
Flavonoids as Potential Antiviral Agents for Porcine Viruses

Abstract
:1. Introduction
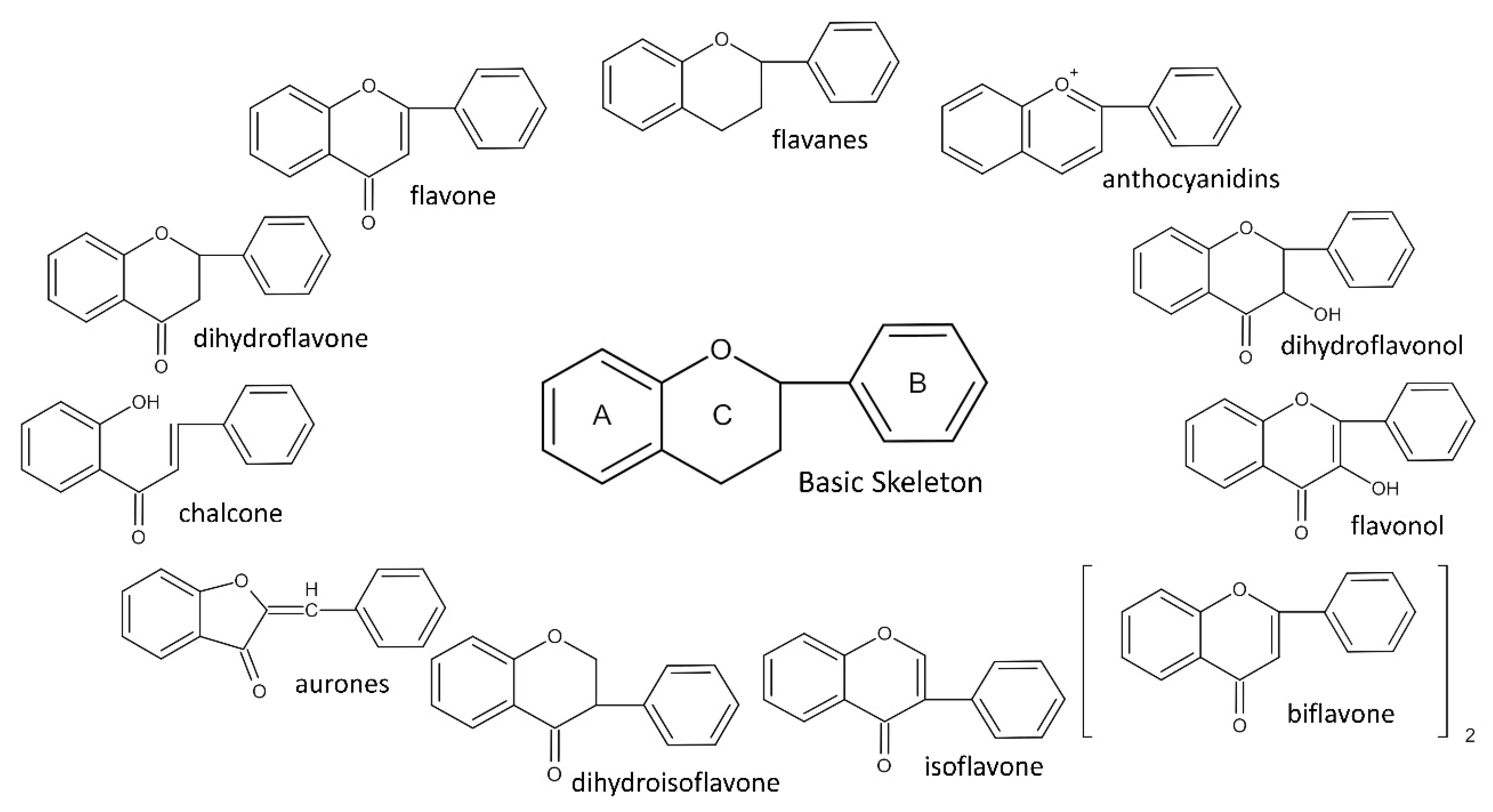
2. Flavonoids and Their Antiviral Mechanism
2.1. Coronaviruses
2.1.1. Transmissible Gastroenteritis Virus (TGEV)
2.1.2. Porcine Epidemic Diarrhea Virus
2.2. Influenza A Virus
2.3. African Swine Fever Virus
2.4. Porcine Reproductive and Respiratory Syndrome Virus
2.5. Porcine Pseudorabies Virus
2.6. Porcine Circovirus 2
3. Limitations of Flavonoids as Antivirals in Pigs
4. Conclusion and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shi, K.; Chen, Y.; Yin, Y.; Long, F.; Feng, S.; Liu, H.; Qu, S.; Si, H. A Multiplex Crystal Digital PCR for Detection of African Swine Fever Virus, Classical Swine Fever Virus, and Porcine Reproductive and Respiratory Syndrome Virus. Front. Vet. Sci. 2022, 9, 926881. [Google Scholar] [CrossRef] [PubMed]
- Rajkhowa, S.; Choudhury, M.; Pegu, S.R.; Sarma, D.K.; Gupta, V.K. Development of a novel one-step triplex PCR assay for the simultaneous detection of porcine circovirus type 2, porcine parvovirus and classical swine fever virus in a single tube. Lett. Appl. Microbiol. 2022, 75, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Liu, Z.; Wang, W.; Tang, D.; Liang, H.; Liu, Z. Establishment and application of a multiplex PCR for rapid and simultaneous detection of six viruses in swine. J. Virol. Methods 2014, 208, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Wang, Y.; Kang, R.; Wu, X.; Lin, H.; Ye, Y.; Yu, J.; Ye, J.; Xie, J.; Cao, Y.; et al. Development and application of a novel Bio-Plex suspension array system for high-throughput multiplexed nucleic acid detection of seven respiratory and reproductive pathogens in swine. J. Virol. Methods 2018, 261, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, T.; Zhang, X.; Liu, X.; Ren, L. Co-Infection of Swine with Porcine Circovirus Type 2 and Other Swine Viruses. Viruses 2019, 11, 185. [Google Scholar] [CrossRef] [PubMed]
- Niu, G.; Chen, S.; Li, X.; Zhang, L.; Ren, L. Advances in Crosstalk between Porcine Circoviruses and Host. Viruses 2022, 14, 1419. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, J.; Wu, X.; Ma, D.; Zhang, X.; Li, R.; Han, C.; Liu, H.; Yin, X.; Du, Q.; et al. PCV2 targets cGAS to inhibit type I interferon induction to promote other DNA virus infection. PLoS Pathog. 2021, 17, e1009940. [Google Scholar] [CrossRef]
- Liu, K.; Ma, G.; Liu, X.; Lu, Y.; Xi, S.; Ou, A.; Wei, J.; Li, B.; Shao, D.; Li, Y.; et al. Porcine reproductive and respiratory syndrome virus counteracts type I interferon-induced early antiviral state by interfering IRF7 activity. Vet. Microbiol. 2019, 229, 28–38. [Google Scholar] [CrossRef]
- Li, C.; Wang, L.; Ren, L. Antiviral mechanisms of candidate chemical medicines and traditional Chinese medicines for SARS-CoV-2 infection. Virus Res. 2020, 286, 198073. [Google Scholar] [CrossRef]
- Wang, Q.; Zhou, L.; Wang, J.; Su, D.; Li, D.; Du, Y.; Yang, G.; Zhang, G.; Chu, B. African Swine Fever Virus K205R Induces ER Stress and Consequently Activates Autophagy and the NF-kappaB Signaling Pathway. Viruses 2022, 14, 394. [Google Scholar] [CrossRef]
- Martin, K.W.; Ernst, E. Antiviral agents from plants and herbs: A systematic review. Antivir. Ther. 2003, 8, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Ninfali, P.; Antonelli, A.; Magnani, M.; Scarpa, E.S. Antiviral Properties of Flavonoids and Delivery Strategies. Nutrients 2020, 12, 2534. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, X.; Wang, C.; He, C.; Ma, Q.; Li, J.; Wang, W.; Xu, Y.T.; Wang, T. Qingwenzhike Prescription Alleviates Acute Lung Injury Induced by LPS via Inhibiting TLR4/NF-kB Pathway and NLRP3 Inflammasome Activation. Front. Pharmacol. 2021, 12, 790072. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Li, X.; Wang, X.; Zhang, L.; Guan, J. Network pharmacological prediction and molecular docking analysis of the combination of Atractylodes macrocephala Koidz. and Paeonia lactiflora Pall. in the treatment of functional constipation and its verification. Anim. Models Exp. Med. 2022, 5, 120–132. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ma, Z.; Li, H.; Li, X. Chinese medicine in the treatment of autoimmune hepatitis: Progress and future opportunities. Anim. Models Exp. Med. 2022, 5, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.C.; Shi, Z.F.; Yan, C.Q.; Tang, Q.X.; Zhao, M.X.; Liu, L.T.; Huang, Q.S.; Ye, L.; Xu, K.; Ke, X. Isolation, X-ray crystal structure of the new diterpene and identification of others lignans and flavonoids from the fresh needles of Pinus massoniana. J. Asian Nat. Prod. Res. 2020, 22, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Feng, Y.; Yu, S.; Fan, Z.; Li, X.; Li, J.; Yin, H. The Flavonoid Biosynthesis Network in Plants. Int. J. Mol. Sci. 2021, 22, 2824. [Google Scholar] [CrossRef]
