
Application of Nanomaterials in the Prevention, Detection, and Treatment of Methicillin-Resistant Staphylococcus aureus (MRSA)

Abstract
:1. Introduction
2. Vaccines and Nanovesicles
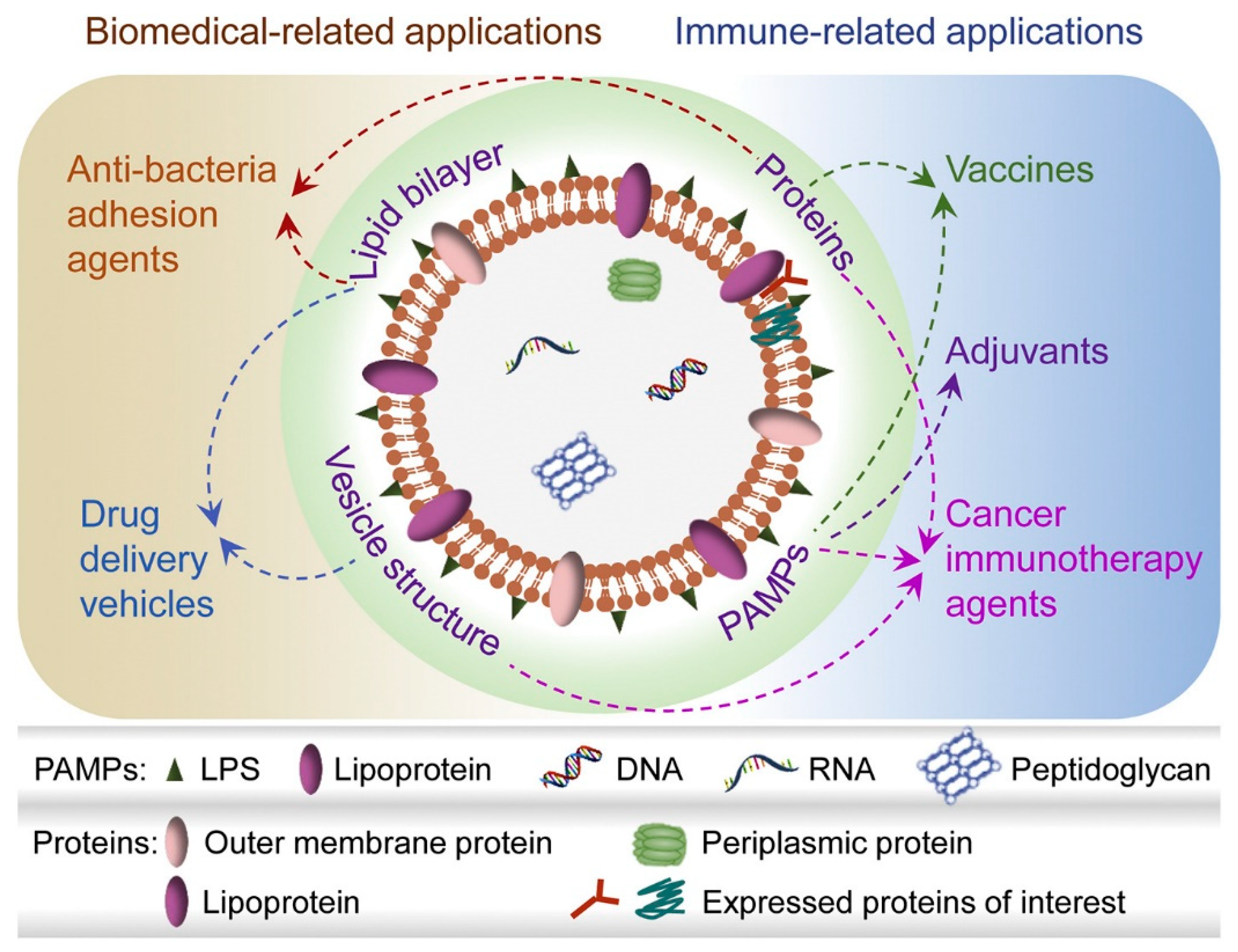
The Role of Gram-Negative and Positive Extracellular Vesicles in Vaccine Development
3. Multiple Roles of Nanomaterials in Rapid MRSA Diagnostics
Graphene Oxide and Fluorescent Nanomaterials
4. Antibacterial Agents
4.1. Metallic Nanoparticles
4.2. Liposomes
4.3. Polymeric Nanoparticles
4.4. Solid Lipid Nanoparticles
4.5. Stealth Coatings (Delivery and Detoxification)
5. Biofilms
6. Clinical Translations
7. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MRSA | methicillin-resistant Staphylococcus aureus |
| AS | antibiotic stewardship |
| AMR | antimicrobial resistance |
| NDM | Delhi Metallo |
| SA | S. aureus |
| VISA | vancomycin-intermediate S. aureus |
| h-VISA | heterogeneous VISA |
| VRSA | vancomycin-resistant S. aureus |
| NSA | narrow-spectrum antibiotics |
| MAD NAAT | multiplexable autonomous disposable nucleic acid amplification tests |
| Hla | a-hemolysin |
| PVL | Panton-Valentine leukocidin |
| PSM | phenol-soluble modulin |
| EV | extracellular vesicles |
| OMVs | outer membrane vesicles |
| PAMPs | pathogen-associated molecular patterns |
| LPS | lipopolysaccharide |
| LukED | leukotoxin ED |
| SelX | staphylococcal enterotoxin-like toxin X |
| PTT | photodynamic therapies |
| HA-MRSA | hospital-acquired MRSA |
| CA-MRSA | community-acquired MRSA |
References
- Nieuwlaat, R.; Mbuagbaw, L.; Mertz, D.; Burrows, L.L.; Bowdish, D.M.E.; Moja, L.; Wright, G.D.; Schünemann, H.J. Coronavirus Disease 2019 and Antimicrobial Resistance: Parallel and Interacting Health Emergencies. Clin. Infect. Dis. 2021, 72, 1657–1659. [Google Scholar] [CrossRef]
- Diekema, D.J.; Pfaller, M.A.; Shortridge, D.; Zervos, M.; Jones, R.N. Twenty-Year Trends in Antimicrobial Susceptibilities among Staphylococcus aureus from the SENTRY Antimicrobial Surveillance Program. Open Forum Infect. Dis. 2019, 6, S47–S53. [Google Scholar] [CrossRef]
- Iwu-Jaja, C.J.; Jaca, A.; Jaja, I.F.; Jordan, P.; Bhengu, P.; Iwu, C.D.; Okeibunor, J.; Karamagi, H.; Tumusiime, P.; Fuller, W.; et al. Preventing and Managing Antimicrobial Resistance in the African Region: A Scoping Review Protocol. PLoS ONE 2021, 16, e0254737. [Google Scholar] [CrossRef]
- Umar, L.W.; Isah, A.; Musa, S.; Umar, B. Prescribing Pattern and Antibiotic Use for Hospitalized Children in a Northern Nigerian Teaching Hospital. Ann. Afr. Med. 2018, 17, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Veeraraghavan, B.; Walia, K. Antimicrobial Susceptibility Profile & Resistance Mechanisms of Global Antimicrobial Resistance Surveillance System (GLASS) Priority Pathogens from India. Indian J. Med. Res. 2019, 149, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Purohit, M.R.; Lindahl, L.F.; Diwan, V.; Marrone, G.; Lundborg, C.S. High Levels of Drug Resistance in Commensal E. coli in a Cohort of Children from Rural Central India. Sci. Rep. 2019, 9, 6682. [Google Scholar] [CrossRef] [PubMed]
- Langford, B.J.; So, M.; Raybardhan, S.; Leung, V.; Westwood, D.; MacFadden, D.R.; Soucy, J.-P.R.; Daneman, N. Bacterial Co-Infection and Secondary Infection in Patients with COVID-19: A Living Rapid Review and Meta-Analysis. Clin. Microbiol. Infect. 2020, 26, 1622–1629. [Google Scholar] [CrossRef] [PubMed]
- Langford, B.J.; So, M.; Raybardhan, S.; Leung, V.; Soucy, J.-P.R.; Westwood, D.; Daneman, N.; MacFadden, D.R. Antibiotic Prescribing in Patients with COVID-19: Rapid Review and Meta-Analysis. Clin. Microbiol. Infect. 2021, 27, 520–531. [Google Scholar] [CrossRef] [PubMed]
- Kampmeier, S.; Tönnies, H.; Correa-Martinez, C.L.; Mellmann, A.; Schwierzeck, V. A Nosocomial Cluster of Vancomycin Resistant Enterococci among COVID-19 Patients in an Intensive Care Unit. Antimicrob. Resist. Infect. Control. 2020, 9, 154. [Google Scholar] [CrossRef]
- Porretta, A.D.; Baggiani, A.; Arzilli, G.; Casigliani, V.; Mariotti, T.; Mariottini, F.; Scardina, G.; Sironi, D.; Totaro, M.; Barnini, S.; et al. Increased Risk of Acquisition of New Delhi Metallo-Beta-Lactamase-Producing Carbapenem-Resistant Enterobacterales (NDM-CRE) among a Cohort of COVID-19 Patients in a Teaching Hospital in Tuscany, Italy. Pathogens 2020, 9, E635. [Google Scholar] [CrossRef] [PubMed]
- Rapacka-Zdonczyk, A.; Wozniak, A.; Nakonieczna, J.; Grinholc, M. Development of Antimicrobial Phototreatment Tolerance: Why the Methodology Matters. Int. J. Mol. Sci. 2021, 22, 2224. [Google Scholar] [CrossRef]
- Nori, P.; Cowman, K.; Chen, V.; Bartash, R.; Szymczak, W.; Madaline, T.; Punjabi Katiyar, C.; Jain, R.; Aldrich, M.; Weston, G.; et al. Bacterial and Fungal Coinfections in COVID-19 Patients Hospitalized during the New York City Pandemic Surge. Infect. Control Hosp. Epidemiol. 2021, 42, 84–88. [Google Scholar] [CrossRef] [PubMed]
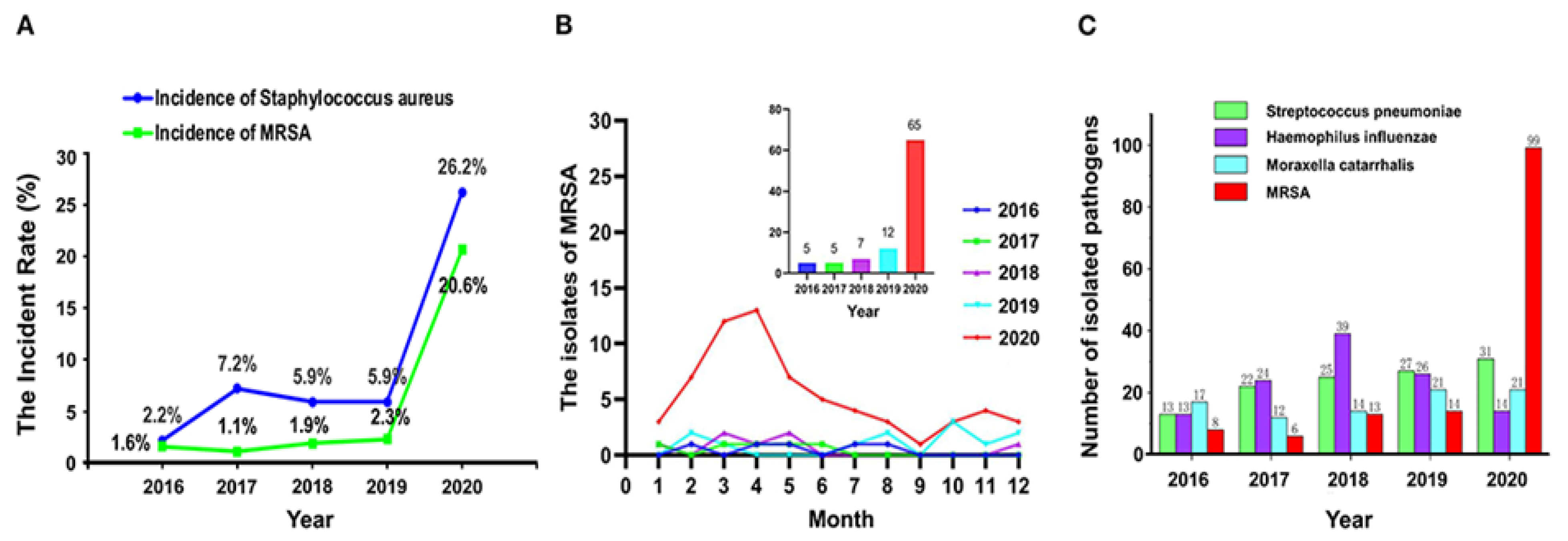
- Yang, M.; Feng, Y.; Yuan, L.; Zhao, H.; Gao, S.; Li, Z. High Concentration and Frequent Application of Disinfection Increase the Detection of Methicillin-Resistant Staphylococcus aureus Infections in Psychiatric Hospitals During the COVID-19 Pandemic. Front. Med. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Sabokroo, N.; Wang, Y.; Hashemian, M.; Karamollahi, S.; Kouhsari, E. Systematic Review and Meta-Analysis of the Epidemiology of Vancomycin-Resistance Staphylococcus aureus Isolates. Antimicrob. Resist. Infect. Control. 2021, 10, 101. [Google Scholar] [CrossRef]
- Diaz, R.; Ramalheira, E.; Afreixo, V.; Gago, B. Evaluation of Vancomycin MIC Creep in Staphylococcus aureus. J. Glob. Antimicrob. Resist. 2017, 10, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Goić-Barišić, I.; Radić, M.; Novak, A.; Rubić, Ž.; Boban, N.; Lukšić, B.; Tonkić, M. Vancomycin-Resistant Enterococcus Faecium COLONIZATION and Clostridium Difficile Infection in a HEMATOLOGIC Patient. Acta Clin. Croat. 2020, 59, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, S.; Samanta, I.; Bhattacharyya, D.; Nanda, P.K.; Kar, D.; Chowdhury, J.; Dandapat, P.; Das, A.K.; Batul, N.; Mondal, B.; et al. Co-Infection of Methicillin-Resistant Staphylococcus Epidermidis, Methicillin-Resistant Staphylococcus aureus and Extended Spectrum β-Lactamase Producing Escherichia Coli in Bovine Mastitis--Three Cases Reported from India. Vet. Q 2015, 35, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, C.; Pagano, P.; Pillar, C.M.; Shinabarger, D.L.; Boulos, R.A. Comparison of the in Vitro Antibacterial Activity of Ramizol, Fidaxomicin, Vancomycin, and Metronidazole against 100 Clinical Isolates of Clostridium Difficile by Broth Microdilution. Diagn. Microbiol. Infect. Dis. 2018, 92, 250–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shariati, A.; Dadashi, M.; Moghadam, M.T.; van Belkum, A.; Yaslianifard, S.; Darban-Sarokhalil, D. Global Prevalence and Distribution of Vancomycin Resistant, Vancomycin Intermediate and Heterogeneously Vancomycin Intermediate Staphylococcus aureus Clinical Isolates: A Systematic Review and Meta-Analysis. Sci. Rep. 2020, 10, 12689. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.Y.C.; Lye, D.C.; Yahav, D.; Sud, A.; Robinson, J.O.; Nelson, J.; Archuleta, S.; Roberts, M.A.; Cass, A.; Paterson, D.L.; et al. Effect of Vancomycin or Daptomycin with vs Without an Antistaphylococcal β-Lactam on Mortality, Bacteremia, Relapse, or Treatment Failure in Patients With MRSA Bacteremia: A Randomized Clinical Trial. JAMA 2020, 323, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, R.; Rayman, G. Point-of-Care Blood Glucose Testing for Diabetes Care in Hospitalized Patients. J. Diabetes Sci. Technol. 2014, 8, 1081–1090. [Google Scholar] [CrossRef]
