
Sex and Circadian Timing Modulate Oxaliplatin Hematological and Hematopoietic Toxicities
 , , and
, , and
Abstract
:1. Introduction
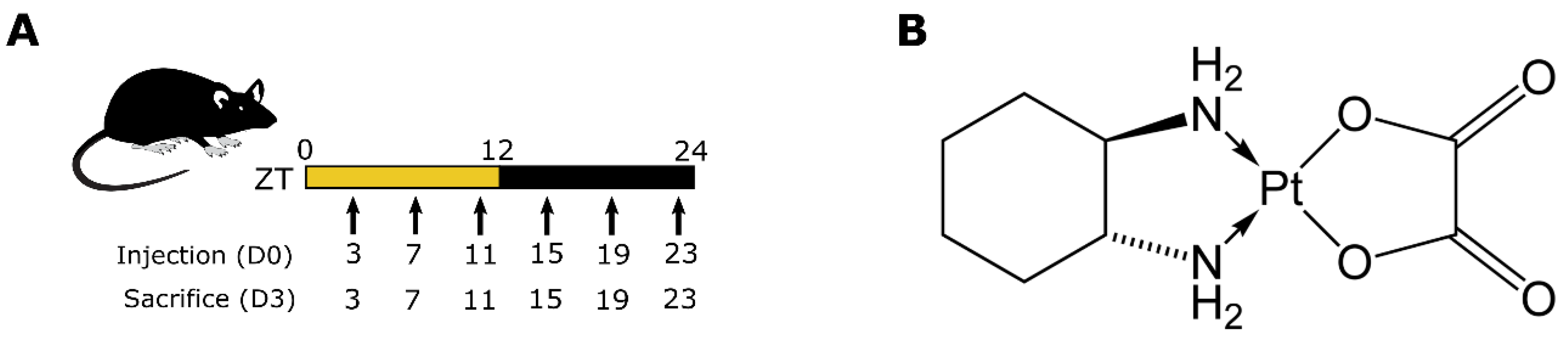
2. Materials and Methods
3. Results
3.1. Circadian Rhythms of Hematopoietic Components under Control Conditions
3.1.1. Circulating Blood Cells
3.1.2. Bone Marrow Cellularity
3.1.3. Bone Marrow Hematopoietic Progenitors
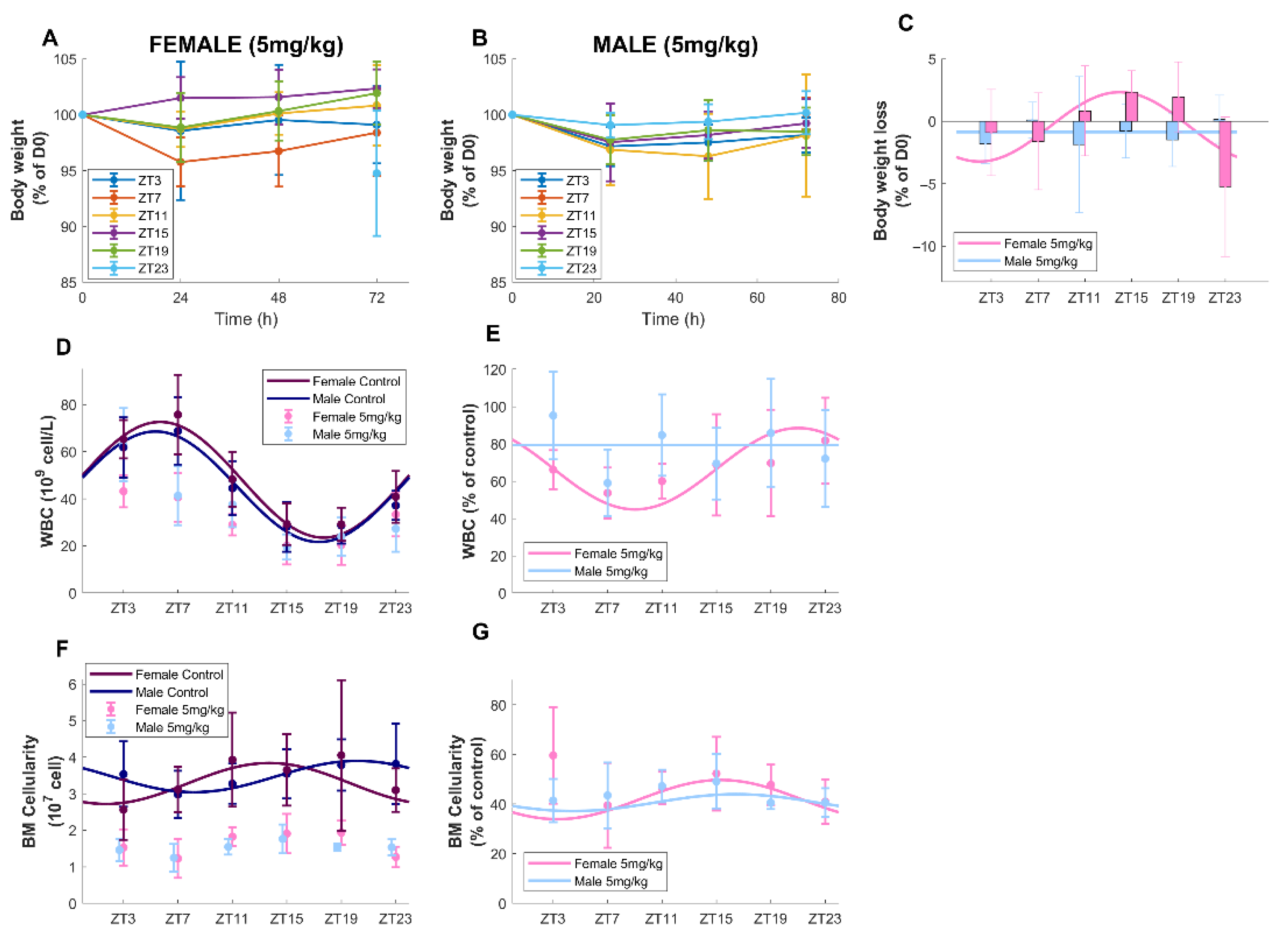
3.2. Timing-Dependent Oxaliplatin Toxicities in Male and Female Mice Treated at Equidose (5 mg/kg)
3.2.1. Survival and Body-Weight Loss
3.2.2. Circulating Blood Cell Counts
3.2.3. Bone Marrow Cellularity
3.2.4. Bone Marrow Hematopoietic Progenitors
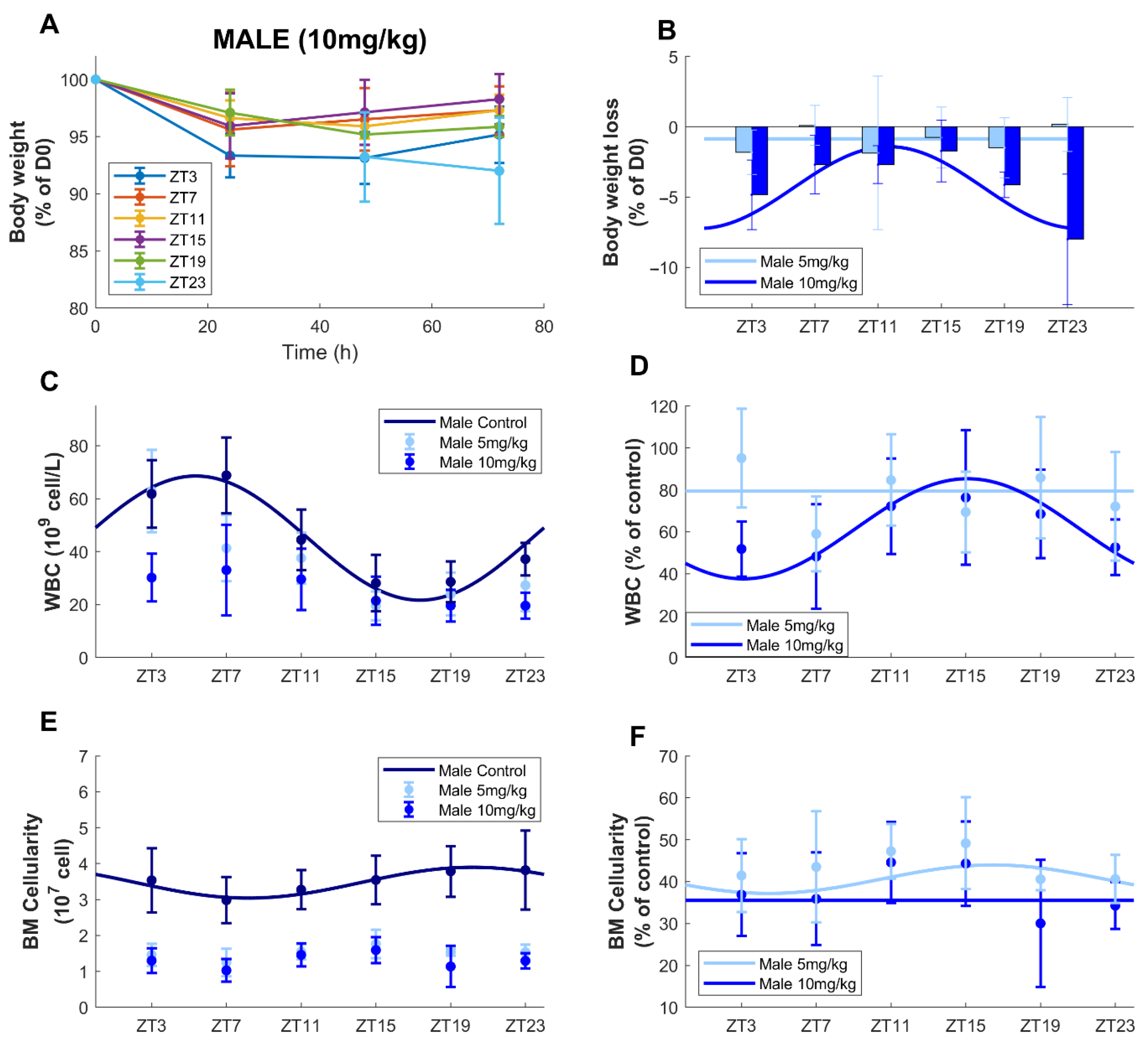
3.3. Circadian Time-Dependent Oxaliplatin Toxicities in Males upon Dose Increase
3.3.1. Survival and Body-Weight Loss
3.3.2. Circulating Blood Cell Counts
3.3.3. Bone Marrow Cellularity
3.3.4. Bone Marrow Hematopoietic Progenitors
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ballesta, A.; Innominato, P.F.; Dallmann, R.; Rand, D.A.; Levi, F.A. Systems Chronotherapeutics. Pharmacol. Rev. 2017, 69, 161–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özdemir, B.C.; Csajka, C.; Dotto, G.-P.; Wagner, A.D. Sex Differences in Efficacy and Toxicity of Systemic Treatments: An Undervalued Issue in the Era of Precision Oncology. J. Clin. Oncol. 2018, 36, 2680–2683. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.B.; McGlynn, K.A.; Devesa, S.S.; Freedman, N.D.; Anderson, W.F. Sex Disparities in Cancer Mortality and Survival. Cancer Epidemiol. Biomark. Prev. 2011, 20, 1629–1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majek, O.; Gondos, A.; Jansen, L.; Emrich, K.; Holleczek, B.; Katalinic, A.; Nennecke, A.; Eberle, A.; Brenner, H. Sex differences in colorectal cancer survival: Population-based analysis of 164,996 colorectal cancer patients in Germany. PLoS ONE 2013, 8, e68077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soldin, O.P.; Mattison, D.R. Sex Differences in Pharmacokinetics and Pharmacodynamics. Clin. Pharmacokinet. 2009, 48, 143–157. [Google Scholar] [CrossRef] [Green Version]
