Dead Fixable Violet Dead Cell Stain (ThermoFisher)
- CD 206-FITC (FITC conjugated anti-mouse CD206, Biolegend);
- CD68-BV605 (Brilliant Violet 605 anti-mouse CD68, Biolegend).
2.4.3. Quantitative RT-PCR Assays
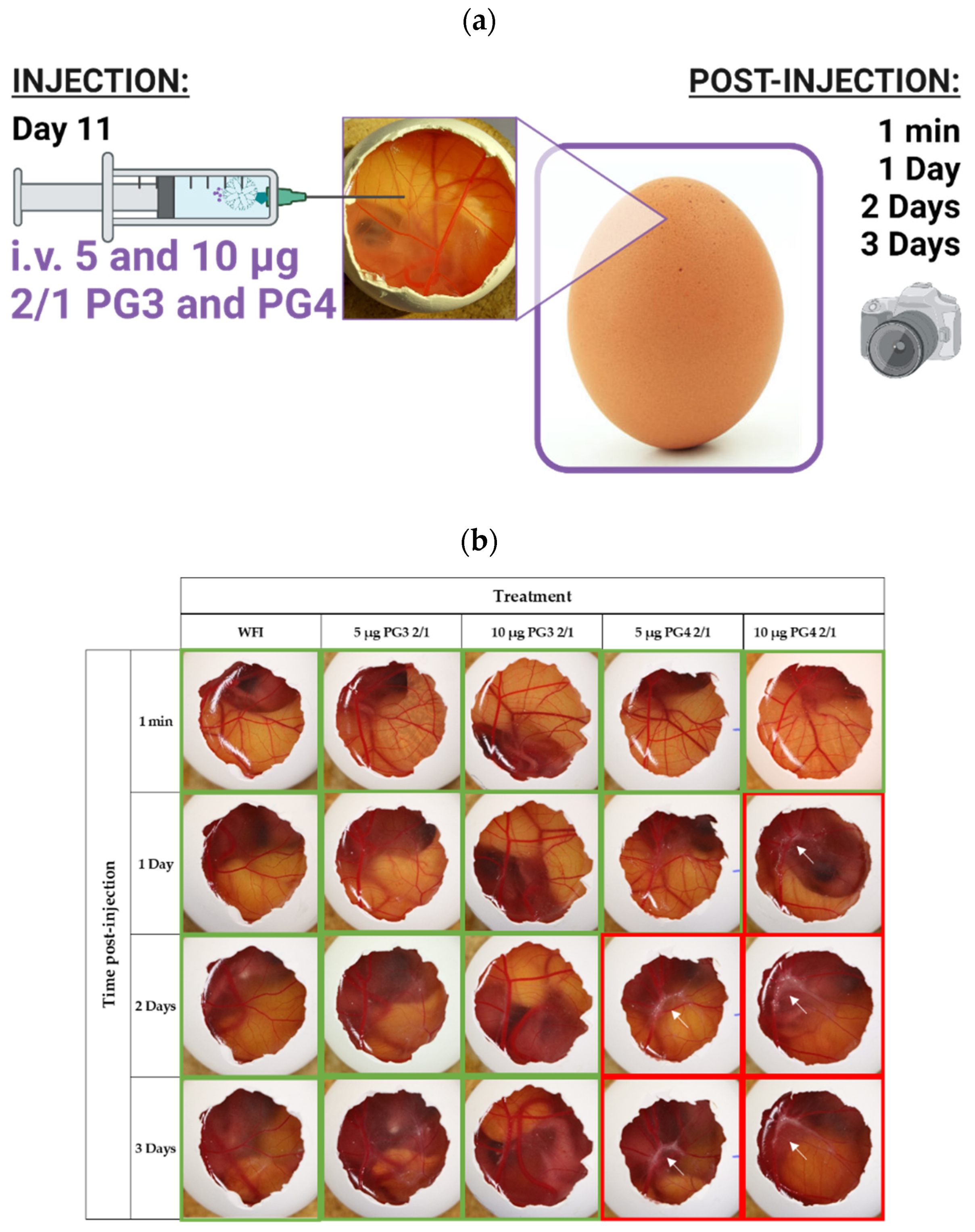
2.5. Hen Egg Chorioallantoic Membrane Model
2.5.1. Ethics Statement
2.5.2. Hen Egg Chorioallantoic Membrane Model
2.5.3. Intravenous Administration
2.5.4. Image Processing
2.6. Statistical Analysis