- Lafleur, L.K.; Bishop, J.D.; Heiniger, E.K.; Gallagher, R.P.; Wheeler, M.D.; Kauffman, P.; Zhang, X.; Kline, E.C.; Buser, J.R.; Kumar, S.; et al. A Rapid, Instrument-Free, Sample-to-Result Nucleic Acid Amplification Test. Lab. Chip. 2016, 16, 3777–3787. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Chen, S.; Chen, Y.; Tian, L.; Wu, Q.; Zheng, M.; Li, Z. Alterations of Fecal Antibiotic Resistome in COVID-19 Patients after Empirical Antibiotic Exposure. Int. J. Hyg. Environ. Heal. 2022, 240, 113882. [Google Scholar] [CrossRef] [PubMed]
- Andreoni, F.; Toyofuku, M.; Menzi, C.; Kalawong, R.; Shambat, S.M.; François, P.; Zinkernagel, A.S.; Eberl, L. Antibiotics Stimulate Formation of Vesicles in Staphylococcus aureus in Both Phage-Dependent and -Independent Fashions and via Different Routes. Antimicrob. Agents Chemother. 2018, 63, e01439-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.W.; Seo, J.-S.; Park, S.B.; Lee, A.R.; Lee, J.S.; Jung, J.W.; Chun, J.H.; Lazarte, J.M.S.; Kim, J.; Kim, J.-H.; et al. Significant Increase in the Secretion of Extracellular Vesicles and Antibiotics Resistance from Methicillin-Resistant Staphylococcus aureus Induced by Ampicillin Stress. Sci. Rep. 2020, 10, 21066. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Yuan, F.; Lu, F.; Yin, Y.; Cao, J. Vancomycin-Induced Biofilm Formation by Methicillin-Resistant Staphylococcus aureus Is Associated with the Secretion of Membrane Vesicles. Microb. Pathog. 2017, 110, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Jansen, K.U.; Anderson, A.S. The Role of Vaccines in Fighting Antimicrobial Resistance (AMR). Hum. Vaccin. Immunother. 2018, 14, 2142–2149. [Google Scholar] [CrossRef] [Green Version]
- Giau, V.V.; Lee, H.; An, S.S.A.; Hulme, J. Recent Advances in the Treatment of C. Difficile Using Biotherapeutic Agents. Infect. Drug Resist. 2019, 12, 1597–1615. [Google Scholar] [CrossRef] [Green Version]
- Geoghegan, J.A.; Foster, T.J. Cell Wall-Anchored Surface Proteins of Staphylococcus aureus: Many Proteins, Multiple Functions. Curr. Top. Microbiol. Immunol. 2017, 409, 95–120. [Google Scholar] [CrossRef] [PubMed]
- Fazal, S.; Lee, R. Biomimetic Bacterial Membrane Vesicles for Drug Delivery Applications. Pharmaceutics 2021, 13, 1430. [Google Scholar] [CrossRef]
- Wang, X.; Koffi, P.F.; English, O.F.; Lee, J.C. Staphylococcus aureus Extracellular Vesicles: A Story of Toxicity and the Stress of 2020. Toxins 2021, 13, 75. [Google Scholar] [CrossRef] [PubMed]
- Raafat, D.; Otto, M.; Reppschläger, K.; Iqbal, J.; Holtfreter, S. Fighting Staphylococcus aureus Biofilms with Monoclonal Antibodies. Trends Microbiol. 2019, 27, 303–322. [Google Scholar] [CrossRef] [PubMed]
- Rupp, M.E.; Holley, H.P.; Lutz, J.; Dicpinigaitis, P.V.; Woods, C.W.; Levine, D.P.; Veney, N.; Fowler, V.G. Phase II, Randomized, Multicenter, Double-Blind, Placebo-Controlled Trial of a Polyclonal Anti-Staphylococcus aureus Capsular Polysaccharide Immune Globulin in Treatment of Staphylococcus aureus Bacteremia. Antimicrob. Agents Chemother. 2007, 51, 4249–4254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNeely, T.B.; Shah, N.A.; Fridman, A.; Joshi, A.; Hartzel, J.S.; Keshari, R.S.; Lupu, F.; DiNubile, M.J. Mortality among Recipients of the Merck V710 Staphylococcus aureus Vaccine after Postoperative S. aureus Infections: An Analysis of Possible Contributing Host Factors. Hum. Vaccin. Immunother. 2014, 10, 3513–3516. [Google Scholar] [CrossRef] [Green Version]
- Begier, E.; Seiden, D.J.; Patton, M.; Zito, E.; Severs, J.; Cooper, D.; Eiden, J.; Gruber, W.C.; Jansen, K.U.; Anderson, A.S.; et al. SA4Ag, a 4-Antigen Staphylococcus aureus Vaccine, Rapidly Induces High Levels of Bacteria-Killing Antibodies. Vaccine 2017, 35, 1132–1139. [Google Scholar] [CrossRef]
- Micoli, F.; Bagnoli, F.; Rappuoli, R.; Serruto, D. The Role of Vaccines in Combatting Antimicrobial Resistance. Nat. Rev. Microbiol. 2021, 19, 287–302. [Google Scholar] [CrossRef]
- Knox, K.W.; Vesk, M.; Work, E. Relation Between Excreted Lipopolysaccharide Complexes and Surface Structures of a Lysine-Limited Culture of Escherichia Coli. J. Bacteriol. 1966, 92, 1206–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, P.C.; Sharyan, A.; Sheikhi Moghaddam, L. Meningococcal Vaccines: Current Status and Emerging Strategies. Vaccines 2018, 6, E12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Zhou, H.; Yang, C.; Wu, Y.; Zhou, X.; Liu, H.; Wang, Y. Bacterial Outer Membrane Vesicles as a Platform for Biomedical Applications: An Update. J. Control. Release 2020, 323, 253–268. [Google Scholar] [CrossRef] [PubMed]
- Yur, D.; Lieser, R.M.; Sullivan, M.O.; Chen, W. Engineering Bionanoparticles for Improved Biosensing and Bioimaging. Curr. Opin. Biotechnol. 2021, 71, 41–48. [Google Scholar] [CrossRef]
- Kim, M.-R.; Hong, S.-W.; Choi, E.-B.; Lee, W.-H.; Kim, Y.-S.; Jeon, S.G.; Jang, M.H.; Gho, Y.S.; Kim, Y.-K. Staphylococcus aureus-Derived Extracellular Vesicles Induce Neutrophilic Pulmonary Inflammation via Both Th1 and Th17 Cell Responses. Allergy 2012, 67, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Lam, O.; Wheeler, J.; Tang, C.M. Thermal Control of Virulence Factors in Bacteria: A Hot Topic. Virulence 2014, 5, 852–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.-W.; Choi, E.-B.; Min, T.-K.; Kim, J.-H.; Kim, M.-H.; Jeon, S.G.; Lee, B.-J.; Gho, Y.S.; Jee, Y.-K.; Pyun, B.-Y.; et al. An Important Role of α-Hemolysin in Extracellular Vesicles on the Development of Atopic Dermatitis Induced by Staphylococcus aureus. PLoS ONE 2014, 9, e100499. [Google Scholar] [CrossRef]
- Li, Y.; Xiao, Q.; Tang, J.; Xiong, L.; Li, L. Extracellular Vesicles: Emerging Therapeutics in Cutaneous Lesions. Int. J. Nanomed. 2021, 16, 6183–6202. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Eagen, W.J.; Lee, J.C. Orchestration of Human Macrophage NLRP3 Inflammasome Activation by Staphylococcus aureus Extracellular Vesicles. Proc. Natl. Acad. Sci. USA 2020, 117, 3174–3184. [Google Scholar] [CrossRef] [PubMed]
- Tartaglia, N.R.; Nicolas, A.; Rodovalho, V.d.R.; Luz, B.S.R.d.; Briard-Bion, V.; Krupova, Z.; Thierry, A.; Coste, F.; Burel, A.; Martin, P.; et al. Extracellular Vesicles Produced by Human and Animal Staphylococcus aureus Strains Share a Highly Conserved Core Proteome. Sci. Rep. 2020, 10, 8467. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, J.; Park, J.; Gho, Y.S. Gram-Negative and Gram-Positive Bacterial Extracellular Vesicles. Semin. Cell Dev. Biol. 2015, 40, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Da Luz, B.S.R.; Nicolas, A.; Chabelskaya, S.; de Rezende Rodovalho, V.; Le Loir, Y.; de Carvalho Azevedo, V.A.; Felden, B.; Guédon, E. Environmental Plasticity of the RNA Content of Staphylococcus aureus Extracellular Vesicles. Front. Microbiol. 2021, 12, 634226. [Google Scholar] [CrossRef]
- Joshi, B.; Singh, B.; Nadeem, A.; Askarian, F.; Wai, S.N.; Johannessen, M.; Hegstad, K. Transcriptome Profiling of Staphylococcus aureus Associated Extracellular Vesicles Reveals Presence of Small RNA-Cargo. Front. Mol. Biosci. 2021, 7, 566207. [Google Scholar] [CrossRef] [PubMed]
- Kumaraswamy, M.; Wiull, K.; Joshi, B.; Sakoulas, G.; Kousha, A.; Vaaje-Kolstad, G.; Johannessen, M.; Hegstad, K.; Nizet, V.; Askarian, F. Bacterial Membrane-Derived Vesicles Attenuate Vancomycin Activity against Methicillin-Resistant Staphylococcus aureus. Microorganisms 2021, 9, 2055. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Thompson, C.D.; Weidenmaier, C.; Lee, J.C. Release of Staphylococcus aureus Extracellular Vesicles and Their Application as a Vaccine Platform. Nat. Commun. 2018, 9, 1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briaud, P.; Frey, A.; Marino, E.C.; Bastock, R.A.; Zielinski, R.E.; Wiemels, R.E.; Keogh, R.A.; Murphy, E.R.; Shaw, L.N.; Carroll, R.K. Temperature Influences the Composition and Cytotoxicity of Extracellular Vesicles in Staphylococcus aureus. mSphere 2021, 6, e0067621. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Hulme, J.P. Recent Advances in the Detection of Antibiotic and Multi-Drug Resistant Salmonella: An Update. Int. J. Mol. Sci. 2021, 22, 3499. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.; Vincent, J.-L.; Adhikari, N.K.J.; Machado, F.R.; Angus, D.C.; Calandra, T.; Jaton, K.; Giulieri, S.; Delaloye, J.; Opal, S.; et al. Sepsis: A Roadmap for Future Research. Lancet Infect. Dis. 2015, 15, 581–614. [Google Scholar] [CrossRef]