- Havsteen, B.H. The biochemistry and medical significance of the flavonoids. Pharmacol. Ther. 2002, 96, 67–202. [Google Scholar] [CrossRef]
- Friedman, M. Overview of antibacterial, antitoxin, antiviral, and antifungal activities of tea flavonoids and teas. Mol. Nutr. Food Res. 2007, 51, 116–134. [Google Scholar] [CrossRef]
- Ciumarnean, L.; Milaciu, M.V.; Runcan, O.; Vesa, S.C.; Rachisan, A.L.; Negrean, V.; Perne, M.G.; Donca, V.I.; Alexescu, T.G.; Para, I.; et al. The Effects of Flavonoids in Cardiovascular Diseases. Molecules 2020, 25, 4320. [Google Scholar] [CrossRef]
- Suebsaard, P.; Charerntantanakul, W. Rutin, alpha-tocopherol, and l-ascorbic acid up-regulate type I interferon-regulated gene and type I and II interferon expressions and reduce inflammatory cytokine expressions in monocyte-derived macrophages infected with highly pathogenic porcine reproductive and respiratory syndrome virus. Vet. Immunol. Immunopathol. 2021, 235, 110231. [Google Scholar] [CrossRef] [PubMed]
- Ge, M.; Xiao, Y.; Chen, H.; Luo, F.; Du, G.; Zeng, F. Multiple antiviral approaches of (-)-epigallocatechin-3-gallate (EGCG) against porcine reproductive and respiratory syndrome virus infection in vitro. Antivir. Res. 2018, 158, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.W.; Fu, P.F.; Zeng, L.; Qi, Y.L.; Li, X.Q.; Wang, Q.; Yang, G.Y.; Li, H.W.; Wang, J.; Chu, B.B.; et al. EGCG Restricts PRRSV Proliferation by Disturbing Lipid Metabolism. Microbiol. Spectr. 2022, 10, e0227621. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Liu, S.; Li, C.; Yang, L.; Zu, Y. In vitro evaluation of the antiviral activity of the synthetic epigallocatechin gallate analog-epigallocatechin gallate (EGCG) palmitate against porcine reproductive and respiratory syndrome virus. Viruses 2014, 6, 938–950. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, R.; Hu, H.; Chen, X.; Yin, Z.; Liang, X.; He, C.; Yin, L.; Ye, G.; Zou, Y.; et al. The antiviral activity of kaempferol against pseudorabies virus in mice. BMC Vet. Res. 2021, 17, 247. [Google Scholar] [CrossRef] [PubMed]
- Huan, C.; Xu, W.; Guo, T.; Pan, H.; Zou, H.; Jiang, L.; Li, C.; Gao, S. (-)-Epigallocatechin-3-Gallate Inhibits the Life Cycle of Pseudorabies Virus In Vitro and Protects Mice Against Fatal Infection. Front. Cell. Infect. Microbiol. 2020, 10, 616895. [Google Scholar] [CrossRef]
- Li, J.; Song, D.; Wang, S.; Dai, Y.; Zhou, J.; Gu, J. Antiviral Effect of Epigallocatechin Gallate via Impairing Porcine Circovirus Type 2 Attachment to Host Cell Receptor. Viruses 2020, 12, 176. [Google Scholar] [CrossRef]
- Xu, M.J.; Liu, B.J.; Wang, C.L.; Wang, G.H.; Tian, Y.; Wang, S.H.; Li, J.; Li, P.Y.; Zhang, R.H.; Wei, D.; et al. Epigallocatechin-3-gallate inhibits TLR4 signaling through the 67-kDa laminin receptor and effectively alleviates acute lung injury induced by H9N2 swine influenza virus. Int. Immunopharmacol. 2017, 52, 24–33. [Google Scholar] [CrossRef]
- Ruansit, W.; Charerntantanakul, W. Oral supplementation of quercetin in PRRSV-1 modified-live virus vaccinated pigs in response to HP-PRRSV-2 challenge. Vaccine 2020, 38, 3570–3581. [Google Scholar] [CrossRef]
- Gao, J.; Xiao, S.; Liu, X.; Wang, L.; Ji, Q.; Mo, D.; Chen, Y. Inhibition of HSP70 reduces porcine reproductive and respiratory syndrome virus replication in vitro. BMC Microbiol. 2014, 14, 64. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Cao, H.; Cheng, Y.; Zhang, X.; Zeng, W.; Sun, Y.; Chen, S.; He, Q.; Han, H. Inhibition of Porcine Epidemic Diarrhea Virus Replication and Viral 3C-Like Protease by Quercetin. Int. J. Mol. Sci. 2020, 21, 8095. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Kim, S.; Shin, D.H.; Kim, M.S. Inhibition of SARS-CoV 3CL protease by flavonoids. J. Enzym. Inhib. Med. Chem. 2020, 35, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Zhu, G.H.; Zhang, Y.N.; Hu, Q.; Wang, H.N.; Yu, H.N.; Qin, X.Y.; Guan, X.Q.; Xiang, Y.W.; Tang, H.; et al. Flavonoids in Ampelopsis grossedentata as covalent inhibitors of SARS-CoV-2 3CL(pro): Inhibition potentials, covalent binding sites and inhibitory mechanisms. Int. J. Biol. Macromol. 2021, 187, 976–987. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Kim, J.H.; Lee, C.H.; Ahn, Y.J.; Song, J.H.; Baek, S.H.; Kwon, D.H. Antiviral activity of quercetin 7-rhamnoside against porcine epidemic diarrhea virus. Antiviral Res. 2009, 81, 77–81. [Google Scholar] [CrossRef]
- Sun, Y.; Li, C.; Li, Z.; Shangguan, A.; Jiang, J.; Zeng, W.; Zhang, S.; He, Q. Quercetin as an antiviral agent inhibits the Pseudorabies virus in vitro and in vivo. Virus Res. 2021, 305, 198556. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xie, X.; Chen, Q.; Yi, S.; Chen, J.; Xiao, Q.; Yu, M.; Wei, Y.; Hu, T. Effects of Quercitrin on PRV-Induced Secretion of Reactive Oxygen Species and Prediction of lncRNA Regulatory Targets in 3D4/2 Cells. Antioxidants 2022, 11, 631. [Google Scholar] [CrossRef]