3. Results and Discussion
3.1. PAMAM-Glu Synthesis
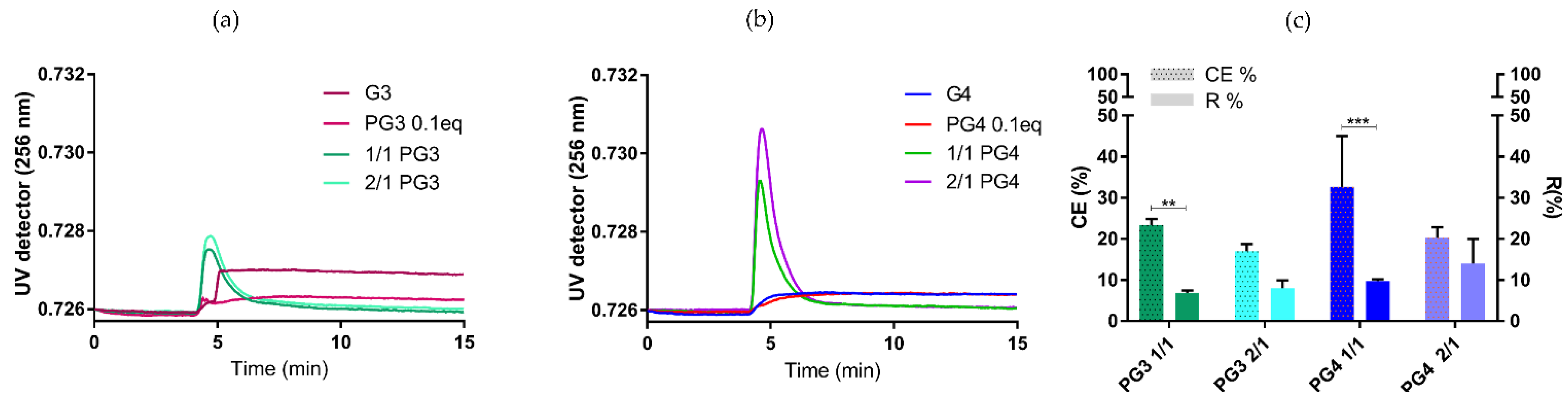
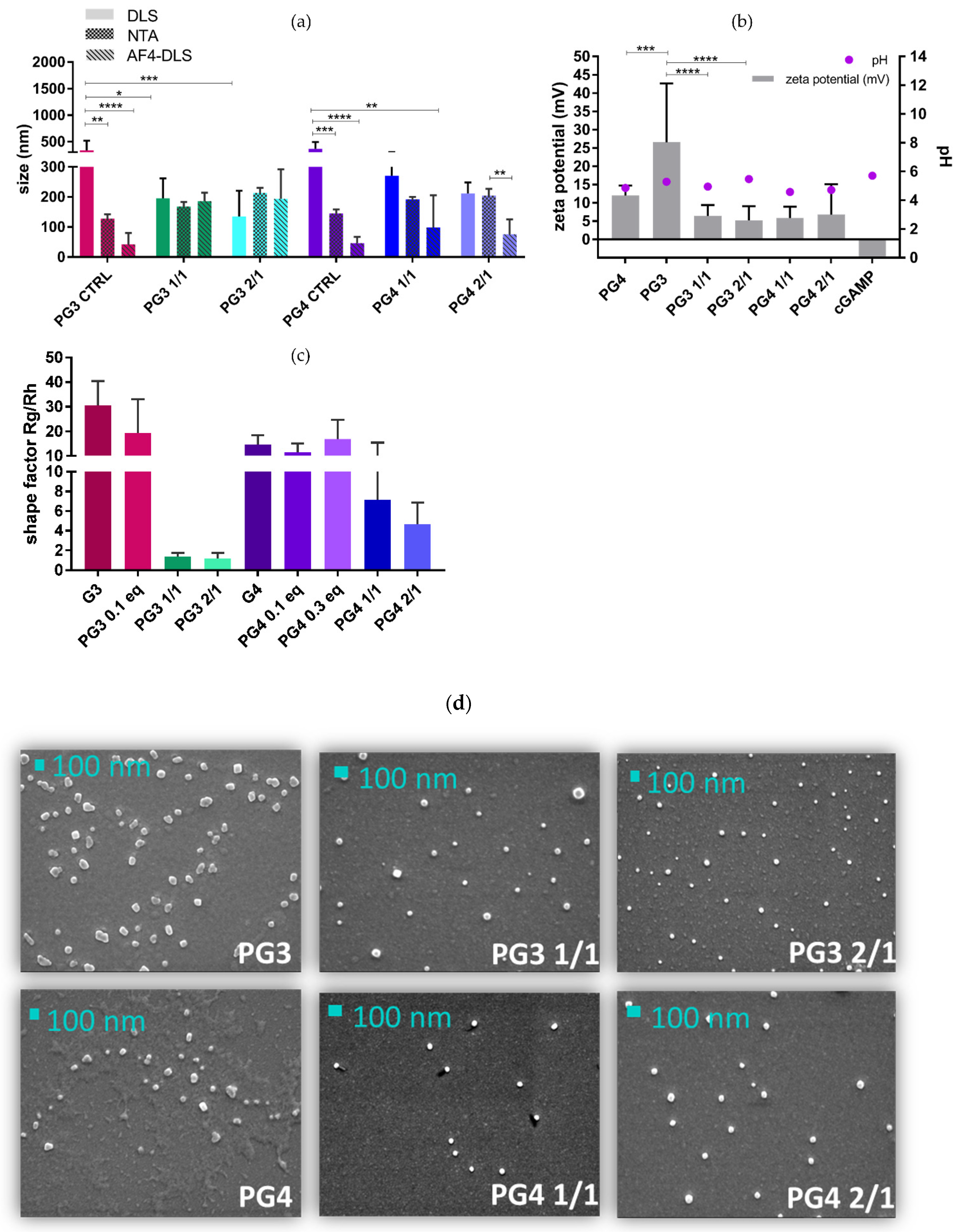
3.2. cGAMP Complexation with PG3/PG4
- -
- The absence of significant differences between the NTA, DLS and AF4-DLS for NCs (PG3 150–200 nm and PG4 200–270 nm) (Figure 5a);
- -
- PG NCs being significantly smaller by DLS compared to PG alone, especially in the case of the 2/1 ratio (PG3 2/1 < PG3 p < 0.0001; PG4 2/1 < PG4 p < 0.01).