- Knikman, J.E.; Gelderblom, H.; Beijnen, J.H.; Cats, A.; Guchelaar, H.J.; Henricks, L.M. Individualized Dosing of Fluoropyrimidine-Based Chemotherapy to Prevent Severe Fluoropyrimidine-Related Toxicity: What Are the Options? Clin. Pharm. 2021, 109, 591–604. [Google Scholar] [CrossRef]
- Deyme, L.; Barbolosi, D.; Gattacceca, F. Population pharmacokinetics of folfirinox: A review of studies and parameters. Cancer Chemother. Pharmacol. 2019, 83, 27–42. [Google Scholar] [CrossRef]
- Dallmann, R.; Okyar, A.; Levi, F. Dosing-Time Makes the Poison: Circadian Regulation and Pharmacotherapy. Trends Mol. Med. 2016, 22, 430–445. [Google Scholar] [CrossRef] [Green Version]
- Sancar, A.; Van Gelder, R.N. Clocks, cancer, and chronochemotherapy. Science 2021, 371, eabb0738. [Google Scholar] [CrossRef]
- Barreto, J.N.; McCullough, K.B.; Ice, L.L.; Smith, J.A. Antineoplastic agents and the associated myelosuppressive effects: A review. J. Pharm. Pract. 2014, 27, 440–446. [Google Scholar] [CrossRef]
- Al-Samkari, H.; Connors, J.M. Managing the competing risks of thrombosis, bleeding, and anticoagulation in patients with malignancy. Blood Adv. 2019, 3, 3770–3779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiozawa, T.; Tadokoro, J.; Fujiki, T.; Fujino, K.; Kakihata, K.; Masatani, S.; Morita, S.; Gemma, A.; Boku, N. Risk factors for severe adverse effects and treatment-related deaths in Japanese patients treated with irinotecan-based chemotherapy: A postmarketing survey. Jpn. J. Clin. Oncol. 2013, 43, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Penninckx, B.; Van de Voorde, W.M.; Casado, A.; Reed, N.; Moulin, C.; Karrasch, M. A systemic review of toxic death in clinical oncology trials: An Achilles’ heel in safety reporting revisited. Br. J. Cancer 2012, 107, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiel, M.J.; Yilmaz, Ö.H.; Iwashita, T.; Yilmaz, O.H.; Terhorst, C.; Morrison, S.J. SLAM Family Receptors Distinguish Hematopoietic Stem and Progenitor Cells and Reveal Endothelial Niches for Stem Cells. Cell 2005, 121, 1109–1121. [Google Scholar] [CrossRef] [Green Version]