- Arabyan, E.; Hakobyan, A.; Hakobyan, T.; Grigoryan, R.; Izmailyan, R.; Avetisyan, A.; Karalyan, Z.; Jackman, J.A.; Ferreira, F.; Elrod, C.C.; et al. Flavonoid Library Screening Reveals Kaempferol as a Potential Antiviral Agent Against African Swine Fever Virus. Front. Microbiol. 2021, 12, 736780. [Google Scholar] [CrossRef]
- Zhang, R.; Ai, X.; Duan, Y.; Xue, M.; He, W.; Wang, C.; Xu, T.; Xu, M.; Liu, B.; Li, C.; et al. Kaempferol ameliorates H9N2 swine influenza virus-induced acute lung injury by inactivation of TLR4/MyD88-mediated NF-kappaB and MAPK signaling pathways. Biomed. Pharmacother. 2017, 89, 660–672. [Google Scholar] [CrossRef]
- Su, M.; Shi, D.; Xing, X.; Qi, S.; Yang, D.; Zhang, J.; Han, Y.; Zhu, Q.; Sun, H.; Wang, X.; et al. Coronavirus Porcine Epidemic Diarrhea Virus Nucleocapsid Protein Interacts with p53 To Induce Cell Cycle Arrest in S-Phase and Promotes Viral Replication. J. Virol. 2021, 95, e0018721. [Google Scholar] [CrossRef]
- Ren, C.Z.; Hu, W.Y.; Li, J.C.; Xie, Y.H.; Jia, N.N.; Shi, J.; Wei, Y.Y.; Hu, T.J. Ethyl acetate fraction of flavonoids from Polygonum hydropiper L. modulates pseudorabies virus-induced inflammation in RAW264.7 cells via the nuclear factor-kappa B and mitogen-activated protein kinase pathways. J. Vet. Med. Sci. 2020, 82, 1781–1792. [Google Scholar] [CrossRef]
- Chen, H.L.; Yang, J.; Fu, Y.F.; Meng, X.N.; Zhao, W.D.; Hu, T.J. Effect of total flavonoids of Spatholobus suberectus Dunn on PCV2 induced oxidative stress in RAW264.7 cells. BMC Complementary Med. Ther. 2017, 17, 244. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.F.; Jiang, L.H.; Zhao, W.D.; Xi-Nan, M.; Huang, S.Q.; Yang, J.; Hu, T.J.; Chen, H.L. Immunomodulatory and antioxidant effects of total flavonoids of Spatholobus suberectus Dunn on PCV2 infected mice. Sci. Rep. 2017, 7, 8676. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; He, L.; Ning, P.; Lin, J.; Li, H.; Lin, Z.; Kang, K.; Zhang, Y. (+)-Catechin inhibition of transmissible gastroenteritis coronavirus in swine testicular cells is involved its antioxidation. Res. Vet. Sci. 2015, 103, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Xie, W.; Liu, Y.; Sui, B.; Zhang, H.; Liu, L.; Tan, Y.; Tong, X.; Fu, Z.F.; Yin, P.; et al. Receptor tyrosine kinase inhibitors block proliferation of TGEV mainly through p38 mitogen-activated protein kinase pathways. Antiviral Res. 2020, 173, 104651. [Google Scholar] [CrossRef]
- Jo, S.; Kim, S.; Shin, D.H.; Kim, M.S. Inhibition of African swine fever virus protease by myricetin and myricitrin. J. Enzym. Inhib. Med. Chem. 2020, 35, 1045–1049. [Google Scholar] [CrossRef]
- Arabyan, E.; Hakobyan, A.; Kotsinyan, A.; Karalyan, Z.; Arakelov, V.; Arakelov, G.; Nazaryan, K.; Simonyan, A.; Aroutiounian, R.; Ferreira, F.; et al. Genistein inhibits African swine fever virus replication in vitro by disrupting viral DNA synthesis. Antiviral Res. 2018, 156, 128–137. [Google Scholar] [CrossRef]
- Hakobyan, A.; Arabyan, E.; Avetisyan, A.; Abroyan, L.; Hakobyan, L.; Zakaryan, H. Apigenin inhibits African swine fever virus infection in vitro. Arch. Virol. 2016, 161, 3445–3453. [Google Scholar] [CrossRef]
- Hakobyan, A.; Arabyan, E.; Kotsinyan, A.; Karalyan, Z.; Sahakyan, H.; Arakelov, V.; Nazaryan, K.; Ferreira, F.; Zakaryan, H. Inhibition of African swine fever virus infection by genkwanin. Antiviral Res. 2019, 167, 78–82. [Google Scholar] [CrossRef]
- Dao, T.T.; Nguyen, P.H.; Lee, H.S.; Kim, E.; Park, J.; Lim, S.I.; Oh, W.K. Chalcones as novel influenza A (H1N1) neuraminidase inhibitors from Glycyrrhiza inflata. Bioorganic Med. Chem. Lett. 2011, 21, 294–298. [Google Scholar] [CrossRef]
- Lunney, J.K.; Fang, Y.; Ladinig, A.; Chen, N.; Li, Y.; Rowland, B.; Renukaradhya, G.J. Porcine Reproductive and Respiratory Syndrome Virus (PRRSV): Pathogenesis and Interaction with the Immune System. Annu. Rev. Anim. Biosci. 2016, 4, 129–154. [Google Scholar] [CrossRef]
- Ryu, Y.B.; Curtis-Long, M.J.; Lee, J.W.; Ryu, H.W.; Kim, J.Y.; Lee, W.S.; Park, K.H. Structural characteristics of flavanones and flavones from Cudrania tricuspidata for neuraminidase inhibition. Bioorg. Med. Chem. Lett. 2009, 19, 4912–4915. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.M.; Liu, T.X.; Wang, T.Y.; Wang, G.; Liu, Y.G.; Liu, S.G.; Tang, Y.D.; Cai, X.H. Isobavachalcone inhibits post-entry stages of the porcine reproductive and respiratory syndrome virus life cycle. Arch. Virol. 2018, 163, 1263–1270. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, T.X.; Wang, T.Y.; Tang, Y.D.; Wei, P. Isobavachalcone inhibits Pseudorabies virus by impairing virus-induced cell-to-cell fusion. Virol. J. 2020, 17, 39. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Song, Z.; Bai, J.; Nauwynck, H.; Zhao, Y.; Jiang, P. Xanthohumol inhibits PRRSV proliferation and alleviates oxidative stress induced by PRRSV via the Nrf2-HMOX1 axis. Vet. Res. 2019, 50, 61. [Google Scholar] [CrossRef]
