
Biomimetic Nanovesicles—Sources, Design, Production Methods, and Applications
 , , ,
, , ,  and
and
Abstract
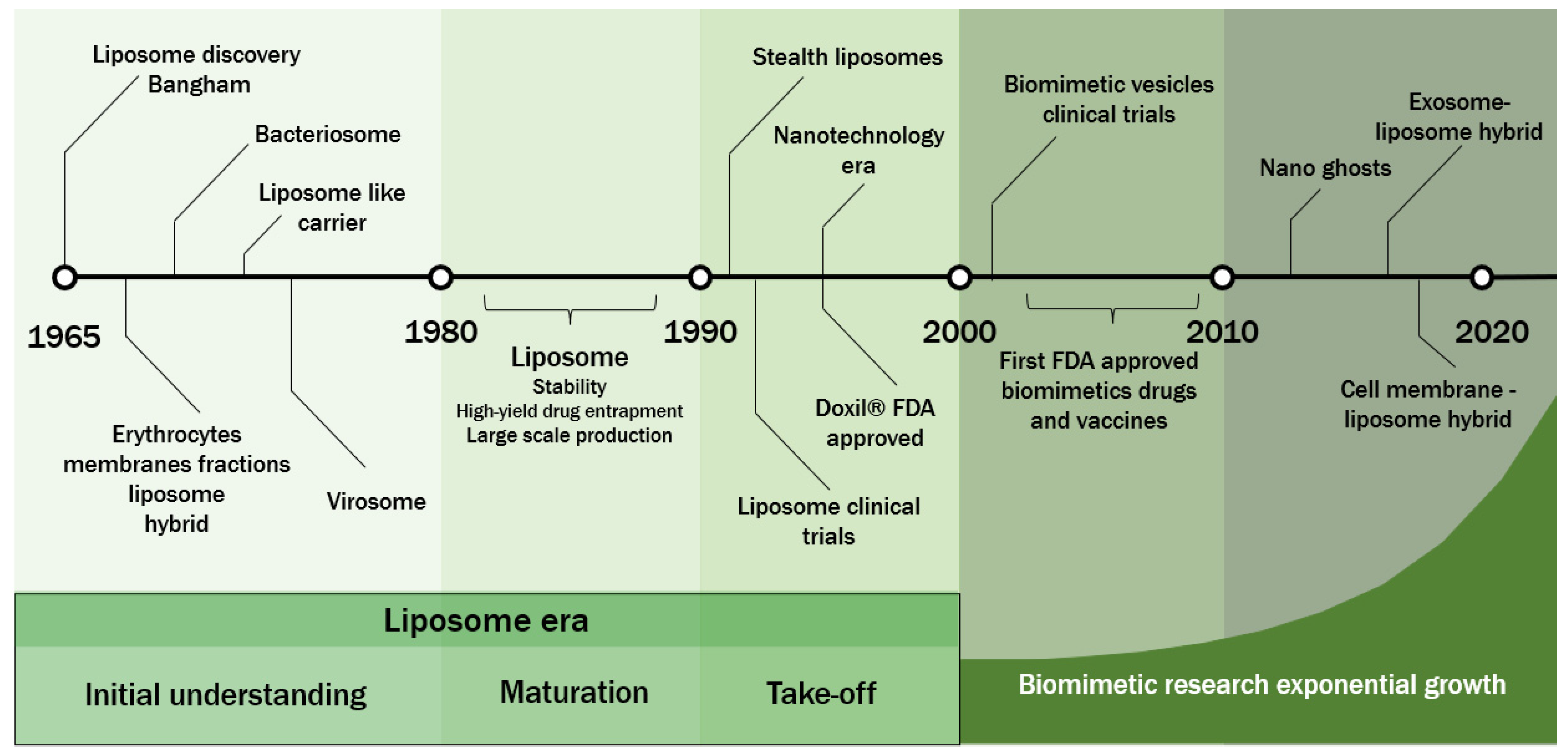
:1. Introduction
2. Natural Membrane Sources
2.1. Virus
2.2. Outer Membrane Vesicles (OMV)
2.3. Bacterial Membrane (Bacterial Ghosts or Bacteriosomes)
2.4. Extracellular Vesicles (EV) from Mammalian Cells
2.5. Mammalian Cell Membranes
3. Design Strategies for Biomimetic Vesicles
3.1. Liposomes Containing Isolated Protein from Cell Membranes
3.2. Native Cell Membranes as a Delivery System

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nanostructure | Membrane Source | Targeting | Important Features | Refs. |
|---|---|---|---|---|
| Nanosized native cell membrane | Mesenchymal stem cells (MSCs) membrane (hypotonic lysis) | Cancer | Requires large number of cells (compare to the hybrid nanovesicle approach) | [48,49,67] |
| Expensive method relative to membrane source | ||||
| The delivery system for cancer treatment can circumvent resistance to therapy | ||||
| Ability to cross biological barriers | ||||
| Native membrane cloaked nanoparticle | Platelet membrane (freeze–thaw process) | Atherosclerosis | Good system evasion (increased adhesion of platelets with endothelial cells) | [50,68,69,70] |
| Less complex extraction protocol (anucleated cells) | ||||
| Requires large number of cells (compare to the hybrid nanovesicle approach) | ||||
| Target for antithrombosis drug delivery | ||||
| Good immune evasion of platelet membranes | ||||
| Monocyte cell membrane (hypotonic lysis) | Cancer | Ability to retain biological targeting properties of the cell of origin | [67] | |
| Higher cellular uptake in cancer cells | ||||
| Good targeting for cancer cells | ||||
| Ability to make intercellular homologous bonds with membrane proteins | ||||
| Good targeting for cancer cells | ||||
| Cancer cell membrane coating improved; sustained release of chemotherapeutic and decreased cellular uptake of macrophages | ||||
| Biomimetic vesicles with isolated proteins (proteolipid) | Leukocyte plasma membrane proteins | Inflammation target | Referred to as leukosomes by the authors | [60,71] |
| Can bypass macrophages (increased circulation) | ||||
| Difficulty obtaining (needs more blood to obtain) | ||||
| Lack of targeting for tumor targeting | ||||
| Compared to conventional liposomes, leukosomes preferentially recognized the inflamed endothelium | ||||
| Genetically engineered hPSC differentiating in neurons plasma membrane proteins | Neurodegeneration | Referred to as neurosomes by the authors | [62] | |
| Ability to mediate protein-protein interactions with target cells | ||||
| Bottom-up microfluidics-based synthesis method to make nanovesicles | ||||
| The ability to circulate in the blood or cross the blood–brain barrier (BBB) has not been evaluated | ||||