- Osawa, M.; Hanada, K.-i.; Hamada, H.; Nakauchi, H. Long-Term Lymphohematopoietic Reconstitution by a Single CD34-Low/Negative Hematopoietic Stem Cell. Science 1996, 273, 242–245. [Google Scholar] [CrossRef] [Green Version]
- Ohkura, N.; Oishi, K.; Sekine, Y.; Atsumi, G.; Ishida, N.; Matsuda, J.; Horie, S. Comparative study of circadian variation in numbers of peripheral blood cells among mouse strains: Unique feature of C3H/HeN mice. Biol. Pharm. Bull. 2007, 30, 1177–1180. [Google Scholar] [CrossRef] [Green Version]
- Stenzinger, M.; Karpova, D.; Unterrainer, C.; Harenkamp, S.; Wiercinska, E.; Hoerster, K.; Pfeffer, M.; Maronde, E.; Bonig, H. Hematopoietic-Extrinsic Cues Dictate Circadian Redistribution of Mature and Immature Hematopoietic Cells in Blood and Spleen. Cells 2019, 8, 1033. [Google Scholar] [CrossRef] [Green Version]
- Méndez-Ferrer, S.; Lucas, D.; Battista, M.; Frenette, P.S. Haematopoietic stem cell release is regulated by circadian oscillations. Nature 2008, 452, 442–447. [Google Scholar] [CrossRef]
- Bauer, M.; Goldstein, M.; Heylmann, D.; Kaina, B. Human monocytes undergo excessive apoptosis following temozolomide activating the ATM/ATR pathway while dendritic cells and macrophages are resistant. PLoS ONE 2012, 7, e39956. [Google Scholar] [CrossRef] [Green Version]
- Levi, F.; Okyar, A.; Dulong, S.; Innominato, P.F.; Clairambault, J. Circadian timing in cancer treatments. Annu. Rev. Pharm. Toxicol. 2010, 50, 377–421. [Google Scholar] [CrossRef]
- Hammel, P.; Vitellius, C.; Boisteau, É.; Wisniewski, M.; Colle, E.; Hilmi, M.; Dengremont, C.; Granier, S.; Turpin, A.; de Mestier, L.; et al. Maintenance therapies in metastatic pancreatic cancer: Present and future with a focus on PARP inhibitors. Adv. Med. Oncol. 2020, 12, 1758835920937949. [Google Scholar] [CrossRef] [PubMed]
- Levi, F.; Metzger, G.; Massari, C.; Milano, G. Oxaliplatin: Pharmacokinetics and chronopharmacological aspects. Clin Pharm. 2000, 38, 1–21. [Google Scholar] [CrossRef]
- Levi, F.; Zidani, R.; Misset, J.L. Randomised multicentre trial of chronotherapy with oxaliplatin, fluorouracil, and folinic acid in metastatic colorectal cancer. International Organization for Cancer Chronotherapy. Lancet 1997, 350, 681–686. [Google Scholar] [CrossRef]