- Wu, H.; Moser, C.; Wang, H.-Z.; Høiby, N.; Song, Z.-J. Strategies for Combating Bacterial Biofilm Infections. Int. J. Oral Sci. 2015, 7, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malhotra-Kumar, S.; Haccuria, K.; Michiels, M.; Ieven, M.; Poyart, C.; Hryniewicz, W.; Goossens, H.; MOSAR WP2 Study Team. Current Trends in Rapid Diagnostics for Methicillin-Resistant Staphylococcus aureus and Glycopeptide-Resistant Enterococcus Species. J. Clin. Microbiol. 2008, 46, 1577–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faron, M.L.; Buchan, B.W.; Vismara, C.; Lacchini, C.; Bielli, A.; Gesu, G.; Liebregts, T.; Bree, A.v.; Jansz, A.; Soucy, G.; et al. Automated Scoring of Chromogenic Media for Detection of Methicillin-Resistant Staphylococcus aureus by Use of WASPLab Image Analysis Software. J. Clin. Microbiol. 2015, 54, 620–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srisrattakarn, A.; Tippayawat, P.; Chanawong, A.; Tavichakorntrakool, R.; Daduang, J.; Wonglakorn, L.; Lulitanond, A. Development of a Prototype Lateral Flow Immunoassay for Rapid Detection of Staphylococcal Protein A in Positive Blood Culture Samples. Diagnostics 2020, 10, E794. [Google Scholar] [CrossRef]
- Urushibara, N.; Aung, M.S.; Kawaguchiya, M.; Kobayashi, N. Novel Staphylococcal Cassette Chromosome Mec (SCCmec) Type XIV (5A) and a Truncated SCCmec Element in SCC Composite Islands Carrying SpeG in ST5 MRSA in Japan. J. Antimicrob. Chemother. 2020, 75, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Vandenesch, F.; Naimi, T.; Enright, M.C.; Lina, G.; Nimmo, G.R.; Heffernan, H.; Liassine, N.; Bes, M.; Greenland, T.; Reverdy, M.-E.; et al. Community-Acquired Methicillin-Resistant Staphylococcus aureus Carrying Panton-Valentine Leukocidin Genes: Worldwide Emergence. Emerg. Infect. Dis. 2003, 9, 978–984. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Kulkarni, K.; Behera, L.; Verma, V. Preparation and Characterization of Maghemite Nanoparticles from Mild Steel for Magnetically Guided Drug Therapy. J. Mater. Sci. Mater. Med. 2017, 28, 116. [Google Scholar] [CrossRef]
- Mocan, T.; Matea, C.T.; Pop, T.; Mosteanu, O.; Buzoianu, A.D.; Puia, C.; Iancu, C.; Mocan, L. Development of Nanoparticle-Based Optical Sensors for Pathogenic Bacterial Detection. J. Nanobiotechnol. 2017, 15, 25. [Google Scholar] [CrossRef] [Green Version]
- Kearns, H.; Goodacre, R.; Jamieson, L.E.; Graham, D.; Faulds, K. SERS Detection of Multiple Antimicrobial-Resistant Pathogens Using Nanosensors. Anal. Chem. 2017, 89, 12666–12673. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wang, C.; Shi, L.; Shao, L.; Fu, P.; Wang, K.; Xiao, R.; Wang, S.; Gu, B. Rapid Identification and Antibiotic Susceptibility Test of Pathogens in Blood Based on Magnetic Separation and Surface-Enhanced Raman Scattering. Mikrochim. Acta 2019, 186, 475. [Google Scholar] [CrossRef] [PubMed]
- Potluri, P.R.; Rajendran, V.K.; Sunna, A.; Wang, Y. Rapid and Specific Duplex Detection of Methicillin-Resistant Staphylococcus aureus Genes by Surface-Enhanced Raman Spectroscopy. Analyst 2020, 145, 2789–2794. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Tang, M.; Liu, Y.; Huang, J.; Liu, Z.; Tian, H.; Zheng, Y.; de la Chapelle, M.L.; Zhang, Y.; Fu, W. Surface-Enhanced Raman Scattering Method for the Identification of Methicillin-Resistant Staphylococcus aureus Using Positively Charged Silver Nanoparticles. Microchim. Acta 2019, 186, 102. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Ge, W.; Li, Q.; Wu, Y.; Jiang, H.; Wang, X. Rapid and Ultrasensitive Electrochemical Detection of Multidrug-Resistant Bacteria Based on Nanostructured Gold Coated ITO Electrode. ACS Appl. Mater. Interfaces 2014, 6, 11025–11031. [Google Scholar] [CrossRef] [PubMed]
- Storhoff, J.J.; Marla, S.S.; Bao, P.; Hagenow, S.; Mehta, H.; Lucas, A.; Garimella, V.; Patno, T.; Buckingham, W.; Cork, W.; et al. Gold Nanoparticle-Based Detection of Genomic DNA Targets on Microarrays Using a Novel Optical Detection System. Biosens. Bioelectron. 2004, 19, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.-S.; Tang, B.S.F.; Boost, M.V.; Chow, C.; Leung, P.H.M. Detection of Methicillin-Resistant Staphylococcus aureus Using a Gold Nanoparticle-Based Colourimetric Polymerase Chain Reaction Assay. Biosens. Bioelectron. 2014, 53, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Abd-El-Hady, H.; El-Said, W.; El-Enbaawy, M.; Taher, A.; Eldin, S. Preparation of mecA Biosensor Based on Gold Nanoparticles to Determine Methicillin Resistant Staphylococcus aureus (MRSA) Strains from Human and Animals. IOSR-JAVS 2014, 7, 64–71. [Google Scholar] [CrossRef]
- Krishnan, T.; Wang, H.-N.; Vo-Dinh, T. Smartphone-Based Device for Colorimetric Detection of MicroRNA Biomarkers Using Nanoparticle-Based Assay. Sensors 2021, 21, 8044. [Google Scholar] [CrossRef] [PubMed]
- Suaifan, G.A.R.Y.; Alhogail, S.; Zourob, M. Rapid and Low-Cost Biosensor for the Detection of Staphylococcus aureus. Biosens. Bioelectron. 2017, 90, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, R.; Buckingham, W.; Domanus, M.; Gieser, L.; Klein, K.; Kunkel, G.; Prokhorova, A.; Riccelli, P.V. Sensitive Assay for Identification of Methicillin-Resistant Staphylococcus aureus, Based on Direct Detection of Genomic DNA by Use of Gold Nanoparticle Probes. Clin. Chem. 2004, 50, 1949–1952. [Google Scholar] [CrossRef]
- Corrigan, D.K.; Schulze, H.; Henihan, G.; Hardie, A.; Ciani, I.; Giraud, G.; Terry, J.G.; Walton, A.J.; Pethig, R.; Ghazal, P.; et al. Development of a PCR-Free Electrochemical Point of Care Test for Clinical Detection of Methicillin Resistant Staphylococcus aureus (MRSA). Analyst 2013, 138, 6997–7005. [Google Scholar] [CrossRef] [PubMed]
- Senok, A.; Monecke, S.; Nassar, R.; Celiloglu, H.; Thyagarajan, S.; Müller, E.; Ehricht, R. Lateral Flow Immunoassay for the Detection of Panton-Valentine Leukocidin in Staphylococcus aureus From Skin and Soft Tissue Infections in the United Arab Emirates. Front. Cell Infect. Microbiol. 2021, 11, 754523. [Google Scholar] [CrossRef]
- Daccache, A.; Haddad, J.; Ghanem, A.; Feghali, E.J.; El Osta, B. Cough-Induced Rib Fracture in a Smoker: A Case Report. J. Med. Case Rep. 2020, 14, 147. [Google Scholar] [CrossRef]
- Yang, A.K.L.; Lu, H.; Wu, S.Y.; Kwok, H.C.; Ho, H.P.; Yu, S.; Cheung, A.K.L.; Kong, S.K. Detection of Panton-Valentine Leukocidin DNA from Methicillin-Resistant Staphylococcus aureus by Resistive Pulse Sensing and Loop-Mediated Isothermal Amplification with Gold Nanoparticles. Anal. Chim. Acta 2013, 782, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-H.; Lien, K.-Y.; Wu, J.-J.; Lee, G.-B. A Magnetic Bead-Based Assay for the Rapid Detection of Methicillin-Resistant Staphylococcus aureus by Using a Microfluidic System with Integrated Loop-Mediated Isothermal Amplification. Lab. Chip. 2011, 11, 1521–1531. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Öçsoy, I.; Yuan, Q.; Wang, R.; You, M.; Zhao, Z.; Song, E.; Zhang, X.; Tan, W. One-Step Facile Surface Engineering of Hydrophobic Nanocrystals with Designer Molecular Recognition. J. Am. Chem. Soc. 2012, 134, 13164–13167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekhon, S.S.; Kaur, P.; Kim, Y.-H.; Sekhon, S.S. 2D Graphene Oxide–Aptamer Conjugate Materials for Cancer Diagnosis. npj 2D Mater. Appl. 2021, 5, 1–19. [Google Scholar] [CrossRef]