| Hybrid nanovesicle Membrane | Platelet membrane (Freeze–thaw process) | Atherosclerosis | Requires less amount of cells compared to coating (low cost) | [50,70] |
| Good targeting of inflammation cells, immune evasion of platelet membranes, biocompatibility and biodegradability when fusing with liposomes, prolonged circulation and ability to encapsulate and release components, | ||||
| Mesenchymal stem cells (MSCs) membrane (Freeze–thaw) | Inflammation tropism to ischemic cerebrum | MSCs can communicate with immune cells like phagocytes and microglia; ability to cross the blood–brain barrier (BBB); prolonged circulation and ability to encapsulate and release components | [55,72] | |
| Ability to reduce uptake by the mononuclear phagocytic system | ||||
| Exosomes (MSC-derived) | Cancer | Good result of polyethylene glycol (PEG)-mediated fusion of cell-derived EVs with liposomes bringing biocompatible compositions | [73] | |
| Can add biogenic functionalities to the liposome for enhanced cellular absorption | ||||
| Promotes increased circulation and improved targeting of permeability and retention | ||||
| Exosomes (macrophage-derived) | Cancer | Good drug delivery to cancer cells under acidic conditions | [74] | |
| Ability to target sick/inflammatory cells due to surface proteins acquired by EV | ||||
| Nanovesicle surface proteins acquired by EV play a significant role in intercellular adhesion | ||||
| Genetically engineered exosomes (CD47-expressed) | Cancer | Able to penetrate the target tumor and release loads under hypothermic conditions of HIPEC after intravenous administration | [75] | |
| May surpass the efficiency of delivering chemotherapeutic drugs to large tumor nodules | ||||
| Two hybrid nanovesicle membranes | Leukocyte–tumor cell plasma membrane (hypotonic lysis) | Cancer | Increased affinity and uptake with tumor cells | [54] |
| Prolonged blood circulation | ||||
| Multiple biofunction for drug delivery | ||||
| High loading capacity for poorly water-soluble anticancer drugs. | ||||
| Two native membrane-cloaked nanoparticles | Erythrocyte (hypotonic lysis) -platelet (freeze–thaw) plasma membrane | Blood circulation | Ability to load features from both source cells | [65,76,77] |
| Circulation for long periods of time | ||||
| Presents immunomodulatory markers, such as CD47, with density similar to original red blood cells |
3.3. Liposome and Native Membrane Hybrid Nanostructure
4. Biomimetic Nanovesicle Production Methods
4.1. Strategy—Top-Down/Bottom-UpTop-Down and Bottom-Up
4.1.1. Top-Down Method
4.1.2. Bottom-Up Method
4.2. Physical Methods
4.2.1. Freeze and Thaw
4.2.2. Sonication
4.2.3. Extrusion
4.2.4. Incubation
4.3. Chemical Methods
4.3.1. Polyethylene Glycol
4.3.2. DOPE—1,2-Dioleoyl-sn-Glycero-3-Phosphoethanolamine
4.3.3. Calcium–Mediate Fusion
4.4. Continuous Flow Methods
Microfluidics