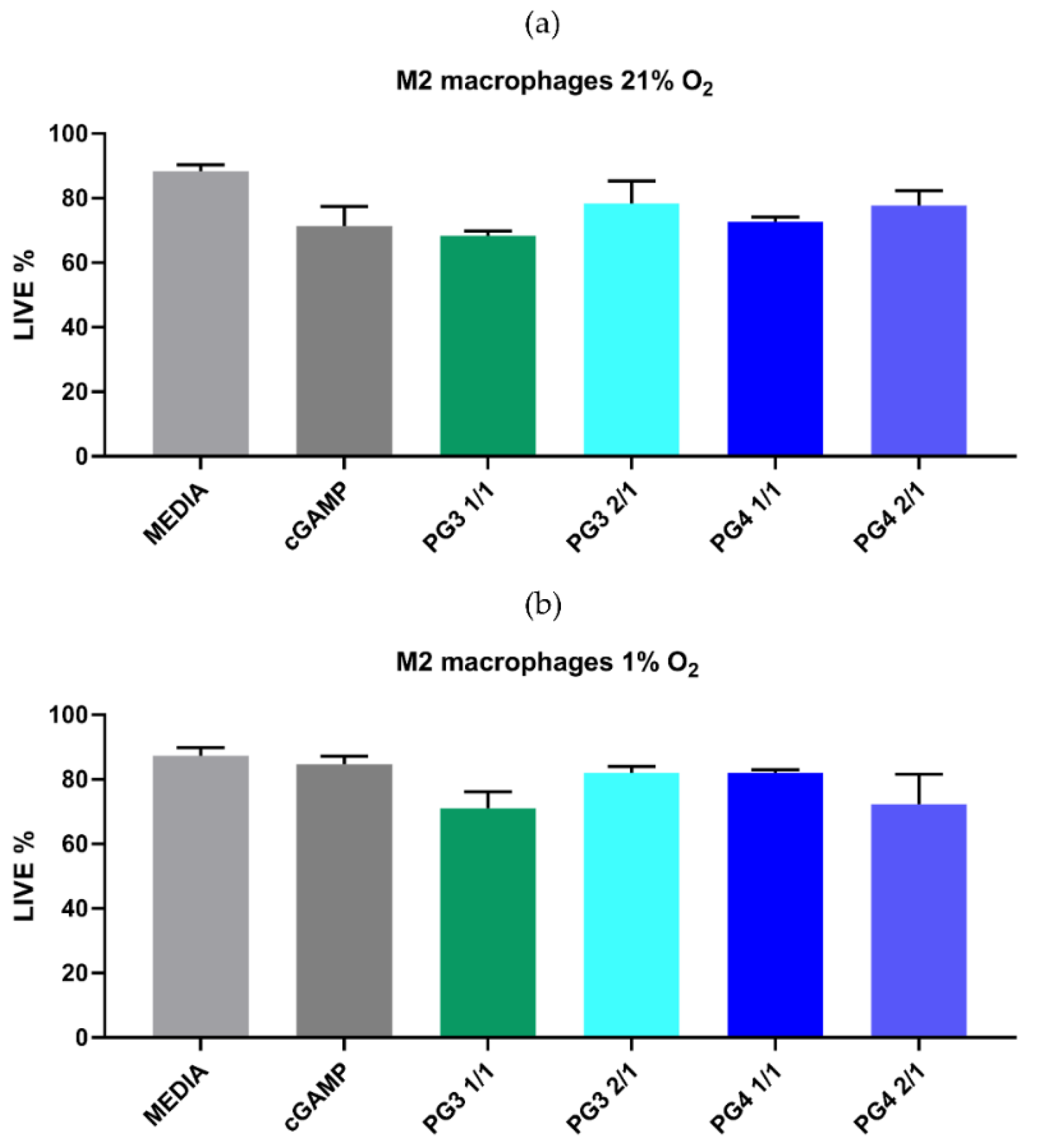
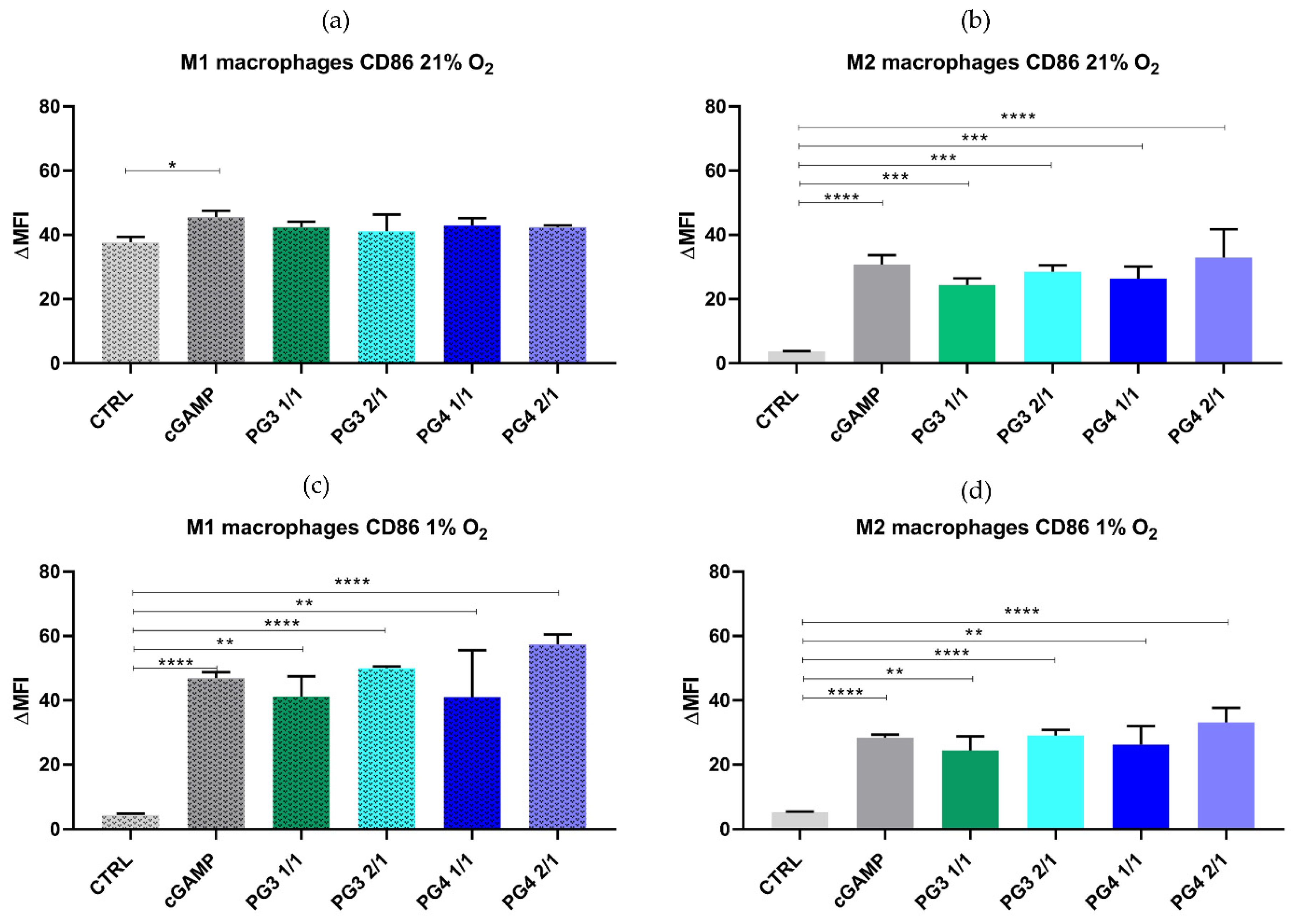
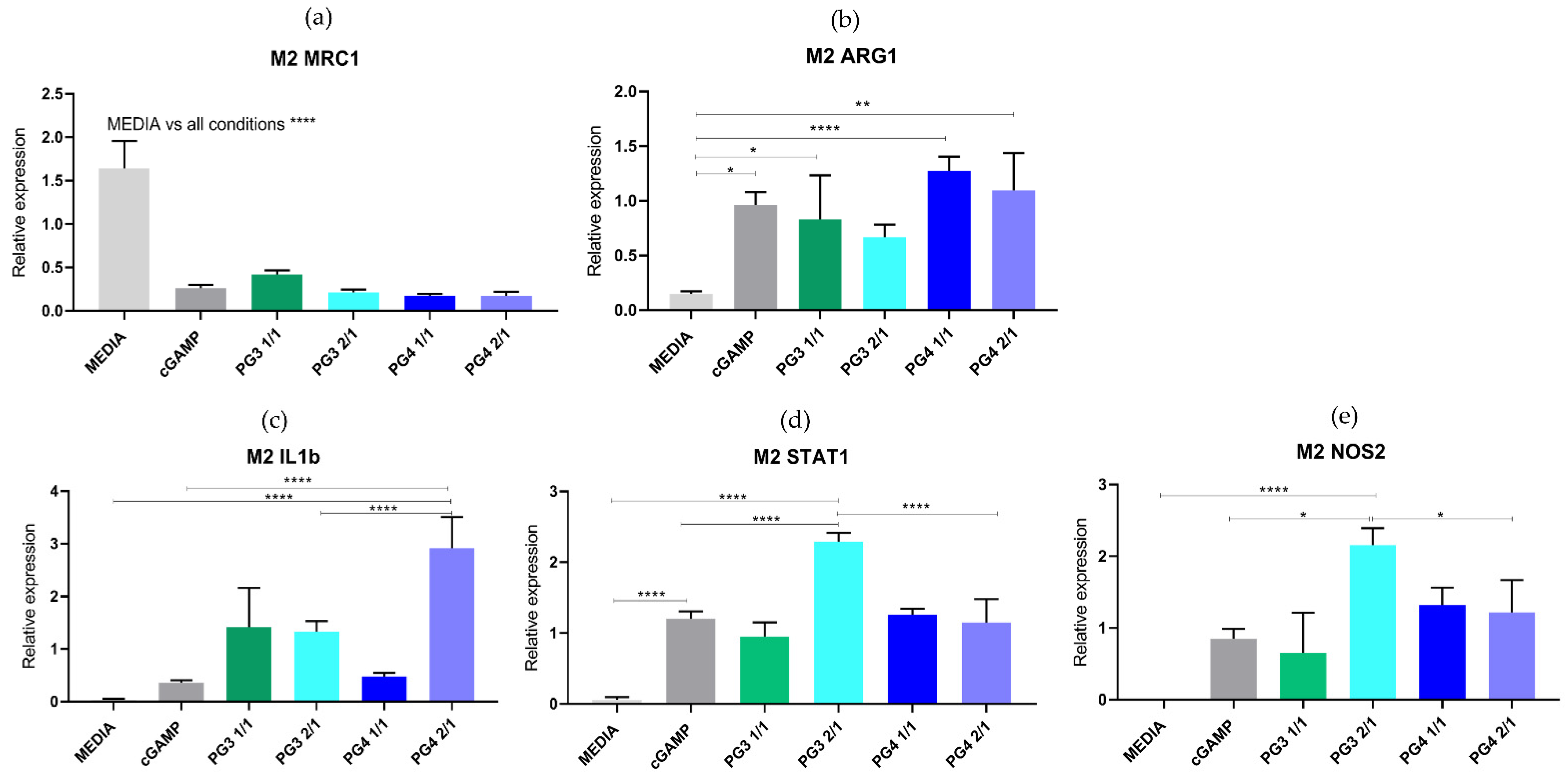
3.3. In Vitro Toxicity and Efficacy of NCs
3.4. In Vivo Toxicity of NCs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| APC | Antigen-presenting cells |
| DC | Dendritic cells |
| TME | Tumor microenvironnement |
| TAM | Tumor-associated macrophages |
| STING | Stimulator of interferon genes |
| IFN | Interferon |
| i.t. | Intratumoral |
| i.v. | Intravenous |
| DS | Degree of substitution |
| NCs | Nanocomplexes |
| N/P | Amino to phosphate group molar charge ratio |
| CE | Complexation efficiency |
| cGAMP | Cyclic GMP-AMP |
| PAMAM | Polyamidoamine |
| EDA | Ethylenediamine |
| G | Generation |
| TLR | Toll-like receptors |
| PAMP | Pathogen-associated molecular patterns |
| DAMP | Damage-associated molecular patterns |
| MRs | Mannose receptor |
| Glu | D-Glucuronic acid |
| PG | PAMAM-Glu |
| PG3 | PAMAM-Glu GENERATION 3 |
| PG4 | PAMAM-Glu GENERATION 4 |
| Da | Daltons |
| DMEM | Dulbecco’s modified Eagle medium |
| EDC | N-ethyl-N′-dimethylaminopropylcarbodiimide |
| CAM | Chorioallantoic membrane |
| EDTA | Ethylenediaminetetraacetic acid |
| FBS | Fetal bovine serum |
| ISO | International Organization for Standardization |
| LVE | Linear viscoelastic region |
| MW | Molecular weight |
| Pa | Pascals |
| Pas | Pascal seconds |
| PBS | Phosphate-buffered saline |
References
- Petrovic, M.; Borchard, G.; Jordan, O. Considerations for the delivery of STING ligands in cancer immunotherapy. J. Control. Release 2021, 339, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Yunna, C.; Mengru, H.; Lei, W.; Weidong, C. Macrophage M1/M2 polarization. Eur. J. Pharmacol. 2020, 877, 173090. [Google Scholar] [CrossRef]