- Kong, R.-M.; Zhang, X.-B.; Chen, Z.; Tan, W. Aptamer-Assembled Nanomaterials for Biosensing and Biomedical Applications. Small 2011, 7, 2428–2436. [Google Scholar] [CrossRef] [PubMed]
- Ocsoy, I.; Yusufbeyoglu, S.; Yılmaz, V.; McLamore, E.S.; Ildız, N.; Ülgen, A. DNA Aptamer Functionalized Gold Nanostructures for Molecular Recognition and Photothermal Inactivation of Methicillin-Resistant Staphylococcus aureus. Colloids Surf. B Biointerfaces 2017, 159, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Wang, L.; Zhao, K.; Li, N.; Shi, Z.; Ge, Z.; Jin, Z. Fabrication, Mechanical Properties, and Biocompatibility of Graphene-Reinforced Chitosan Composites. Biomacromolecules 2010, 11, 2345–2351. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Wan, J.; Zhang, S.; Zhang, Y.; Lee, S.-T.; Liu, Z. In Vivo Pharmacokinetics, Long-Term Biodistribution, and Toxicology of PEGylated Graphene in Mice. ACS Nano 2011, 5, 516–522. [Google Scholar] [CrossRef]
- Wan, C.; Chen, B. Poly(ε-Caprolactone)/Graphene Oxide Biocomposites: Mechanical Properties and Bioactivity. Biomed. Mater. 2011, 6, 055010. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Qiu, Z. Crystallization Behaviors of Biodegradable Poly(l-Lactic Acid)/Graphene Oxide Nanocomposites from the Amorphous State. Thermochim. Acta 2011, 526, 229–236. [Google Scholar] [CrossRef]
- Morimune, S.; Nishino, T.; Goto, T. Poly(Vinyl Alcohol)/Graphene Oxide Nanocomposites Prepared by a Simple Eco-Process. Polym. J. 2012, 44, 1056–1063. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Zhang, J.; Chen, P.; Zhou, X.; Yang, Y.; Wu, S.; Niu, L.; Han, Y.; Wang, L.; Chen, P.; et al. Label-Free, Electrochemical Detection of Methicillin-Resistant Staphylococcus aureus DNA with Reduced Graphene Oxide-Modified Electrodes. Biosens. Bioelectron. 2011, 26, 3881–3886. [Google Scholar] [CrossRef]
- Ning, Y.; Gao, Q.; Zhang, X.; Wei, K.; Chen, L. A Graphene Oxide–Based Sensing Platform for the Determination of Methicillin-Resistant Staphylococcus aureus Based on Strand-Displacement Polymerization Recycling and Synchronous Fluorescent Signal Amplification. J. Biomol. Screen 2016, 21, 851–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ning, Y.; Zou, L.; Gao, Q.; Hu, J.; Lu, F. Graphene Oxide-Based Fluorometric Determination of Methicillin-Resistant Staphylococcus aureus by Using Target-Triggered Chain Reaction and Deoxyribonuclease-Assisted Recycling. Microchim. Acta 2018, 185, 183. [Google Scholar] [CrossRef]
- Liu, J.; Cheng, J.; Zhang, Y. Upconversion Nanoparticle Based LRET System for Sensitive Detection of MRSA DNA Sequence. Biosens. Bioelectron. 2013, 43, 252–256. [Google Scholar] [CrossRef]
- Venkataramana, M.; Kurkuri, M.D.; Balakrishna, R.G. Simple quantum dot bioprobe/label for sensitive detection of Staphylococcus aureus TNase. Sensors Actuators B Chem. 2016, 222, 1201–1208. [Google Scholar] [CrossRef]
- Huh, A.J.; Kwon, Y.J. “Nanoantibiotics”: A New Paradigm for Treating Infectious Diseases Using Nanomaterials in the Antibiotics Resistant Era. J. Control. Release 2011, 156, 128–145. [Google Scholar] [CrossRef] [PubMed]
- Gurunathan, S.; Han, J.W.; Dayem, A.A.; Eppakayala, V.; Kim, J.-H. Oxidative Stress-Mediated Antibacterial Activity of Graphene Oxide and Reduced Graphene Oxide in Pseudomonas Aeruginosa. Int. J. Nanomed. 2012, 7, 5901–5914. [Google Scholar] [CrossRef] [Green Version]
- Leung, Y.H.; Ng, A.M.C.; Xu, X.; Shen, Z.; Gethings, L.A.; Wong, M.T.; Chan, C.M.N.; Guo, M.Y.; Ng, Y.H.; Djurišić, A.B.; et al. Mechanisms of Antibacterial Activity of MgO: Non-ROS Mediated Toxicity of MgO Nanoparticles towards Escherichia Coli. Small 2014, 10, 1171–1183. [Google Scholar] [CrossRef] [PubMed]
- Zakharova, O.V.; Godymchuk, A.Y.; Gusev, A.A.; Gulchenko, S.I.; Vasyukova, I.A.; Kuznetsov, D.V. Considerable Variation of Antibacterial Activity of Cu Nanoparticles Suspensions Depending on the Storage Time, Dispersive Medium, and Particle Sizes. Biomed. Res. Int. 2015, 2015, 412530. [Google Scholar] [CrossRef] [PubMed]
- Morones, J.R.; Elechiguerra, J.L.; Camacho, A.; Holt, K.; Kouri, J.B.; Ramírez, J.T.; Yacaman, M.J. The Bactericidal Effect of Silver Nanoparticles. Nanotechnology 2005, 16, 2346–2353. [Google Scholar] [CrossRef] [Green Version]
- Nabikhan, A.; Kandasamy, K.; Raj, A.; Alikunhi, N.M. Synthesis of Antimicrobial Silver Nanoparticles by Callus and Leaf Extracts from Saltmarsh Plant, Sesuvium Portulacastrum L. Colloids Surf. B Biointerfaces 2010, 79, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Dospivova, D.; Hynek, D.; Kopel, P.; Bezdekov, A.; Sochor, J.; Krizkova, S.; Vojtech, A.; Trnkova, L.; Hubalek, J.; Babula, P.; et al. Voltammetry of Adiponectin and its Interactions with Collagen on a Carbon Paste Electrode at Femtogram Level. Int. J. Electrochem. Sci. 2012, 7. Available online: http://www.electrochemsci.org/list12.htm (accessed on 1 March 2022).
- Lambert, P.A. Cellular Impermeability and Uptake of Biocides and Antibiotics in Gram-Positive Bacteria and Mycobacteria. J. Appl. Microbiol. 2002, 92, 46S–54S. [Google Scholar] [CrossRef] [PubMed]
- Grzelak, A.; Wojewódzka, M.; Meczynska-Wielgosz, S.; Zuberek, M.; Wojciechowska, D.; Kruszewski, M. Crucial Role of Chelatable Iron in Silver Nanoparticles Induced DNA Damage and Cytotoxicity. Redox Biol. 2018, 15, 435–440. [Google Scholar] [CrossRef]
- Holmila, R.J.; Vance, S.A.; King, S.B.; Tsang, A.W.; Singh, R.; Furdui, C.M. Silver Nanoparticles Induce Mitochondrial Protein Oxidation in Lung Cells Impacting Cell Cycle and Proliferation. Antioxidants 2019, 8, 552. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, C.A.; Seckler, M.M.; Ingle, A.P.; Gupta, I.; Galdiero, S.; Galdiero, M.; Gade, A.; Rai, M. Silver Nanoparticles: Therapeutical Uses, Toxicity, and Safety Issues. J. Pharm. Sci. 2014, 103, 1931–1944. [Google Scholar] [CrossRef]
- Strauch, B.M.; Niemand, R.K.; Winkelbeiner, N.L.; Hartwig, A. Comparison between Micro- and Nanosized Copper Oxide and Water Soluble Copper Chloride: Interrelationship between Intracellular Copper Concentrations, Oxidative Stress and DNA Damage Response in Human Lung Cells. Part. Fibre Toxicol. 2017, 14, 28. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.A.; Khan, H.M.; Khan, A.A.; Sultan, A.; Azam, A. Characterization of Clinical Strains of MSSA, MRSA and MRSE Isolated from Skin and Soft Tissue Infections and the Antibacterial Activity of ZnO Nanoparticles. World J. Microbiol. Biotechnol. 2012, 28, 1605–1613. [Google Scholar] [CrossRef] [PubMed]
- Ahmar Rauf, M.; Owais, M.; Rajpoot, R.; Ahmad, F.; Khan, N.; Zubair, S. Biomimetically Synthesized ZnO Nanoparticles Attain Potent Antibacterial Activity against Less Susceptible S. aureus Skin Infection in Experimental Animals. RSC Adv. 2017, 7, 36361–36373. [Google Scholar] [CrossRef] [Green Version]
- Umamageswari, S.S.M.; Manipriya, B.; Kalyani, M. Evaluation of Antibacterial Activity of Zinc Oxide Nanoparticles against Biofilm Producing Methicillin Resistant Staphylococcus aureus (MRSA). Res. J. Pharm. Technol. 2018, 11, 1884–1888. [Google Scholar] [CrossRef]
- Kadiyala, U.; Turali-Emre, E.S.; Bahng, J.H.; Kotov, N.A.; VanEpps, J.S. Unexpected Insights into Antibacterial Activity of Zinc Oxide Nanoparticles against Methicillin Resistant Staphylococcus aureus (MRSA). Nanoscale 2018, 10, 4927–4939. [Google Scholar] [CrossRef]
- Roy, A.S.; Parveen, A.; Koppalkar, A.R.; Prasad, M.V.N.A. Effect of Nano-Titanium Dioxide with Different Antibiotics against Methicillin-Resistant Staphylococcus aureus. J. Biomater. Nanobiotechnol. 2010, 1, 37–41. [Google Scholar] [CrossRef] [Green Version]