5. Applications for Drug and Gene Delivery, and Vaccine
5.1. Cancer Therapy
5.2. Gene Delivery
5.3. Brain Delivery
5.4. Vaccines
6. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ApoE | Apolipoprotein E |
| BBB | Blood–brain barrier |
| BG | Bacterial ghost |
| BNV | Biomimetic Nanovesicles |
| CDV | Cell-delivered plasma membrane vesicles |
| COVID-19 | Coronavirus Disease 2019 |
| DAP | 1,2-dipalmitoyl-3-dimethylammonium-propane |
| DNA | Deoxyribonucleic acid |
| DOP | 1,2-dioleoyl-sn-glycero-3-phospho-L-serine (DOPS) |
| DOPC | 1,2-dioleoyl-sn-glycero-3 -phosphocholine |
| DOPE | Dioleoylphosphatidylethanolamine |
| Dox | Doxorubicin |
| DOXIL | Doxorubicin (liposomal) |
| DPPC | Dipalmitoylphosphatidylcholine |
| DSPC | 1,2-Distearoyl-sn-glycero-3-phosphocholine |
| DSPE-PEG2000 | 1,2-distearoyl-sn-glycero-3-phosphoethanolamine-N-[amino(polyethylene glycol)-2000] |
| EPC | Egg Phosphatidylcholine (1,2-dioleoyl-sn-glycero-3-ethylphosphocholine) |
| ESEs | Early sorting endosomes |
| EV | Extracellular vesicles |
| FDA | Food and Drug Administration |
| FRET | Förster resonance energy transfer |
| HIV | Human immunodeficiency virus |
| hPSCs | Human pluripotent stem cells |
| HPV16 | Human papillomavirus 16 |
| HSV-1 | Herpes simplex virus type 1 |
| ICG | Indocyanine green |
| IGg | Immunoglobin G |
| ILVs | Intraluminal vesicles |
| LPS | Lipopolysaccharide |
| LSE | Late sorting endosomes |
| MAP | Model amphiphilic peptide |
| MSCs | Mesenchymal stem cells membrane |
| MVB | Multivesicular bodies |
| NP | Nanoparticle |
| OMV | Outer Membrane Vesicles |
| PC S100 | Soya Phosphatidylcholine |
| PE | Phosphatidyl ethanolamine |
| PEG | Polyethylene glycol |
| PG | Phosphatidyl glycerol |
| PLGA | Poly(DL-lactide-co-glycolide) |
| PNP | PLGA nanoparticles |
| POPC | 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine |
| PPG | Peptidoglycan |
| PTX | Paclitaxel |
| RNA | Ribonucleic acid |
| sEVs | Small extracellular vesicles |
| VLP | Virus-like particles |
References
- Gregoriadis, G. Liposomes in Drug Delivery: How It All Happened. Pharmaceutics 2016, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Bangham, A.D.; Standish, M.M.; Watkins, J.C. Diffusion of univalent ions across the lamellae of swollen phospholipids. J. Mol. Biol. 1965, 13, 238–252. [Google Scholar] [CrossRef]
- Bangham, A.D. Surrogate cells or trojan horses. The discovery of liposomes. BioEssays 1995, 17, 1081–1088. [Google Scholar] [CrossRef] [PubMed]
- de La Torre, L.G.; de Pinho, S.C. Lipid Matrices for Nanoencapsulation in Food: Liposomes and Lipid Nanoparticles. In Food Nanoscience and Nanotechnology; Food Engineering Series; Springer: Cham, Switzeland, 2015; pp. 99–143. [Google Scholar] [CrossRef]
- Almeida, B.; Nag, O.K.; Rogers, K.E.; Delehanty, J.B. Recent Progress in Bioconjugation Strategies for Liposome-Mediated Drug Delivery. Molecules 2020, 25, 5672. [Google Scholar] [CrossRef]
