

Achyranthes aspera Extracts as Adjuvants for the Redressal of Antibiotic Resistance

 ,
,  ,
, .jpg) , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material Collection
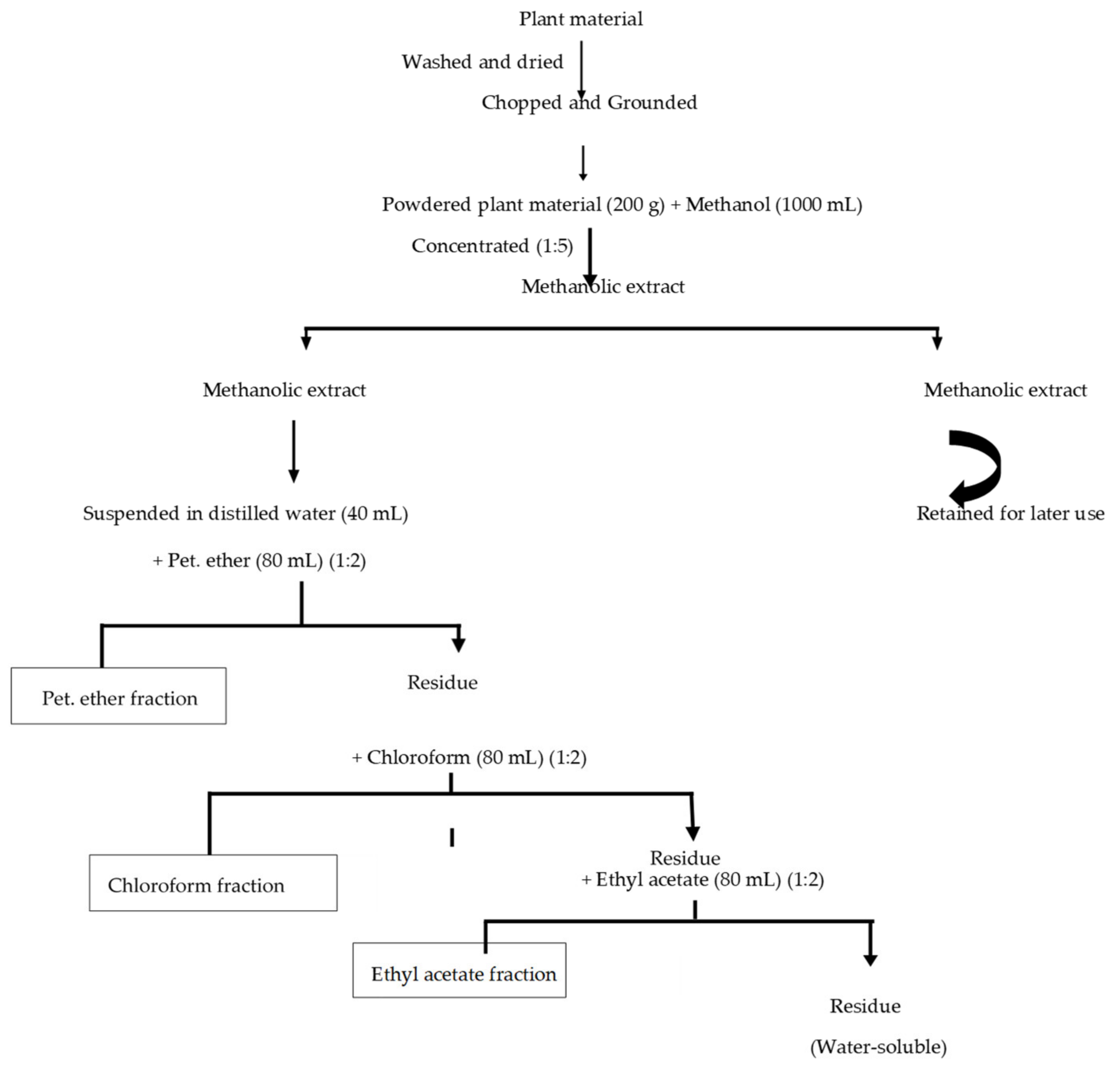
2.2. Preparation of Plant Extracts
2.2.1. Cleaning and Grinding of Plant Material
2.2.2. Crude Extract Preparation
2.2.3. Fractionation of Crude Extract
2.2.4. Stock Preparation
2.3. Phytochemical Screening
2.4. Bacterial Strains Collection
2.5. Confirmation of Resistant Bacterial Strains
2.6. Antibiotic Susceptibility Test for Plant Extracts
2.7. Antibiotic Resistance Redressal Activity
3. Results and Discussion
3.1. Phytochemical Screening
3.1.1. For Achyranthes aspera (Seed)
3.1.2. For Achyranthes aspera Leaves
3.2. Confirmation of Resistant Bacterial Strains
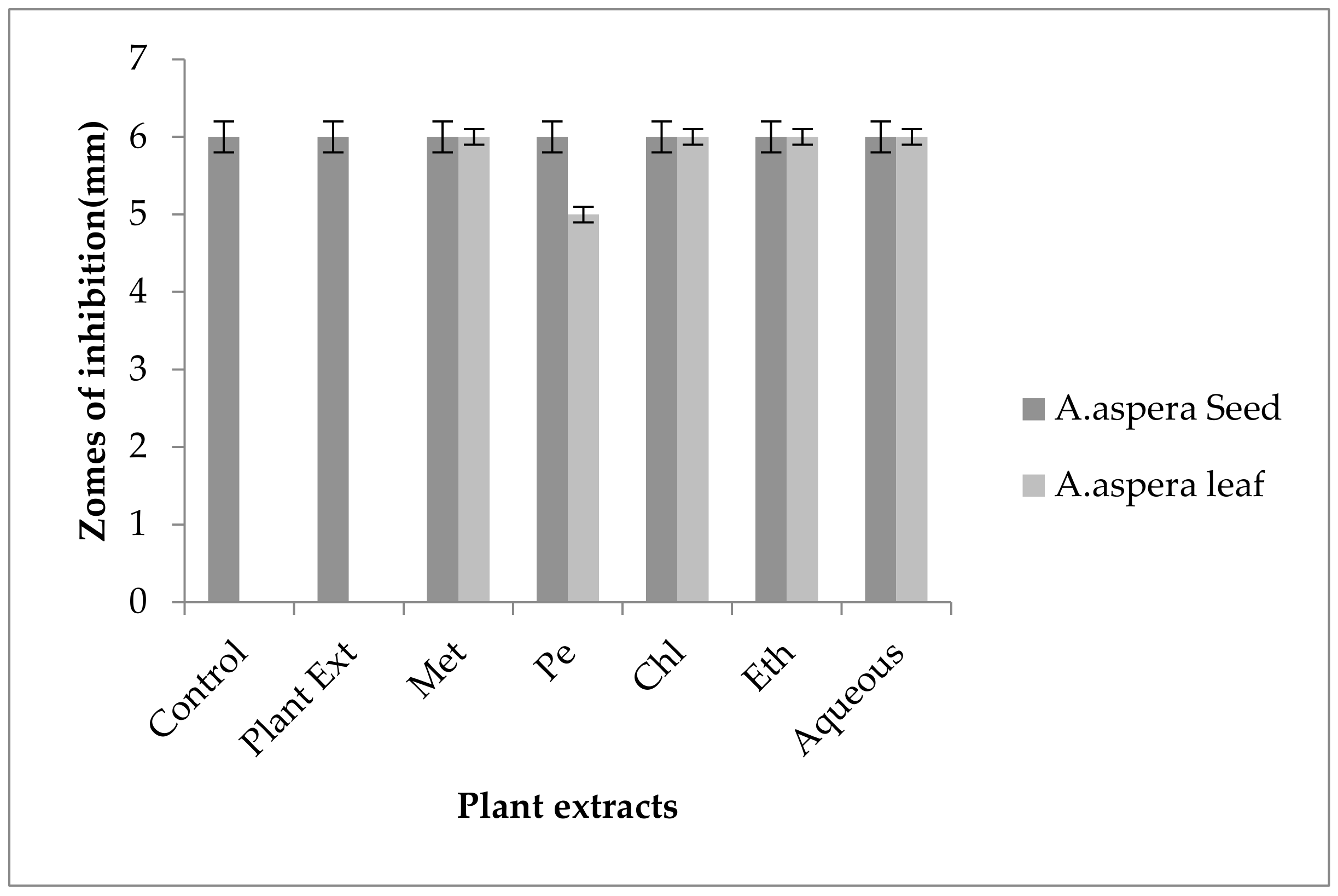
3.2.1. AST Test of Staphylococcus aureus