- Liu, X.; Bai, J.; Jiang, C.; Song, Z.; Zhao, Y.; Nauwynck, H.; Jiang, P. Therapeutic effect of Xanthohumol against highly pathogenic porcine reproductive and respiratory syndrome viruses. Vet. Microbiol. 2019, 238, 108431. [Google Scholar] [CrossRef]
- Smith, B.N.; Oelschlager, M.L.; Abdul Rasheed, M.S.; Dilger, R.N. Dietary soy isoflavones reduce pathogen-related mortality in growing pigs under porcine reproductive and respiratory syndrome viral challenge. J. Anim. Sci. 2020, 98, skaa024. [Google Scholar] [CrossRef]
- Rochell, S.J.; Alexander, L.S.; Rocha, G.C.; Van Alstine, W.G.; Boyd, R.D.; Pettigrew, J.E.; Dilger, R.N. Effects of dietary soybean meal concentration on growth and immune response of pigs infected with porcine reproductive and respiratory syndrome virus. J. Anim. Sci. 2015, 93, 2987–2997. [Google Scholar] [CrossRef]
- Smith, B.N.; Morris, A.; Oelschlager, M.L.; Connor, J.; Dilger, R.N. Effects of dietary soy isoflavones and soy protein source on response of weanling pigs to porcine reproductive and respiratory syndrome viral infection. J. Anim. Sci. 2019, 97, 2989–3006. [Google Scholar] [CrossRef]
- Wu, M.; Yi, D.; Zhang, Q.; Wu, T.; Yu, K.; Peng, M.; Wang, L.; Zhao, D.; Hou, Y.; Wu, G. Puerarin enhances intestinal function in piglets infected with porcine epidemic diarrhea virus. Sc.i Rep. 2021, 11, 6552. [Google Scholar] [CrossRef]
- Wu, M.; Zhang, Q.; Yi, D.; Wu, T.; Chen, H.; Guo, S.; Li, S.; Ji, C.; Wang, L.; Zhao, D.; et al. Quantitative Proteomic Analysis Reveals Antiviral and Anti-inflammatory Effects of Puerarin in Piglets Infected With Porcine Epidemic Diarrhea Virus. Front. Immunol. 2020, 11, 169. [Google Scholar] [CrossRef]
- Yan, Q.; Liu, X.; Sun, Y.; Zeng, W.; Li, Y.; Zhao, F.; Wu, K.; Fan, S.; Zhao, M.; Chen, J.; et al. Swine Enteric Coronavirus: Diverse Pathogen-Host Interactions. Int. J. Mol. Sci. 2022, 23, 3953. [Google Scholar] [CrossRef] [PubMed]
- Thakor, J.C.; Dinesh, M.; Manikandan, R.; Bindu, S.; Sahoo, M.; Sahoo, D.; Dhawan, M.; Pandey, M.K.; Tiwari, R.; Emran, T.B.; et al. Swine coronaviruses (SCoVs) and their emerging threats to swine population, inter-species transmission, exploring the susceptibility of pigs for SARS-CoV-2 and zoonotic concerns. Vet. Q. 2022, 42, 125–147. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhang, X.; Lian, K.; Tian, X.; Zhang, M.; Wang, S.; Chen, C.; Nie, C.; Pan, Y.; Han, F.; et al. Antiviral effects of Bovine antimicrobial peptide against TGEV in vivo and in vitro. J. Vet. Sci. 2020, 21, e80. [Google Scholar] [CrossRef]
- Shimizu, M.; Adachi, S.; Masuda, M.; Kozawa, O.; Moriwaki, H. Cancer chemoprevention with green tea catechins by targeting receptor tyrosine kinases. Mol. Nutr. Food Res. 2011, 55, 832–843. [Google Scholar] [CrossRef] [PubMed]
- Bernatova, I. Biological activities of (-)-epicatechin and (-)-epicatechin-containing foods: Focus on cardiovascular and neuropsychological health. Biotechnol. Adv. 2018, 36, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Saif, L.J.; Wang, Q. Porcine epidemic diarrhea virus (PEDV): An update on etiology, transmission, pathogenesis, and prevention and control. Virus Res. 2020, 286, 198045. [Google Scholar] [CrossRef]
- Park, J.; Lee, C. Emergence and evolution of novel G2b-like porcine epidemic diarrhea virus inter-subgroup G1b recombinants. Arch. Virol. 2020, 165, 2471–2478. [Google Scholar] [CrossRef]
- Dhanya, R. Quercetin for managing type 2 diabetes and its complications, an insight into multitarget therapy. Biomed. Pharmacother. 2022, 146, 112560. [Google Scholar] [CrossRef]
- Wang, Q.; Wei, H.C.; Zhou, S.J.; Li, Y.; Zheng, T.T.; Zhou, C.Z.; Wan, X.H. Hyperoside: A review on its sources, biological activities, and molecular mechanisms. Phytother. Res. 2022, 36, 2779–2802. [Google Scholar] [CrossRef]
- Zhang, L. Pharmacokinetics and drug delivery systems for puerarin, a bioactive flavone from traditional Chinese medicine. Drug Deliv. 2019, 26, 860–869. [Google Scholar] [CrossRef] [Green Version]
- Skelton, R.M.; Huber, V.C. Comparing Influenza Virus Biology for Understanding Influenza D Virus. Viruses 2022, 14, 1036. [Google Scholar] [CrossRef] [PubMed]