- Ohkuri, T.; Kosaka, A.; Ishibashi, K.; Kumai, T.; Hirata, Y.; Ohara, K.; Nagato, T.; Oikawa, K.; Aoki, N.; Harabuchi, Y.; et al. Intratumoral administration of cGAMP transiently accumulates potent macrophages for anti-tumor immunity at a mouse tumor site. Cancer Immunol. Immunother. 2017, 66, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Bergholz, J.S.; Ding, L.; Lin, Z.; Kabraji, S.K.; Hughes, M.E.; He, X.; Xie, S.; Jiang, T.; Wang, W.; et al. STING agonism reprograms tumor-associated macrophages and overcomes resistance to PARP inhibition in BRCA1-deficient models of breast cancer. Nat. Commun. 2022, 13, 3022. [Google Scholar] [CrossRef] [PubMed]
- Guillot, B.; Portalès, P.; Thanh, A.D.; Merlet, S.; Dereure, O.; Clot, J.; Corbeau, P. The expression of cytotoxic mediators is altered in mononuclear cells of patients with melanoma and increased by interferon-alpha treatment. Br. J. Dermatol. 2005, 152, 690–696. [Google Scholar] [CrossRef] [PubMed]
- Diner, E.J.; Burdette, D.L.; Wilson, S.C.; Monroe, K.M.; Kellenberger, C.A.; Hyodo, M.; Hayakawa, Y.; Hammond, M.C.; Vance, R.E. The Innate Immune DNA Sensor cGAS Produces a Noncanonical Cyclic Dinucleotide that Activates Human STING. Cell Rep. 2013, 3, 1355–1361. [Google Scholar] [CrossRef]
- Tomalia, D.A.; Baker, H.; Dewald, J.; Hall, M.; Kallos, G.; Martin, S.; Roeck, J.; Ryder, J.; Smith, P. A New Class of Polymers: Starburst-Dendritic Macromolecules. Polym. J. 1985, 17, 117–132. [Google Scholar] [CrossRef]
- Li, J.; Liang, H.; Liu, J.; Wang, Z. Poly (amidoamine) (PAMAM) dendrimer mediated delivery of drug and pDNA/siRNA for cancer therapy. Int. J. Pharm. 2018, 546, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Braun, C.S.; Vetro, J.A.; Tomalia, D.A.; Koe, G.S.; Koe, J.G.; Middaugh, C.R. Structure/function relationships of polyamidoamine/DNA dendrimers as gene delivery vehicles. J. Pharm. Sci. 2005, 94, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Araújo, R.V.; Santos, S.D.S.; Igne Ferreira, E.; Giarolla, J. New Advances in General Biomedical Applications of PAMAM Dendrimers. Molecules 2018, 23, 2849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavan, G.M.; Albertazzi, L.; Danani, A. Ability to adapt: Different generations of PAMAM dendrimers show different behaviors in binding siRNA. J. Phys. Chem. B 2010, 114, 2667–2675. [Google Scholar] [CrossRef] [PubMed]