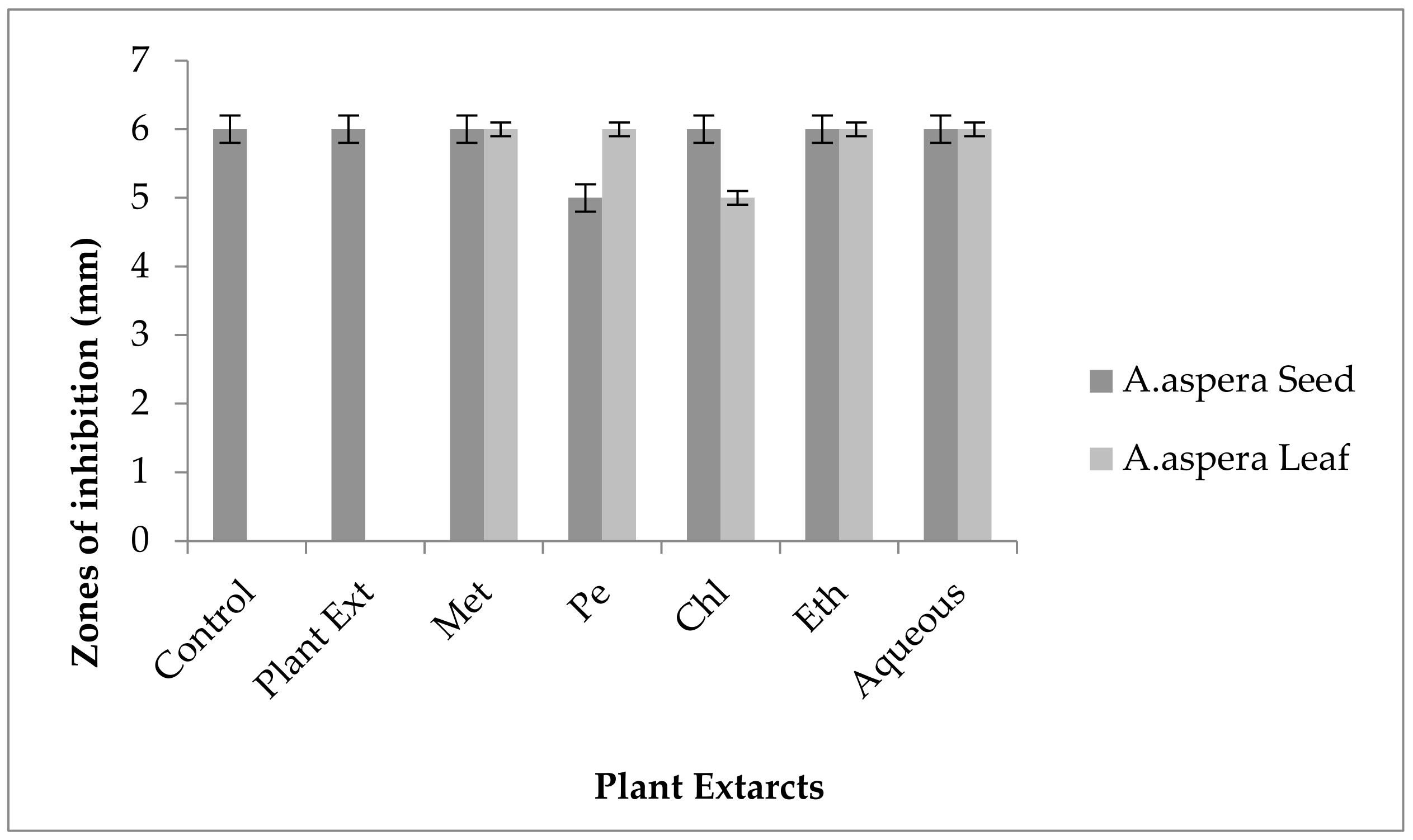
3.2.2. AST Test of Enterococcus faecalis
3.2.3. AST of Acinetobacter baumannii
3.2.4. AST of Klebsiella pneumoniae
3.2.5. AST of Pseudomonas aeruginosa
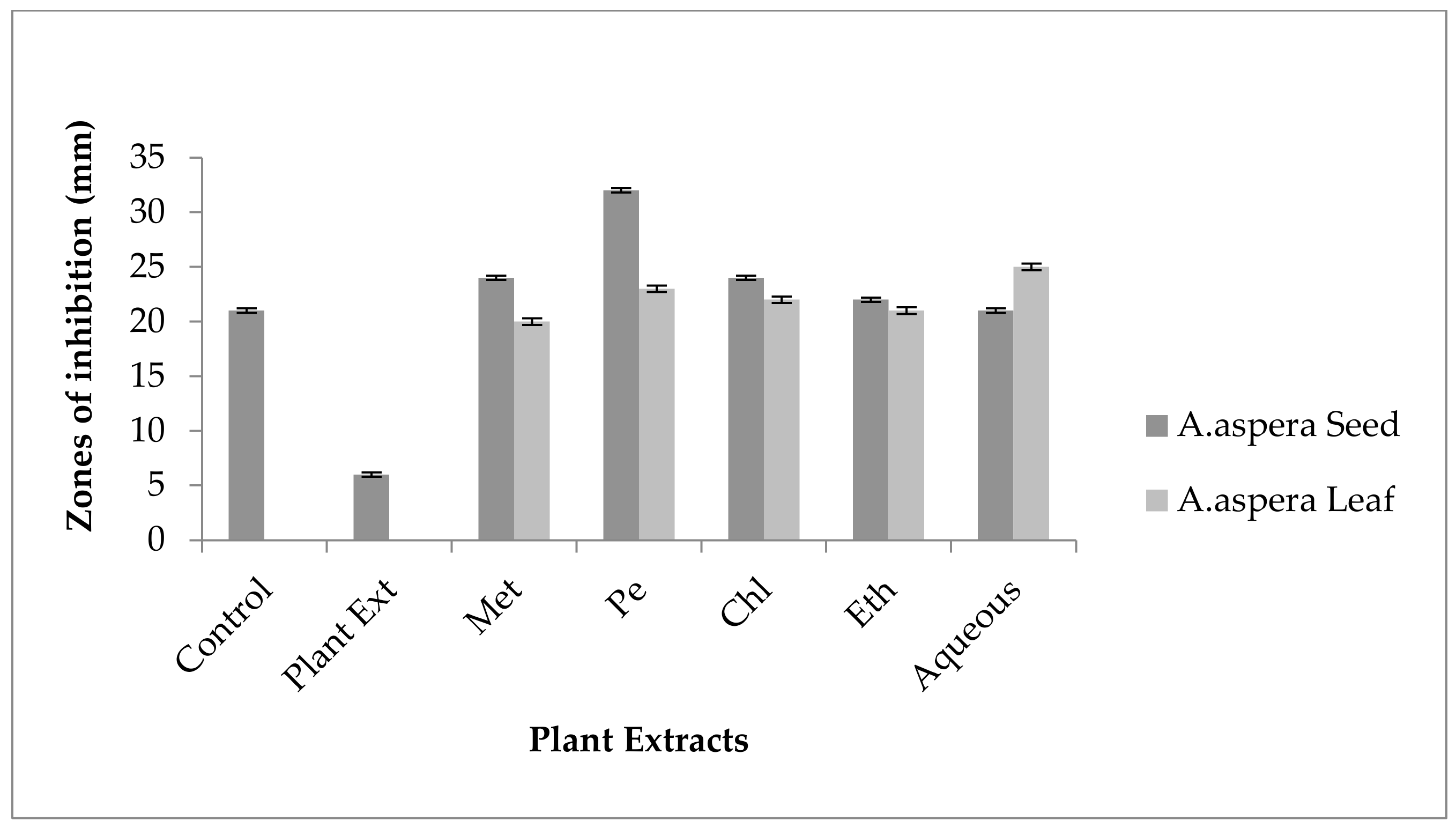
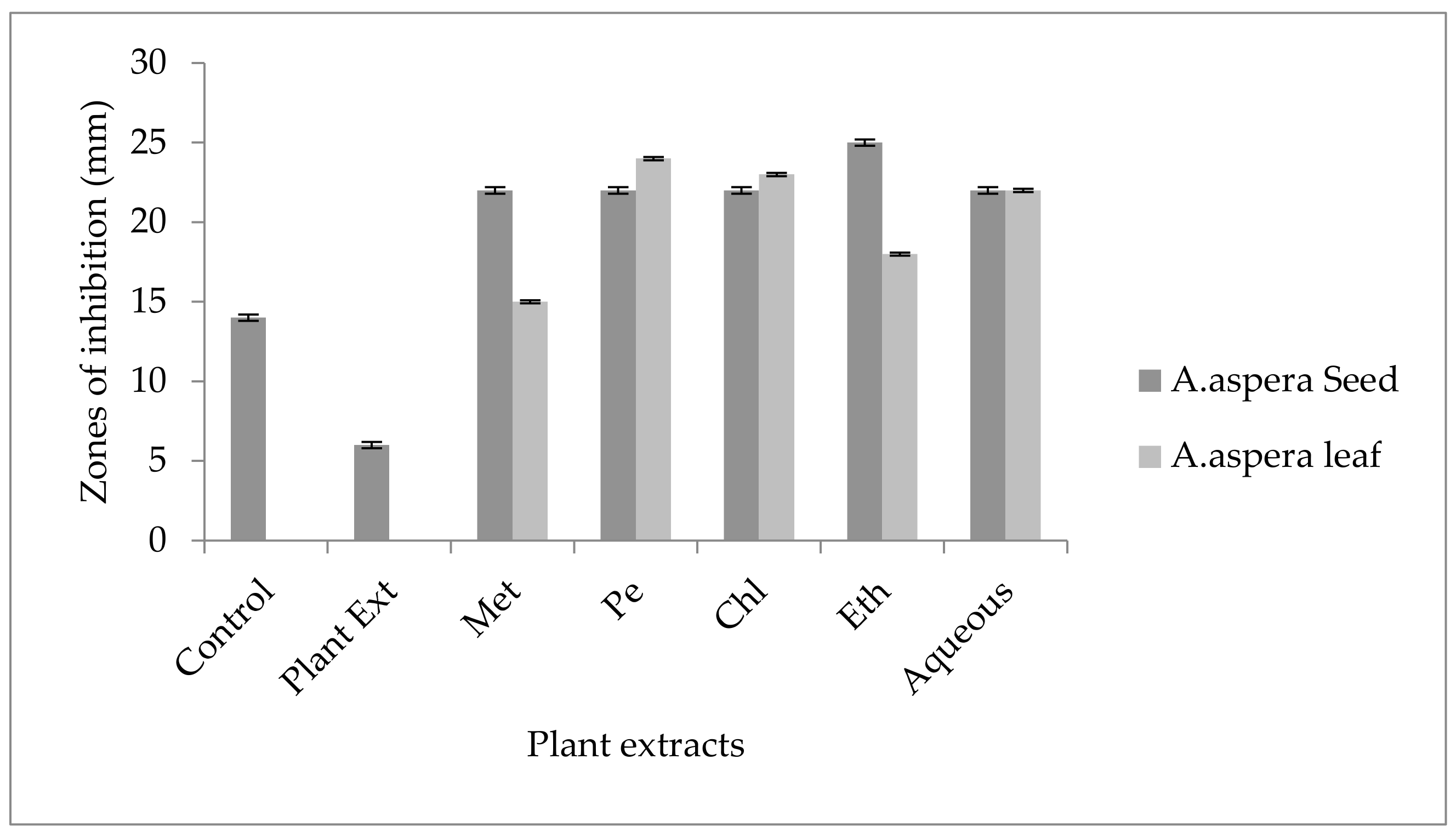
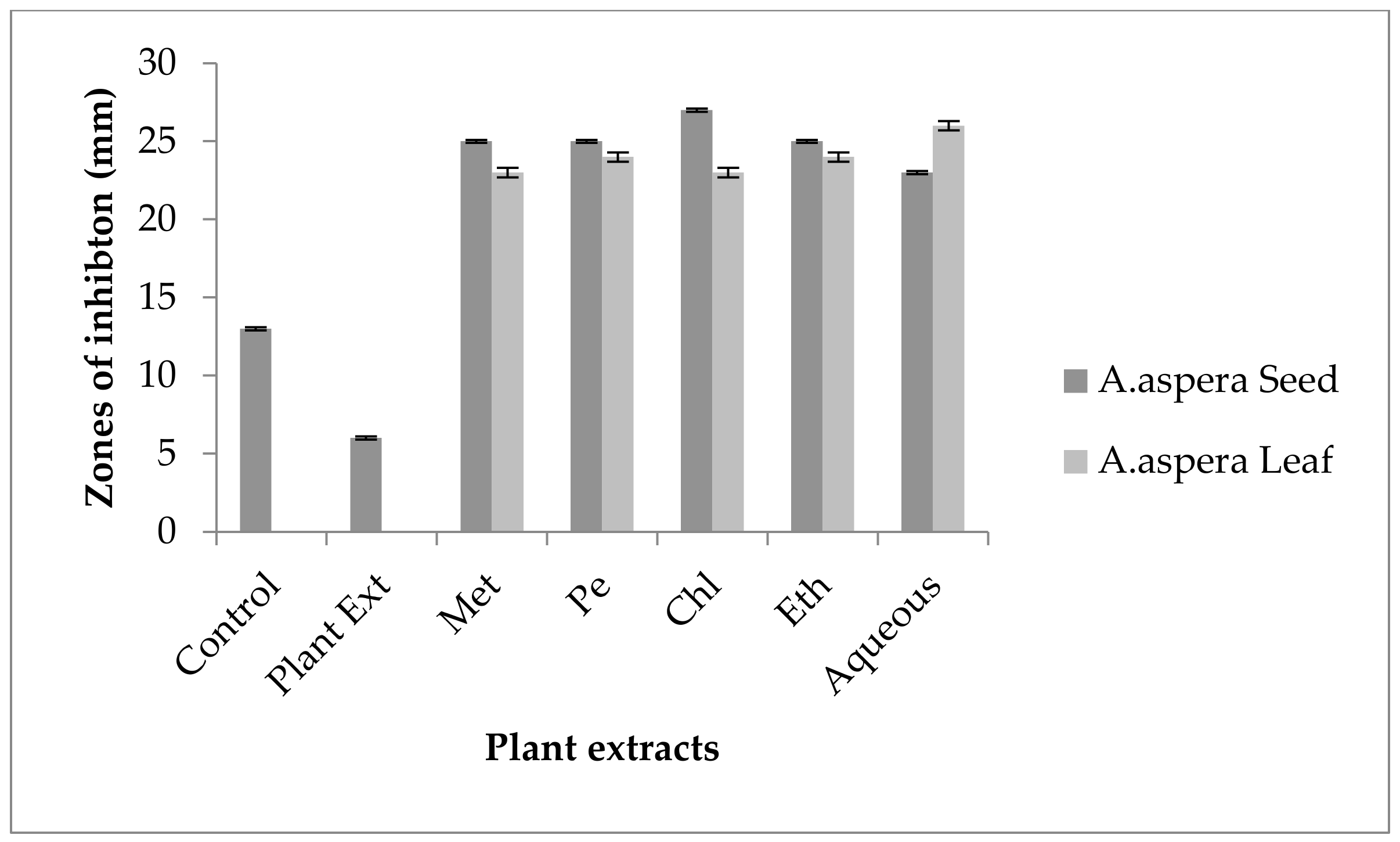
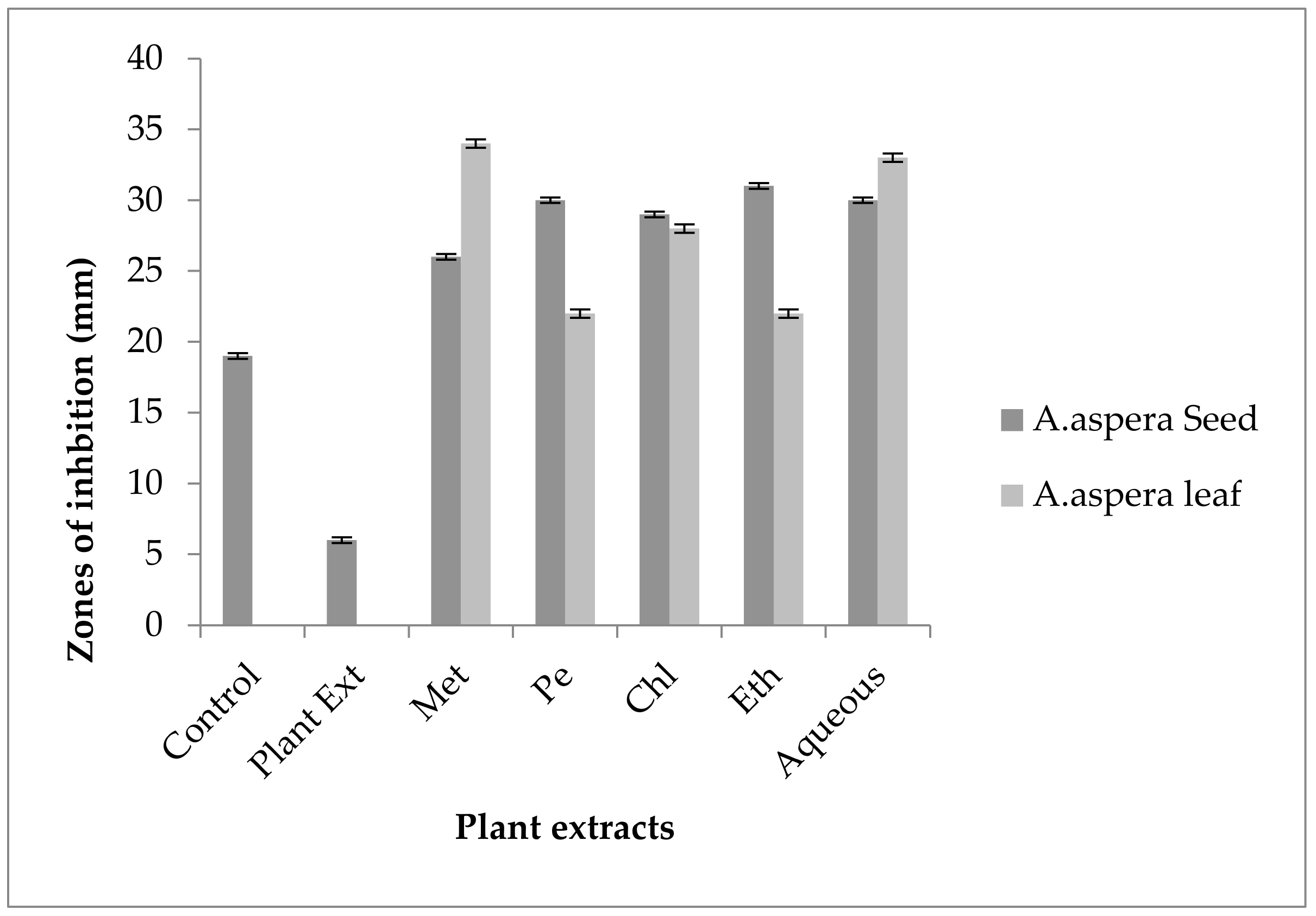
3.3. AST (Antibiotic Susceptibility Test) for Plant Extracts
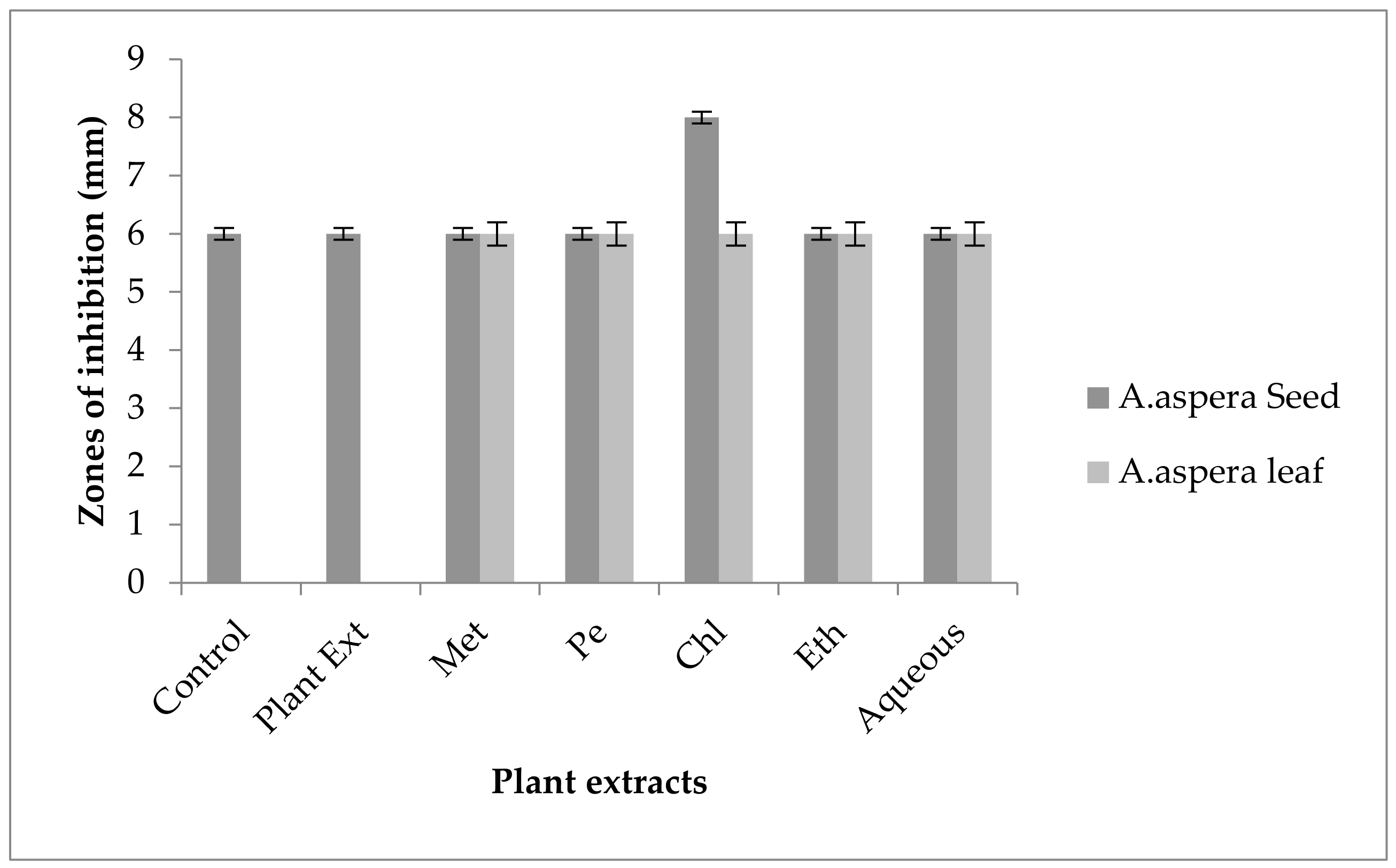
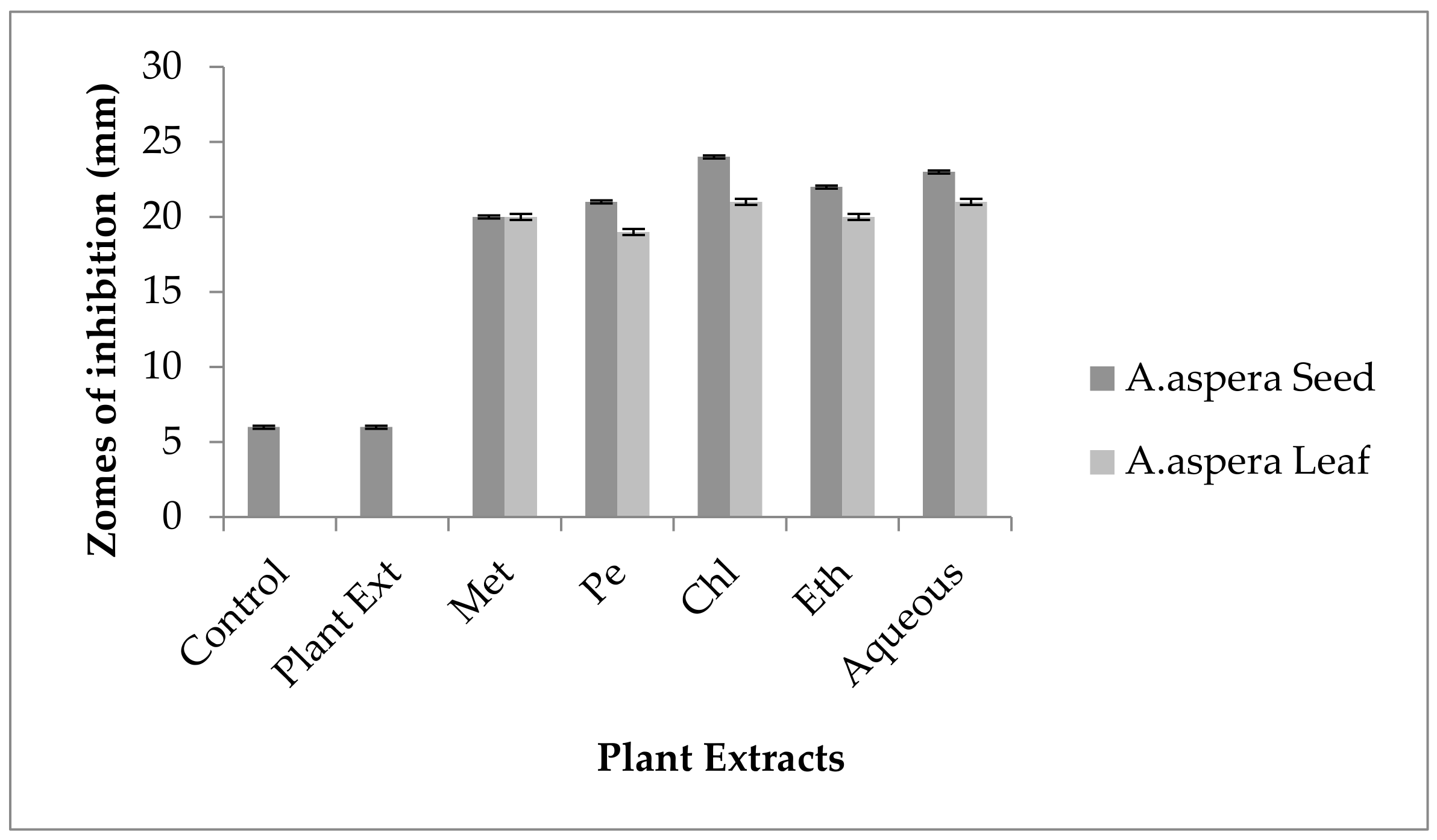
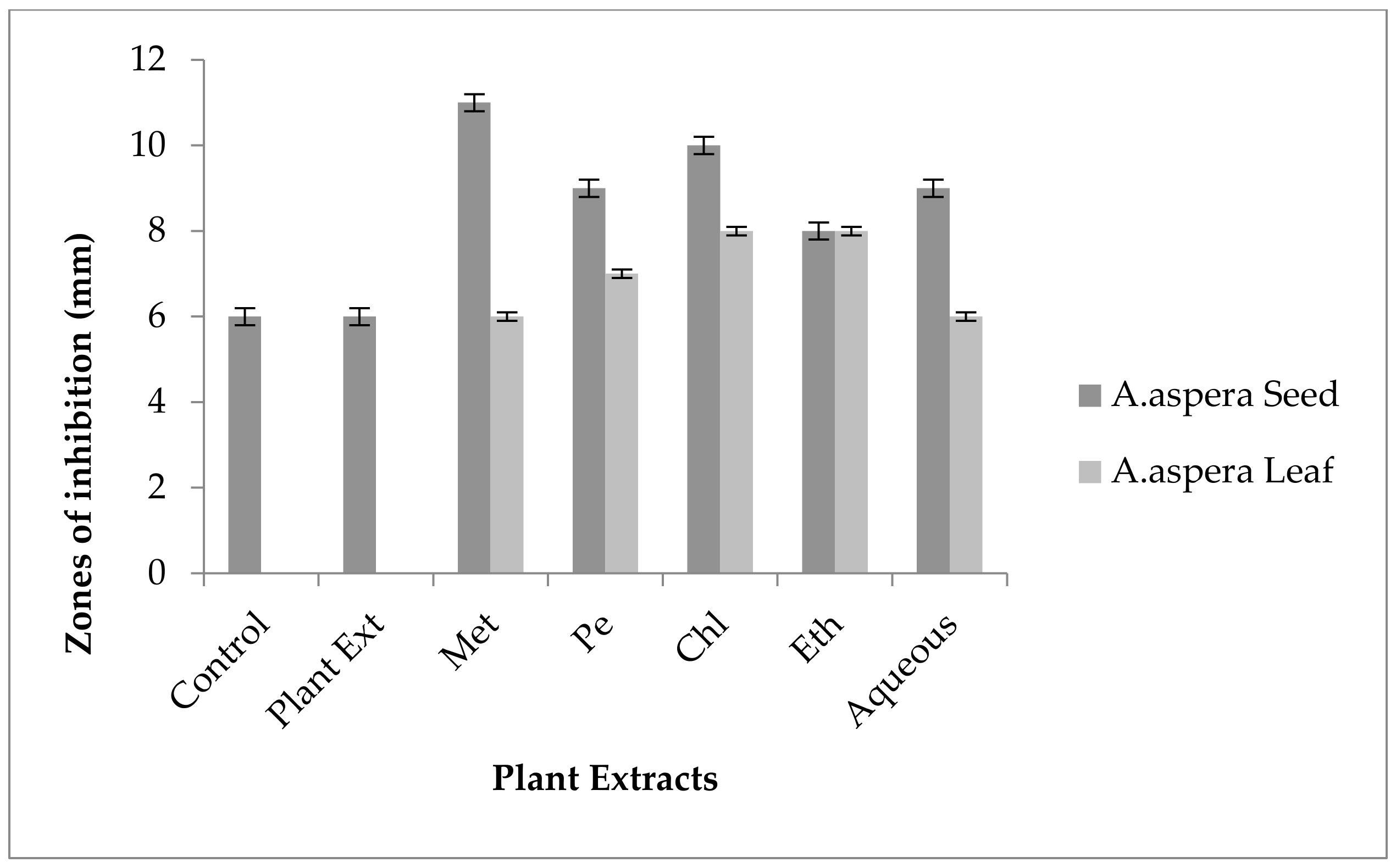
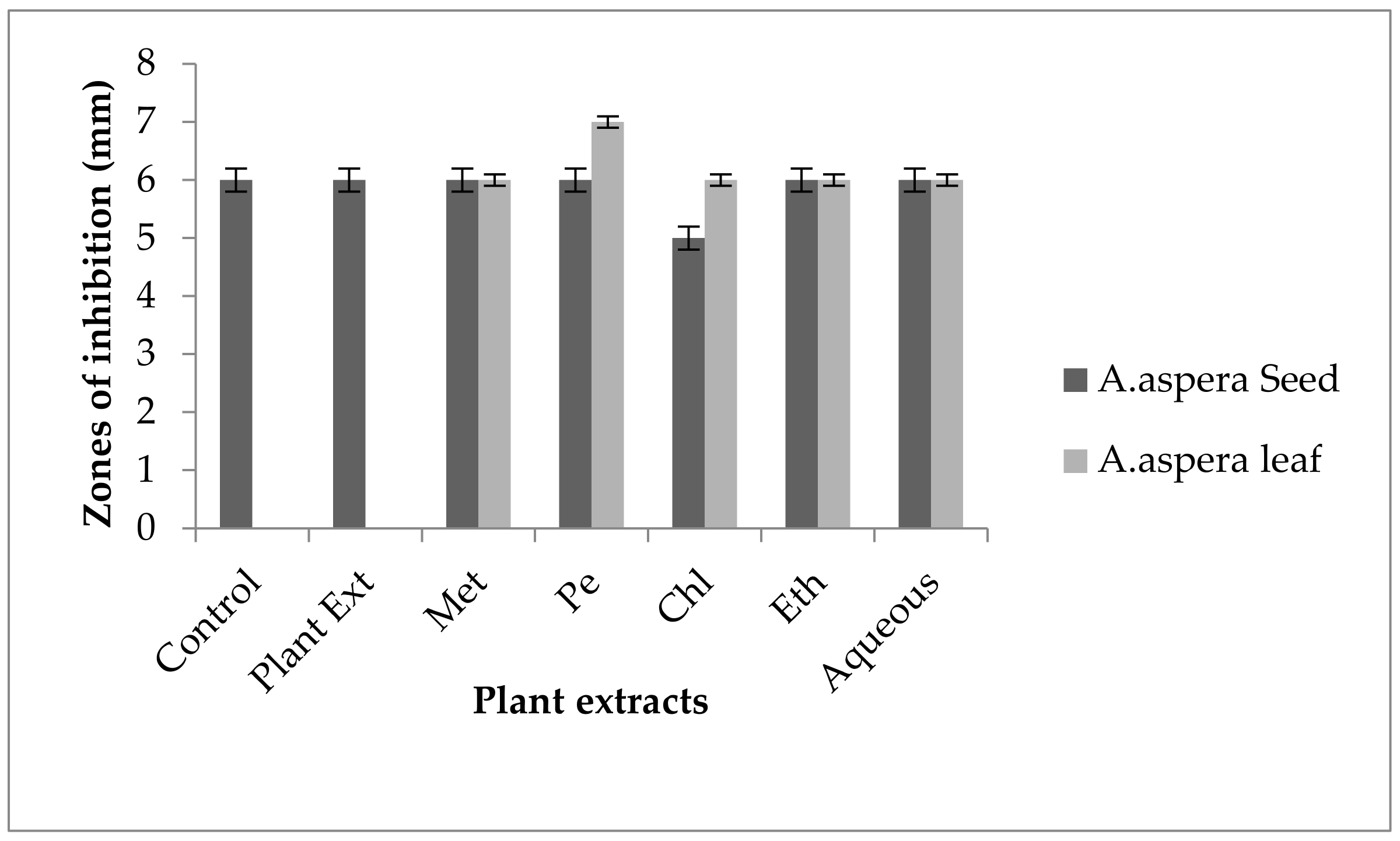
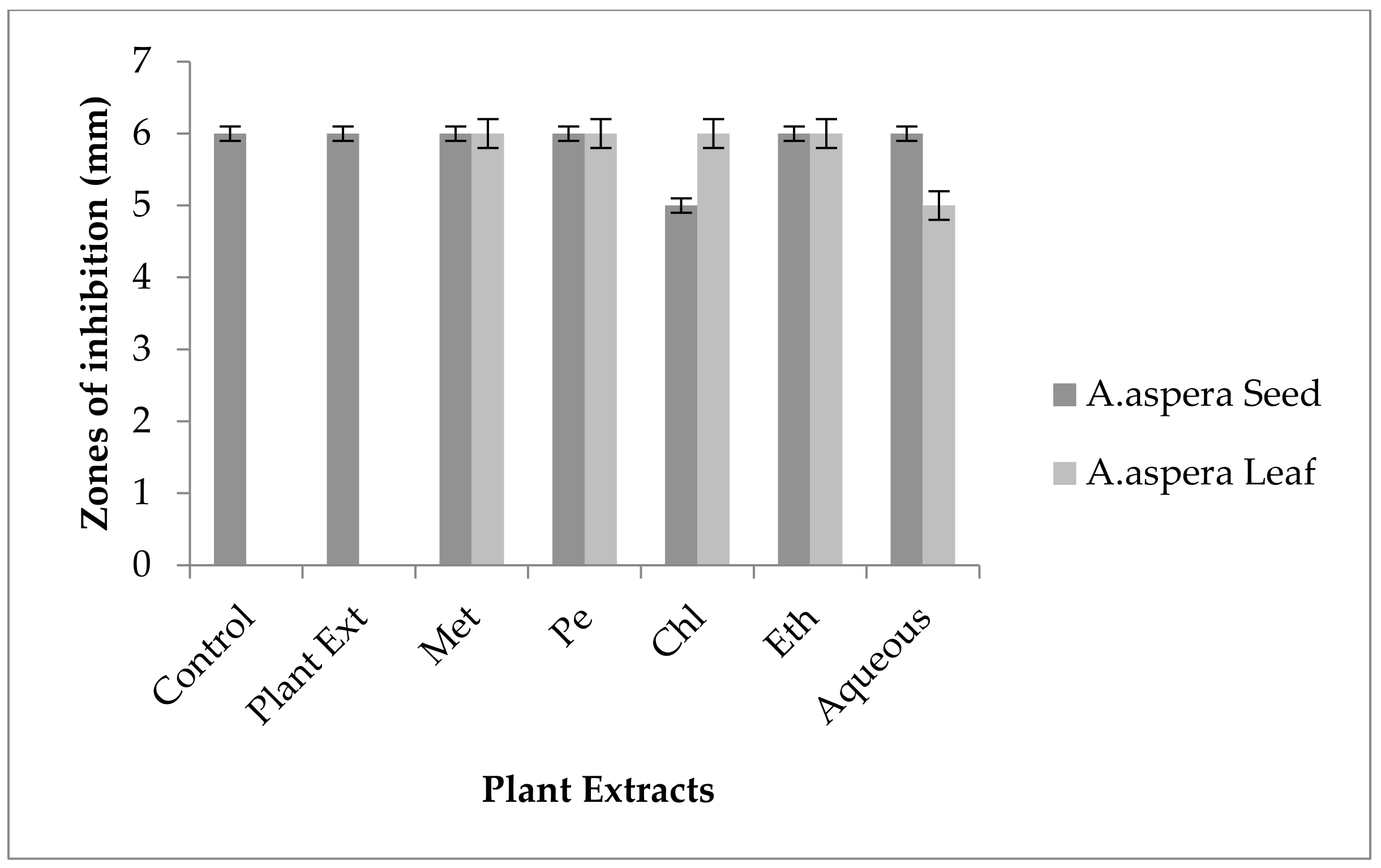
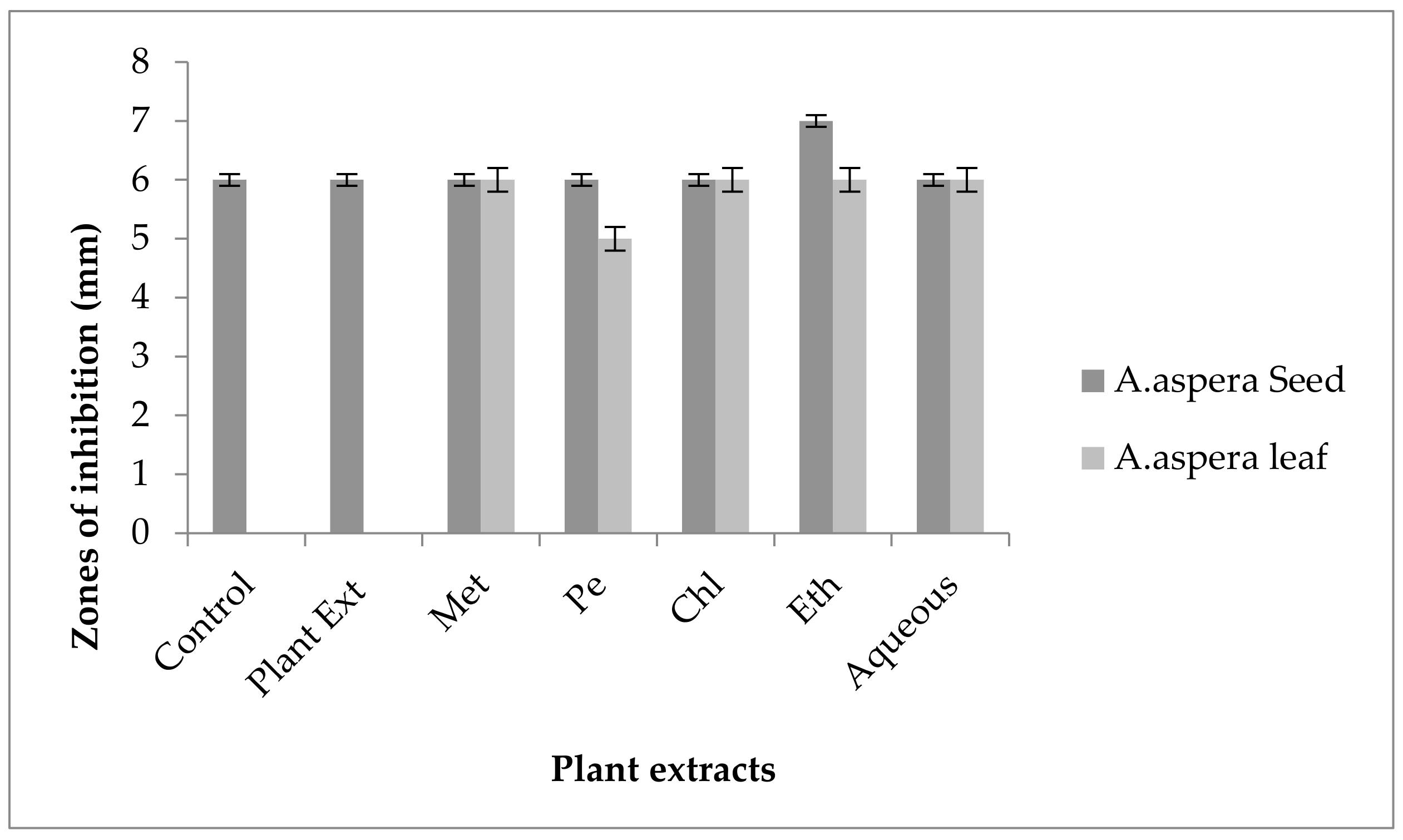
3.3.1. AST Results for Achyranthes aspera Seed
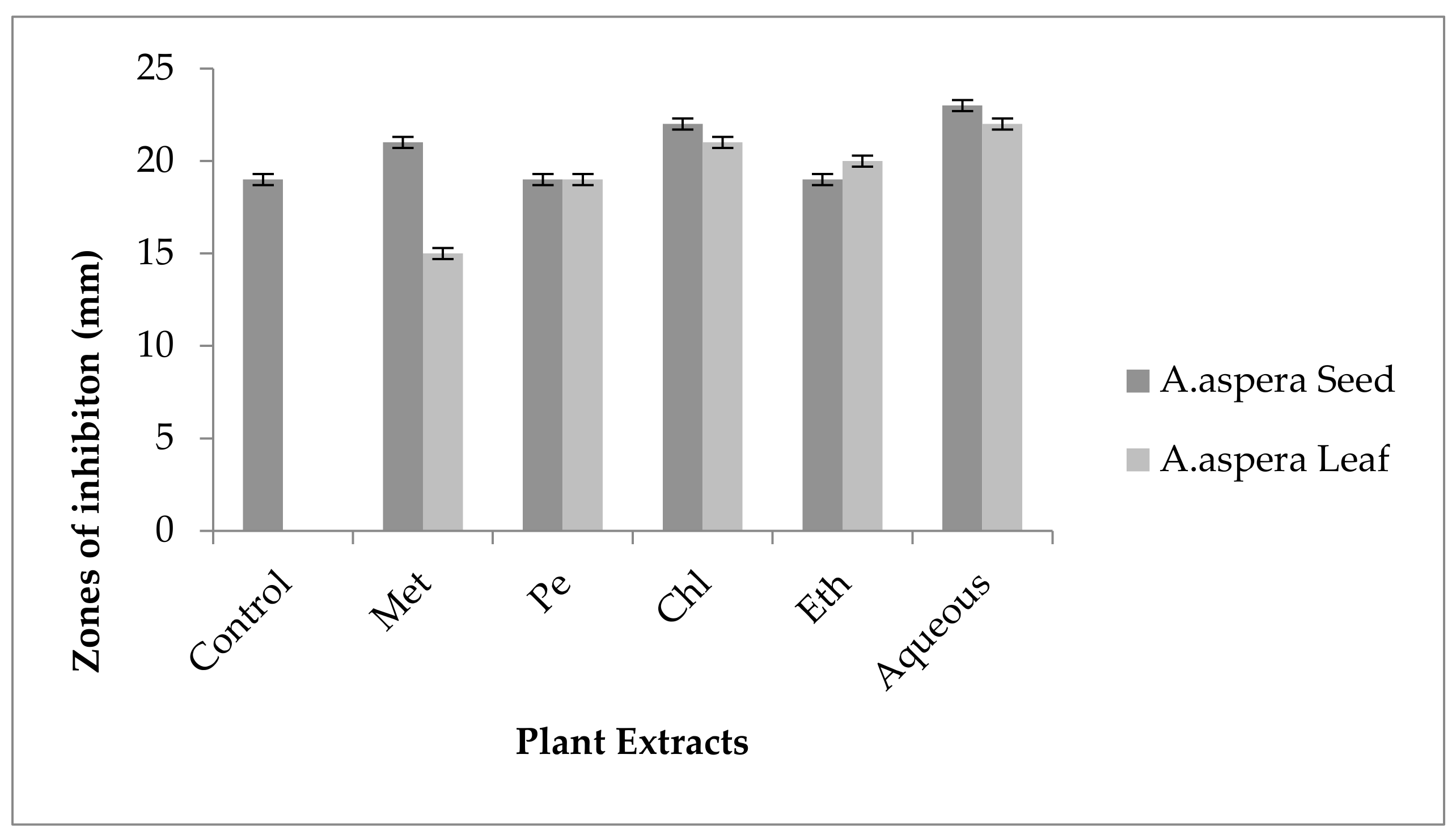
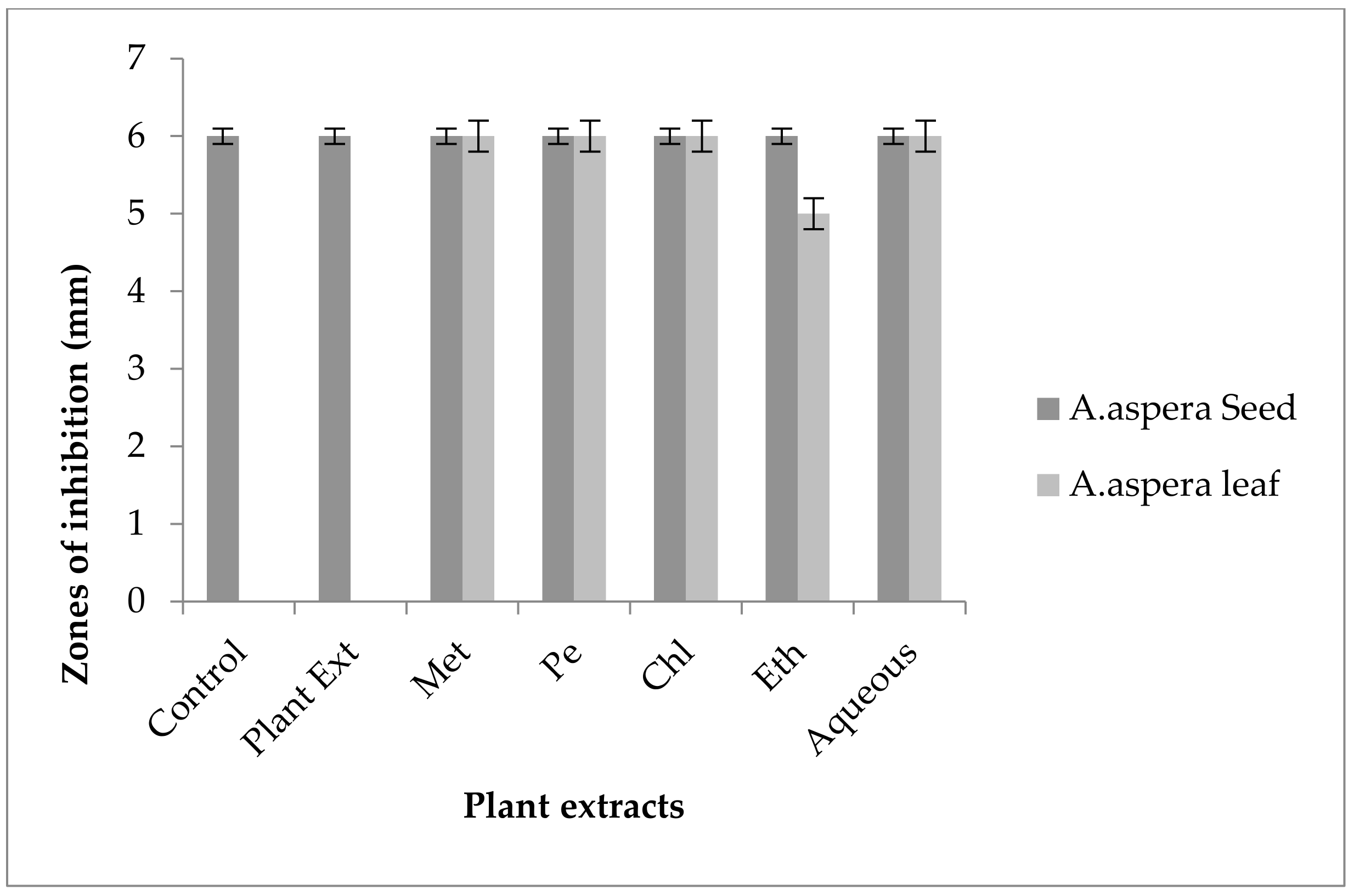
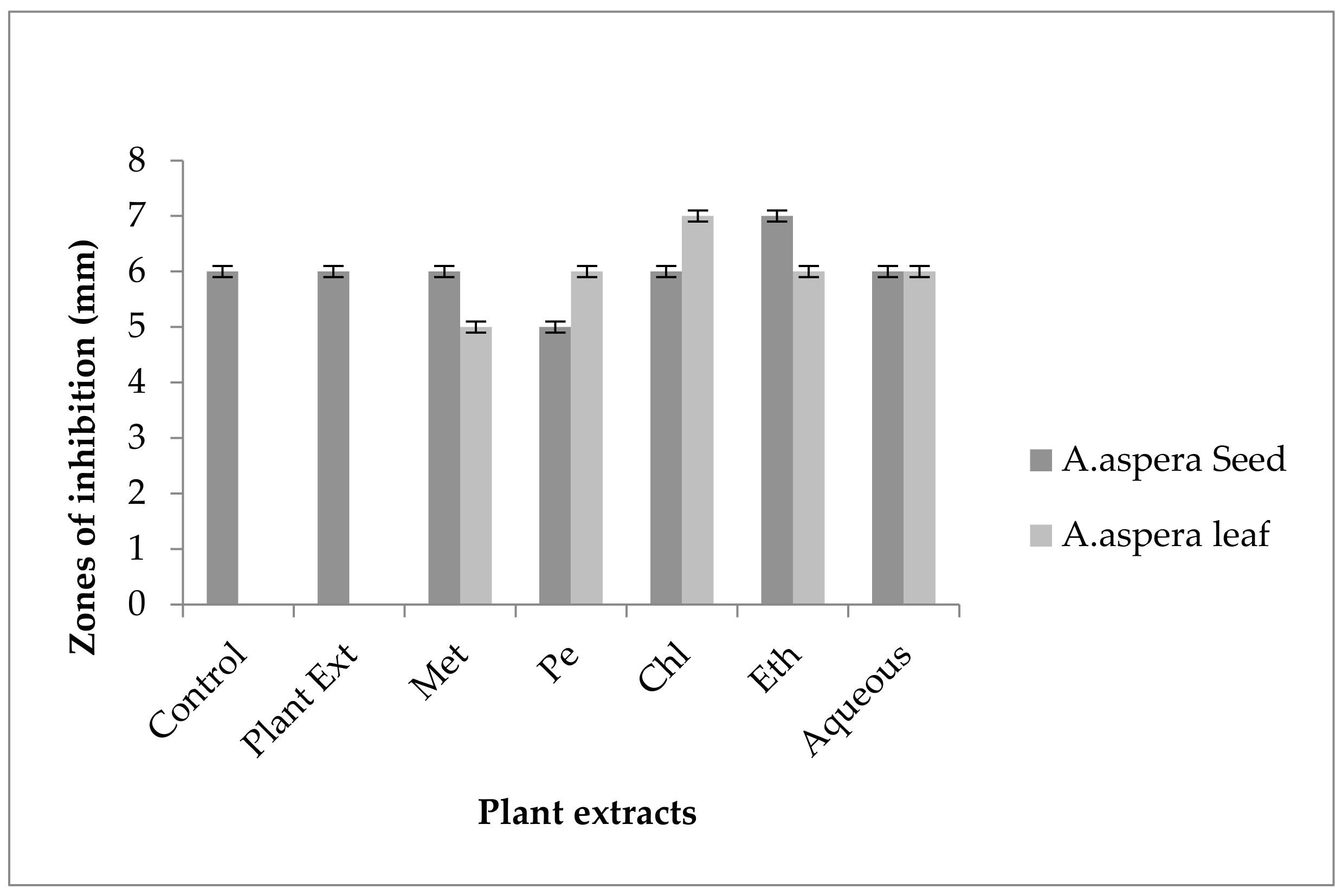
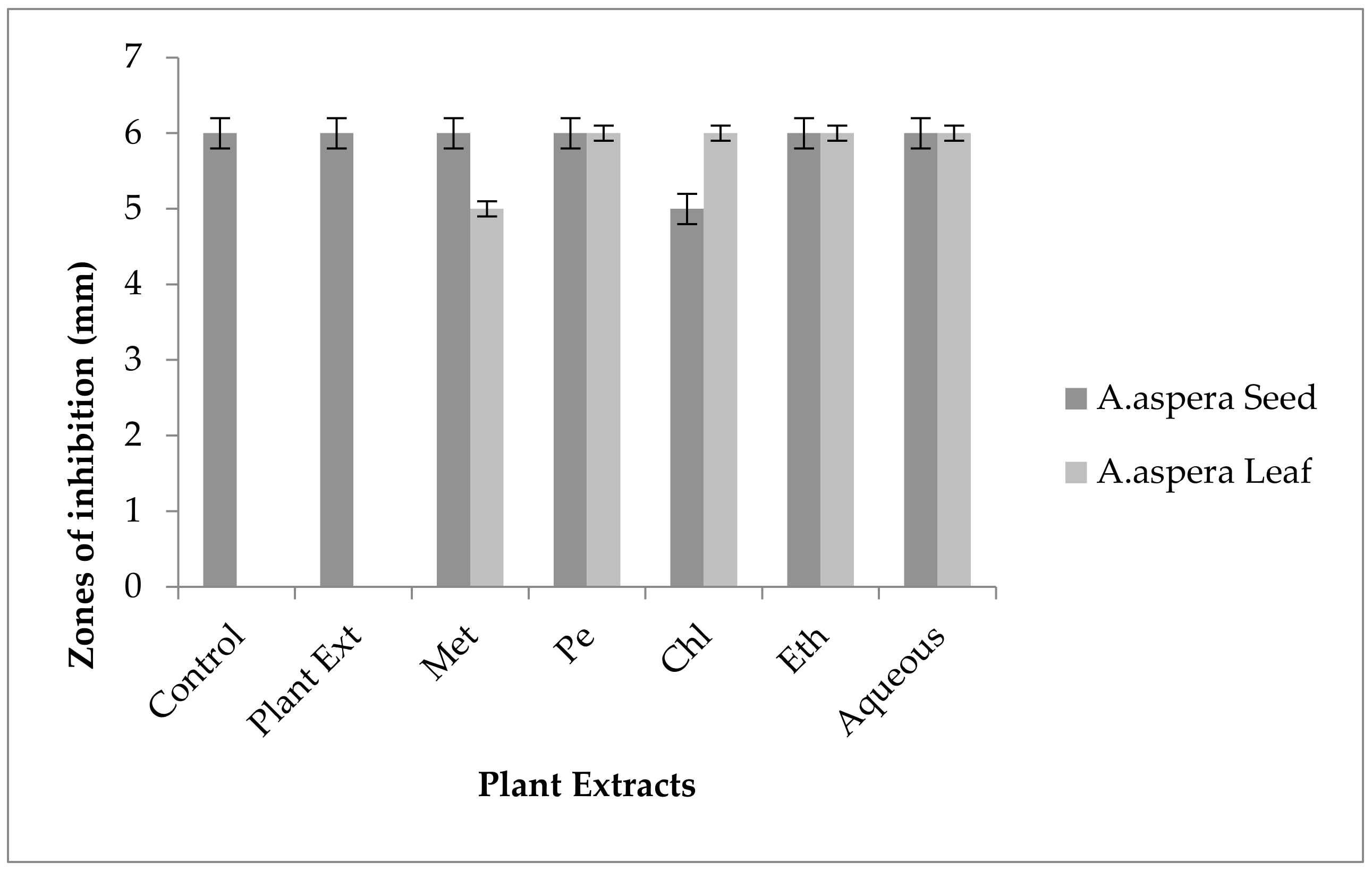