- Sui, J.; Hwang, W.C.; Perez, S.; Wei, G.; Aird, D.; Chen, L.M.; Santelli, E.; Stec, B.; Cadwell, G.; Ali, M.; et al. Structural and functional bases for broad-spectrum neutralization of avian and human influenza A viruses. Nat. Struct. Mol. Biol. 2009, 16, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, E.C. Influenza Virus. Trends Microbiol. 2018, 26, 809–810. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, R.P.; Gordon, M.L. A systematic review of influenza A virus prevalence and transmission dynamics in backyard swine populations globally. Porc. Health Manag. 2022, 8, 10. [Google Scholar] [CrossRef]
- Anderson, T.K.; Chang, J.; Arendsee, Z.W.; Venkatesh, D.; Souza, C.K.; Kimble, J.B.; Lewis, N.S.; Davis, C.T.; Vincent, A.L. Swine Influenza A Viruses and the Tangled Relationship with Humans. Cold Spring Harb. Perspect. Med. 2021, 11, a038737. [Google Scholar] [CrossRef]
- Russell, C.J. Hemagglutinin Stability and Its Impact on Influenza A Virus Infectivity, Pathogenicity, and Transmissibility in Avians, Mice, Swine, Seals, Ferrets, and Humans. Viruses 2021, 13, 746. [Google Scholar] [CrossRef]
- Saito, T.; Sakuma, S.; Mine, J.; Uchida, Y.; Hangalapura, B.N. Genetic Diversity of the Hemagglutinin Genes of Influenza a Virus in Asian Swine Populations. Viruses 2022, 14, 747. [Google Scholar] [CrossRef]
- Rudrapal, M.; Khan, J.; Dukhyil, A.A.B.; Alarousy, R.; Attah, E.I.; Sharma, T.; Khairnar, S.J.; Bendale, A.R. Chalcone Scaffolds, Bioprecursors of Flavonoids: Chemistry, Bioactivities, and Pharmacokinetics. Molecules 2021, 26, 7177. [Google Scholar] [CrossRef]
- Elkhalifa, D.; Al-Hashimi, I.; Al Moustafa, A.E.; Khalil, A. A comprehensive review on the antiviral activities of chalcones. J. Drug Target. 2021, 29, 403–419. [Google Scholar] [CrossRef]
- Hameed, A.; Abdullah, M.I.; Ahmed, E.; Sharif, A.; Irfan, A.; Masood, S. Anti-HIV cytotoxicity enzyme inhibition and molecular docking studies of quinoline based chalcones as potential non-nucleoside reverse transcriptase inhibitors (NNRT). Bioorg. Chem. 2016, 65, 175–182. [Google Scholar] [CrossRef]
- Mathaiyan, M.; Suresh, A.; Balamurugan, R. Binding property of HIV p24 and Reverse transcriptase by chalcones from Pongamia pinnata seeds. Bioinformation 2018, 14, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.; Patil, S.; Sanchez, T.W.; Neamati, N.; Schinazi, R.F.; Buolamwini, J.K. Synthesis, biological evaluation and 3D-QSAR studies of 3-keto salicylic acid chalcones and related amides as novel HIV-1 integrase inhibitors. Bioorg. Med. Chem. 2011, 19, 2030–2045. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Sanchez, T.; Al-Mawsawi, L.Q.; Dayam, R.; Yunes, R.A.; Garofalo, A.; Bolger, M.B.; Neamati, N. Discovery of structurally diverse HIV-1 integrase inhibitors based on a chalcone pharmacophore. Bioorg. Med. Chem. 2007, 15, 4985–5002. [Google Scholar] [CrossRef] [PubMed]
- Malbari, K.; Gonsalves, H.; Chintakrindi, A.; Gohil, D.; Joshi, M.; Kothari, S.; Srivastava, S.; Chowdhary, A.; Kanyalkar, M. In search of effective H1N1 neuraminidase inhibitor by molecular docking, antiviral evaluation and membrane interaction studies using NMR. Acta Virol. 2018, 62, 179–190. [Google Scholar] [CrossRef]
- Yang, M.; Li, N.; Li, F.; Zhu, Q.; Liu, X.; Han, Q.; Wang, Y.; Chen, Y.; Zeng, X.; Lv, Y.; et al. Xanthohumol, a main prenylated chalcone from hops, reduces liver damage and modulates oxidative reaction and apoptosis in hepatitis C virus infected Tupaia belangeri. Int. Immunopharmacol. 2013, 16, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Quon, M.J.; Kim, J.A. New insights into the mechanisms of polyphenols beyond antioxidant properties; lessons from the green tea polyphenol, epigallocatechin 3-gallate. Redox Biol. 2014, 2, 187–195. [Google Scholar] [CrossRef]
- Romano, A.; Martel, F. The Role of EGCG in Breast Cancer Prevention and Therapy. Mini-Rev. Med. Chem. 2021, 21, 883–898. [Google Scholar] [CrossRef]
- Rahul; Siddique, Y.H. Neurodegenerative Diseases and Flavonoids: Special Reference to Kaempferol. CNS Neurol. Disord. Drug Targets 2021, 20, 327–342. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Y.; An, Y.; Fang, G. The mechanism of anticancer action and potential clinical use of kaempferol in the treatment of breast cancer. Biomed. Pharmacother. 2019, 117, 109086. [Google Scholar] [CrossRef]
- Imran, M.; Rauf, A.; Shah, Z.A.; Saeed, F.; Imran, A.; Arshad, M.U.; Ahmad, B.; Bawazeer, S.; Atif, M.; Peters, D.G.; et al. Chemo-preventive and therapeutic effect of the dietary flavonoid kaempferol: A comprehensive review. Phytother. Res. 2019, 33, 263–275. [Google Scholar] [CrossRef]
- Huang, Z.; Guo, S.; Fu, C.; Zhou, W.; Stalin, A.; Zhang, J.; Liu, X.; Jia, S.; Wu, C.; Lu, S.; et al. Identification of molecular mechanisms underlying the therapeutic effects of Xintong granule in coronary artery disease by a network pharmacology and molecular docking approach. Medicine (Baltimore) 2022, 101, e29829. [Google Scholar] [CrossRef] [PubMed]