- Díaz, R.; Aparicio, J.; Molina, J.; Palomar, L.; Giménez, A.; Ponce, J.; Segura, Á.; Gómez-Codina, J. Clinical predictors of severe toxicity in patients treated with combination chemotherapy with irinotecan and/or oxaliplatin for metastatic colorectal cancer. Med. Oncol. 2006, 23, 347–357. [Google Scholar] [CrossRef]
- Wettergren, Y.; Carlsson, G.; Odin, E.; Gustavsson, B. Pretherapeutic uracil and dihydrouracil levels of colorectal cancer patients are associated with sex and toxic side effects during adjuvant 5-fluorouracil-based chemotherapy. Cancer 2012, 118, 2935–2943. [Google Scholar] [CrossRef]
- Li, X.M.; Mohammad-Djafari, A.; Dumitru, M.; Dulong, S.; Filipski, E.; Siffroi-Fernandez, S.; Mteyrek, A.; Scaglione, F.; Guettier, C.; Delaunay, F.; et al. A circadian clock transcription model for the personalization of cancer chronotherapy. Cancer Res. 2013, 73, 7176–7188. [Google Scholar] [CrossRef] [Green Version]
- Levi, F.; Focan, C.; Karaboue, A.; de la Valette, V.; Focan-Henrard, D.; Baron, B.; Kreutz, F.; Giacchetti, S. Implications of circadian clocks for the rhythmic delivery of cancer therapeutics. Adv. Drug Deliv. Rev. 2007, 59, 1015–1035. [Google Scholar] [CrossRef]
- Giacchetti, S.; Dugue, P.A.; Innominato, P.F.; Bjarnason, G.A.; Focan, C.; Garufi, C.; Tumolo, S.; Coudert, B.; Iacobelli, S.; Smaaland, R.; et al. Sex moderates circadian chemotherapy effects on survival of patients with metastatic colorectal cancer: A meta-analysis. Ann. Oncol. 2012, 23, 3110–3116. [Google Scholar] [CrossRef]
- Innominato, P.F.; Ballesta, A.; Huang, Q.; Focan, C.; Chollet, P.; Karaboué, A.; Giacchetti, S.; Bouchahda, M.; Adam, R.; Garufi, C.; et al. Sex-dependent least toxic timing of irinotecan combined with chronomodulated chemotherapy for metastatic colorectal cancer: Randomized multicenter EORTC 05011 trial. Cancer Med. 2020, 9, 4148–4159. [Google Scholar] [CrossRef] [Green Version]