3.3.2. AST Results for Achyranthes aspera Leaves
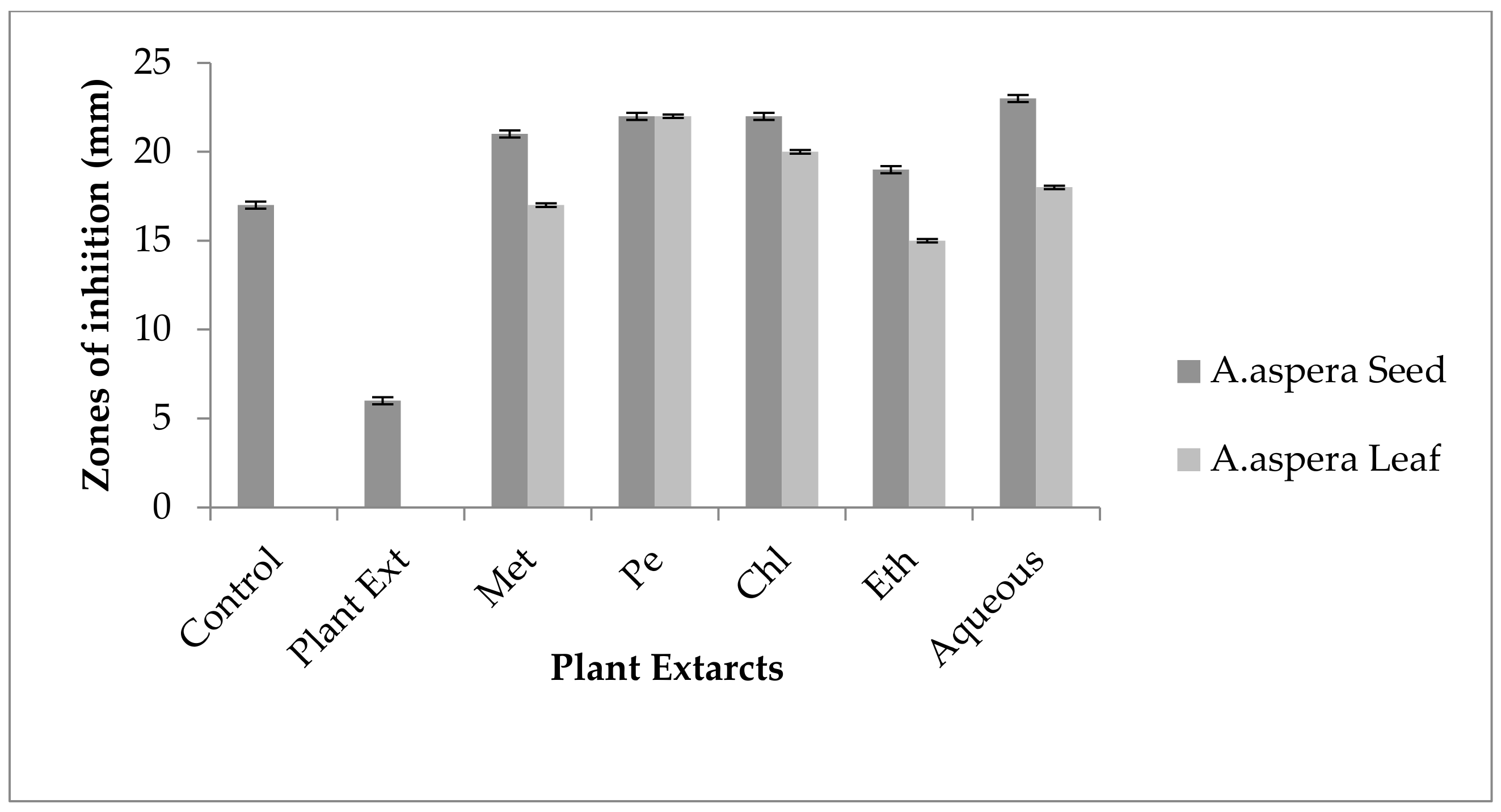
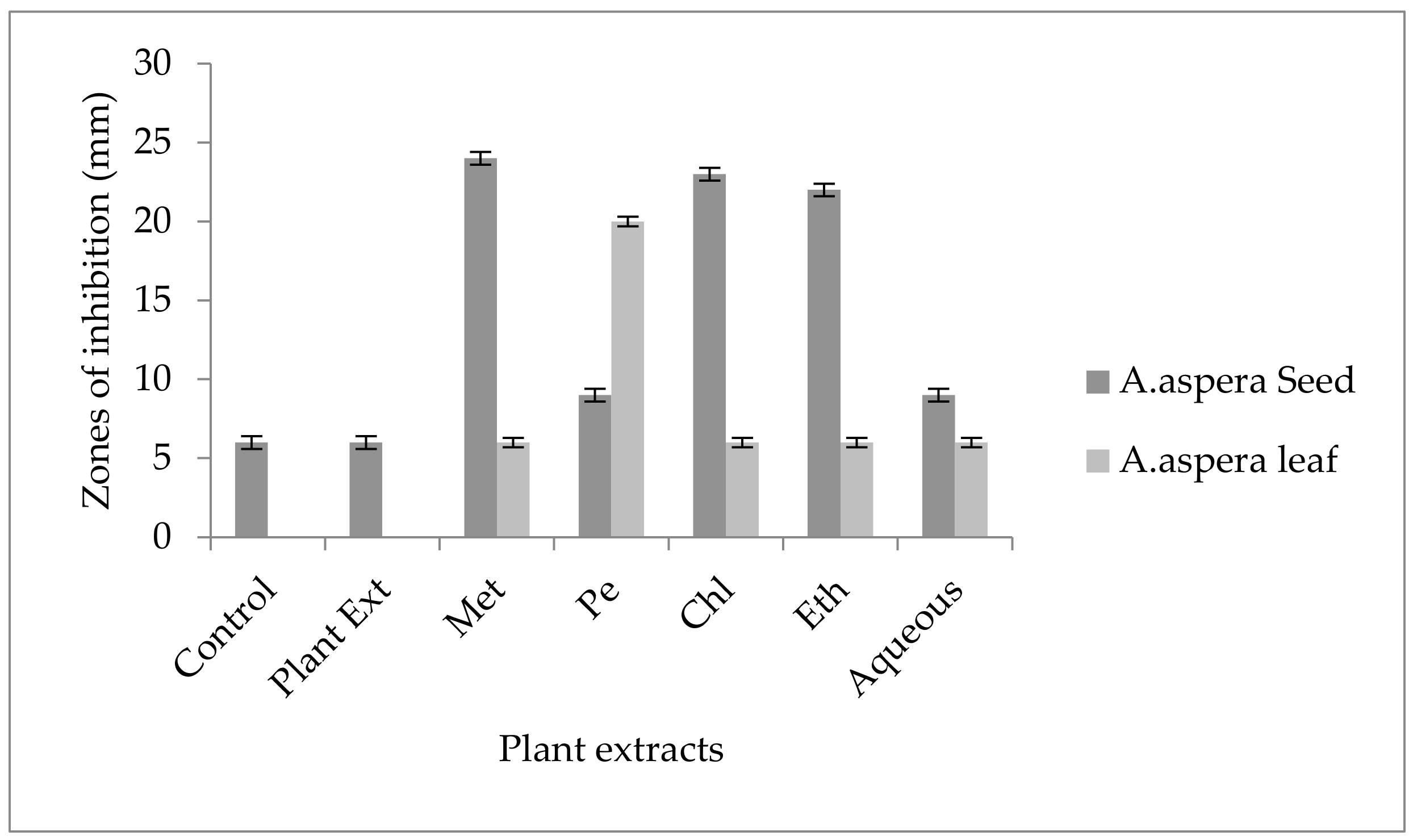
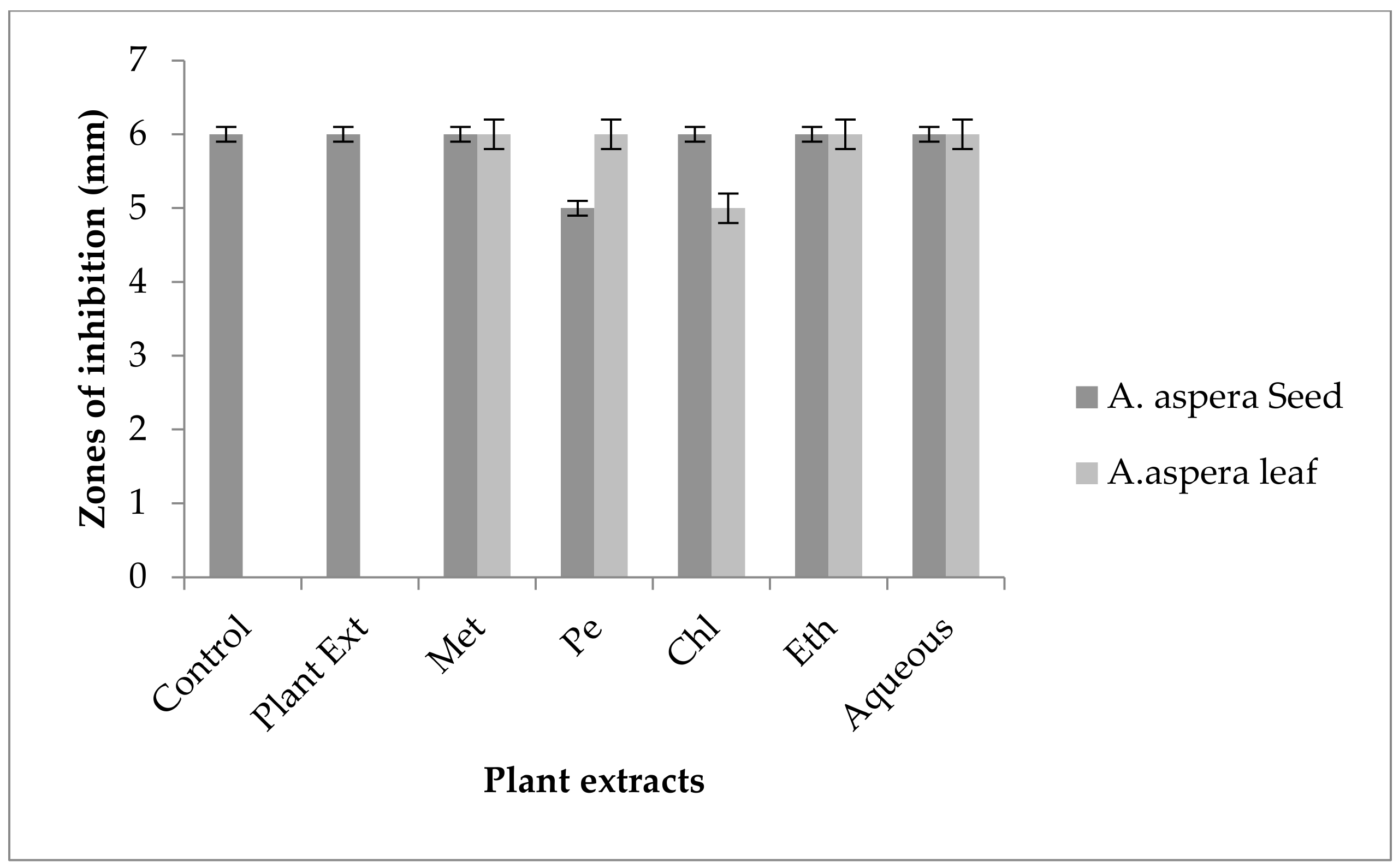
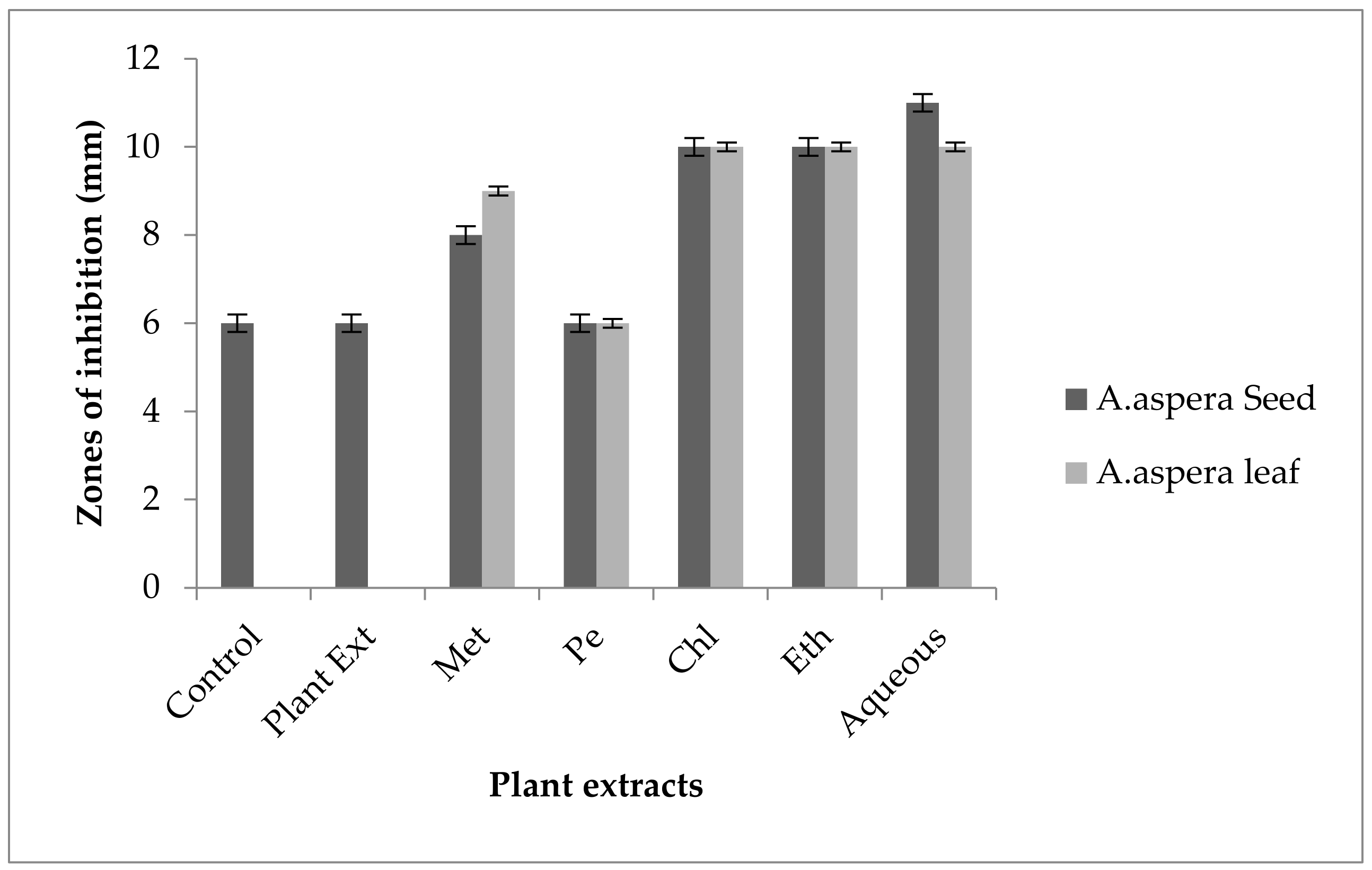
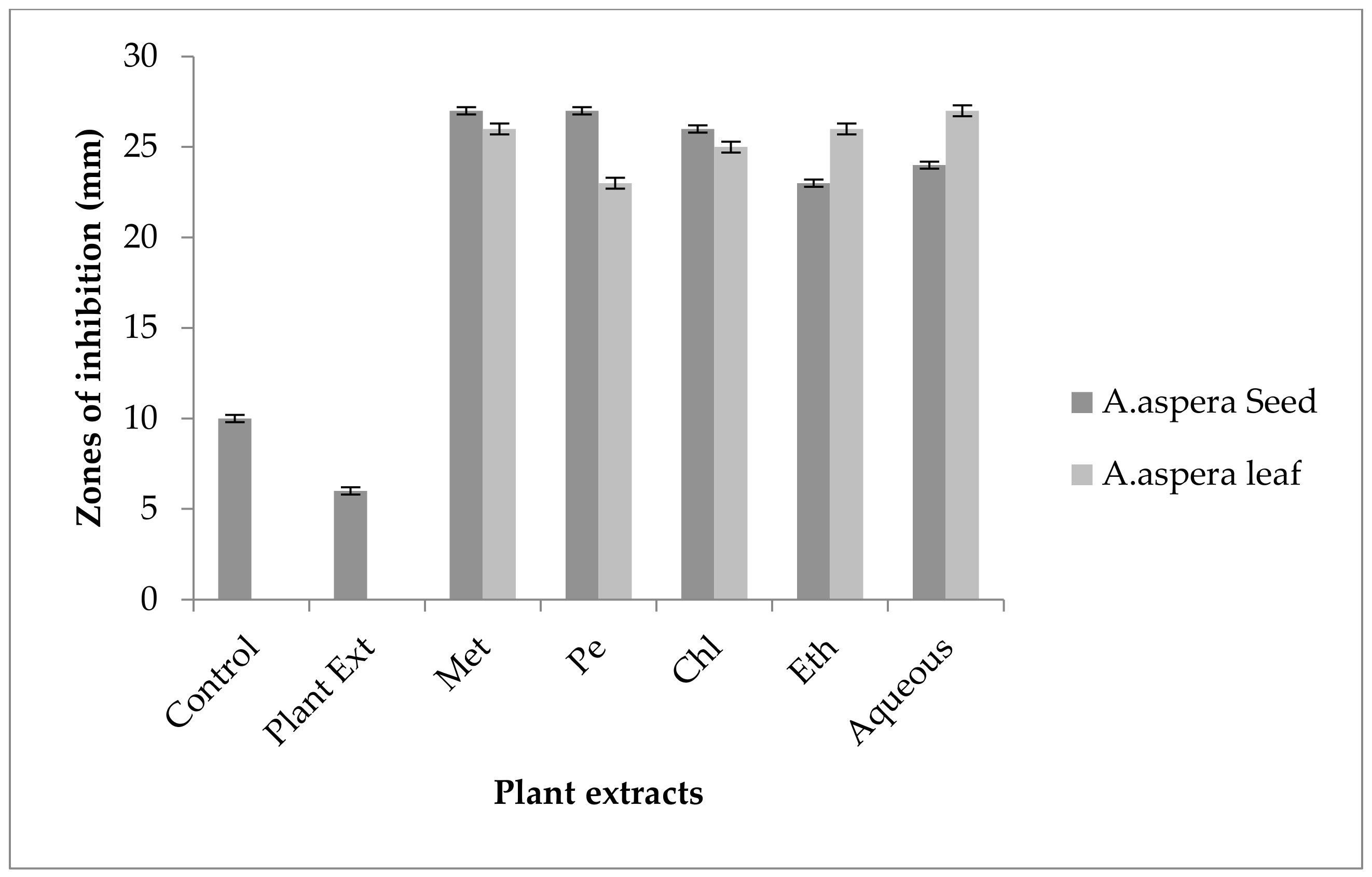
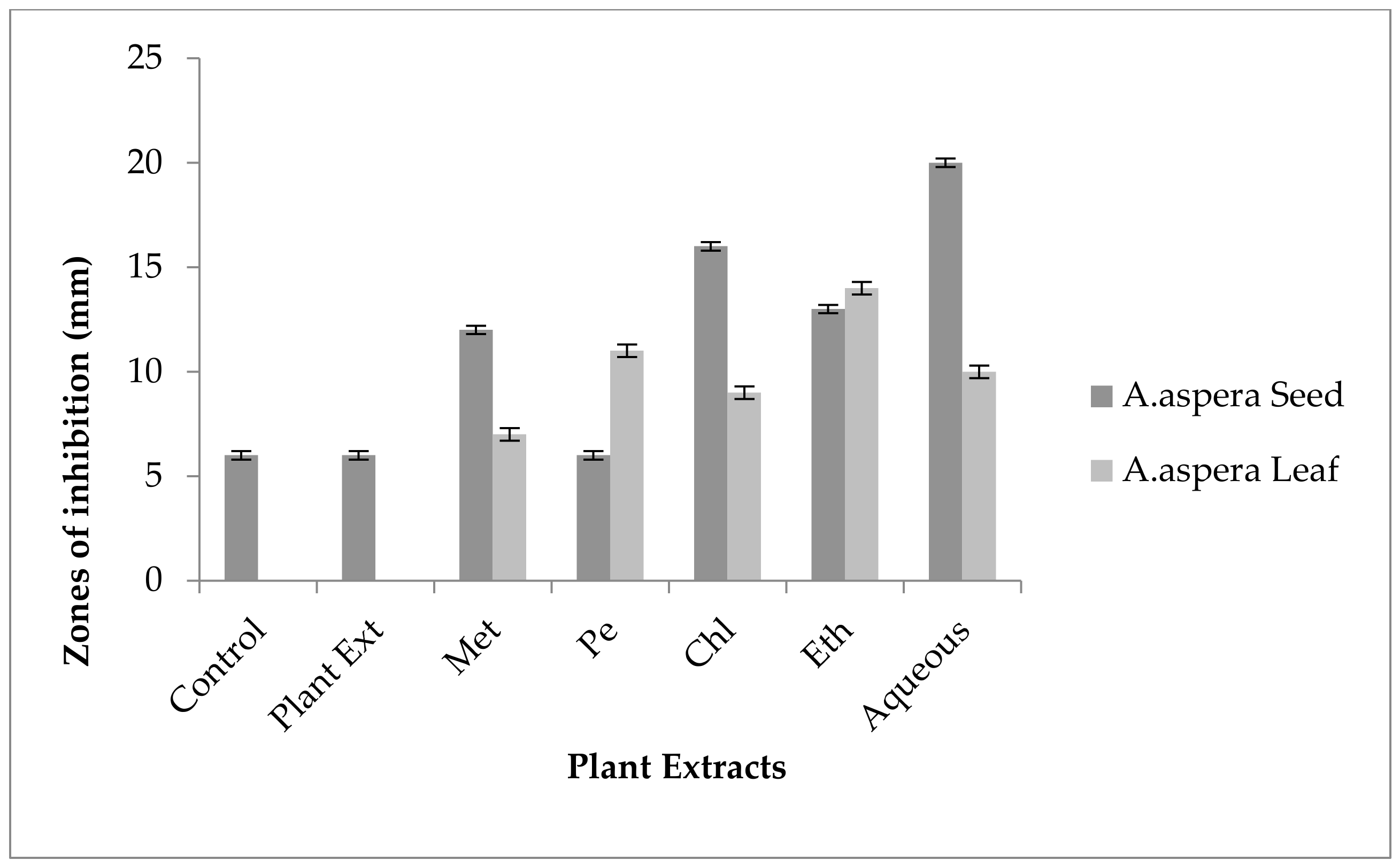
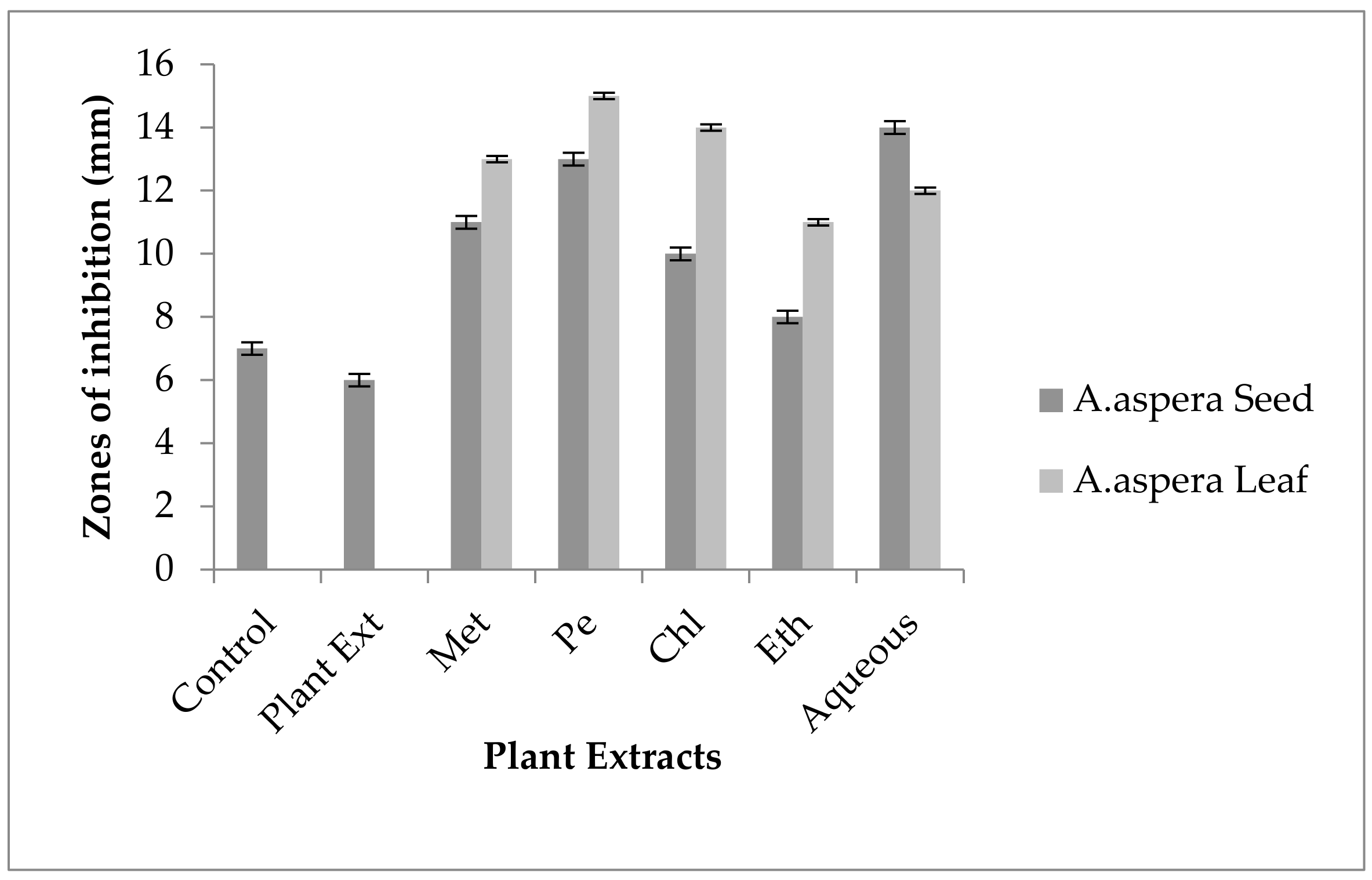
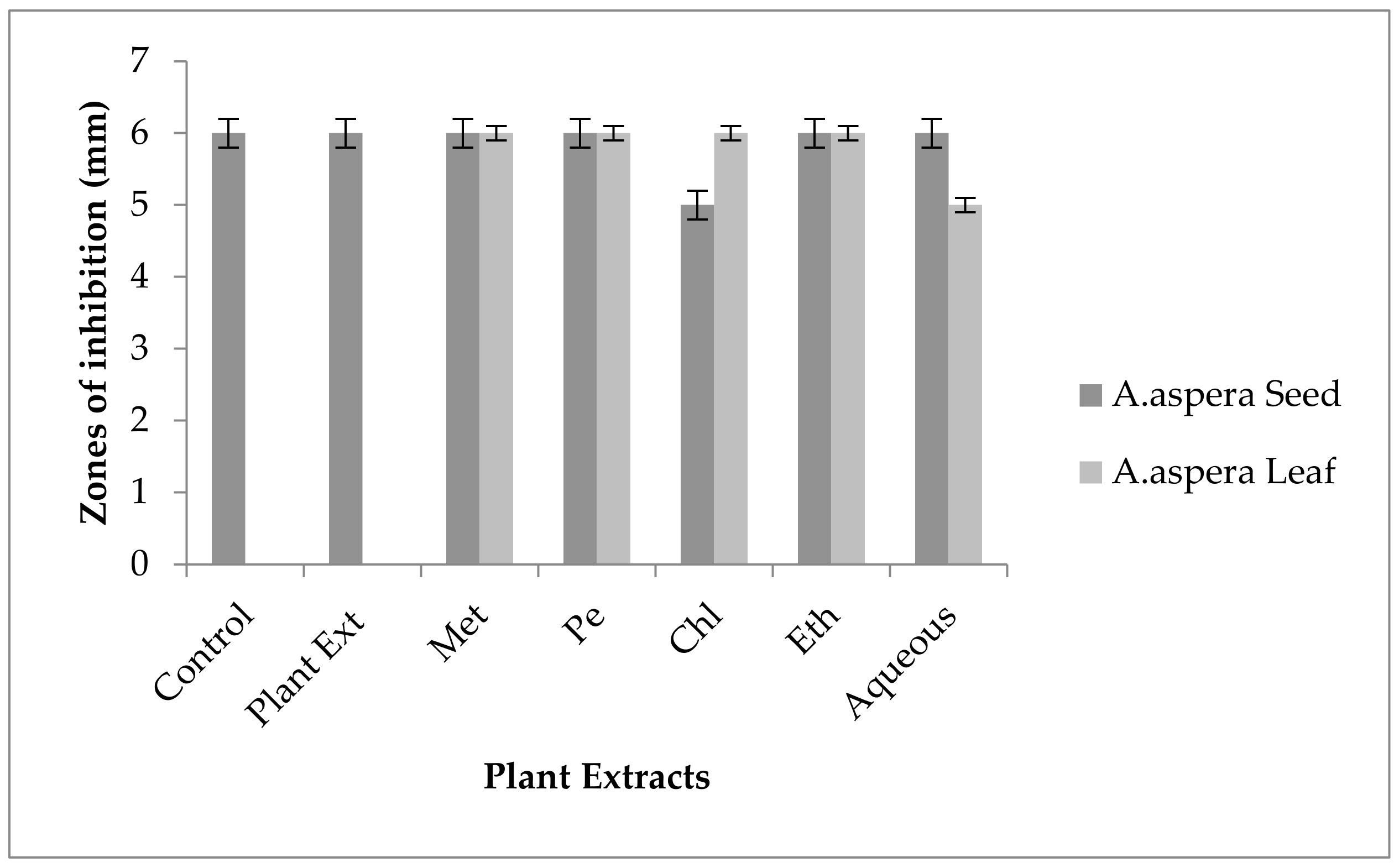
3.4. Redressal Activity of Antibiotic Resistance for MRSA
3.4.1. Ciprofloxacin
3.4.2. Amikacin
3.4.3. Cefoxitin
3.4.4. Levofloxacin
3.4.5. Penicillin
3.4.6. Linezolid
3.4.7. Co-trimoxazole
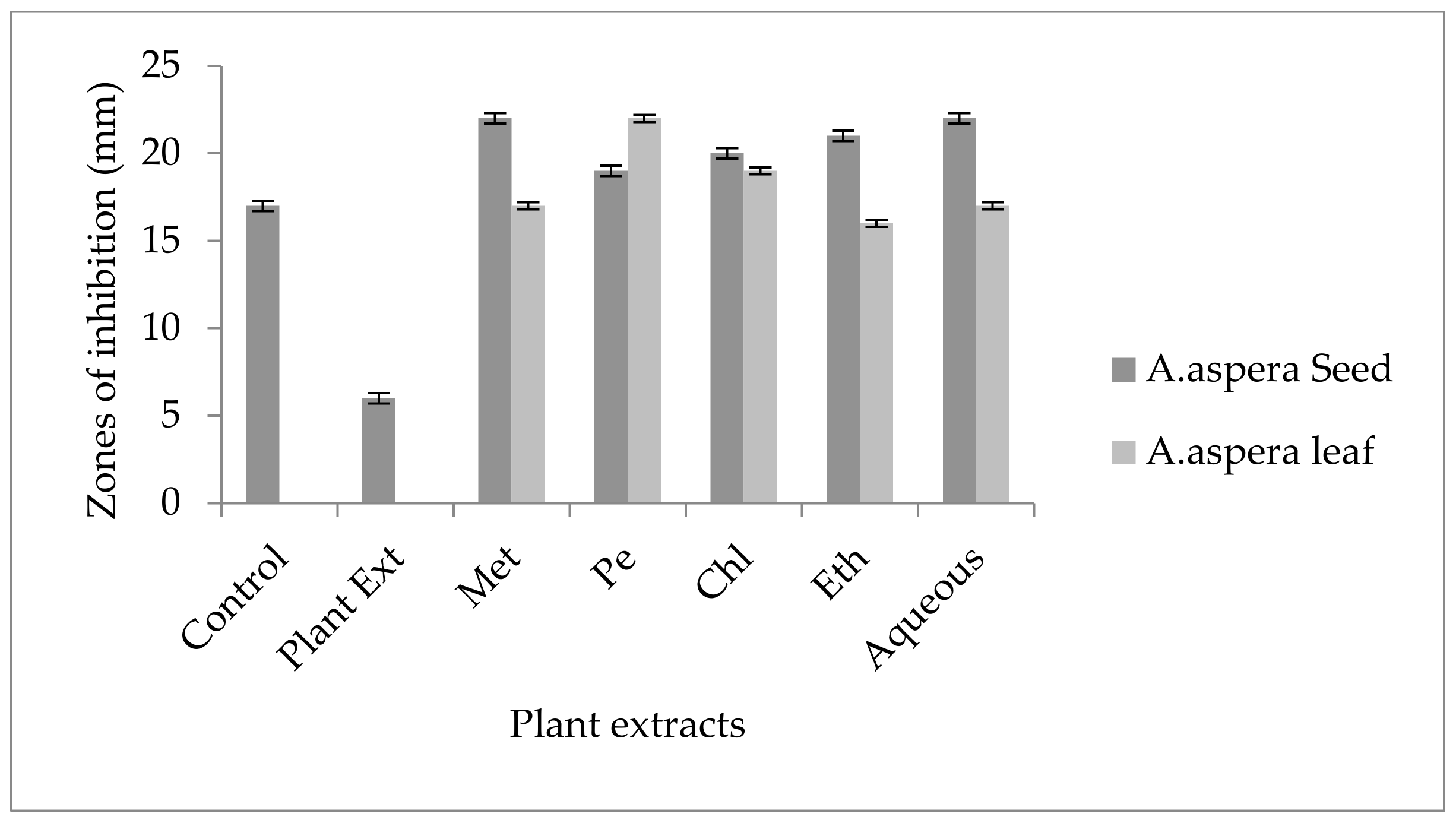
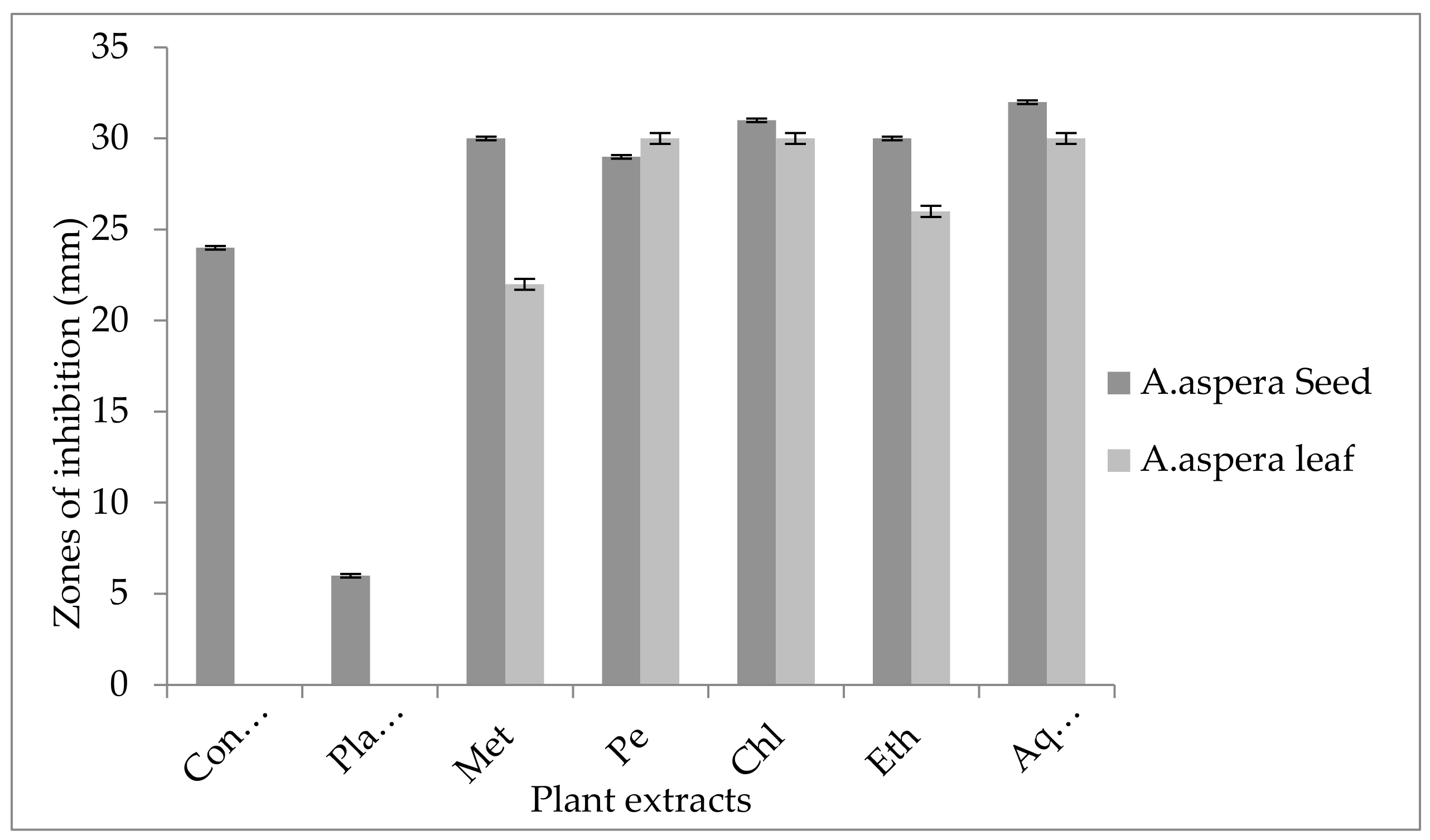
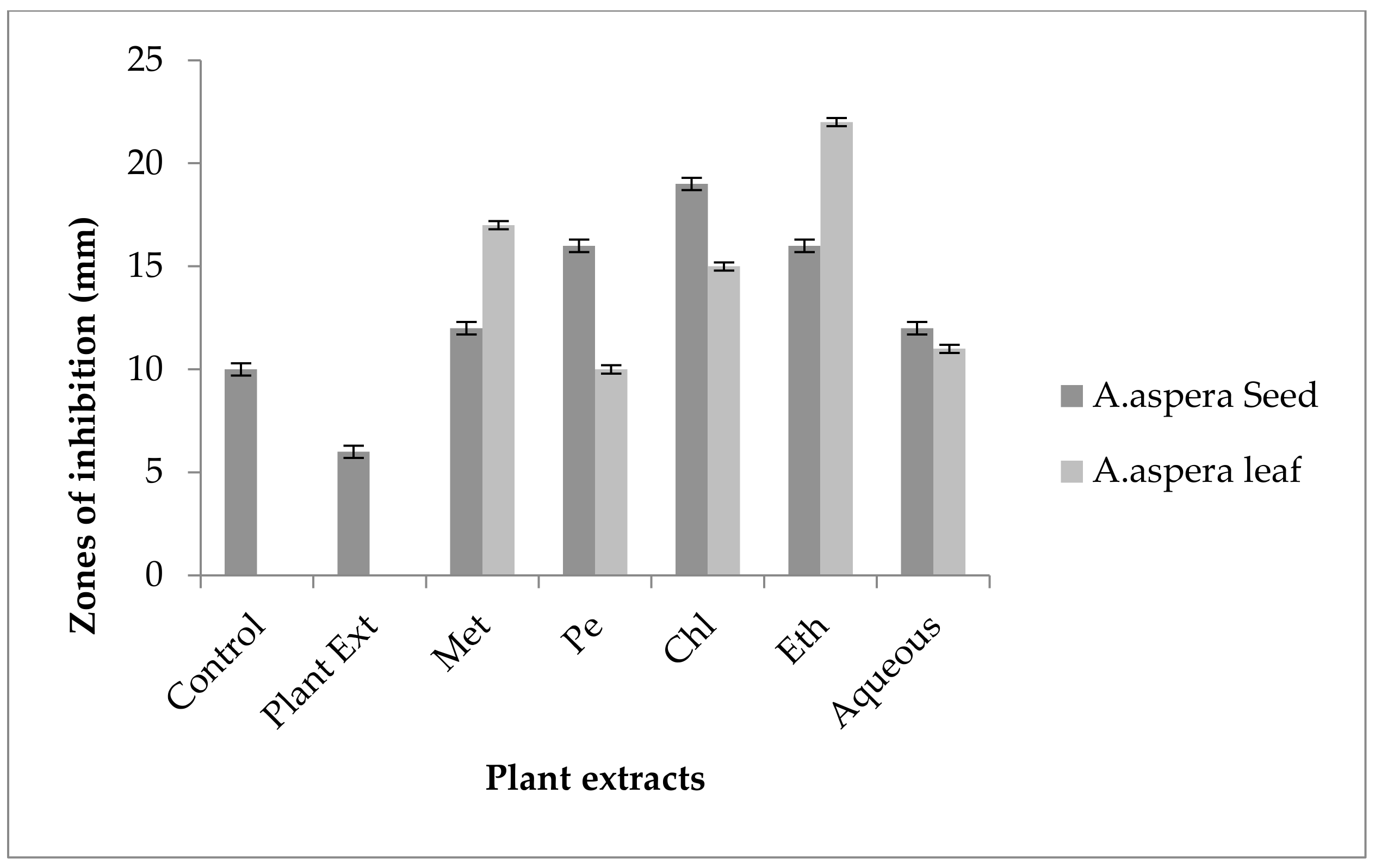
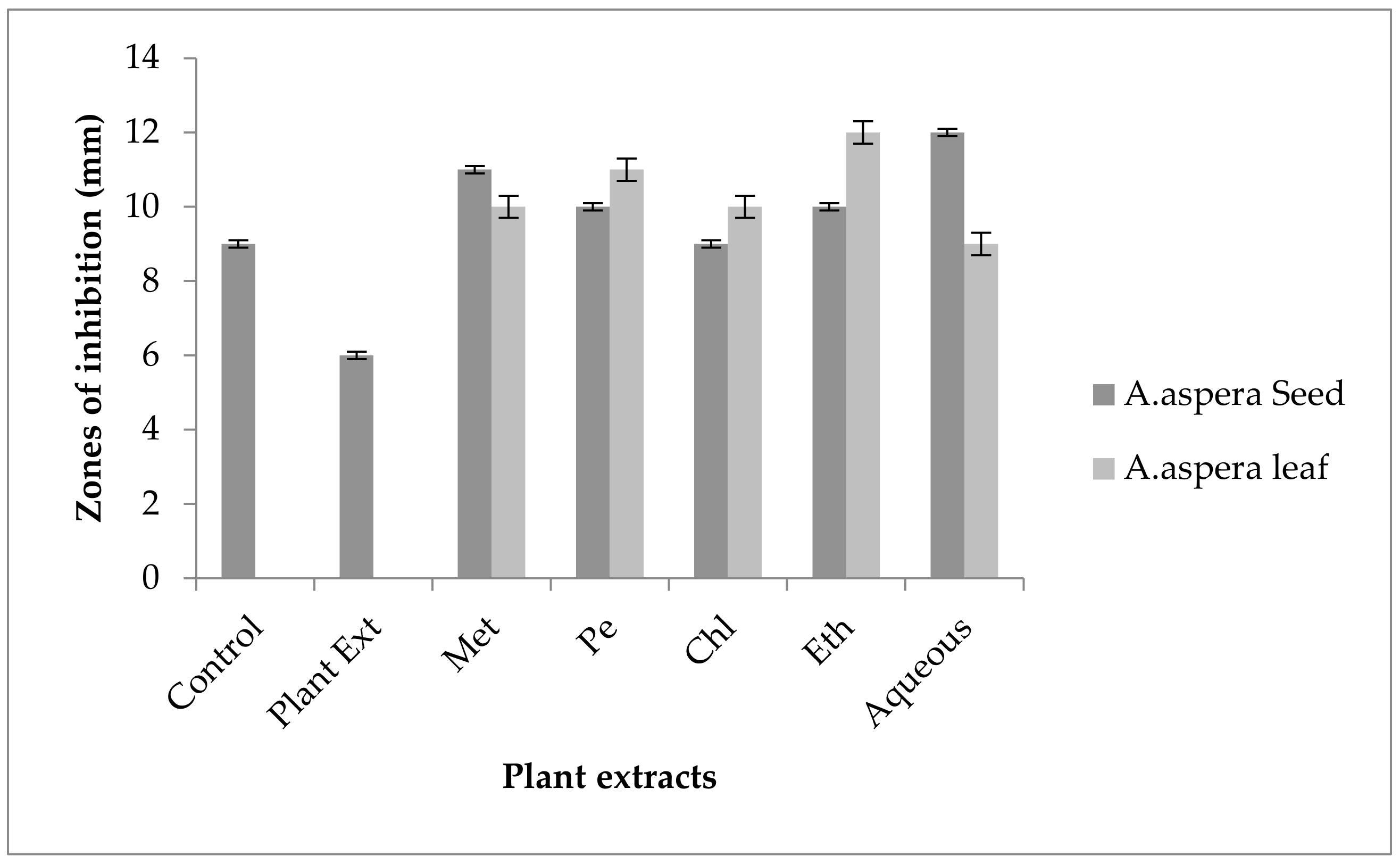
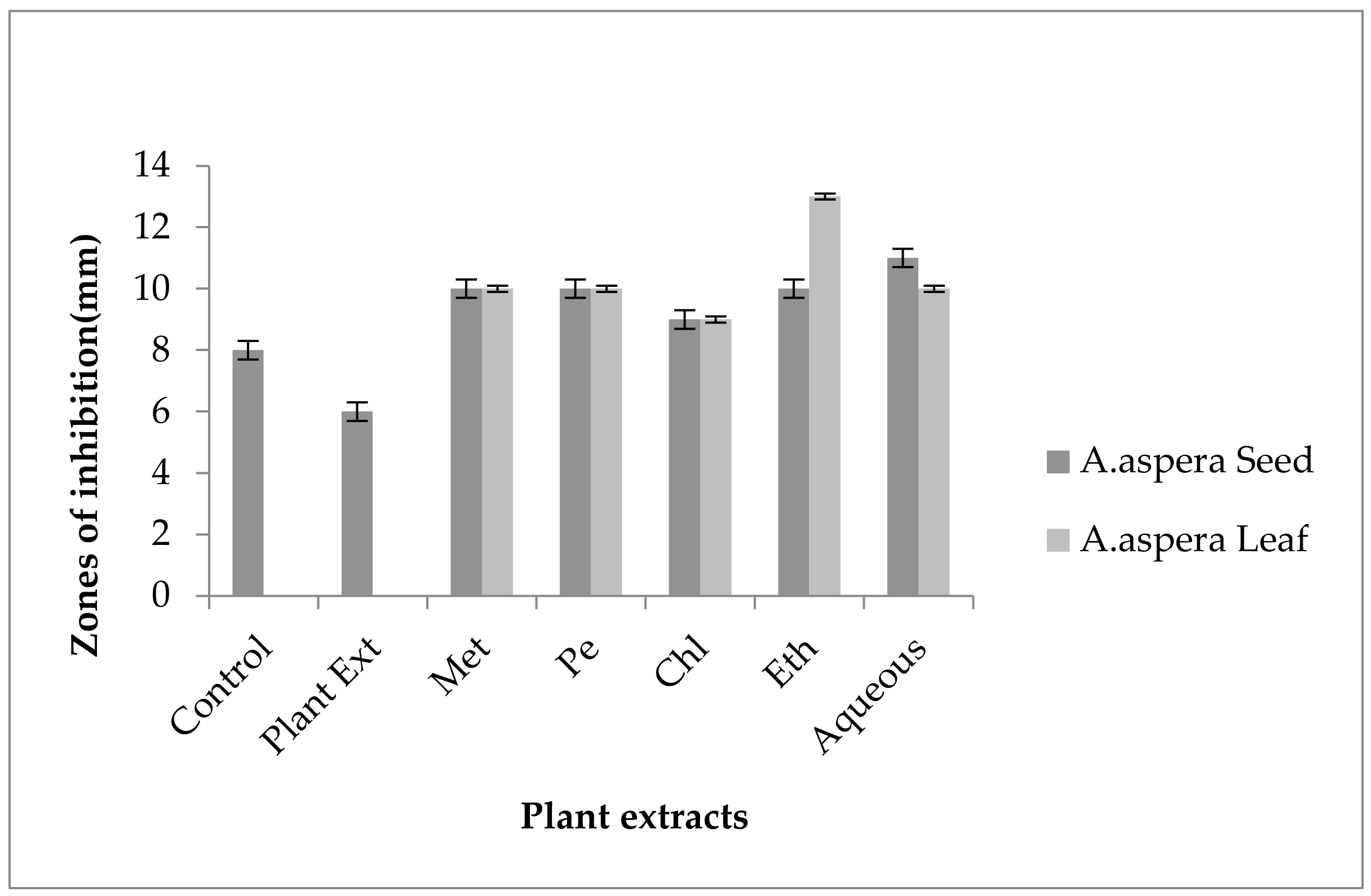
3.5. Antibiotic Resistant Redressal Activity for Enterococcus faecalis
3.5.1. Ciprofloxacin