- Devi, K.P.; Malar, D.S.; Nabavi, S.F.; Sureda, A.; Xiao, J.; Nabavi, S.M.; Daglia, M. Kaempferol and inflammation: From chemistry to medicine. Pharmacol. Res. 2015, 99, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Xie, M.; Wu, W.; Chen, Z. Structures and Functional Diversities of ASFV Proteins. Viruses 2021, 13, 2124. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kang, W.; Yang, W.; Zhang, J.; Li, D.; Zheng, H. Structure of African Swine Fever Virus and Associated Molecular Mechanisms Underlying Infection and Immunosuppression: A Review. Front. Immunol. 2021, 12, 715582. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ai, Q.; Huang, S.; Ou, Y.; Gao, Y.; Tong, T.; Fan, H. Immune Escape Mechanism and Vaccine Research Progress of African Swine Fever Virus. Vaccines 2022, 10, 344. [Google Scholar] [CrossRef]
- Jiang, M.; Zhu, M.; Wang, L.; Yu, S. Anti-tumor effects and associated molecular mechanisms of myricetin. Biomed. Pharmacother. 2019, 120, 109506. [Google Scholar] [CrossRef]
- Jang, J.Y.; Sung, B.; Kim, N.D. Role of Induced Programmed Cell Death in the Chemopreventive Potential of Apigenin. Int. J. Mol. Sci. 2022, 23, 3757. [Google Scholar] [CrossRef]
- Zhang, L.; Cheng, X.; Gao, Y.; Zheng, J.; Xu, Q.; Sun, Y.; Guan, H.; Yu, H.; Sun, Z. Apigenin induces autophagic cell death in human papillary thyroid carcinoma BCPAP cells. Food Funct. 2015, 6, 3464–3472. [Google Scholar] [CrossRef]
- Hsu, M.C.; Guo, B.C.; Chen, C.H.; Hu, P.A.; Lee, T.S. Apigenin ameliorates hepatic lipid accumulation by activating the autophagy-mitochondria pathway. J. Food Drug Anal. 2021, 29, 240–254. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, X.; Nie, Y.; Li, H.; Chen, W.; Lin, W.; Chen, F.; Xie, Q. African Swine Fever Virus Protein E199L Promotes Cell Autophagy through the Interaction of PYCR2. Virol. Sin. 2021, 36, 196–206. [Google Scholar] [CrossRef]
- Sun, M.; Yu, S.; Ge, H.; Wang, T.; Li, Y.; Zhou, P.; Pan, L.; Han, Y.; Yang, Y.; Sun, Y.; et al. The A137R Protein of African Swine Fever Virus Inhibits Type I Interferon Production via the Autophagy-Mediated Lysosomal Degradation of TBK1. J. Virol. 2022, 96, e0195721. [Google Scholar] [CrossRef] [PubMed]
- Mas-Bargues, C.; Borras, C.; Vina, J. The multimodal action of genistein in Alzheimer’s and other age-related diseases. Free. Radic. Biol. Med. 2022, 183, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Bocard, L.V.; Kick, A.R.; Hug, C.; Lischer, H.E.L.; Kaser, T.; Summerfield, A. Systems Immunology Analyses Following Porcine Respiratory and Reproductive Syndrome Virus Infection and Vaccination. Front. Immunol. 2021, 12, 779747. [Google Scholar] [CrossRef]
- Muvhulawa, N.; Dludla, P.V.; Ziqubu, K.; Mthembu, S.X.H.; Mthiyane, F.; Nkambule, B.B.; Mazibuko-Mbeje, S.E. Rutin ameliorates inflammation and improves metabolic function: A comprehensive analysis of scientific literature. Pharmacol. Res. 2022, 178, 106163. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Qin, X.; Li, P.; Zhang, H.; Lin, T.; Miao, Z.; Ma, S. Isobavachalcone isolated from Psoralea corylifolia inhibits cell proliferation and induces apoptosis via inhibiting the AKT/GSK-3beta/beta-catenin pathway in colorectal cancer cells. Drug Des. Dev. Ther. 2019, 13, 1449–1460. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Lin, L.; Lu, J.J.; Chen, X. Pharmacological review of isobavachalcone, a naturally occurring chalcone. Pharmacol. Res. 2021, 165, 105483. [Google Scholar] [CrossRef]
- Limbach, J.R.; Espinosa, C.D.; Perez-Calvo, E.; Stein, H.H. Effect of dietary crude protein level on growth performance, blood characteristics, and indicators of intestinal health in weanling pigs. J. Anim. Sci. 2021, 99, skab166. [Google Scholar] [CrossRef]
- Payne, R.L.; Bidner, T.D.; Southern, L.L.; Geaghan, J.P. Effects of dietary soy isoflavones on growth, carcass traits, and meat quality in growing-finishing pigs. J. Anim. Sci. 2001, 79, 1230–1239. [Google Scholar] [CrossRef]
- Hariri, M.; Baradaran, H.R.; Gholami, A. The effect of soy isoflavones and soy isoflavones plus soy protein on serum concentration of tumor necrosis factor-alpha? A systematic review and meta-analysis of randomized controlled trials. Phytother. Res. 2021, 35, 3575–3589. [Google Scholar] [CrossRef]
- Smith, B.N.; Dilger, R.N. Immunomodulatory potential of dietary soybean-derived isoflavones and saponins in pigs. J. Anim. Sci. 2018, 96, 1288–1304. [Google Scholar] [CrossRef]
- Bo, Z.; Li, X. A Review of Pseudorabies Virus Variants: Genomics, Vaccination, Transmission, and Zoonotic Potential. Viruses 2022, 14, 1003. [Google Scholar] [CrossRef] [PubMed]