- Boughattas, N.A.; Hecquet, B.; Fournier, C.; Bruguerolle, B.; Trabelsi, H.; Bouzouita, K.; Omrane, B.; Levi, F. Comparative pharmacokinetics of oxaliplatin (L-OHP) and carboplatin (CBDCA) in mice with reference to circadian dosing time. Biopharm. Drug Dispos. 1994, 15, 761–773. [Google Scholar] [CrossRef]
- Boughattas, N.A.; Levi, F.; Fournier, C.; Lemaigre, G.; Roulon, A.; Hecquet, B.; Mathe, G.; Reinberg, A. Circadian rhythm in toxicities and tissue uptake of 1,2-diamminocyclohexane(trans-1)oxalatoplatinum(II) in mice. Cancer Res. 1989, 49, 3362–3368. [Google Scholar] [PubMed]
- Schindler, J.W.; Van Buren, D.; Foudi, A.; Krejci, O.; Qin, J.; Orkin, S.H.; Hock, H. TEL-AML1 corrupts hematopoietic stem cells to persist in the bone marrow and initiate leukemia. Cell Stem Cell 2009, 5, 43–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smaaland, R.; Sothern, R.B.; Laerum, O.D.; Abrahamsen, J.F. Rhythms in human bone marrow and blood cells. Chronobiol. Int. 2002, 19, 101–127. [Google Scholar] [CrossRef] [PubMed]
- Scheiermann, C.; Kunisaki, Y.; Lucas, D.; Chow, A.; Jang, J.E.; Zhang, D.; Hashimoto, D.; Merad, M.; Frenette, P.S. Adrenergic nerves govern circadian leukocyte recruitment to tissues. Immunity 2012, 37, 290–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, G.R.; Al-Obaidi, M.; Rower, J.; Harmon, C.; Dai, C.; Acosta, E.; Giri, S.; Zamboni, W.; Lucas, A.T.; Shachar, S.S.; et al. Does oxaliplatin pharmacokinetics (PKs) explain associations between body composition and chemotherapy toxicity risk in older adults with gastrointestinal (GI) cancers? J. Clin. Oncol. 2021, 39, 3095. [Google Scholar] [CrossRef]
- Graham, M.A.; Lockwood, G.F.; Greenslade, D.; Brienza, S.; Bayssas, M.; Gamelin, E. Clinical Pharmacokinetics of Oxaliplatin: A Critical Review. Clin. Cancer Res. 2000, 6, 1205–1218. [Google Scholar]
- He, W.; Holtkamp, S.; Hergenhan, S.M.; Kraus, K.; de Juan, A.; Weber, J.; Bradfield, P.; Grenier, J.M.P.; Pelletier, J.; Druzd, D.; et al. Circadian Expression of Migratory Factors Establishes Lineage-Specific Signatures that Guide the Homing of Leukocyte Subsets to Tissues. Immunity 2018, 49, 1175–1190.e1177. [Google Scholar] [CrossRef] [Green Version]
- Sletvold, O. Circadian rhythms of peripheral blood leukocytes in aging mice. Mech. Ageing Dev. 1987, 39, 251–261. [Google Scholar] [CrossRef]