- Bono, N.; Pennetta, C.; Bellucci, M.C.; Sganappa, A.; Malloggi, C.; Tedeschi, G.; Candiani, G.; Volonterio, A. Role of Generation on Successful DNA Delivery of PAMAM–(Guanidino)Neomycin Conjugates. ACS Omega 2019, 4, 6796–6807. [Google Scholar] [CrossRef]
- Urbiola, K.; Blanco-Fernández, L.; Ogris, M.; Rödl, W.; Wagner, E.; Tros de Ilarduya, C. Novel PAMAM-PEG-Peptide Conjugates for siRNA Delivery Targeted to the Transferrin and Epidermal Growth Factor Receptors. J. Pers. Med. 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Ying, W.; Cheruku, P.S.; Bazer, F.W.; Safe, S.H.; Zhou, B. Investigation of macrophage polarization using bone marrow derived macrophages. J. Vis. Exp. 2013, 76, e50323. [Google Scholar] [CrossRef]
- Rőszer, T. Understanding the Mysterious M2 Macrophage through Activation Markers and Effector Mechanisms. Mediat. Inflamm. 2015, 2015, 816460. [Google Scholar] [CrossRef]
- Valdez, H.A.; Marin Franco, J.L.; Gorgojo, J.P.; Alvarez Hayes, J.; Balboa, L.; Fernandez Lahore, M.; Sasiain, M.C.; Rodriguez, M.E. Human macrophage polarization shapes B. pertussis intracellular persistence. J. Leukoc. Biol. 2021, 112, 173–184. [Google Scholar] [CrossRef]
- Hossain, M.K.; Wall, K.A. Use of Dendritic Cell Receptors as Targets for Enhancing Anti-Cancer Immune Responses. Cancers 2019, 11, 418. [Google Scholar] [CrossRef]
- Stahl, P.D.; Ezekowitz, R.A.B. The mannose receptor is a pattern recognition receptor involved in host defense. Curr. Opin. Immunol. 1998, 10, 50–55. [Google Scholar] [CrossRef]
- Szolnoky, G.; Bata-Csörgö, Z.; Kenderessy, A.S.; Kiss, M.; Pivarcsi, A.; Novák, Z.; Nagy Newman, K.; Michel, G.; Ruzicka, T.; Maródi, L.; et al. A Mannose-Binding Receptor is Expressed on Human Keratinocytes and Mediates Killing of Candida albicans. J. Investig. Dermatol. 2001, 117, 205–213. [Google Scholar] [CrossRef]