- Pomeranz, L.E.; Reynolds, A.E.; Hengartner, C.J. Molecular biology of pseudorabies virus: Impact on neurovirology and veterinary medicine. Microbiol. Mol. Biol. Rev. 2005, 69, 462–500. [Google Scholar] [CrossRef] [PubMed]
- Ai, J.W.; Weng, S.S.; Cheng, Q.; Cui, P.; Li, Y.J.; Wu, H.L.; Zhu, Y.M.; Xu, B.; Zhang, W.H. Human Endophthalmitis Caused By Pseudorabies Virus Infection, China, 2017. Emerg. Infect. Dis. 2018, 24, 1087–1090. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Han, H.; Wang, H.; Cui, Y.; Liu, H.; Ding, S. A Case of Human Viral Encephalitis Caused by Pseudorabies Virus Infection in China. Front. Neurol. 2019, 10, 534. [Google Scholar] [CrossRef]
- Wang, D.; Tao, X.; Fei, M.; Chen, J.; Guo, W.; Li, P.; Wang, J. Human encephalitis caused by pseudorabies virus infection: A case report. J. Neurovirol. 2020, 26, 442–448. [Google Scholar] [CrossRef]
- Sun, W.; Liu, S.; Lu, A.; Yang, F.; Duan, J. In vitro anti-PRV activity of dihydromyricetin from Ampelopsis grossedentata. Nat. Prod. Res. 2021, 1–4. [Google Scholar] [CrossRef]
- Liu, C.W.; Lin, H.W.; Yang, D.J.; Chen, S.Y.; Tseng, J.K.; Chang, T.J.; Chang, Y.Y. Luteolin inhibits viral-induced inflammatory response in RAW264.7 cells via suppression of STAT1/3 dependent NF-kappaB and activation of HO-1. Free. Radic. Biol. Med. 2016, 95, 180–189. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, S.; Yang, S.; Chen, C.; Yang, Y.; Lin, M.; Liu, C.; Wang, W.; Zhou, X.; Ai, Q.; et al. Mechanism of Dihydromyricetin on Inflammatory Diseases. Front. Pharmacol. 2021, 12, 794563. [Google Scholar] [CrossRef]
- Fan, L.; Zhao, X.; Tong, Q.; Zhou, X.; Chen, J.; Xiong, W.; Fang, J.; Wang, W.; Shi, C. Interactions of Dihydromyricetin, a Flavonoid from Vine Tea (Ampelopsis grossedentata) with Gut Microbiota. J. Food Sci. 2018, 83, 1444–1453. [Google Scholar] [CrossRef]
- Ye, Y.; Huang, Z.; Chen, M.; Mo, Y.; Mo, Z. Luteolin Potentially Treating Prostate Cancer and COVID-19 Analyzed by the Bioinformatics Approach: Clinical Findings and Drug Targets. Front. Endocrinol. 2021, 12, 802447. [Google Scholar] [CrossRef]
- Xie, Y.Z.; Peng, C.W.; Su, Z.Q.; Huang, H.T.; Liu, X.H.; Zhan, S.F.; Huang, X.F. A Practical Strategy for Exploring the Pharmacological Mechanism of Luteolin Against COVID-19/Asthma Comorbidity: Findings of System Pharmacology and Bioinformatics Analysis. Front. Immunol. 2021, 12, 769011. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, R.; Yuan, B.; Liu, Y.; Liu, C. The antiviral and antimicrobial activities of licorice, a widely-used Chinese herb. Acta Pharm. Sin. B 2015, 5, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Adnan, A.; Allaudin, Z.N.; Hani, H.; Loh, H.S.; Khoo, T.J.; Ting, K.N.; Abdullah, R. Virucidal activity of Garcinia parvifolia leaf extracts in animal cell culture. BMC Complementary Med. Ther. 2019, 19, 169. [Google Scholar] [CrossRef] [PubMed]
- Machado, V.B.; Marostica de Sa, J.; Miranda Prado, A.K.; Alves de Toledo, K.; Regasini, L.O.; Pereira de Souza, F.; Caruso, I.P.; Fossey, M.A. Biophysical and flavonoid-binding studies of the G protein ectodomain of group A human respiratory syncytial virus. Heliyon 2019, 5, e01394. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Noll, L.; Lu, N.; Porter, E.; Stoy, C.; Zheng, W.; Liu, X.; Peddireddi, L.; Niederwerder, M.; Bai, J. Genetic diversity and prevalence of porcine circovirus type 3 (PCV3) and type 2 (PCV2) in the Midwest of the USA during 2016–2018. Transbound. Emerg. Dis. 2020, 67, 1284–1294. [Google Scholar] [CrossRef]
- Opriessnig, T.; Karuppannan, A.K.; Halbur, P.G.; Calvert, J.G.; Nitzel, G.P.; Matzinger, S.R.; Meng, X.J. Porcine circovirus type 2a or 2b based experimental vaccines provide protection against PCV2d/porcine parvovirus 2 co-challenge. Vaccine 2020, 38, 1975–1981. [Google Scholar] [CrossRef]
- Kang, L.; Wahaab, A.; Shi, K.; Mustafa, B.E.; Zhang, Y.; Zhang, J.; Li, Z.; Qiu, Y.; Li, B.; Liu, K.; et al. Molecular Epidemic Characteristics and Genetic Evolution of Porcine Circovirus Type 2 (PCV2) in Swine Herds of Shanghai, China. Viruses 2022, 14, 289. [Google Scholar] [CrossRef]
- Doan, H.T.T.; Do, R.T.; Thao, P.T.P.; Le, X.T.K.; Nguyen, K.T.; Hien, N.T.T.; Duc, L.M.; Pham, L.T.K.; Le, T.H. Molecular genotypic analysis of porcine circovirus type 2 reveals the predominance of PCV2d in Vietnam (2018–2020) and the association between PCV2h, the recombinant forms, and Vietnamese vaccines. Arch. Virol. 2022, 167, 2011–2026. [Google Scholar] [CrossRef]
- Yi, J.; Chen, C.; Liu, X.; Kang, Q.; Hao, L.; Huang, J.; Lu, J. Radioprotection of EGCG based on immunoregulatory effect and antioxidant activity against (60)Cogamma radiation-induced injury in mice. Food Chem. Toxicol. 2020, 135, 111051. [Google Scholar] [CrossRef]