- Alving, C.R.; Kinsky, S.C.; Haxby, J.A.; Kinsky, C.B. Antibody binding and complement fixation by a liposomal model membrane. Biochemistry 1969, 8, 1582–1587. [Google Scholar] [CrossRef]
- Kataoka, T.; Inoue, K.; Luderitz, O.; Kinsky, S.C. Antibody- and Complement-Dependent Damage to Liposomes Prepared with Bacterial Lipopolysaccharides. JBIC J. Biol. Inorg. Chem. 1971, 21, 80–85. [Google Scholar] [CrossRef]
- Kataoka, T.; Inoue, K.; Galanos, C.; Kinsky, S.C. Detection and Specificity of Lipid A Antibodies Using Liposomes Sensitized with Lipid A and Bacterial Lipopolysaccharides. JBIC J. Biol. Inorg. Chem. 1971, 24, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Almeida, J.; Edwards, D.; Brand, C.; Heath, T. Formation of Virosomes from Influenza Subunits and Liposomes. Lancet 1975, 306, 899–901. [Google Scholar] [CrossRef]
- Sharma, V.K.; Agrawal, M.K. A historical perspective of liposomes-a bio nanomaterial. Mater. Today Proc. 2021, 45, 2963–2966. [Google Scholar] [CrossRef]
- Gregoriadis, G. Liposome research in drug delivery: The early days. J. Drug Target. 2008, 16, 520–524. [Google Scholar] [CrossRef]
- Lee, J.; Kim, O.Y.; Gho, Y.S. Proteomic profiling of Gram-negative bacterial outer membrane vesicles: Current perspectives. Proteom. Clin. Appl. 2016, 10, 897–909. [Google Scholar] [CrossRef]
- Garland, S.M.; Hernandez-Avila, M.; Wheeler, C.M.; Perez, G.; Harper, D.M.; Leodolter, S.; Tang, G.W.; Ferris, D.G.; Steben, M.; Bryan, J.; et al. Quadrivalent Vaccine against Human Papillomavirus to Prevent Anogenital Diseases. N. Engl. J. Med. 2007, 356, 1928–1943. [Google Scholar] [CrossRef] [PubMed]
- Leroux-Roels, G. Unmet needs in modern vaccinology: Adjuvants to improve the immune response. Vaccine 2010, 28, C25–C36. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Hsiao, S.C.; Carrico, Z.M.; Francis, M.B. Genome-Free Viral Capsids as Multivalent Carriers for Taxol Delivery. Angew. Chem. Int. Ed. 2009, 48, 9493–9497. [Google Scholar] [CrossRef] [PubMed]
- Waelti, E.; Wegmann, N.; Schwaninger, R.; Wetterwald, A.; Wingenfeld, C.; Rothen-Rutishauser, B.; Gimmi, C.D. Targeting HER-2/Neu with Antirat Neu Virosomes for Cancer Therapy. Cancer Res. 2002, 62, 437–444. [Google Scholar]
- Yoo, J.-W.; Irvine, D.J.; Discher, D.E.; Mitragotri, S. Bio-inspired, bioengineered and biomimetic drug delivery carriers. Nat. Rev. Drug Discov. 2011, 10, 521–535. [Google Scholar] [CrossRef] [PubMed]
- de Lima, P.H.C.; Butera, A.P.; Cabeça, L.F.; Ribeiro-Viana, R.M. Liposome surface modification by phospholipid chemical reactions. Chem. Phys. Lipids 2021, 237, 105084. [Google Scholar] [CrossRef]