3.5.2. Amoxicillin–Clavulanate
3.5.3. Linezolid
3.5.4. Penicillin
3.5.5. Levofloxacin
3.5.6. Vancomycin
3.5.7. Imipenem
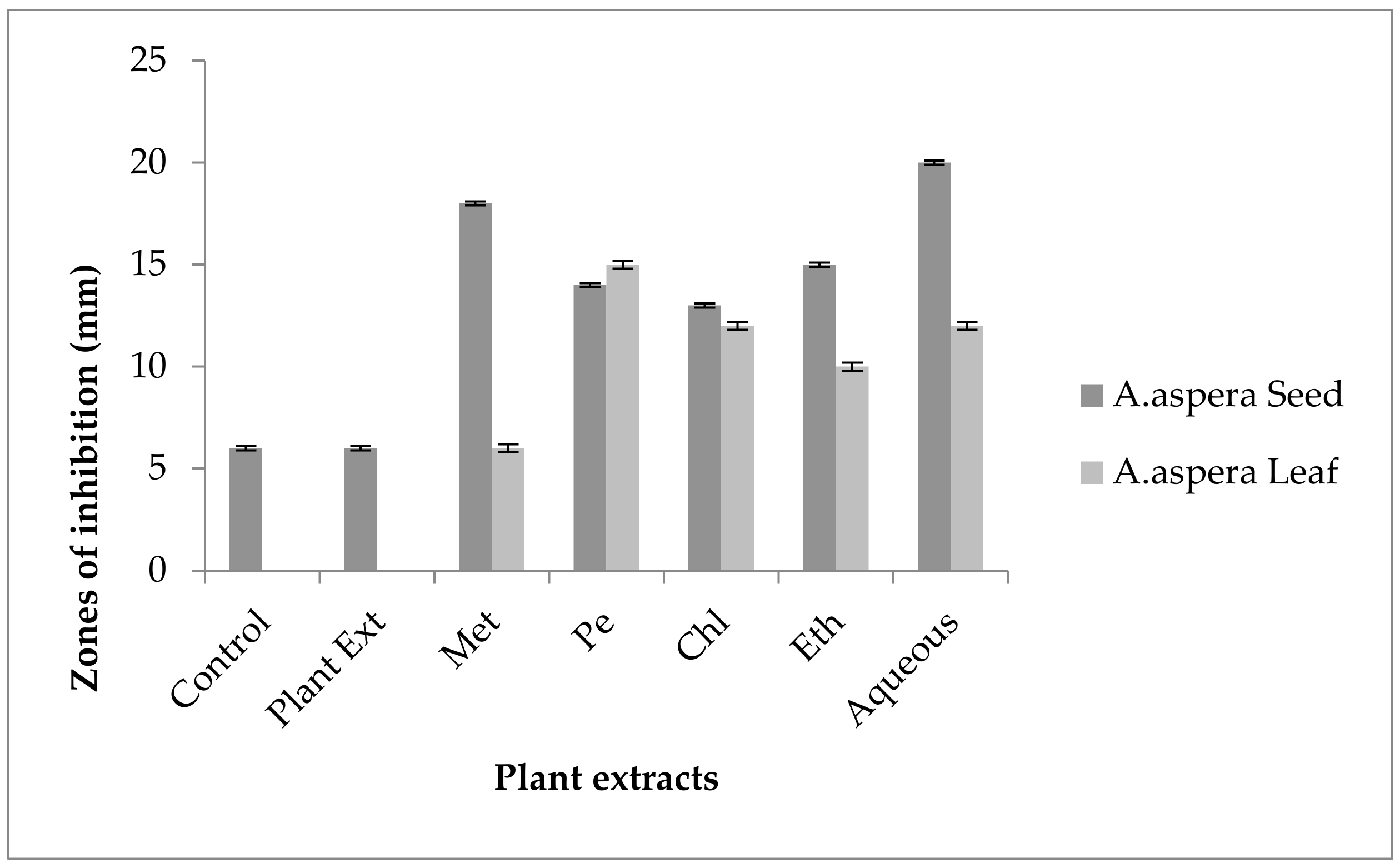
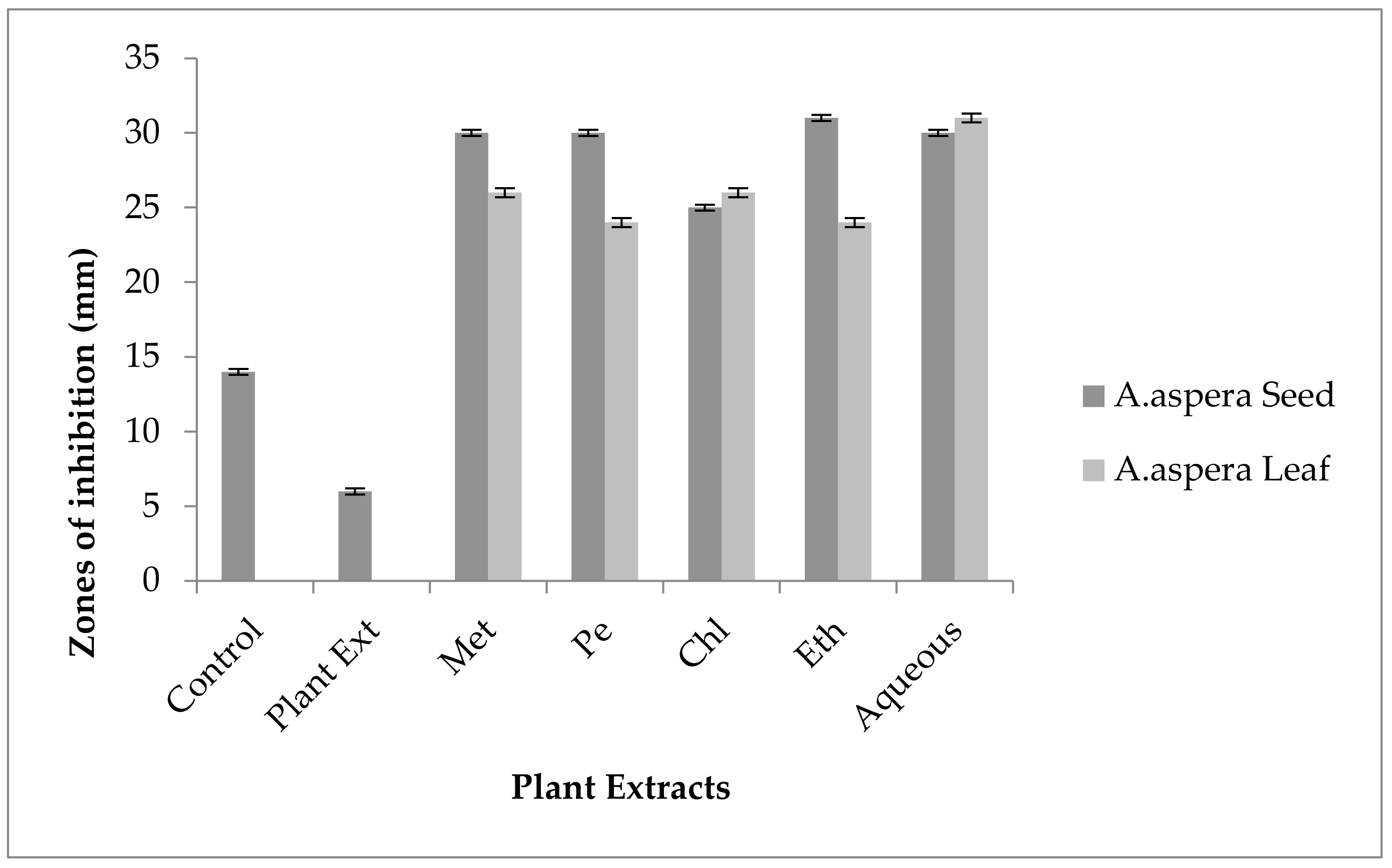
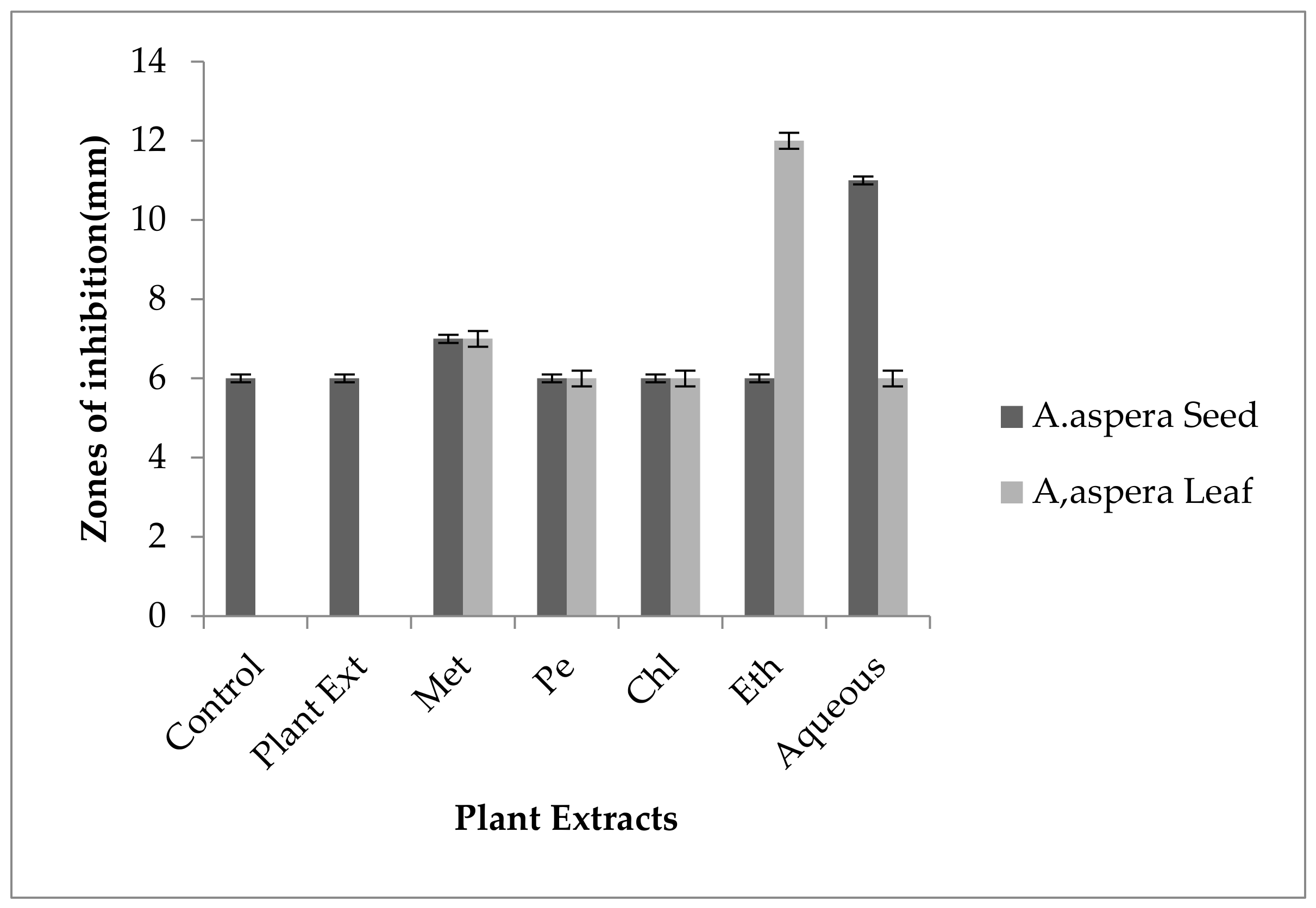
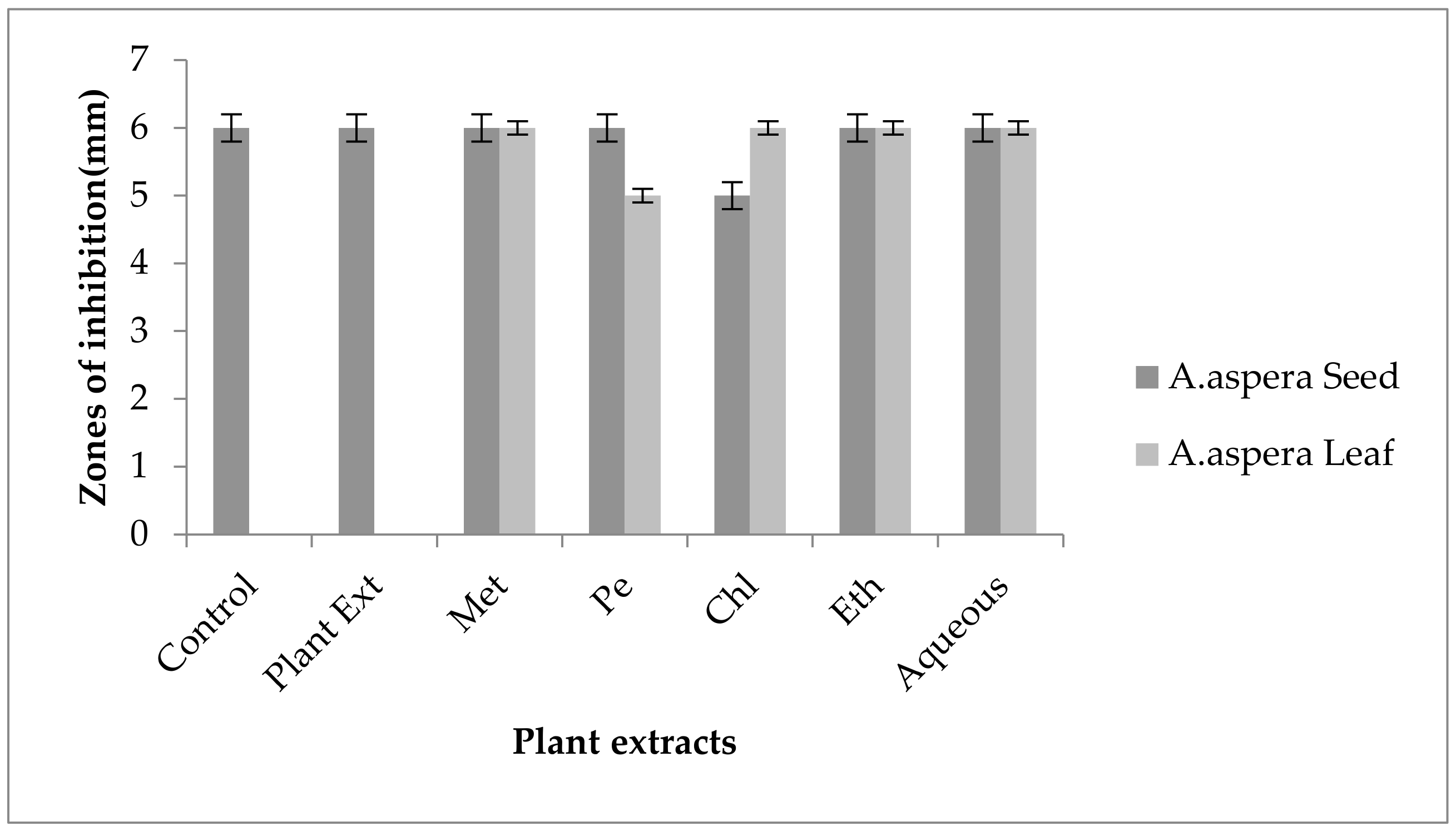
3.6. Antibiotic Resistant Redressal Activity for Acinetobacter baumannii
3.6.1. Ciprofloxacin
3.6.2. Amikacin
3.6.3. Ceftriaxone
3.6.4. Levofloxacin
3.6.5. Ceftazidime
3.6.6. Imipenem
3.6.7. Co-trimoxazole
3.7. Antibiotic Resistant Redressal Activity for Klebsiella pneumoniae
3.7.1. Ciprofloxacin
3.7.2. Amikacin
3.7.3. Ceftriaxone
3.7.4. Levofloxacin
3.7.5. Imipenem
3.7.6. Ceftazidime
3.7.7. Co-trimoxazole
3.8. Antibiotic Resistant Redressal Activity for Pseudomonas aeruginosa
3.8.1. Ciprofloxacin
3.8.2. Amikacin
3.8.3. Ceftriaxone
3.8.4. Levofloxacin
3.8.5. Imipenem
3.8.6. Co-trimoxazole