- Wahab, R.; Siddiqui, M.A.; Saquib, Q.; Dwivedi, S.; Ahmad, J.; Musarrat, J.; Al-Khedhairy, A.A.; Shin, H.-S. ZnO Nanoparticles Induced Oxidative Stress and Apoptosis in HepG2 and MCF-7 Cancer Cells and Their Antibacterial Activity. Colloids Surf. B Biointerfaces 2014, 117, 267–276. [Google Scholar] [CrossRef]
- Antoine, T.E.; Hadigal, S.R.; Yakoub, A.M.; Mishra, Y.K.; Bhattacharya, P.; Haddad, C.; Valyi-Nagy, T.; Adelung, R.; Prabhakar, B.S.; Shukla, D. Intravaginal Zinc Oxide Tetrapod Nanoparticles as Novel Immunoprotective Agents against Genital Herpes. J. Immunol. 2016, 196, 4566–4575. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, J.; Wahab, R.; Siddiqui, M.A.; Saquib, Q.; Al-Khedhairy, A.A. Cytotoxicity and cell death induced by engineered nanostructures (quantum dots and nanoparticles) in human cell lines. JBIC J. Biol. Inorg. Chem. 2020, 25, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Abebe, B.; Zereffa, E.A.; Tadesse, A.; Murthy, H.C.A. A Review on Enhancing the Antibacterial Activity of ZnO: Mechanisms and Microscopic Investigation. Nanoscale Res. Lett. 2020, 15, 190. [Google Scholar] [CrossRef]
- Khalid, A.; Ahmad, P.; Alharthi, A.I.; Muhammad, S.; Khandaker, M.U.; Faruque, M.R.I.; Din, I.U.; Alotaibi, M.A.; Khan, A. Synergistic Effects of Cu-Doped ZnO Nanoantibiotic against Gram-Positive Bacterial Strains. PLoS ONE 2021, 16, e0251082. [Google Scholar] [CrossRef] [PubMed]
- Khalid, A.; Ahmad, P.; Alharthi, A.I.; Muhammad, S.; Khandaker, M.U.; Faruque, M.R.I.; Khan, A.; Din, I.U.; Alotaibi, M.A.; Alzimami, K.; et al. Enhanced Optical and Antibacterial Activity of Hydrothermally Synthesized Cobalt-Doped Zinc Oxide Cylindrical Microcrystals. Materials 2021, 14, 3223. [Google Scholar] [CrossRef] [PubMed]
- Goldmann, O.; Cern, A.; Müsken, M.; Rohde, M.; Weiss, W.; Barenholz, Y.; Medina, E. Liposomal Mupirocin Holds Promise for Systemic Treatment of Invasive Staphylococcus aureus Infections. J. Control. Release 2019, 316, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Hajiahmadi, F.; Alikhani, M.Y.; Shariatifar, H.; Arabestani, M.R.; Ahmadvand, D. The Bactericidal Effect of Liposomal Vancomycin as a Topical Combating System against Methicillin-Resistant Staphylococcus aureus Skin Wound Infection in Mice. Med. J. Islam Repub. Iran 2019, 33, 153. [Google Scholar] [CrossRef] [PubMed]
- Immordino, M.L.; Dosio, F.; Cattel, L. Stealth Liposomes: Review of the Basic Science, Rationale, and Clinical Applications, Existing and Potential. Int. J. Nanomed. 2006, 1, 297–315. [Google Scholar]
- Sande, L.; Sanchez, M.; Montes, J.; Wolf, A.J.; Morgan, M.A.; Omri, A.; Liu, G.Y. Liposomal Encapsulation of Vancomycin Improves Killing of Methicillin-Resistant Staphylococcus aureus in a Murine Infection Model. J. Antimicrob. Chemother. 2012, 67, 2191–2194. [Google Scholar] [CrossRef] [Green Version]
- Serri, A.; Mahboubi, A.; Zarghi, A.; Moghimi, H. Investigating the Antimicrobial Efficacy of Liposomal Vancomycin in Gram-Positive and Gram-Negative Bacteria- A Preliminary Mechanistic Study. Iran. J. Pharm. Sci. 2018, 14, 13–24. [Google Scholar] [CrossRef]
- Abrishami, M.; Motamed Shariati, M.; Malaekeh-Nikouei, B.; Tajani, A.S.; Mahmoudi, A.; Abrishami, M.; Khameneh, B. Preparation and in Vivo Evaluation of Nanoliposomes Containing Vancomycin after Intravitreal Injection in Albino Rabbits. Iran J. Basic Med. Sci. 2020, 23, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Xiao, F.; Xing, D.; Hu, X. Polyprodrug Antimicrobials: Remarkable Membrane Damage and Concurrent Drug Release to Combat Antibiotic Resistance of Methicillin-Resistant Staphylococcus aureus. Small 2018, 14, e1802008. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Xu, G.; Pranantyo, D.; Xu, L.Q.; Neoh, K.-G.; Kang, E.-T. PH-Sensitive Zwitterionic Polymer as an Antimicrobial Agent with Effective Bacterial Targeting. ACS Biomater. Sci. Eng. 2018, 4, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Omolo, C.A.; Megrab, N.A.; Kalhapure, R.S.; Agrawal, N.; Jadhav, M.; Mocktar, C.; Rambharose, S.; Maduray, K.; Nkambule, B.; Govender, T. Liposomes with PH Responsive “on and off” Switches for Targeted and Intracellular Delivery of Antibiotics. J. Liposome Res. 2021, 31, 45–63. [Google Scholar] [CrossRef] [PubMed]
- Makhathini, S.S.; Kalhapure, R.S.; Jadhav, M.; Waddad, A.Y.; Gannimani, R.; Omolo, C.A.; Rambharose, S.; Mocktar, C.; Govender, T. Novel Two-Chain Fatty Acid-Based Lipids for Development of Vancomycin PH-Responsive Liposomes against Staphylococcus aureus and Methicillin-Resistant Staphylococcus aureus (MRSA). J. Drug Target. 2019, 27, 1094–1107. [Google Scholar] [CrossRef] [PubMed]
- Scriboni, A.B.; Couto, V.M.; Ribeiro, L.N.d.M.; Freires, I.A.; Groppo, F.C.; de Paula, E.; Franz-Montan, M.; Cogo-Müller, K. Fusogenic Liposomes Increase the Antimicrobial Activity of Vancomycin Against Staphylococcus aureus Biofilm. Front. Pharmacol. 2019, 10, 1401. [Google Scholar] [CrossRef] [PubMed]
- Lister, J.L.; Horswill, A.R. Staphylococcus aureus Biofilms: Recent Developments in Biofilm Dispersal. Front. Cell. Infect. Microbiol. 2014, 4, 178. [Google Scholar] [CrossRef] [Green Version]
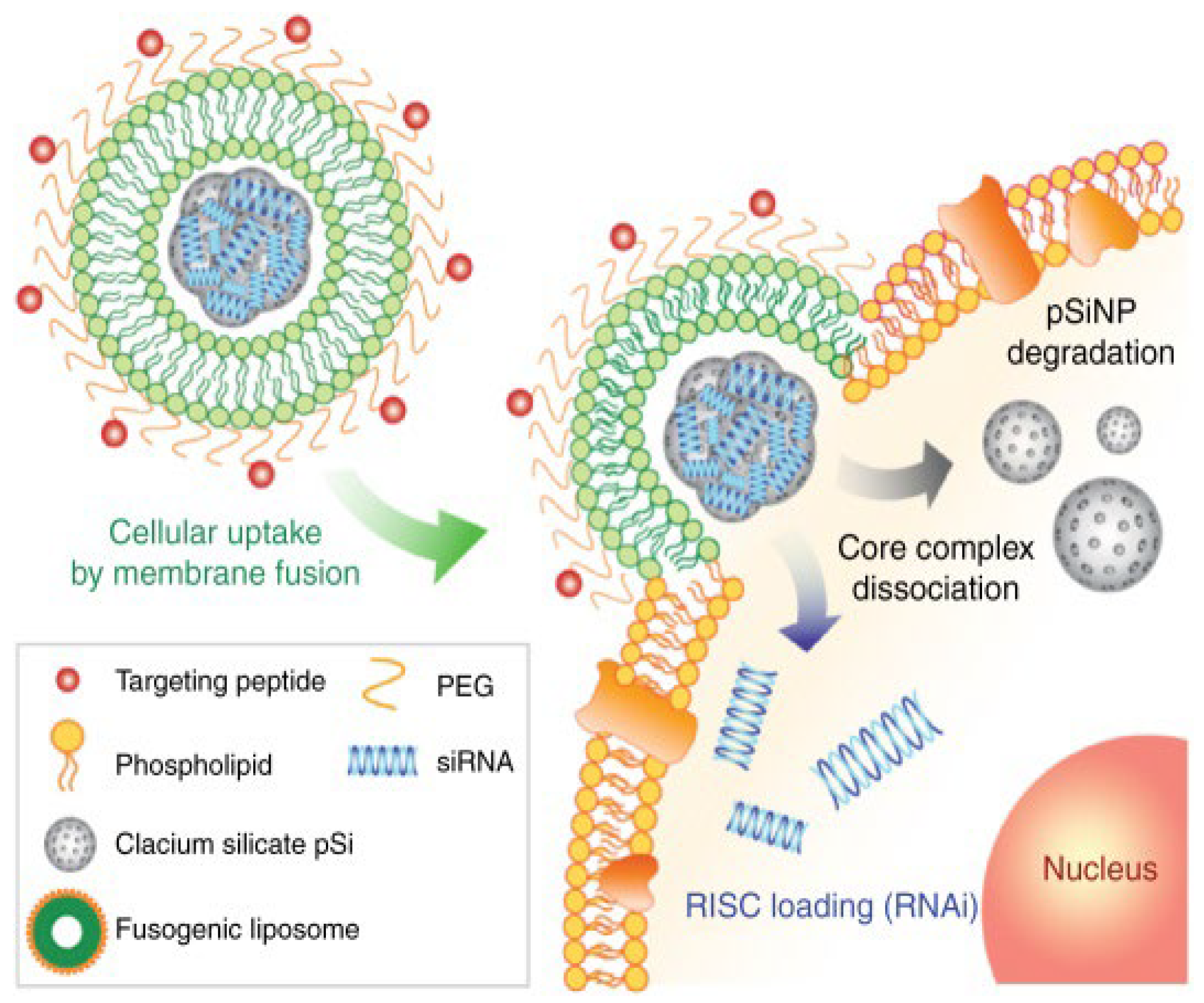
- Kim, B.; Pang, H.-B.; Kang, J.; Park, J.-H.; Ruoslahti, E.; Sailor, M.J. Immunogene Therapy with Fusogenic Nanoparticles Modulates Macrophage Response to Staphylococcus aureus. Nat. Commun. 2018, 9, 1969. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhang, L.; Liu, H.; Cheng, K. Delivery Strategies of the CRISPR-Cas9 Gene-Editing System for Therapeutic Applications. J. Control Release 2017, 266, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Gründling, A.; Schneewind, O. Cross-Linked Peptidoglycan Mediates Lysostaphin Binding to the Cell Wall Envelope of Staphylococcus aureus. J. Bacteriol. 2006, 188, 2463–2472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pornpattananangkul, D.; Zhang, L.; Olson, S.; Aryal, S.; Obonyo, M.; Vecchio, K.; Huang, C.-M.; Zhang, L. Bacterial Toxin-Triggered Drug Release from Gold Nanoparticle-Stabilized Liposomes for the Treatment of Bacterial Infection. J. Am. Chem. Soc. 2011, 133, 4132–4139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajiahmadi, F.; Alikhani, M.Y.; Shariatifar, H.; Arabestani, M.R.; Ahmadvand, D. The Bactericidal Effect of Lysostaphin Coupled with Liposomal Vancomycin as a Dual Combating System Applied Directly on Methicillin-Resistant Staphylococcus aureus Infected Skin Wounds in Mice. Int. J. Nanomed. 2019, 14, 5943–5955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nwabuife, J.C.; Pant, A.M.; Govender, T. Liposomal Delivery Systems and Their Applications against Staphylococcus aureus and Methicillin-Resistant Staphylococcus aureus. Adv. Drug Deliv. Rev. 2021, 178, 113861. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.; Ogren, M.; Dias, J.N.R.; Silva, M.; Gil, S.; Tavares, L.; Aires-da-Silva, F.; Gaspar, M.M.; Aguiar, S.I. Liposomes as Antibiotic Delivery Systems: A Promising Nanotechnological Strategy against Antimicrobial Resistance. Molecules 2021, 26, 2047. [Google Scholar] [CrossRef] [PubMed]
- Mat Rani, N.N.I.; Mustafa Hussein, Z.; Mustapa, F.; Azhari, H.; Sekar, M.; Chen, X.Y.; Mohd Amin, M.C.I. Exploring the Possible Targeting Strategies of Liposomes against Methicillin-Resistant Staphylococcus aureus (MRSA). Eur. J. Pharm. Biopharm. 2021, 165, 84–105. [Google Scholar] [CrossRef]
- Rukavina, Z.; Šegvić Klarić, M.; Filipović-Grčić, J.; Lovrić, J.; Vanić, Ž. Azithromycin-Loaded Liposomes for Enhanced Topical Treatment of Methicillin-Resistant Staphyloccocus Aureus (MRSA) Infections. Int. J. Pharm. 2018, 553, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Nicolle, L.; Journot, C.M.A.; Gerber-Lemaire, S. Chitosan Functionalization: Covalent and Non-Covalent Interactions and Their Characterization. Polymers 2021, 13, 4118. [Google Scholar] [CrossRef] [PubMed]