- Weger, M.; Diotel, N.; Dorsemans, A.C.; Dickmeis, T.; Weger, B.D. Stem cells and the circadian clock. Dev. Biol. 2017, 431, 111–123. [Google Scholar] [CrossRef]
- Lucas, D.; Battista, M.; Shi, P.A.; Isola, L.; Frenette, P.S. Mobilized Hematopoietic Stem Cell Yield Depends on Species-Specific Circadian Timing. Cell Stem. Cell 2008, 3, 364–366. [Google Scholar] [CrossRef] [Green Version]
- Nakada, D.; Oguro, H.; Levi, B.P.; Ryan, N.; Kitano, A.; Saitoh, Y.; Takeichi, M.; Wendt, G.R.; Morrison, S.J. Oestrogen increases haematopoietic stem-cell self-renewal in females and during pregnancy. Nature 2014, 505, 555–558. [Google Scholar] [CrossRef]
- Foudi, A.; Hochedlinger, K.; Van Buren, D.; Schindler, J.W.; Jaenisch, R.; Carey, V.; Hock, H. Analysis of histone 2B-GFP retention reveals slowly cycling hematopoietic stem cells. Nat. Biotechnol. 2009, 27, 84–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.H.; Broxmeyer, H.E. In vitro behavior of hematopoietic progenitor cells under the influence of chemoattractants: Stromal cell-derived factor-1, steel factor, and the bone marrow environment. Blood 1998, 91, 100–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheshier, S.H.; Morrison, S.J.; Liao, X.; Weissman, I.L. In vivo proliferation and cell cycle kinetics of long-term self-renewing hematopoietic stem cells. Proc. Natl. Acad. Sci. USA 1999, 96, 3120–3125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietras, E.M.; Reynaud, D.; Kang, Y.A.; Carlin, D.; Calero-Nieto, F.J.; Leavitt, A.D.; Stuart, J.M.; Göttgens, B.; Passegué, E. Functionally Distinct Subsets of Lineage-Biased Multipotent Progenitors Control Blood Production in Normal and Regenerative Conditions. Cell Stem Cell 2015, 17, 35–46. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Female Control | Male Control | Female 5 mg/kg | Male 5 mg/kg | Male 10 mg/kg | |||
|---|---|---|---|---|---|---|---|
| WBC | Mesor (109 cell/L) | 48.07 ± 1.82 | 45.12 ± 1.86 | Mesor (% of control) | 66.65 ± 6.16 | 79.40 ± 15.59 | 61.44 ± 5.58 |
| Ampl. (% of Mesor) | 51.08 ± 5.485.79 | 51.97 ± 6.14 | Ampl. (% of Mesor) | 32.74 ± 13.42 | NS | 38.87± 13.58 | |