3.8.7. Ceftazidime
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carver, D.C.; Kuehn, S.B.; Weinlein, J.C. Role of Systemic and Local Antibiotics in the Treatment of Open Fractures. Orthop. Clin. N. Am. 2017, 48, 137–153. [Google Scholar] [CrossRef] [PubMed]
- Hollyer, I.; Ison, M.G. The challenge of urinary tract infections in renal transplant recipients. Transpl. Infect. Dis. 2018, 20, e12828. [Google Scholar] [CrossRef] [PubMed]
- Katz, L.; Baltz, R.H. Natural product discovery: Past, present, and future. J. Ind. Microbiol. Biotechnol. 2016, 43, 155–176. [Google Scholar] [CrossRef] [PubMed]
- Bebell, L.M.; Muiru, A.N. Antibiotic Use and Emerging Resistance: How Can Resource-Limited Countries Turn the Tide? Glob. Hear. 2014, 9, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Prescott, J.F. The resistance tsunami, antimicrobial stewardship, and the golden age of microbiology. Veter. Microbiol. 2014, 171, 273–278. [Google Scholar] [CrossRef]
- Ahmad, A.; Ghosh, A.; Schal, C.; Zurek, L. Insects in confined swine operations carry a large antibiotic resistant and potentially virulent enterococcal community. BMC Microbiol. 2011, 11, 23. [Google Scholar] [CrossRef] [Green Version]
- Silbergeld, E.K.; Graham, J.; Price, L.B. Industrial Food Animal Production, Antimicrobial Resistance, and Human Health. Annu. Rev. Public Health 2008, 29, 151–169. [Google Scholar] [CrossRef]
- Wielinga, P.R.; Schlundt, J. Combating the Risk of Antimicrobial Resistance in Animals for the Benefit of Human Health in Denmark. A Case Study of Emerging Risks Related to AMR for the International Risk Governance Council. 2012. Available online: http://www.irgc.org/wp-content/uploads/2012/04/P (accessed on 25 December 2021).