- Garg, U.; Chauhan, S.; Nagaich, U.; Jain, N. Current Advances in Chitosan Nanoparticles Based Drug Delivery and Targeting. Adv. Pharm. Bull. 2019, 9, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Jamil, B.; Habib, H.; Abbasi, S.A.; Ihsan, A.; Nasir, H.; Imran, M. Development of Cefotaxime Impregnated Chitosan as Nano-Antibiotics: De Novo Strategy to Combat Biofilm Forming Multi-Drug Resistant Pathogens. Front. Microbiol. 2016, 7, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharifi, A.; Mohammadzadeh, A.; Salehi, T.Z.; Mahmoodi, P.; Nourian, A. Cuminum Cyminum L. Essential Oil: A Promising Antibacterial and Antivirulence Agent Against Multidrug-Resistant Staphylococcus aureus. Front. Microbiol. 2021, 12, 667833. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Kim, M.-C.; Kang, O.-H.; Kwon, D.-Y. The Mechanism of Bisdemethoxycurcumin Enhances Conventional Antibiotics against Methicillin-Resistant Staphylococcus aureus. Int. J. Mol. Sci. 2020, 21, E7945. [Google Scholar] [CrossRef]
- Nair, A.; Mallya, R.; Suvarna, V.; Khan, T.A.; Momin, M.; Omri, A. Nanoparticles-Attractive Carriers of Antimicrobial Essential Oils. Antibiotics 2022, 11, 108. [Google Scholar] [CrossRef] [PubMed]
- Jamil, B.; Abbasi, R.; Abbasi, S.; Imran, M.; Khan, S.U.; Ihsan, A.; Javed, S.; Bokhari, H.; Imran, M. Encapsulation of Cardamom Essential Oil in Chitosan Nano-Composites: In-Vitro Efficacy on Antibiotic-Resistant Bacterial Pathogens and Cytotoxicity Studies. Front. Microbiol. 2016, 7, 1580. [Google Scholar] [CrossRef]
- Qiu, Y.; Wu, Y.; Lu, B.; Zhu, G.; Gong, T.; Wang, R.; Peng, Q.; Li, Y. Inhibition of Methicillin-Resistant Staphylococcus aureus (MRSA) Biofilm by Cationic Poly (D, L-Lactide-Co-Glycolide) Nanoparticles. Biofouling 2020, 36, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ding, Y.; Fan, B.; Wang, Y.; Mao, Z.; Wang, W.; Wu, J. Inflammation-Targeting Polymeric Nanoparticles Deliver Sparfloxacin and Tacrolimus for Combating Acute Lung Sepsis. J. Control Release 2020, 321, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Thiyagarajan, D.; Das, G.; Ramesh, A. Amphiphilic Cargo-Loaded Nanocarrier Enhances Antibiotic Uptake and Perturbs Efflux: Effective Synergy for Mitigation of Methicillin-Resistant Staphylococcus aureus. ChemMedChem 2017, 12, 1125–1132. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y.; Mohamed, M.F.; Seleem, M.N.; Yeo, Y. Particle Engineering for Intracellular Delivery of Vancomycin to Methicillin-Resistant Staphylococcus aureus (MRSA)-Infected Macrophages. J. Control Release 2017, 267, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.; Moreira, M.L.A.; Costa, I.F.d.J.B.; de Sousa, V.P.; Rodrigues, C.R.; da Rocha E Lima, L.M.T.; Sisnande, T.; do Carmo, F.A.; Leal, I.C.R.; Dos Santos, K.R.N.; et al. Vancomycin-Loaded Nanoparticles against Vancomycin Intermediate and Methicillin Resistant Staphylococcus aureus Strains. Nanotechnology 2020, 31, 375101. [Google Scholar] [CrossRef]
- Haddadzadegan, S.; Dorkoosh, F.; Bernkop-Schnürch, A. Oral Delivery of Therapeutic Peptides and Proteins: Technology Landscape of Lipid-Based Nanocarriers. Adv. Drug Deliv. Rev. 2022, 182, 114097. [Google Scholar] [CrossRef]
- Kalhapure, R.S.; Sikwal, D.R.; Rambharose, S.; Mocktar, C.; Singh, S.; Bester, L.; Oh, J.K.; Renukuntla, J.; Govender, T. Enhancing Targeted Antibiotic Therapy via PH Responsive Solid Lipid Nanoparticles from an Acid Cleavable Lipid. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 2067–2077. [Google Scholar] [CrossRef]
- Mhule, D.; Kalhapure, R.S.; Jadhav, M.; Omolo, C.A.; Rambharose, S.; Mocktar, C.; Singh, S.; Waddad, A.Y.; Ndesendo, V.M.K.; Govender, T. Synthesis of an Oleic Acid Based PH-Responsive Lipid and Its Application in Nanodelivery of Vancomycin. Int. J. Pharm. 2018, 550, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Hassan, D.; Omolo, C.A.; Fasiku, V.O.; Mocktar, C.; Govender, T. Novel chitosan-based pH-responsive lipid-polymer hybrid nanovesicles (OLA-LPHVs) for delivery of vancomycin against methicillin-resistant Staphylococcus aureus infections. Int. J. Biol. Macromol. 2020, 147, 385–398. [Google Scholar] [CrossRef]
- Maji, R.; Omolo, C.A.; Agrawal, N.; Maduray, K.; Hassan, D.; Mokhtar, C.; Mackhraj, I.; Govender, T. PH-Responsive Lipid–Dendrimer Hybrid Nanoparticles: An Approach to Target and Eliminate Intracellular Pathogens. Mol. Pharm. 2019, 16, 4594–4609. [Google Scholar] [CrossRef]
- Dubrac, S.; Boneca, I.G.; Poupel, O.; Msadek, T. New Insights into the WalK/WalR (YycG/YycF) Essential Signal Transduction Pathway Reveal a Major Role in Controlling Cell Wall Metabolism and Biofilm Formation in Staphylococcus aureus. J. Bacteriol. 2007, 189, 8257–8269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hibbitts, A.; Lucía, A.; Serrano-Sevilla, I.; De Matteis, L.; McArthur, M.; de la Fuente, J.M.; Aínsa, J.A.; Navarro, F. Co-Delivery of Free Vancomycin and Transcription Factor Decoy-Nanostructured Lipid Carriers Can Enhance Inhibition of Methicillin Resistant Staphylococcus aureus (MRSA). PLoS ONE 2019, 14, e0220684. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yang, R.; Yuan, B.; Liu, Y.; Liu, C. The Antiviral and Antimicrobial Activities of Licorice, a Widely-Used Chinese Herb. Acta Pharm. Sin. B 2015, 5, 310–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaglal, Y.; Osman, N.; Omolo, C.A.; Mocktar, C.; Devnarain, N.; Govender, T. Formulation of PH-Responsive Lipid-Polymer Hybrid Nanoparticles for Co-Delivery and Enhancement of the Antibacterial Activity of Vancomycin and 18β-Glycyrrhetinic Acid. J. Drug Deliv. Sci. Technol. 2021, 64, 102607. [Google Scholar] [CrossRef]
- Giraudo, A.T.; Mansilla, C.; Chan, A.; Raspanti, C.; Nagel, R. Studies on the Expression of Regulatory Locus Sae in Staphylococcus aureus. Curr. Microbiol. 2003, 46, 246–250. [Google Scholar] [CrossRef] [PubMed]
- Long, D.R.; Mead, J.; Hendricks, J.M.; Hardy, M.E.; Voyich, J.M. 18β-Glycyrrhetinic Acid Inhibits Methicillin-Resistant Staphylococcus aureus Survival and Attenuates Virulence Gene Expression. Antimicrob. Agents Chemother. 2013, 57, 241–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, M.; Abu Lila, A.S.; Shimizu, T.; Alaaeldin, E.; Hussein, A.; Sarhan, H.A.; Szebeni, J.; Ishida, T. PEGylated Liposomes: Immunological Responses. Sci. Technol. Adv. Mater. 2019, 20, 710–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szebeni, J. Complement Activation-Related Pseudoallergy: A Stress Reaction in Blood Triggered by Nanomedicines and Biologicals. Mol. Immunol. 2014, 61, 163–173. [Google Scholar] [CrossRef]
- Szebeni, J.; Jiskoot, W. Immunological Issues with Nanomedicines: Immunogenicity, Hypersensitivity, Accelerated Clearance and Immune Suppression. In Handbook of Nanobiomedical Research: Fundamentals, Applications and Recent Developments; World Scientific Publishing Company: New Jersey, NJ, USA, 2013; Volume 3, pp. 45–73. ISBN 978-981-4520-64-5. [Google Scholar]
- Patkó, Z.; Szebeni, J. Blood Cell Changes in Complement Activation-Related Pseudoallergy. Eur. J. Nanomed. 2015, 7, 233–244. [Google Scholar] [CrossRef]
- Lila, A.S.A.; Uehara, Y.; Ishida, T.; Kiwada, H. Application of Polyglycerol Coating to Plasmid DNA Lipoplex for the Evasion of the Accelerated Blood Clearance Phenomenon in Nucleic Acid Delivery. J. Pharm. Sci. 2014, 103, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Hong, L.; Wang, Z.; Wei, X.; Shi, J.; Li, C. Antibodies against Polyethylene Glycol in Human Blood: A Literature Review. J. Pharmacol. Toxicol. Methods 2020, 102, 106678. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Liang, C.; Song, X.; Chen, Q.; Jin, Q.; Wang, C.; Liu, Z. Erythrocyte-Membrane-Enveloped Perfluorocarbon as Nanoscale Artificial Red Blood Cells to Relieve Tumor Hypoxia and Enhance Cancer Radiotherapy. Adv. Mater. 2017, 29, 1701429. [Google Scholar] [CrossRef] [PubMed]