- Liu, D.R.; Guan, Q.L.; Gao, M.T.; Jiang, L.; Kang, H.X. Mannose receptor as a potential biomarker for gastric cancer: A pilot study. Int. J. Biol. Markers 2017, 32, e278–e283. [Google Scholar] [CrossRef] [Green Version]
- Räihä, M.R.; Puolakkainen, P.A. Tumor-associated macrophages (TAMs) as biomarkers for gastric cancer: A review. Chronic Dis. Transl. Med. 2018, 4, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Feinberg, H.; Jégouzo, S.A.F.; Lasanajak, Y.; Smith, D.F.; Drickamer, K.; Weis, W.I.; Taylor, M.E. Structural analysis of carbohydrate binding by the macrophage mannose receptor CD206. J. Biol. Chem. 2021, 296, 100368. [Google Scholar] [CrossRef] [PubMed]
- Van der Zande, H.J.P.; Nitsche, D.; Schlautmann, L.; Guigas, B.; Burgdorf, S. The Mannose Receptor: From Endocytic Receptor and Biomarker to Regulator of (Meta)Inflammation. Front. Immunol. 2021, 12, 765034. [Google Scholar] [CrossRef] [PubMed]
- Shadrack, D.M.; Mubofu, E.B.; Nyandoro, S.S. Synthesis of Polyamidoamine Dendrimer for Encapsulating Tetramethylscutellarein for Potential Bioactivity Enhancement. Int. J. Mol. Sci. 2015, 16, 26363–26377. [Google Scholar] [CrossRef]
- Markowicz, M.; Szymański, P.; Ciszewski, M.; Kłys, A.; Mikiciuk-Olasik, E. Evaluation of poly(amidoamine) dendrimers as potential carriers of iminodiacetic derivatives using solubility studies and 2D-NOESY NMR spectroscopy. J. Biol. Phys. 2012, 38, 637–656. [Google Scholar] [CrossRef]
- Gomez, M.V.; Guerra, J.; Velders, A.H.; Crooks, R.M. NMR characterization of fourth-generation PAMAM dendrimers in the presence and absence of palladium dendrimer-encapsulated nanoparticles. J. Am. Chem. Soc. 2009, 131, 341–350. [Google Scholar] [CrossRef]
- H NMR Spectrum (1D, 500 MHz, H2O, Experimental) of D-Glucuronic Acid. Available online: https://hmdb.ca/spectra/nmr_one_d/1098 (accessed on 15 July 2022).