- Richet, H.M.; Mohammed, J.; McDonald, L.C.; Jarvis, W.R. Building communication networks: International network for the study and prevention of emerging antimicrobial resistance. Emerg. Infect. Dis. 2001, 7, 319. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.D.; Birdi, T.J. Development of botanicals to combat antibiotic resistance. J. Ayurveda Integr. Med. 2017, 8, 266–275. [Google Scholar] [CrossRef]
- Davies, J.; Davies, D. Origins and Evolution of Antibiotic Resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef]
- Kumar, S.; Singh, B.R. An overview of mechanisms and emergence of antimicrobials drug resistance. Adv. Anim. Vet. Sci. 2013, 1, 7–14. [Google Scholar]
- Wenzel, M.; Prochnow, P.; Mowbray, C.; Vuong, C.; Höxtermann, S.; Stepanek, J.J.; Albada, H.B.; Hall, J.; Metzler-Nolte, N.; Bandow, J.E. Towards Profiles of Resistance Development and Toxicity for the Small Cationic Hexapeptide RWRWRW-NH2. Front. Cell Dev. Biol. 2016, 4, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendelson, M. Role of antibiotic stewardship in extending the age of modern medicine. S. Afr. Med. J. 2015, 105, 414. [Google Scholar] [CrossRef] [Green Version]
- Grundmann, H.; Aires-De-Sousa, M.; Boyce, J.; Tiemersma, E. Emergence and resurgence of meticillin-resistant Staphylococcus aureus as a public-health threat. Lancet 2006, 368, 874–885. [Google Scholar] [CrossRef] [Green Version]
- Todar, K. Todar’s Online Textbook of Bacteriology Listeria monocytogenes. 2011. Available online: http://textbookofbacteriology.net (accessed on 25 December 2021).
- Bolla, J.M.; Alibert, S.; Handzlik, J.; Chevalier, J.; Mahamoud, A.; Boyer, G.; Kiec-Kononowicz, K.; Pagès, J.-M. Strategies for bypassing the membrane barrier in multidrug resistant Gram-negative bacteria. FEBS Lett. 2011, 585, 1682–1690. [Google Scholar] [CrossRef]
- McCusker, M.P.; Ferreira, D.A.; Cooney, D.; Alves, B.M.; Fanning, S.; Pagès, J.M.; Martins, M.; Davin-Regli, A. Modulation of antimicrobial resistance in clinical isolates of Enterobacter aerogenes: A strategy combining Combining antibiotics and Chemosensirizers. J. Glob. Antimicrob. Resist. 2019, 16, 187–198. [Google Scholar] [CrossRef]
- Wright, G.D. Antibiotic Adjuvants: Rescuing Antibiotics from Resistance. Trends Microbiol. 2016, 24, 862–871. [Google Scholar] [CrossRef]
- Arzanlou, M.; Chai, W.C.; Venter, H. Intrinsic, adaptive and acquired antimicrobial resistance in Gram-negative bacteria. Essays Biochem. 2017, 61, 49–59. [Google Scholar]
- Sully, E.K.; Geller, B.L. Antisense antimicrobial therapeutics. Curr. Opin. Microbiol. 2016, 33, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Pursey, E.; Sünderhauf, D.; Gaze, W.H.; Westra, E.R.; Van Houte, S. CRISPR-Cas antimicrobials: Challenges and future prospects. PLOS Pathog. 2018, 14, e1006990. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, I.; Beg, A.Z. Antimicrobial and phytochemical studies on 45 Indian medicinal plants against multi-drug resistant human pathogens. J. Ethnopharmacol. 2001, 74, 113–123. [Google Scholar] [CrossRef]
- Narayanan, A.S.; Raja, S.S.S.; Ponmurugan, K.; Kandekar, S.C.; Natarajaseenivasan, K.; Maripandi, A.; Mandeel, A.Q. Antibacterial activity of selected medicinal plants against multiple antibiotic resistant uropathogens: A study from Kolli Hills, Tamil Nadu, India. Benef. Microbes 2011, 2, 235–2433. [Google Scholar] [CrossRef] [PubMed]
- Potroz, M.G.; Cho, N.-J. Natural Products for the Treatment of Trachoma and Chlamydia trachomatis. Molecules 2015, 20, 4180–4203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, A.; Krishnamurthy, R. Swine flu and its herbal remedies. Int. J. Eng. Sci. 2013, 2, 68–78. [Google Scholar]
- Okmen, A.S. Antibacterial activities of melissa officinalis l. Extracts against various micrococcus species isolated from football player’s shoes and its antioxidant activities. Pharmacophore 2017, 8, 8–14. [Google Scholar]
- Wink, M.; Ashour, M.L.; El-Readi, M.Z. Secondary Metabolites from Plants Inhibiting ABC Transporters and Reversing Resistance of Cancer Cells and Microbes to Cytotoxic and Antimicrobial Agents. Front. Microbiol. 2012, 3, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wink, M. Evolutionary Advantage and Molecular Modes of Action of Multi-Component Mixtures Used in Phytomedicine. Curr. Drug Metab. 2008, 9, 996–1009. [Google Scholar] [CrossRef]
- Savoia, D. Plant-derived antimicrobial compounds: Alternatives to antibiotics. Future Microbiol. 2012, 7, 979–990. [Google Scholar] [CrossRef] [Green Version]
- Kumar, G.; Karthik, L.; Rao, K.V.B. Phytochemical composition and in vitro antioxidant activity of aqueous extract of Aerva lanata (L.) Juss. ex Schult. Stem (Amaranthaceae). Asian Pac. J. Trop. Med. 2013, 6, 180–187. [Google Scholar] [CrossRef] [Green Version]
- Sharma, N.; Kaushik, P. Medicinal, biological and pharmacological aspects of Plumbago zeylanica (Linn.). J. Pharmacogn. Phytochem. 2014, 3, 117–120. [Google Scholar]
- Shriram, V.; Jahagirdar, S.; Latha, C.; Kumar, V.; Dhakephalkar, P.; Rojatkar, S.; Shitole, M.G. Antibacterial & antiplasmid activities of Helicteres isora L. Indian J. Med. Res. 2010, 132, 94–99. [Google Scholar] [PubMed]
- Khder, A.K.; Muhammed, S.A. Potential of Aqueous and Alcohol Extracts of Quercus infectoria, Linusm usitatissium and Cinnamomum zeylanicium as Antimicrobials and Curing of Antibiotic Resistance in E. coli. Curr. Res. J. Biol. Sci. 2010, 2, 333–337. [Google Scholar]
- Shendkar, C.D.; Chandrachood, P.S.; Pawar, A.B.; Lavate, S.M.; Deshpande, N.R. Quantitative estimation of macro, micro nutrients and trace elements by X-ray fluorescence spectroscopy (XRF) from Achyranthes aspera Linn. Int. J. Chem. Tech. Res. 2011, 9630, 610–613. [Google Scholar]
- Yasmin, H.; Kaiser, A.; Sarker, M.R.; Rahman, M.S.; Rashid, A.M. Preliminary Anti-bacterial Activity of Some Indigenous Plants of Bangladesh. Dhaka Univ. J. Pharm. Sci. 2009, 8, 61–65. [Google Scholar] [CrossRef]
- Mushtaq, A.; Anwar, R.; Ahmad, M. Lavandula stoechas L alleviates dementia by preventing oxidative damage of cholinergic neurons in mice brain. Trop. J. Pharm. Res. 2018, 17, 1539. [Google Scholar] [CrossRef] [Green Version]
- Alastruey-Izquierdo, A.; Gómez-López, A.; Arendrup, M.C.; Lass-Florl, C.; Hope, W.W.; Perlin, D.S.; Rodriguez-Tudela, J.L.; Cuenca-Estrella, M. Comparison of Dimethyl Sulfoxide and Water as Solvents for Echinocandin Susceptibility Testing by the EUCAST Methodology. J. Clin. Microbiol. 2012, 50, 2509–2512. [Google Scholar] [CrossRef] [Green Version]
- Ismail, A.M.; Mohamed, E.A.; Marghany, M.R.; Abdel-Motaal, F.F.; Abdel-Farid, I.B.; El-Sayed, M.A. Preliminary phytochemical screening, plant growth inhibition and antimicrobial activity studies of Faidherbia albida legume extracts. J. Saudi Soc. Agric. Sci. 2016, 15, 112–117. [Google Scholar] [CrossRef]
- Nakkuntod, M.; Prapatsorn, J. Potential of thai herbal weeds for antimicrobial activity using agar disc diffusion method. Plant Cell Biotechnol. Mol. Biol. 2021, 22, 25–26. [Google Scholar]
- Sharma, V.; Chaudhary, U.; Singh, R.; Agarwal, A. Achyranthes aspera: Phytochemical estimation. Am. J. Pharmtech. Res. 2013, 3, 242–251. [Google Scholar]
- Mannas, B.; Rajesh, Y.; Kumar, V.R.; Praveen, B.; Mangamma, K. Extraction, phytochemical screening and in-vitro evaluation of anti-oxidant properties of Commicarpus chinesis (aqueous leaf extract). Int. J. Pharm. Bio. Sci. 2010, 1, 537–547. [Google Scholar]
- Ezeonu, C.S.; Ejikeme, C.M. Qualitative and Quantitative Determination of Phytochemical Contents of Indigenous Nigerian Softwoods. New J. Sci. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Khandelwal, K. Practical Pharmacognosy; Pragati Books Pvt. Ltd.: Nagpur, India, 2008. [Google Scholar]
- Abbas, M.N.; Rana, S.A.; Mahmood-Ul-Hassan, M.; Rana, N.; Iqbal, M. Phytochemical constituents of weeds: Baseline study in mixed crop zone agroecosystem. Pak. J. Weed Sci. Res. 2013, 19, 231–238. [Google Scholar]
- Sawant, R.S.; Godghate, A.G. Qualitative phytochemical screening of rhizomes of Curcuma longa Linn. Int. J. Sci. Environ. Technol. 2013, 2, 634–641. [Google Scholar]
- Shahid-Ud-Daula, A.F.M.; Basher, M.A. Phytochemical screening, plant growth inhibition, and antimicrobial activity studies of Xylocarpus granatum. Malays. J. Pharm. Sci. 2009, 7, 9–21. [Google Scholar]
- Gul, R.; Jan, S.U.; Faridullah, S.; Sherani, S.; Jahan, N. Preliminary phytochemical screening, quantitative analysis of alkaloids, and antioxidant activity of crude plant extracts from Ephedra intermedia indigenous to Balochistan. Sci. World J. 2017, 2017, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Yadav, M.; Chatterji, S.; Gupta, S.K.; Watal, G. Preliminary phytochemical screening of six medicinal plants used in traditional medicine. Int. J. Pharm. Sci. 2014, 6, 539–542. [Google Scholar]
- Bulbul, I.J.; Nahar, L.; Ripa, F.A.; Haque, O. Antibacterial, Cytotoxic and Antioxidant activity of Chloroform, n-hexane and Ethyl Acetate extract of plant Amaranthus spinosus. Int. J. Pharmtech. Res. 2011, 3, 1675–1680. [Google Scholar]
- Cheesbrough, M. District Laboratory Practice in Tropical Countries, Part 2; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Haslam, E. Natural Polyphenols (Vegetable Tannins) as Drugs: Possible Modes of Action. J. Nat. Prod. 1996, 59, 205–215. [Google Scholar] [CrossRef]
- Khanbabaee, K.; Van Ree, T. Tannins: Classification and definition. Nat. Prod. Rep. 2001, 18, 641–649. [Google Scholar]
- Kashiwada, Y.; Huang, L.; Kilkuskie, R.E.; Bodner, A.J.; Lee, K.-H. New hexahydroxydiphenyl derivatives as potent inhibitors of HIV replication in H9 lymphocytes. Bioorg. Med. Chem. Lett. 1992, 2, 235–238. [Google Scholar] [CrossRef]
- Del-Rio, A.O.B.G.; BObdululio, G.; Casfillo, J.; Marin, F.G.; Ortuno, A. Uses and properties of citrus flavonoids. J. Agric. Food Chem. 1997, 45, 4505–4515. [Google Scholar]
- Braunwald, E.; Bloodwell, R.D.; Goldberg, L.I.; Morrow, A.G. Studies on digitalis. IV. observations in man on the effects of digitalis preparations on the contractility of the non-failing heart and on total vascular resistance. J. Clin. Investig. 1961, 40, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Dillard, C.J.; German, J.B. Phytochemicals: Nutraceuticals and human health. J. Sci. Food Agric. 2000, 80, 1744–1756. [Google Scholar] [CrossRef]
- Ejikeme, C.; Ezeonu, C.S.; Eboatu, A.N. Determination of Physical and Phytochemical Constituents of some Tropical Timbers Indigenous to nigerdelta area of nigeria. Eur. Sci. J. 2014, 10, 247–270. [Google Scholar]
- Cushnie, T.T.; Cushnie, B.; Lamb, A.J. Alkaloids: An overview of their antibacterial, antibiotic-enhancing and antivirulence activities. Int. J. Antimicrob. Agents 2014, 44, 377–386. [Google Scholar] [CrossRef]
- Shi, Q.I.U.; Hui, S.U.N.; Zhang, A.H.; Hong-Ying, X.U.; Guang-Li, Y.A.N.; Ying, H.A.N.; Xi-Jun, W.A.N. Natural alkaloids: Basic aspects, biological roles, and future perspectives. Chin. J. Nat. Med. 2014, 12, 401–406. [Google Scholar]
- Rabi, T.; Bishayee, A. Terpenoids and breast cancer chemoprevention. Breast Cancer Res. Treat. 2008, 115, 223–239. [Google Scholar] [CrossRef]
- Sultana, N.; Ata, A. Oleanolic acid and related derivatives as medicinally important compounds. J. Enzym. Inhib. Med. Chem. 2008, 23, 739–756. [Google Scholar] [CrossRef]
- Shah, B.A.; Qazi, G.N.; Taneja, S.C. Boswellic acids: A group of medicinally important compounds. Nat. Prod. Rep. 2008, 26, 72–89. [Google Scholar] [CrossRef]
- Setchell, K.D.R.; Cassidy, A. Dietary Isoflavones: Biological Effects and Relevance to Human Health. J. Nutr. 1999, 129, 758S–767S. [Google Scholar] [CrossRef] [Green Version]
- Theis, N.; Lerdau, M. The Evolution of Function in Plant Secondary Metabolites. Bot. Gaz. 2003, 164, S93–S102. [Google Scholar] [CrossRef]
- Ojala, T.; Remes, S.; Haansuu, P.; Vuorela, H.; Hiltunen, R.; Haahtela, K.; Vuorela, P. Antimicrobial activity of some coumarin containing herbal plants growing in Finland. J. Ethnopharmacol. 2000, 73, 299–305. [Google Scholar] [CrossRef]
- Talreja, T.; Goswami, A.; Sharma, T. Preliminary phytochemical analysis of Achyranthes aspera and Cissus quadrangularis. Int. J. Pharmacogn. Phytochem. Res. 2016, 5, 362. [Google Scholar]
- Priya, K.; Krishnakumari, S. Phytochemical analysis of Achyranthes aspera and its activity on sesame oil induced lipid peroxidation. Anc. Sci. Life 2007, 27, 6–10. [Google Scholar] [PubMed]
- Somagari, D.R.; Basappa, K.; Rolla, S.; Jithendar, P. Phytochemical investigation of seeds of Achyranthes aspera Linn. J. Pharmacogn. Phytochem. 2014, 43, 190–193. [Google Scholar]
- Kokila, A.P.; Sarju, N.P.; Vaishali, V.C.; Chetan, R.P. Preliminary Phytochemical, Pharmacognostical and Microbial Screening of Achyranthes aspera (Amaranthaceae). J. Nat. Prod. Plant Resour. 2013, 3, 15–17. [Google Scholar]
- Sodipo, A.O.; Akinniyi, A.J.; Ogunbameru, J.V. Studies on certain characteristics of extracts of bark of Pausinystalia johimbe and Pausinystalia macroceras (K Schum) Pierre ex Beille. Glob. J. Pure Appl. Sci. 2000, 6, 83–88. [Google Scholar] [CrossRef] [Green Version]
- Performance Standards for Antimicrobial Susceptibility Testing. Available online: https://www.nih.org.pk/wp-content/uploads/2021/02/CLSI-2020.pdf (accessed on 25 December 2021).