- Xuan, M.; Shao, J.; Zhao, J.; Li, Q.; Dai, L.; Li, J. Magnetic Mesoporous Silica Nanoparticles Cloaked by Red Blood Cell Membranes: Applications in Cancer Therapy. Angew. Chem. Int. Ed. 2018, 57, 6049–6053. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.-M.J.; Zhang, L.; Aryal, S.; Cheung, C.; Fang, R.H.; Zhang, L. Erythrocyte Membrane-Camouflaged Polymeric Nanoparticles as a Biomimetic Delivery Platform. Proc. Natl. Acad. Sci. USA 2011, 108, 10980–10985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
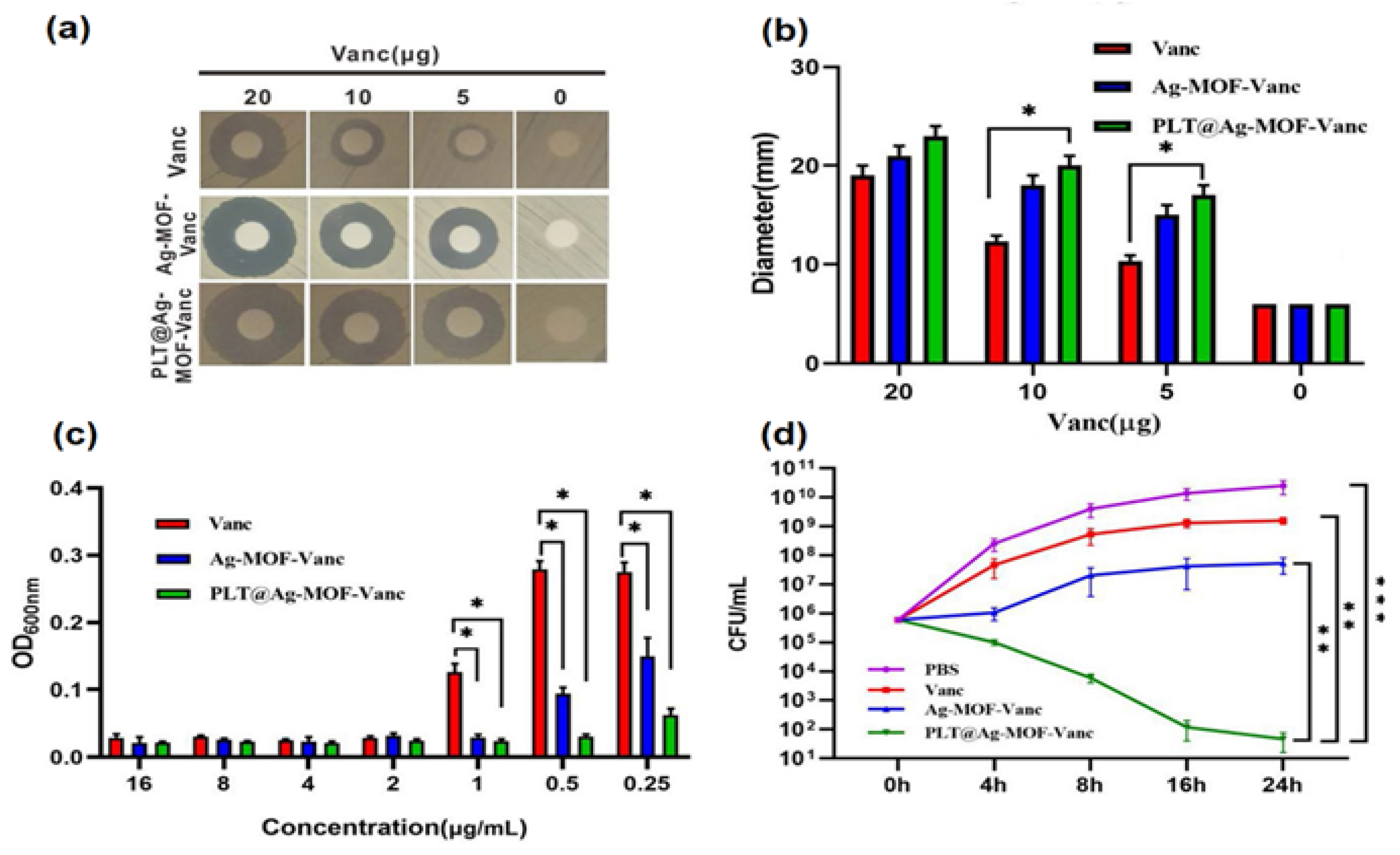
- Huang, R.; Cai, G.-Q.; Li, J.; Li, X.-S.; Liu, H.-T.; Shang, X.-L.; Zhou, J.-D.; Nie, X.-M.; Gui, R. Platelet Membrane-Camouflaged Silver Metal-Organic Framework Drug System against Infections Caused by Methicillin-Resistant Staphylococcus aureus. J. Nanobiotechnology 2021, 19, 229. [Google Scholar] [CrossRef] [PubMed]
- Olsson, M.; Oldenborg, P.-A. CD47 on Experimentally Senescent Murine RBCs Inhibits Phagocytosis Following Fcγ Receptor–Mediated but Not Scavenger Receptor–Mediated Recognition by Macrophages. Blood 2008, 112, 4259–4267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalman, L.S.; Wood, L.M.; Müller-Eberhard, H.J. Isolation of a Human Erythrocyte Membrane Protein Capable of Inhibiting Expression of Homologous Complement Transmembrane Channels. Proc. Natl. Acad. Sci. USA 1986, 83, 6975–6979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson-Weller, A.; Burge, J.; Austen, K.F. Purification from Guinea Pig Erythrocyte Stroma of a Decay-Accelerating Factor for the Classical C3 Convertase, C4b,2a. J. Immunol. 1981, 127, 2035–2039. [Google Scholar]
- Freysdottir, J.; Sigfusson, A. A Flow Cytometric Assay for Measuring Complement Receptor 1 (CR1) and the Complement Fragments C3d and C4d on Erythrocytes. J. Immunol. Methods 1991, 142, 45–52. [Google Scholar] [CrossRef]
- Zhang, X.; Angsantikul, P.; Ying, M.; Zhuang, J.; Zhang, Q.; Wei, X.; Jiang, Y.; Zhang, Y.; Dehaini, D.; Chen, M.; et al. Remote Loading of Small Molecule Therapeutics into Cholesterol-Enriched Cell Membrane-Derived Vesicles. Angew. Chem. Int. Ed. Engl. 2017, 56, 14075–14079. [Google Scholar] [CrossRef] [PubMed]
- Peraro, M.D.; van der Goot, F.G. Pore-Forming Toxins: Ancient, but Never Really out of Fashion. Nat. Rev. Microbiol. 2016, 14, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Greaney, A.J.; Leppla, S.H.; Moayeri, M. Bacterial Exotoxins and the Inflammasome. Front. Immunol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, J.; Chen, W.; Angsantikul, P.; Spiekermann, K.A.; Fang, R.H.; Gao, W.; Zhang, L. Erythrocyte Membrane-Coated Nanogel for Combinatorial Antivirulence and Responsive Antimicrobial Delivery against Staphylococcus aureus Infection. J. Control Release 2017, 263, 185–191. [Google Scholar] [CrossRef]
- Xue, B.; Qu, Y.; Shi, K.; Zhou, K.; He, X.; Chu, B.; Qian, Z. Advances in the Application of Injectable Thermosensitive Hydrogel Systems for Cancer Therapy. J. Biomed. Nanotechnol. 2020, 16, 1427–1453. [Google Scholar] [CrossRef]
- Zou, S.; He, Q.; Wang, Q.; Wang, B.; Liu, G.; Zhang, F.; Cheng, X.; Wang, B.; Zhang, L. Injectable Nanosponge-Loaded Pluronic F127 Hydrogel for Pore-Forming Toxin Neutralization. Int. J. Nanomed. 2021, 16, 4239–4250. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Bai, Y.; Li, Z.; Wang, F.; Fan, X.; Zhou, X. Bacterial Extracellular Vesicle-Coated Multi-Antigenic Nanovaccines Protect against Drug-Resistant Staphylococcus aureus Infection by Modulating Antigen Processing and Presentation Pathways. Theranostics 2020, 10, 7131–7149. [Google Scholar] [CrossRef]
- Gao, F.; Xu, L.; Yang, B.; Fan, F.; Yang, L. Kill the Real with the Fake: Eliminate Intracellular Staphylococcus aureus Using Nanoparticle Coated with Its Extracellular Vesicle Membrane as Active-Targeting Drug Carrier. ACS Infect. Dis. 2019, 5, 218–227. [Google Scholar] [CrossRef]
- Schrank, C.L.; Minbiole, K.P.C.; Wuest, W.M. Are Quaternary Ammonium Compounds, the Workhorse Disinfectants, Effective against Severe Acute Respiratory Syndrome-Coronavirus-2? ACS Infect. Dis. 2020, 6, 1553–1557. [Google Scholar] [CrossRef] [PubMed]
- Sabry, W.; Al-Dakhakhni, M.Y.S.; Al-hoseiny, I.M.; Abdel Tawab, R.A.R. Detection of Biocides Resistance Genes among Clinical Isolates of Both Methicillin-Resistant and Methicillin-Sensitive Strains of Staphylococcus aureus. Int. J. Med. Arts 2021, 3, 1602–1610. [Google Scholar] [CrossRef]
- Chen, B.; Han, J.; Dai, H.; Jia, P. Biocide-Tolerance and Antibiotic-Resistance in Community Environments and Risk of Direct Transfers to Humans: Unintended Consequences of Community-Wide Surface Disinfecting during COVID-19? Environ. Pollut. 2021, 283, 117074. [Google Scholar] [CrossRef]
- Merchel Piovesan Pereira, B.; Wang, X.; Tagkopoulos, I. Biocide-Induced Emergence of Antibiotic Resistance in Escherichia Coli. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Buzón-Durán, L.; Alonso-Calleja, C.; Riesco-Peláez, F.; Capita, R. Effect of Sub-Inhibitory Concentrations of Biocides on the Architecture and Viability of MRSA Biofilms. Food Microbiol. 2017, 65, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Speck, S.; Wenke, C.; Feßler, A.T.; Kacza, J.; Geber, F.; Scholtzek, A.D.; Hanke, D.; Eichhorn, I.; Schwarz, S.; Rosolowski, M.; et al. Borderline Resistance to Oxacillin in Staphylococcus aureus after Treatment with Sub-Lethal Sodium Hypochlorite Concentrations. Heliyon 2020, 6, e04070. [Google Scholar] [CrossRef] [PubMed]
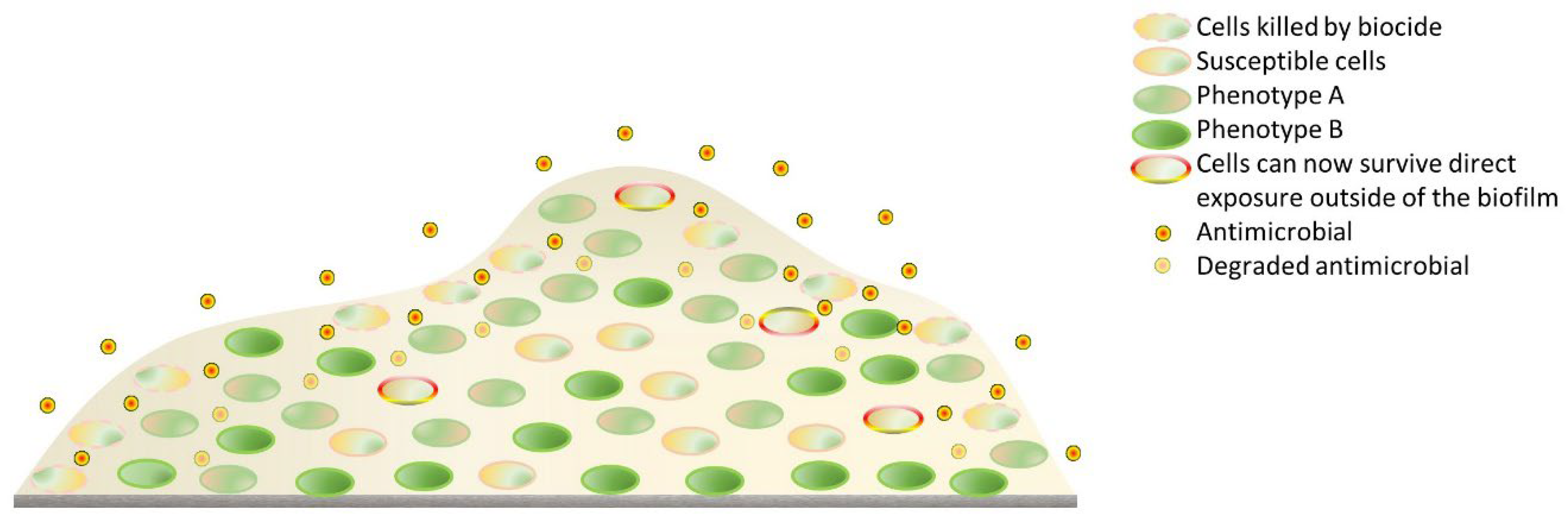
- Flemming, H.-C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An Emergent Form of Bacterial Life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Oyama, T.; Miyazaki, M.; Yoshimura, M.; Takata, T.; Ohjimi, H.; Jimi, S. Biofilm-Forming Methicillin-Resistant Staphylococcus aureus Survive in Kupffer Cells and Exhibit High Virulence in Mice. Toxins 2016, 8, 198. [Google Scholar] [CrossRef] [Green Version]
- Mosselhy, D.A.; Assad, M.; Sironen, T.; Elbahri, M. Nanotheranostics: A Possible Solution for Drug-Resistant Staphylococcus aureus and Their Biofilms? Nanomaterials 2021, 11, 82. [Google Scholar] [CrossRef] [PubMed]
- Ozcelik, B.; Pasic, P.; Sangwan, P.; Be, C.L.; Glattauer, V.; Thissen, H.; Boulos, R.A. Evaluation of the Novel Antimicrobial BCP3 in a Coating for Endotracheal Tubes. ACS Omega 2020, 5, 10288–10296. [Google Scholar] [CrossRef] [PubMed]
- Ledwoch, K.; Dancer, S.J.; Otter, J.A.; Kerr, K.; Roposte, D.; Rushton, L.; Weiser, R.; Mahenthiralingam, E.; Muir, D.D.; Maillard, J.-Y. Beware Biofilm! Dry Biofilms Containing Bacterial Pathogens on Multiple Healthcare Surfaces; a Multi-Centre Study. J. Hosp. Infect. 2018, 100, e47–e56. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcal Infections: Mechanisms of Biofilm Maturation and Detachment as Critical Determinants of Pathogenicity. Annu. Rev. Med. 2013, 64, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.; Allan, R.N.; Howlin, R.P.; Stoodley, P.; Hall-Stoodley, L. Targeting microbial biofilms: Current and prospective therapeutic strategies. Nat. Rev. Microbiol. 2017, 15, 740–755. [Google Scholar] [CrossRef] [PubMed]
- Hickok, N.J.; Shapiro, I.M. Immobilized Antibiotics to Prevent Orthopaedic Implant Infections. Adv. Drug Deliv. Rev. 2012, 64, 1165–1176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, E.P.; Truong, V.K.; Wang, J.Y.; Berndt, C.C.; Jones, R.T.; Yusuf, I.I.; Peake, I.; Schmidt, H.W.; Fluke, C.; Barnes, D.; et al. Impact of Nanoscale Roughness of Titanium Thin Film Surfaces on Bacterial Retention. Langmuir 2010, 26, 1973–1982. [Google Scholar] [CrossRef] [PubMed]