| Phase (ZT) | 5 h 42 ± 24 min | 5 h 21 ± 26 min | Phase (ZT) | 21 h 02 ±1 h 31 | NS | 15 h 06 ± 1 h 15 | |
| pvalues | <0.001 | <0.001 | pvalues | 0.049 | 0.75 | 0.016 | |
| BM cellularity | Mesor (107 cell/L) | 3.28 ± 0.14 | 3.47 ± 0.12 | Mesor (% of control) | 41.81 ± 1.22 | 40.58 ± 0.91 | 35.52 ± 0.88 |
| Ampl. (% of Mesor) | 17.06 ± 6.14 | 12.35 ± 4.85 | Ampl. (% of Mesor) | 18.87 ± 4.15 | 8.36 ± 3.19 | NS | |
| Phase (ZT) | 13 h 42 ± 1 h 22 | 20 h 10 ± 1 h 28 | Phase (ZT) | 15 h 19 ± 51 min | 16 h 27 ± 1 h 26 | NS | |
| pvalues | 0.027 | 0.045 | pvalues | <0.001 | 0.04 | 0.26 | |
| LK | Mesor (105 cell/L) | 3.18 ± 0.17 | 2.26 ± 0.56 | Mesor (% of control) | 29.22 ± 1.46 | 1.59 ± 0.001 | 0.44 ± 0.0001 |
| Ampl. (% of Mesor) | 38.84 ± 7.44 | NS | Ampl. (% of Mesor) | 27.55 ± 7.37 | NS | NS | |
| Phase (ZT) | 3 h 22 ± 47 min | NS | Phase (ZT) | 15 h 27 ± 59 min | NS | NS | |
| pvalues | <0.001 | 0.51 | pvalues | 0.002 | 0.89 | 0.22 | |
| LKS | Mesor (104 cell/L) | 7.56 ± 0.43 | 5.96 ± 0.19 | Mesor (% of control) | 35.07 ± 1.53 | 2.67 ± 0.008 | 1.43 ± 0.002 |
| Ampl. (% of Mesor) | 40.23 ± 8.09 | 14.35 ± 4.53 | Ampl. (% of Mesor) | 35.21 ± 6.47 | NS | NS | |
| Phase (ZT) | 3 h 20 ± 49 min | 21 h 39 ± 1 h 14 | Phase (ZT) | 14 h30 ± 40 min | NS | NS | |
| pvalues | <0.001 | 0.009 | pvalues | <0.001 | 0.27 | 0.32 | |
| MPP1 | Mesor (104 cell/L) | 4.32 ± 0.26 | 3.77 ± 0.14 | Mesor (% of control) | 24.31 ± 0.83 | 1.25 ± 0.002 | 1.02 ± 0.002 |
| Ampl. (% of Mesor) | 43.72 ± 8.68 | NS | Ampl. (% of Mesor) | 21.62 ± 4.92 | NS | NS | |
| Phase (ZT) | 3 h 15 ± 48 min | NS | Phase (ZT) | 14 h 10 ± 51 min | NS | NS | |
| pvalues | <0.001 | 0.077 | pvalues | <0.001 | 0.35 | 0.37 | |
| MPP2 | Mesor (103 cell/L) | 6.24 ± 0.68 | 4.78 ± 0.31 | Mesor (% of control) | 80.15 ± 23.51 | 7.63 ± 0.08 | 2.24 ± 0.007 |
| Ampl. (% of Mesor) | 52.17 ± 15.15 | NS | Ampl. (% of Mesor) | 229.67 ± 80.87 | NS | NS | |
| Phase (ZT) | 2 h 16 ± 1 h 09 | NS | Phase (ZT) | 13 h 52 ± 41 min | NS | NS | |
| pvalues | 0.003 | 0.12 | pvalues | <0.001 | 0.24 | 0.80 | |