- Ganora, L. Herbal Constituents: Foundations of Phytochemistry; Herbalchem Press: Louisville, CA, USA, 2009. [Google Scholar]
- Sethi, A.; Sharma, R.A. Antioxidant activity with total phenolic constituents from Aerva tomentosa Forsk. Int. J. Pharm. Bio. Sci. 2011, 2, 596–603. [Google Scholar]
- Al-Saiym, R.A.; Al-Kamali, H.H.; Al-Magboul, A.Z. Synergistic Antibacterial Interaction between Trachyspermum ammi, Senna alexandrina Mill and Vachellia nilotica spp. Nilotica Extract and Antibiotics. Pak. J. Biol. Sci. 2015, 18, 115–121. [Google Scholar] [CrossRef] [Green Version]
- O’Jensen, S.; Lyon, B.R. Genetics of antimicrobial resistance in Staphylococcus aureus. Future Microbiol. 2009, 4, 565–582. [Google Scholar]
- Ambrose, K.D.; Nisbet, R.; Stephens, D.S. Macrolide Efflux in Streptococcus pneumoniae Is Mediated by a Dual Efflux Pump (mel and mef) and Is Erythromycin Inducible. Antimicrob. Agents Chemother. 2005, 49, 4203–4209. [Google Scholar] [CrossRef] [Green Version]
- Moore, D. Antibiotic Classification & Mechanism. Available online: https://www.orthobullets.com/basic-science/9059/antibiotic-classification-and-mechanism (accessed on 25 December 2021).
- Rehman, A.; Patrick, W.M.; Lamont, I.L. Mechanisms of ciprofloxacin resistance in Pseudomonas aeruginosa: New approaches to an old problem. J. Med. Microbiol. 2019, 68, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, M.S.; Tolmasky, M.E. Amikacin: Uses, Resistance, and Prospects for Inhibition. Molecules 2017, 22, 2267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skov, R.; Larsen, A.R.; Kearns, A.; Holmes, M.; Teale, C.; Edwards, G.; Hill, R. Phenotypic detection of mecC-MRSA: Cefoxitin is more reliable than oxacillin. J. Antimicrob. Chemother. 2014, 69, 133–135. [Google Scholar] [CrossRef] [PubMed]
- Venter, H. Reversing resistance to counter antimicrobial resistance in the World Health Organisation’s critical priority of most dangerous pathogens. Biosci. Rep. 2019, 39, BSR20180474. [Google Scholar] [CrossRef] [Green Version]
- Thakuria, B.; Lahon, K. The beta lactam antibiotics as an empirical therapy in a developing country: An update on their current status and recommendations to counter the resistance against them. J. Clin. Diagn. Res. 2013, 7, 1207. [Google Scholar] [CrossRef]
- Bachoon, D.S.; Dustman, W.A. Exercise 8: Selective and Differential Media for Isolation; Microbiological Laboratory Manual: Mason, OH, USA, 2008. [Google Scholar]
- Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance. Microbiol. Spectrum 2016, 4, 4.2.15. [Google Scholar] [CrossRef] [Green Version]
- Lowy, F.D. Antimicrobial resistance: The example of Staphylococcus aureus. J. Clin. Investig. 2003, 111, 1265–1273. [Google Scholar] [CrossRef]
- Shore, A.C.; Lazaris, A.; Kinnevey, P.M.; Brennan, O.M.; Brennan, G.I.; O’Connell, B.; Feßler, A.T.; Schwarz, S.; Coleman, D.C. First report of cfr-carrying plasmids in the pandemic sequence type 22 methicillin-resistant Staphylococcus aureus staphylococcal cassette chromosome mec type IV clone. Antimicrob. Agents Chemother. 2016, 60, 3007–3015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moo, C.L.; Yang, S.-K.; Yusoff, K.; Ajat, M.; Thomas, W.; Abushelaibi, A.; Lim, S.-H.-E.; Lai, K.-S. Mechanisms of Antimicrobial Resistance (AMR) and Alternative Approaches to Overcome AMR. Curr. Drug Discov. Technol. 2020, 17, 430–447. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, E.; Bishara, J. Contemporary unconventional clinical use of co-trimoxazole. Clin. Microbiol. Infect. 2012, 18, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, Y.; Ishii, M.; Hasegawa, S.; Sekimizu, K. Enterococcus faecalis YM0831 suppresses sucrose-induced hyperglycemia in a silkworm model and in humans. Commun. Biol. 2019, 2, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.-H.; Cohen, T.; Grad, Y.H.; Hanage, W.P.; O’Brien, T.F.; Lipsitch, M. Origin and Proliferation of Multiple-Drug Resistance in Bacterial Pathogens. Microbiol. Mol. Biol. Rev. 2015, 79, 101–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selim, S. Mechanisms of gram-positive vancomycin resistance (Review). Biomed. Rep. 2022, 16, 7. [Google Scholar] [CrossRef]
- Miller, W.R.; Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance in enterococci. Expert Rev. Anti-Infect. Ther. 2014, 12, 1221–1236. [Google Scholar] [CrossRef] [PubMed]
- Papich, M.G. Saunders Handbook of Veterinary Drugs; Saunders Elsevier: St. Louis, MO, USA, 2007. [Google Scholar]
- Lee, C.-R.; Lee, J.H.; Park, M.; Park, K.S.; Bae, I.K.; Kim, Y.B.; Cha, C.-J.; Jeong, B.C.; Lee, S.H. Biology of Acinetobacter baumannii: Pathogenesis, Antibiotic Resistance Mechanisms, and Prospective Treatment Options. Front. Cell. Infect. Microbiol. 2017, 7, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, V.B.; Rajamohan, G.; Gebreyes, W.A. Role of AbeS, a novel efflux pump of the SMR family of transporters, in resistance to antimicrobial agents in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2009, 53, 5312–5316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallego, L.; Towner, K.J. Carriage of class 1 integrons and antibiotic resistance in clinical isolates of Acinetobacter baumannii from Northern Spain. J. Med. Microbiol. 2001, 50, 71–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranjani, S.; Begum, I.F.; Santhoshini, J.; Kumar, N.S.; Ruckmani, K.; Hemalatha, S. Mimosa pudica floral nanoparticles: A potent antibiotic resistance breaker. Inorg. Nano-Met. Chem. 2020, 2020, 429. [Google Scholar]
- Foye, W.O. Foye’s Principles of Medicinal Chemistry; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2008. [Google Scholar]
- Nagano, N.; Nagano, Y.; Cordevant, C.; Shibata, N.; Arakawa, Y. Nosocomial Transmission of CTX-M-2 β-Lactamase-Producing Acinetobacter baumannii in a Neurosurgery Ward. J. Clin. Microbiol. 2004, 42, 3978–3984. [Google Scholar] [CrossRef] [Green Version]
- Kolayli, F.; Gacar, G.; Karadenizli, A.; Sanic, A.; Vahaboglu, H.; Study Group. PER-1 is still widespread in Turkish hospitals among Pseudomonas aeruginosa and Acinetobacter spp. FEMS Microbiol. Lett. 2005, 249, 241–245. [Google Scholar] [CrossRef] [Green Version]
- Turton, J.F.; Woodford, N.; Glover, J.; Yarde, S.; Kaufmann, M.E.; Pitt, T.L. Identification of Acinetobacter baumannii by Detection of the bla OXA-51-like Carbapenemase Gene Intrinsic to This Species. J. Clin. Microbiol. 2006, 44, 2974–2976. [Google Scholar] [CrossRef] [Green Version]
- Magnet, S.; Courvalin, P.; Lambert, T. Resistance-Nodulation-Cell Division-Type Efflux Pump Involved in Aminoglycoside Resistance in Acinetobacter baumannii Strain BM4454. Antimicrob. Agents Chemother. 2001, 45, 3375–3380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Weinjie, Y.; Ping, L.; Na, W.; Haixia, R.; Xuequn, Z. Antibiotic resistance of Klebsiella pneumoniae through β-arrestin recruitment-induced β- lactamase signaling pathway. Exp. Therapeut. Med. 2018, 15, 2247–2254. [Google Scholar]
- Bhatnagar, K.; Hinz, A.; Kohlman, M.; Wong, A. An sRNA Screen for Reversal of Quinolone Resistance in Escherichia coli. G3 Genes|Genomes|Genet. 2020, 10, 79–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas, L.J.; Marshall, S.; Yasmin, M.; Rudin, S.D.; Perez, F.; Donskey, C.; Hecker, M.; A Bonomo, R. 234. Reversal of Carbapenem and Amikacin Susceptibilities in Isogenic Klebsiella pneumoniae From a Patient with Persistent Bacteriuria. Open Forum Infect. Dis. 2019, 6, 134. [Google Scholar] [CrossRef]
- Wekesa, Y.N.; Namusoke, F.; Sekikubo, M.; Mango, D.W.; Bwanga, F. Ceftriaxone- and ceftazidime-resistant Klebsiella species, Escherichia coli, and methicillin-resistant Staphylococcus aureus dominate caesarean surgical site infections at Mulago Hospital, Kampala, Uganda. SAGE Open Med. 2020, 8, 1–12. [Google Scholar] [CrossRef]
- Papa, A.; Dalla, V.; Papadimitriou, E.; Kartalis, G.N.; Antoniadis, A. Emergence of Crimean–Congo haemorrhagic fever in Greece. Clin. Microbiol. Infect. 2010, 16, 843–847. [Google Scholar] [CrossRef] [Green Version]
- Patilaya, P.; Husori, D.I.; Marhafanny, L. Susceptibility of Klebsiella Pneumoniae Isolated from Pus Specimens of Post-Surgery Patients in Medan, Indonesia to Selected Antibiotics. Open Access Maced. J. Med. Sci. 2019, 7, 3861–3864. [Google Scholar] [CrossRef] [Green Version]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- Dupont, M.; Pagès, J.M.; Lafitte, D.; Siroy, A.; Bollet, C. Identification of an OprD Homologue in Acinetobacter baumannii. J. Proteome Res. 2005, 4, 2386–2390. [Google Scholar] [CrossRef] [PubMed]





































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tests for | Extracts of Achyranthes aspera (Seeds) | ||||
|---|---|---|---|---|---|
| Methanolic Extract | Petroleum Ether Extract | Chloroform Extract | Ethyl Acetate Extract | Aqueous Extract | |
| Alkaloids | |||||
| • Wagner’s test | ++ | - | - | +++ | - |
| • Mayer’s test | + | - | - | ++ | - |
| • Tannic acid test | - | - | ++ | - | ++ |
| Carbohydrates | - | ||||
| • Barfoed’s test | - | - | - | ++ | - |
| • Fehling test | + | - | ++ | +++ | + |
| • Molish test | ++ | ++ | ++ | ++ | ++ |
| Glycosides | |||||
| • Cardiac test | - | +++ | +++ | ++ | - |
| • Legal’s test | - | ++ | +++ | - | ++ |
| Proteins and amino acids | |||||
| • Biuret test | - | - | - | - | - |
| • Ninhydrin test | - | +++ | - | - | ++ |
| Fixed oils and fat | |||||
| • Spot test | - | - | - | +++ | +++ |
| Phenolics | |||||
| • Ferric chloride test | + | - | - | ++ | + |
| Tannins | |||||
| • Gelatin test | ++ | - | - | - | - |
| Terpenes | ++ | ++ | + | ++ | +++ |
| Flavonoids | - | - | - | - | - |
| Saponins | ++ | - | +++ | ++ | - |
| Steroids | |||||
| • Liberman–Burchard reaction | ++ | +++ | +++ | +++ | ++ |
| Anthraquinones | - | - | - | - | - |
| Coumarin | +++ | ++ | ++ | +++ | + |
| Phlobatannins | +++ | - | - | - | - |
| Emodins | - | - | - | - | - |
| Anthocyanins | - | - | - | - | - |
| Tests for | Extracts of Achyranthes aspera (Leaves) | ||||
|---|---|---|---|---|---|
| Methanolic Extract | Petroleum Ether Extract | Chloroform Extract | Ethyl Acetate Extract | Aqueous Extract | |
| Alkaloids | |||||
| • Wagner’s test | - | ++ | - | - | - |
| • Mayer’s test | - | - | - | - | - |
| • Tannic acid test | + | ++ | ++ | - | - |
| Carbohydrates | |||||
| • Barfoed’s test | - | - | - | - | - |
| • Fehling test | ++ | + | +++ | + | +++ |
| • Molish test | - | - | ++ | ++ | - |
| Glycosides | |||||
| • Cardiac test | - | - | - | +++ | ++ |
| • Legal’s test | - | - | - | +++ | - |
| Proteins and amino acids | |||||
| • Biuret test | - | - | - | - | - |
| • Ninhydrin test | - | - | - | - | - |
| Fixed oils and fat | |||||
| • Spot test | ++ | +++ | - | - | + |
| Phenolics | |||||
| • Ferric chloride test | ++ | - | ++ | ++ | ++ |
| Tannins | |||||
| • Gelatin test | ++ | - | - | - | - |
| Terpenes | ++ | + | ++ | +++ | +++ |
| Flavonoids | - | - | - | - | - |
| Saponins | +++ | - | - | - | ++ |
| Steroids | |||||
| • Liberman–Burchard reaction | - | - | - | ++ | ++ |
| Anthraquinones | - | - | - | - | - |
| Coumarins | - | ++ | ++ | +++ | + |
| Phlobatannins | - | - | - | - | - |
| Emodins | - | - | - | - | - |
| Anthocyanins | - | - | - | - | - |
| SR# | Antibiotics Used for Staphylococcus aureus | Zones of Inhibition for Staphylococcus aureus (mm) | Zones of Inhibition Sensitive CLSI (mm) | Zones of Inhibition Intermediate CLSI (mm) | Zones of Inhibition Resistant CLSI (mm) |
|---|---|---|---|---|---|
| 1 | Ciprofloxacin (5 µg) | 21 ± 0.1 | ≥21 | 16–20 | ≤15 |
| 2 | Levofloxacin (5 µg) | 14 ± 0.3 | ≥15 | 13–14 | ≤12 |
| 3 | Amikacin (30 µg) | 17 ± 0.2 | ≥18 | 14–17 | ≤13 |
| 4 | Cefoxitin (30 µg) | 17 ± 0.1 | ≥22 | - | ≤21 |
| 5 | Penicillin (10 µg) | 6 ± 0.5 | ≥15 | 13–14 | ≤12 |
| 6 | Linezolid (10 µg) | 24 ± 0.2 | ≥21 | - | ≤20 |
| 7 | Co-trimoxazole (25 µg) | 6 ± 0.4 | ≥16 | 11–15 | ≤10 |
| SR# | Antibiotics Used for Enterococcus faecalis | Zones of Inhibition for Enterococcus faecalis (mm) | Zones of Inhibition Sensitive CLSI (mm) | Zones of Inhibition Intermediate CLSI (mm) | Zones of Inhibition Resistant CLSI (mm) |
|---|---|---|---|---|---|
| 1 | Ciprofloxacin (5 µg) | 13 ± 0.2 | ≥21 | 16–20 | ≤15 |
| 2 | Levofloxacin (5 µg) | 10 ± 0.4 | ≥17 | 14–16 | ≤13 |
| 3 | Penicillin (10 µg) | 8 ± 0.5 | ≥15 | 13–14 | ≤12 |
| 4 | Amoxicillin (10 µg) | 6 ± 0.1 | ≥18 | 14–17 | ≤13 |
| 5 | Linezolid (30µg) | 19 ± 0.3 | ≥23 | 21–22 | ≤20 |
| 6 | Imipenem (10 µg) | 14 ± 0.2 | ≥21 | 19–21 | ≤18 |
| 7 | Vancomycin (30 µg) | 10 ± 0.1 | ≥17 | 15–16 | ≤14 |
| SR# | Antibiotics Used for Acinetobacter baumannii | Zones of Inhibition for Acinetobacter baumannii (mm) | Zones of Inhibition Sensitive CLSI (mm) | Zones of Inhibition Intermediate CLSI (mm) | Zones of Inhibition Resistant CLSI (mm) |
|---|---|---|---|---|---|
| 1 | Ciprofloxacin (5 µg) | 6 ± 0.3 | ≥26 | 22–25 | ≤21 |
| 2 | Levofloxacin (5 µg) | 9 ± 0.1 | ≥21 | 17–20 | ≤16 |
| 3 | Amikacin (30 µg) | 18 ± 0.3 | ≥18 | 14–17 | ≤13 |
| 4 | Ceftriaxone (30 µg) | 6 ± 0.2 | ≥23 | 20–22 | ≤19 |
| 5 | Ceftazidime (30µg) | 6 ± 0.4 | ≥21 | 18–20 | ≤17 |
| 6 | Imipenem (10 µg) | 6 ± 0.3 | ≥23 | 20–22 | ≤19 |
| 7 | Co-trimoxazole (25 µg) | 19 ± 0.1 | ≥16 | 11–15 | ≤10 |
| SR# | Antibiotics Used for Klebsiella pneumoniae | Zones of Inhibition for Klebsiella pneumoniae (mm) | Zones of Inhibition Sensitive CLSI (mm) | Zones of Inhibition Intermediate CLSI (mm) | Zones of Inhibition Resistant CLSI (mm) |
|---|---|---|---|---|---|
| 1 | Ciprofloxacin (5 µg) | 6 ± 0.4 | ≥26 | 22–25 | ≤21 |
| 2 | Levofloxacin (5 µg) | 6 ± 0.3 | ≥21 | 17–20 | ≤16 |
| 3 | Amikacin (30 µg) | 6 ± 0.2 | ≥18 | 14–17 | ≤13 |
| 4 | Ceftriaxone (30 µg) | 6 ± 0.3 | ≥23 | 20–22 | ≤19 |
| 5 | Ceftazidime (30 µg) | 8 ± 0.1 | ≥21 | 18–20 | ≤17 |
| 6 | Imipenem (10 µg) | 7 ± 0.1 | ≥23 | 20–22 | ≤19 |
| 7 | Co-trimoxazole (25 µg) | 6 ± 0.2 | ≥16 | 11–15 | ≤10 |
| SR# | Antibiotics Used for Pseudomonas Aeruginosa | Zones of Inhibition for Pseudomonas aeruginosa (mm) | Zones of Inhibition Sensitive CSLI (mm) | Zones of Inhibition Intermediate CSLI (mm) | Zones of Inhibition Resistant CSLI (mm) |
|---|---|---|---|---|---|
| 1 | Ciprofloxacin (5 µg) | 6 ± 0.4 | ≥26 | 22–25 | ≤21 |
| 2 | Levofloxacin (5 µg) | 6 ± 0.3 | ≥21 | 17–20 | ≤16 |
| 3 | Amikacin (30 µg) | 6 ± 0.2 | ≥18 | 14–17 | ≤13 |
| 4 | Ceftriaxone (30 µg) | 6 ± 0.3 | ≥23 | 20–22 | ≤19 |
| 5 | Ceftazidime (30µg) | 6 ± 0.1 | ≥21 | 18–20 | ≤17 |
| 6 | Imipenem (10 µg) | 6 ± 0.1 | ≥23 | 20–22 | ≤19 |
| 7 | Co-trimoxazole (25 µg) | 6 ± 0.2 | ≥16 | 11–15 | ≤10 |
| Achyranthes aspera Seed Extracts | Zone of Inhibition MRSA (mm) | Zone of Inhibition E. faecalis (mm) | Zone of Inhibition A. baumannii (mm) | Zone of Inhibition K. pneumoniae (mm) | Zone of Inhibition P. aeruginosa (mm) |
|---|---|---|---|---|---|
| Methanolic extract | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 |
| Petroleum ether extract | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 |
| Chloroform extract | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 |
| Ethyl acetate extract | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 |
| Aqueous extract | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 |
| Achyranthes aspera Leaf Extracts | Zone of Inhibition MRSA (mm) | Zone of Inhibition E. faecalis (mm) | Zone of Inhibition A. baumannii (mm) | Zone of Inhibition K. pneumoniae (mm) | Zone of Inhibition P. aeruginosa (mm) |
|---|---|---|---|---|---|
| Methanolic extract | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 |
| Petroleum ether extract | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 |
| Chloroform extract | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 |
| Ethyl acetate extract | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 |
| Aqueous extract | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 | 6 ± 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, H.; Gohar, U.F.; Mukhtar, H.; Zia-UI-Haq, M.; Marc, R.A.; Irimie, M.; Marceanu, L.G.; Gavris, C.M. Achyranthes aspera Extracts as Adjuvants for the Redressal of Antibiotic Resistance. Pharmaceutics 2022, 14, 2219. https://doi.org/10.3390/pharmaceutics14102219
Ahmad H, Gohar UF, Mukhtar H, Zia-UI-Haq M, Marc RA, Irimie M, Marceanu LG, Gavris CM. Achyranthes aspera Extracts as Adjuvants for the Redressal of Antibiotic Resistance. Pharmaceutics. 2022; 14(10):2219. https://doi.org/10.3390/pharmaceutics14102219
Chicago/Turabian StyleAhmad, Hamna, Umar Farooq Gohar, Hamid Mukhtar, Muhammad Zia-UI-Haq, Romina Alina Marc, Marius Irimie, Luigi Geo Marceanu, and Claudia Mihaela Gavris. 2022. "Achyranthes aspera Extracts as Adjuvants for the Redressal of Antibiotic Resistance" Pharmaceutics 14, no. 10: 2219. https://doi.org/10.3390/pharmaceutics14102219