- Xu, Z.; Xu, T.; Cheng, Y.; Ma, M.; Xu, P.; Qu, H.; Wen, L. Colorimetric Determination of Polyamidoamine Dendrimers and their Derivates using a Simple and Rapid Ninhydrin Assay. Anal. Lett. 2008, 41, 444–455. [Google Scholar] [CrossRef]
- Weng, J.; Tong, H.H.Y.; Chow, S.F. In Vitro Release Study of the Polymeric Drug Nanoparticles: Development and Validation of a Novel Method. Pharmaceutics 2020, 12, 732. [Google Scholar] [CrossRef]
- Wagner, J.; Gößl, D.; Ustyanovska, N.; Xiong, M.; Hauser, D.; Zhuzhgova, O.; Hočevar, S.; Taskoparan, B.; Poller, L.; Datz, S.; et al. Mesoporous Silica Nanoparticles as pH-Responsive Carrier for the Immune-Activating Drug Resiquimod Enhance the Local Immune Response in Mice. ACS Nano 2021, 15, 4450–4466. [Google Scholar] [CrossRef]
- Helft, J.; Böttcher, J.; Chakravarty, P.; Zelenay, S.; Huotari, J.; Schraml, B.U.; Goubau, D.; Reis e Sousa, C. GM-CSF Mouse Bone Marrow Cultures Comprise a Heterogeneous Population of CD11c(+)MHCII(+) Macrophages and Dendritic Cells. Immunity 2015, 42, 1197–1211. [Google Scholar] [CrossRef] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, Research0034. [Google Scholar] [CrossRef] [PubMed]
- Maiti, P.K.; Çaǧın, T.; Lin, S.-T.; Goddard, W.A. Effect of Solvent and pH on the Structure of PAMAM Dendrimers. Macromolecules 2005, 38, 979–991. [Google Scholar] [CrossRef]
- Topp, A.; Bauer, B.J.; Tomalia, D.A.; Amis, E.J. Effect of Solvent Quality on the Molecular Dimensions of PAMAM Dendrimers. Macromolecules 1999, 32, 7232–7237. [Google Scholar] [CrossRef]
- Fox, L.J.; Richardson, R.M.; Briscoe, W.H. PAMAM dendrimer—Cell membrane interactions. Adv. Colloid Interface Sci. 2018, 257, 1–18. [Google Scholar] [CrossRef]
- Liu, Y.; Bryantsev, V.S.; Diallo, M.S.; Goddard Iii, W.A. PAMAM Dendrimers Undergo pH Responsive Conformational Changes without Swelling. J. Am. Chem. Soc. 2009, 131, 2798–2799. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Haddour, N.; Frenea-Robin, M.; Chevolot, Y.; Monnier, V. Polyamidoamine Dendrimers as Crosslinkers for Efficient Electron Transfer between Redox Probes onto Magnetic Nanoparticles. ChemistrySelect 2018, 3, 2823–2829. [Google Scholar] [CrossRef]
- Wang, T.; Yang, W.-L.; Hong, Y.; Hou, Y.-L. Magnetic nanoparticles grafted with amino-riched dendrimer as magnetic flocculant for efficient harvesting of oleaginous microalgae. Chem. Eng. J. 2016, 297, 304–314. [Google Scholar] [CrossRef]
- Petrovic, M.; Borchard, G.; Jordan, O. Polyethylenimine/cGAMP Nanocomplexes for STING-Mediated Cancer Immunotherapy: Formulation and Characterization Using Orthogonal Techniques. Processes 2022, 10, 882. [Google Scholar] [CrossRef]
- Mintzer, M.A.; Simanek, E.E. Nonviral Vectors for Gene Delivery. Chem. Rev. 2009, 109, 259–302. [Google Scholar] [CrossRef]
- Van Gaal, E.V.; van Eijk, R.; Oosting, R.S.; Kok, R.J.; Hennink, W.E.; Crommelin, D.J.; Mastrobattista, E. How to screen non-viral gene delivery systems in vitro? J. Control. Release 2011, 154, 218–232. [Google Scholar] [CrossRef]
- Guo, P.; Li, Y.; An, J.; Shen, S.; Dou, H. Study on structure-function of starch by asymmetrical flow field-flow fractionation coupled with multiple detectors: A review. Carbohydr. Polym. 2019, 226, 115330. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, K.K.; Simon-Santamaria, J.; McCuskey, R.S.; Smedsrød, B. Liver Sinusoidal Endothelial Cells. Compr. Physiol. 2011, 5, 1751–1774. [Google Scholar] [CrossRef]
- Magaña-Maldonado, R.; Chávez-Cortez, E.G.; Olascoaga-Arellano, N.K.; López-Mejía, M.; Maldonado-Leal, F.M.; Sotelo, J.; Pineda, B. Immunological Evasion in Glioblastoma. Biomed Res. Int. 2016, 2016, 7487313. [Google Scholar] [CrossRef] [PubMed]