- Chaves, J.O.; de Souza, M.C.; da Silva, L.C.; Lachos-Perez, D.; Torres-Mayanga, P.C.; Machado, A.; Forster-Carneiro, T.; Vazquez-Espinosa, M.; Gonzalez-de-Peredo, A.V.; Barbero, G.F.; et al. Extraction of Flavonoids From Natural Sources Using Modern Techniques. Front. Chem. 2020, 8, 507887. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
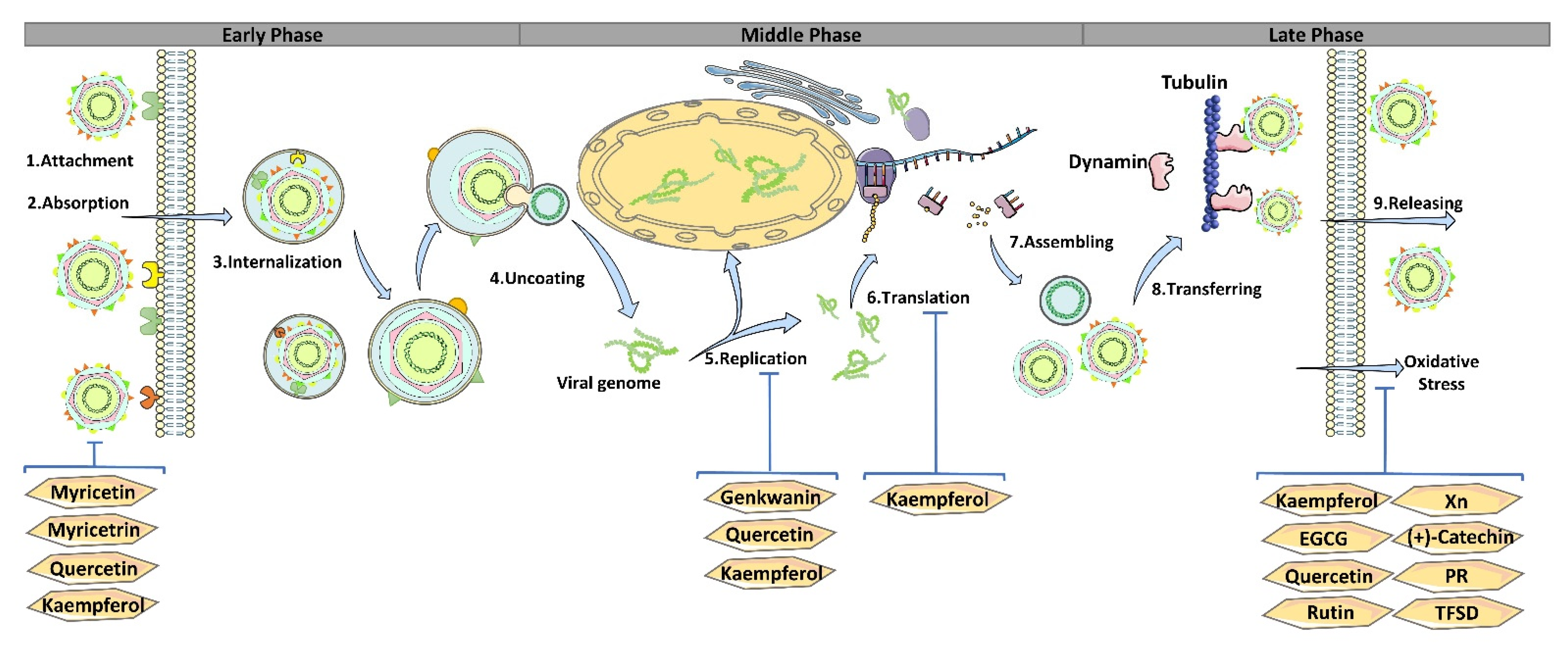
| Flavonoid | Structure | Virus | Treatment Time | Experimental Model | Effective Concentration | Inhibitory Phase | Mechanism | Reference |
|---|---|---|---|---|---|---|---|---|
| epigallocatechin-3-gallate (EGCG) |  | PRRSV | Pre | In vitro | 125 μM | Early phase | Block PRRSV binding to cells, reduce pro-inflammatory factors, and disturb lipid metabolism | [21,22,23,24] |
| PRV | Pre | In vivo/in vitro | 50 μM (in vitro) 40 mg/kg (in vivo) | Multiple steps | Inhibit PRV adsorption, entry, and replication | [25,26] | ||
| PCV2 | Pre | In silico/in vitro | 100 μM | Early phase | Interacts with heparan sulfate to competitively inhibit capsid binding | [27] | ||
| H9N2 | Post | In vivo | 10 mg/kg | Multiple steps | Reduce Organs damage, inflammation, and virus titer | [28] | ||
| quercetin |  | PRRSV | Post | In vivo | 10 mg/kg 100 μM in vivo | Multiple steps | Cross protective efficacy and inhibit the activity of Hsp70 | [29,30] |
| PEDV | Pre | In vitro | 100 μM | Early phase | Inhibit the activity of PEDV 3CL protease | [31,32,33,34] | ||
| PRV | Pre Simultaneity | In vitro In vivo | 50 μM (in vitro) 1.51 μg (in vivo) | Early phase | Interacting with the viral gD protein. Reducing the secretion of reactive oxygen species induced by PRV. | [35,36] | ||
| kaempferol |  | ASFV | Simultaneity Post | In vitro | 20 μg/mL | Early phase | Induced autophagy | [37] |
| PRV | In vitro | 240 mg/kg | Early phase | Reduced the expression level of viral IE180 and inhibit viral replication | [25] | |||
| H9N2 | Post | In vivo | 15 mg/kg | Later phase | Inhibit the NF-κB and MAPKs pathways mediated by TLR4/MyD88 NF-κB | [38] | ||
| hyperoside |  | PEDV | Pre | in vitro | 20 μM | Later phase | Inhibited N protein-induced S phase cell cycle arrest | [39] |
| rutin |  | PRRSV | Post | In vitro | 7.8 μg/mL | Later phase | Regulate inflammation and suppress PRRSV replication | [21] |
| PRV | Post | In vitro | 40 μg/mL | Later phase | Inhibit the activation of NF-κB and MAPK pathways | [40] | ||
| total flavonoids of S. suberectus Dunn (TFSD) | - | PCV2 | Post | In vitro/In vivo | 50–100 μg/mL (in vitro)/ 50–100 mg/kg (in vivo) | Later phase | Anti-oxidation and immunomodulatory effects | [41,42] |
| (+)-catechin |  | TGEV | Post | In vitro | 80 μM | Later phase | Anti-oxidation | [43,44] |
| myricetin |  | ASFV | Simultaneity | In vitro | 20 μM | Early phase | Interaction of 3, 4, 5-trihydroxyphenyl with ASFV protease | [45] |
| myricitrin |  | ASFV | Simultaneity | In vitro | 40 μM | Later phase | Interaction of 3, 4, 5-trihydroxyphenyl with ASFV protease | [45] |
| genistein |  | ASFV | Pre | In vitro | 50 μM | Early phase | Disrupt the synthesis of viral DNA | [46] |
| genkwanin |  | ASFV | Simultaneity | In silico/in vitro | 40 μM | Early phase, later phase | Inhibition of ASFV entry and release phases | [47,48] |
| apigenin |  | ASFV | Pre | In vitro | 50 μM | Early phase, later phase | Expression of 25-kD virus protein was inhibited | [47,48] |
| quercetin 7-rhamnoside (Q7R) | - | PEDV | Simultaneity | In vitro | 10 μg/mL | Early phase | Inhibit the early stage of viral replication | [34] |
| chalcone | - | H1N1 | Pre | In vitro | 2.49 ± 0.14 μg/mL | Later phase | Noncompetitive inhibitors of H1N1 neuraminidase | [49,50,51] |
| Isobavachalcone (IBC) |  | PRRSV | Post | In vitro | <15 μM | Later phase | Interference with RNA synthesis | [52] |
| PRV | Simultaneity | In vitro | 25.6 μM | Later phase | PRV replication was inhibited at the intercellular fusion stage | [53] | ||
| Xanthohumol (Xn) |  | PRRSV | Pre | In vivo/in vitro | 15 μM | Later phase | Nrf2-HMOX1 axis | [54,55] |
| Isoflavones (ISF) | PRRSV | Post | In vivo | 1600 mg/kg | Later phase | Supported immune responses | [56,57,58] | |
| Puerarin | PEDV | Post | In vivo | 0.5 mg/kg | Later phase | Alleviate systemic inflammation | [59,60] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Chen, S.; Li, X.; Zhang, L.; Ren, L. Flavonoids as Potential Antiviral Agents for Porcine Viruses. Pharmaceutics 2022, 14, 1793. https://doi.org/10.3390/pharmaceutics14091793
Zhang X, Chen S, Li X, Zhang L, Ren L. Flavonoids as Potential Antiviral Agents for Porcine Viruses. Pharmaceutics. 2022; 14(9):1793. https://doi.org/10.3390/pharmaceutics14091793
Chicago/Turabian StyleZhang, Xinwei, Si Chen, Xue Li, Liying Zhang, and Linzhu Ren. 2022. "Flavonoids as Potential Antiviral Agents for Porcine Viruses" Pharmaceutics 14, no. 9: 1793. https://doi.org/10.3390/pharmaceutics14091793