- Badica, P.; Batalu, N.D.; Burdusel, M.; Grigoroscuta, M.A.; Aldica, G.; Enculescu, M.; Gradisteanu Pircalabioru, G.; Popa, M.; Marutescu, L.G.; Dumitriu, B.G.; et al. Antibacterial Composite Coatings of MgB2 Powders Embedded in PVP Matrix. Sci. Rep. 2021, 11, 9591. [Google Scholar] [CrossRef]
- Kazemzadeh-Narbat, M.; Lai, B.F.L.; Ding, C.; Kizhakkedathu, J.N.; Hancock, R.E.W.; Wang, R. Multilayered Coating on Titanium for Controlled Release of Antimicrobial Peptides for the Prevention of Implant-Associated Infections. Biomaterials 2013, 34, 5969–5977. [Google Scholar] [CrossRef] [PubMed]
- Passos, T.F.; Souza, M.T.; Zanotto, E.D.; de Souza, C.W.O. Bactericidal Activity and Biofilm Inhibition of F18 Bioactive Glass against Staphylococcus aureus. Mater. Sci. Eng. C 2021, 118, 111475. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, V.d.C.; Souza, M.T.; Zanotto, E.D.; Watanabe, E.; Coraça-Huber, D. Biofilm Formation and Expression of Virulence Genes of Microorganisms Grown in Contact with a New Bioactive Glass. Pathogens 2020, 9, 927. [Google Scholar] [CrossRef] [PubMed]
- David, N.; Nallaiyan, R. Biologically Anchored Chitosan/Gelatin-SrHAP Scaffold Fabricated on Titanium against Chronic Osteomyelitis Infection. Int. J. Biol. Macromol. 2018, 110, 206–214. [Google Scholar] [CrossRef]
- Bala Subramaniyan, S.; Megarajan, S.; Vijayakumar, S.; Mariappan, M.; Anbazhagan, V. Evaluation of the Toxicities of Silver and Silver Sulfide Nanoparticles against Gram-Positive and Gram-Negative Bacteria. IET Nanobiotechnol. 2019, 13, 326–331. [Google Scholar] [CrossRef]
- Gao, C.; Cheng, H.; Xu, N.; Li, Y.; Chen, Y.; Wei, Y.; Gao, B.; Fu, J.; Huo, K.; Xiong, W. Poly(Dopamine) and Ag Nanoparticle-Loaded TiO2 Nanotubes with Optimized Antibacterial and ROS-Scavenging Bioactivities. Nanomedicine 2019, 14, 803–818. [Google Scholar] [CrossRef] [PubMed]
- Guan, M.; Chen, Y.; Wei, Y.; Song, H.; Gao, C.; Cheng, H.; Li, Y.; Huo, K.; Fu, J.; Xiong, W. Long-Lasting Bactericidal Activity through Selective Physical Puncture and Controlled Ions Release of Polydopamine and Silver Nanoparticles-Loaded TiO2 Nanorods in Vitro and in Vivo. Int. J. Nanomed. 2019, 14, 2903–2914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Wang, Y.; Hua, J.; Lyu, B. [Study on antibacterial property of silver loaded titanium dioxide antibacterial coated endotracheal intubation tube]. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2014, 26, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Kollef, M.H.; Afessa, B.; Anzueto, A.; Veremakis, C.; Kerr, K.M.; Margolis, B.D.; Craven, D.E.; Roberts, P.R.; Arroliga, A.C.; Hubmayr, R.D.; et al. Silver-Coated Endotracheal Tubes and Incidence of Ventilator-Associated Pneumonia: The NASCENT Randomized Trial. JAMA 2008, 300, 805–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berra, L.; Curto, F.; Li, B.G.; Laquerriere, P.; Pitts, B.; Baccarelli, A.; Kolobow, T. Antimicrobial-coated endotracheal tubes: An experimental study. Intensive Care Med. 2008, 34, 1020–1029. [Google Scholar] [CrossRef] [PubMed]
- Marassi, V.; Di Cristo, L.; Smith, S.G.J.; Ortelli, S.; Blosi, M.; Costa, A.L.; Reschiglian, P.; Volkov, Y.; Prina-Mello, A. Silver Nanoparticles as a Medical Device in Healthcare Settings: A Five-Step Approach for Candidate Screening of Coating Agents. R. Soc. Open Sci. 2018, 5, 171113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, D.; Kang, E.-T.; Neoh, K.G. Antimicrobial Copper-Based Materials and Coatings: Potential Multifaceted Biomedical Applications. ACS Appl. Mater. Interfaces 2020, 12, 21159–21182. [Google Scholar] [CrossRef]
- Jiao, J.; Zhang, S.; Qu, X.; Yue, B. Recent Advances in Research on Antibacterial Metals and Alloys as Implant Materials. Front. Cell. Infect. Microbiol. 2021, 11, 693939. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, J.; Attarilar, S.; Wang, C.; Tamaddon, M.; Yang, C.; Xie, K.; Yao, J.; Wang, L.; Liu, C.; et al. Nano-Modified Titanium Implant Materials: A Way Toward Improved Antibacterial Properties. Front. Bioeng. Biotechnol. 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.K.; Syed, P.; Lehtinen, L.; Leivo, J.; Gidwani, K.; Wittfooth, S.; Pettersson, K.; Lamminmäki, U. A Nanoparticle-Based Approach for the Detection of Extracellular Vesicles. Sci. Rep. 2019, 9, 10038. [Google Scholar] [CrossRef]
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering Precision Nanoparticles for Drug Delivery. Nat. Rev. Drug Discov. 2021, 20, 101–124. [Google Scholar] [CrossRef] [PubMed]
- Abdellatif, A.A.H.; Alsowinea, A.F. Approved and Marketed Nanoparticles for Disease Targeting and Applications in COVID-19. Nanotechnol. Rev. 2021, 10, 1941–1977. [Google Scholar] [CrossRef]
- Anselmo, A.C.; Mitragotri, S. Nanoparticles in the Clinic: An Update. Bioeng. Transl. Med. 2019, 4, e10143. [Google Scholar] [CrossRef] [Green Version]
- Golčić, M.; Simetić, L.; Majnarić, T.; Golčić, G.; Herceg, D. Could Fecal Microbial Transplantation Offer a New Potential in the Treatment of Metastatic Pancreatic Ductal Adenocarcinoma? Med. Hypotheses 2022, 161, 110801. [Google Scholar] [CrossRef]
- Elsawy, S.; Elsherif, W.M.; Hamed, R. Effect of Silver Nanoparticles on Vancomycin Resistant Staphylococcus aureus Infection in Critically Ill Patients. Pathog. Glob Health 2021, 115, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.S.; de Lencastre, H.; Garau, J.; Kluytmans, J.; Malhotra-Kumar, S.; Peschel, A.; Harbarth, S. Methicillin-Resistant Staphylococcus aureus. Nat. Rev. Dis. Primers 2018, 4, 1–23. [Google Scholar] [CrossRef]
- Huang, W.; Zhang, Q.; Li, W.; Yuan, M.; Zhou, J.; Hua, L.; Chen, Y.; Ye, C.; Ma, Y. Development of Novel Nanoantibiotics Using an Outer Membrane Vesicle-Based Drug Efflux Mechanism. J. Control. Release 2020, 317, 1–22. [Google Scholar] [CrossRef]
- ADhawan, A.P. Editorial Trends and Challenges in Translation of Point-of-Care Technologies in Healthcare. IEEE J. Transl. Eng. Heal. Med. 2018, 6, 0200108. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Technique | Nanoparticulate | LOD | Detection Target | Assay Time | Ref. |
|---|---|---|---|---|---|
| SERS | Ag NPs | 10 CFU/mL | MRSA | / | [64] |
| SERS | AgNPs+ | / | MRSA | 45 min | [67] |
| Colourimetric PCR | AuNPs | 500 ng | mecA | <25 min | [70] |
| Colourimetric | AuNPs | 100 ng | mecA | <100 min | [71] |
| Colourimetric | AuNPs | 500 ng DNA | mecA | <60 min | [73] |
| Resistive pulse sensing | AuNPs | 530 copies | PVL gene | 120 min | [77] |
| Fluorescence | GO | 0.02 nM | MRSA 16S rRNA | / | [90] |
| LRET | UCNs | 0.18 nM | mec-Tar | / | [91] |
| FRET | CdTe QD | 0.5 ng/mL | Antibodies | / | [92] |
| Carrier | Cargo | MIC | MIC Free Form | Ref. |
|---|---|---|---|---|
| Liposome | Van | 0.3 mg/L | 1.25 mg/L | [119] |
| Liposome | Van | 0.48 µg/mL | 7.68 µg/mL | [124] |
| OLA-LPHVs | Van | 0.59 µg/mL | 31.25 μg/mL | [152] |
| LDH-NPs | Van | 3.90 μg/mL | 31.25 mg/mL | [154] |
| LPHNPs | Van & 18β-glycyrrhetinic acid | 0.48 μg/mL | 7.81 mg/mL | [157] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hulme, J. Application of Nanomaterials in the Prevention, Detection, and Treatment of Methicillin-Resistant Staphylococcus aureus (MRSA). Pharmaceutics 2022, 14, 805. https://doi.org/10.3390/pharmaceutics14040805
Hulme J. Application of Nanomaterials in the Prevention, Detection, and Treatment of Methicillin-Resistant Staphylococcus aureus (MRSA). Pharmaceutics. 2022; 14(4):805. https://doi.org/10.3390/pharmaceutics14040805
Chicago/Turabian StyleHulme, John. 2022. "Application of Nanomaterials in the Prevention, Detection, and Treatment of Methicillin-Resistant Staphylococcus aureus (MRSA)" Pharmaceutics 14, no. 4: 805. https://doi.org/10.3390/pharmaceutics14040805