- Pyonteck, S.M.; Akkari, L.; Schuhmacher, A.J.; Bowman, R.L.; Sevenich, L.; Quail, D.F.; Olson, O.C.; Quick, M.L.; Huse, J.T.; Teijeiro, V.; et al. CSF-1R inhibition alters macrophage polarization and blocks glioma progression. Nat. Med. 2013, 19, 1264–1272. [Google Scholar] [CrossRef] [PubMed]
- Cunha, L.D.; Yang, M.; Carter, R.; Guy, C.; Harris, L.; Crawford, J.C.; Quarato, G.; Boada-Romero, E.; Kalkavan, H.; Johnson, M.D.L.; et al. LC3-Associated Phagocytosis in Myeloid Cells Promotes Tumor Immune Tolerance. Cell 2018, 175, 429–441.e416. [Google Scholar] [CrossRef] [PubMed]
- Downey, C.M.; Aghaei, M.; Schwendener, R.A.; Jirik, F.R. DMXAA causes tumor site-specific vascular disruption in murine non-small cell lung cancer, and like the endogenous non-canonical cyclic dinucleotide STING agonist, 2’3’-cGAMP, induces M2 macrophage repolarization. PLoS ONE 2014, 9, e99988. [Google Scholar] [CrossRef] [PubMed]
- Amouzegar, A.; Chelvanambi, M.; Filderman, J.N.; Storkus, W.J.; Luke, J.J. STING Agonists as Cancer Therapeutics. Cancers 2021, 13, 2695. [Google Scholar] [CrossRef] [PubMed]
- Shae, D.; Becker, K.W.; Christov, P.; Yun, D.S.; Lytton-Jean, A.K.R.; Sevimli, S.; Ascano, M.; Kelley, M.; Johnson, D.B.; Balko, J.M.; et al. Endosomolytic polymersomes increase the activity of cyclic dinucleotide STING agonists to enhance cancer immunotherapy. Nat. Nanotechnol. 2019, 14, 269–278. [Google Scholar] [CrossRef]
- Murray, P.J.; Allen, J.E.; Biswas, S.K.; Fischer, E.A.; Gilroy, D.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A.; Ivashkiv, L.B.; Lawrence, T.; et al. Macrophage activation and polarization: Nomenclature and experimental guidelines. Immunity 2014, 41, 14–20. [Google Scholar] [CrossRef]
- Ishikawa, H.; Barber, G.N. STING is an endoplasmic reticulum adaptor that facilitates innate immune signalling. Nature 2008, 455, 674–678. [Google Scholar] [CrossRef] [Green Version]
- Kind, C. The development of the circulating blood volume of the chick embryo. Anat. Embryol. 1975, 147, 127–132. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FOCUS STEP | TIP flow | 0.20 mL/min | 3 min |
| Cross flow | 1 mL/min | 3 min | |
| Focus flow | 1.30 mL/min | 0.2 min | |
| ELUTION STEP | Constant cross flow | 1 mL/min | 0.2 min |
| Exponential cross flow power 0.2 | 1–0.1 mL/min | 10 min | |
| Linear cross flow | 0.1–0 mL/min | 10 min | |
| RINSE STEP | TIP flow | 0.1 mL/min | 0.5 min |
| Focus flow | 0.1 mL/min | 0.5 min |
| Controls | PG/cGAMP = Nanocomplexes (NCs) | |||
|---|---|---|---|---|
| 1/1 or 2/1 PG3/cGAMP | 1/1 or 2/1 PG4/cGAMP | |||
| cGAMP | PG3 | PG4 | PG3 1/1 or 2/1 | PG4 1/1 or 2/1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrovic, M.; Porcello, A.; Tankov, S.; Majchrzak, O.; Kiening, M.; Laingoniaina, A.C.; Jbilou, T.; Walker, P.R.; Borchard, G.; Jordan, O. Synthesis, Formulation and Characterization of Immunotherapeutic Glycosylated Dendrimer/cGAMP Complexes for CD206 Targeted Delivery to M2 Macrophages in Cold Tumors. Pharmaceutics 2022, 14, 1883. https://doi.org/10.3390/pharmaceutics14091883
Petrovic M, Porcello A, Tankov S, Majchrzak O, Kiening M, Laingoniaina AC, Jbilou T, Walker PR, Borchard G, Jordan O. Synthesis, Formulation and Characterization of Immunotherapeutic Glycosylated Dendrimer/cGAMP Complexes for CD206 Targeted Delivery to M2 Macrophages in Cold Tumors. Pharmaceutics. 2022; 14(9):1883. https://doi.org/10.3390/pharmaceutics14091883
Chicago/Turabian StylePetrovic, Marija, Alexandre Porcello, Stoyan Tankov, Oliwia Majchrzak, Martin Kiening, Annick Clara Laingoniaina, Tayeb Jbilou, Paul R. Walker, Gerrit Borchard, and Olivier Jordan. 2022. "Synthesis, Formulation and Characterization of Immunotherapeutic Glycosylated Dendrimer/cGAMP Complexes for CD206 Targeted Delivery to M2 Macrophages in Cold Tumors" Pharmaceutics 14, no. 9: 1883. https://doi.org/10.3390/pharmaceutics14091883