| ST-HSC | Mesor (104 cell/L) | 1.22 ± 0.06 | 0.87 ± 0.041 | Mesor (% of contro) | 23.30 ± 0.82 | 1.68 ± 0.003 | 1.74 ± 0.005 |
| Ampl. (% of Mesor) | 25.95 ± 7.61 | 22.38 ± 6.89 | Ampl. (% of Mesor) | 20.59 ± 5.11 | NS | NS | |
| Phase (ZT) | 4 h 22 ± 1 h 11 | 19 h 53 ± 1 h 09 | Phase (ZT) | 14 h 27 ± 55 min | NS | NS | |
| pvalues | 0.005 | 0.007 | pvalues | <0.001 | 0.29 | 0.14 | |
| HSC-SLAM | Mesor (104 cell/L) | 1.32 ±0.11 | 0.82 ± 0.028 | Mesor (% of control) | 61.87 ± 8.23 | 7.77 ± 0.06 | 2.53 ± 0.006 |
| Ampl. (% of Mesor) | 37.89 ± 11.28 | 18.24 ± 4.93 | Ampl. (% of Mesor) | 103.38 ± 23.88 | NS | NS | |
| Phase (ZT) | 3 h 37 ± 1 h 13 | 19 h 40 ± 1 h 01 | Phase (ZT) | 14 h57 ± 41 min | NS | NS | |
| pvalues | 0.005 | 0.002 | pvalues | <0.001 | 0.062 | 0.70 | |
| LT-HSC | Mesor (103 cell/L) | 4.91± 0.25 | 3.05 ± 0.14 | Mesor (% of control) | 33.13 ± 2.10 | 1.89 ± 0.004 | 2.16 ± 0.005 |
| Ampl. (% of Mesor) | 42.08 ± 7.35 | NS | Ampl. (% of Mesor) | 45.03 ± 9.66 | 0.8 ± 0.3 | NS | |
| Phase (ZT) | 4 h 09 ± 42 min | NS | Phase (ZT) | 15 h 28 ± 45 min | 8 h 16 ± 1 h 28 | NS | |
| pvalues | <0.001 | 0.58 | pvalues | <0.001 | 0.04 | 0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dulong, S.; de Souza, L.E.B.; Machowiak, J.; Peuteman, B.; Duvallet, G.; Boyenval, D.; Roth, E.; Asgarova, A.; Chang, Y.; Li, X.-M.; et al. Sex and Circadian Timing Modulate Oxaliplatin Hematological and Hematopoietic Toxicities. Pharmaceutics 2022, 14, 2465. https://doi.org/10.3390/pharmaceutics14112465
Dulong S, de Souza LEB, Machowiak J, Peuteman B, Duvallet G, Boyenval D, Roth E, Asgarova A, Chang Y, Li X-M, et al. Sex and Circadian Timing Modulate Oxaliplatin Hematological and Hematopoietic Toxicities. Pharmaceutics. 2022; 14(11):2465. https://doi.org/10.3390/pharmaceutics14112465
Chicago/Turabian StyleDulong, Sandrine, Lucas Eduardo Botelho de Souza, Jean Machowiak, Benoit Peuteman, Gaelle Duvallet, Déborah Boyenval, Elise Roth, Afag Asgarova, Yunhua Chang, Xiao-Mei Li, and et al. 2022. "Sex and Circadian Timing Modulate Oxaliplatin Hematological and Hematopoietic Toxicities" Pharmaceutics 14, no. 11: 2465. https://doi.org/10.3390/pharmaceutics14112465