- Cao, Y.; Dong, X.; Chen, X. Polymer-Modified Liposomes for Drug Delivery: From Fundamentals to Applications. Pharmaceutics 2022, 14, 778. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Chen, L.; Li, K.; Lou, B.; Liu, Y.; Liu, Z. Yeast as carrier for drug delivery and vaccine construction. J. Control. Release 2022, 346, 358–379. [Google Scholar] [CrossRef]
- Zhou, J.; Krishnan, N.; Jiang, Y.; Fang, R.H.; Zhang, L. Nanotechnology for virus treatment. Nano Today 2020, 36, 101031. [Google Scholar] [CrossRef]
- Martins, P.; Machado, D.; Theizen, T.H.; Guarnieri, J.P.O.; Bernardes, B.G.; Gomide, G.P.; Corat, M.A.F.; Abbehausen, C.; Proenca-Modena, J.L.; Melo, C.F.O.R.; et al. Outer Membrane Vesicles from Neisseria Meningitidis (Proteossome) Used for Nanostructured Zika Virus Vaccine Production. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Liao, Z.; Tu, L.; Li, X.; Liang, X.-J.; Huo, S. Virus-inspired nanosystems for drug delivery. Nanoscale 2021, 13, 18912–18924. [Google Scholar] [CrossRef] [PubMed]
- Somiya, M.; Liu, Q.; Kuroda, S. Current Progress of Virus-mimicking Nanocarriers for Drug Delivery. Nanotheranostics 2017, 1, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Loo, Y.S.; Bose, R.J.; McCarthy, J.R.; Azmi, I.D.M.; Madheswaran, T. Biomimetic bacterial and viral-based nanovesicles for drug delivery, theranostics, and vaccine applications. Drug Discov. Today 2020, 26, 902–915. [Google Scholar] [CrossRef]
- Parodi, A.; Molinaro, R.; Sushnitha, M.; Evangelopoulos, M.; Martinez, J.O.; Arrighetti, N.; Corbo, C.; Tasciotti, E. Bio-inspired engineering of cell- and virus-like nanoparticles for drug delivery. Biomaterials 2017, 147, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Moser, C.; Amacker, M.; Kammer, A.R.; Rasi, S.; Westerfeld, N.; Zurbriggen, R. Influenza virosomes as a combined vaccine carrier and adjuvant system for prophylactic and therapeutic immunizations. Expert Rev. Vaccines 2007, 6, 711–721. [Google Scholar] [CrossRef]
- Li, R.; Liu, Q. Engineered Bacterial Outer Membrane Vesicles as Multifunctional Delivery Platforms. Front. Mater. 2020, 7, 202. [Google Scholar] [CrossRef]
- Schwechheimer, C.; Kuehn, M.J. Outer-membrane vesicles from Gram-negative bacteria: Biogenesis and functions. Nat. Rev. Microbiol. 2015, 13, 605–619. [Google Scholar] [CrossRef]
- Berleman, J.; Auer, M. The Role of Bacterial Outer Membrane Vesicles for Intra- and Interspecies Delivery. Environ. Microbiol. 2013, 15, 347–354. [Google Scholar] [CrossRef]
- Chatterjee, S.N.; Das, J. Electron Microscopic Observations on the Excretion of Cell-wall Material by Vibrio cholerae. J. Gen. Microbiol. 1967, 49, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Gao, J.; Wang, Z. Outer Membrane Vesicles for Vaccination and Targeted Drug Delivery. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2019, 11, e1523. [Google Scholar] [CrossRef]
- Li, M.; Zhou, H.; Yang, C.; Wu, Y.; Zhou, X.; Liu, H.; Wang, Y. Bacterial outer membrane vesicles as a platform for biomedical applications: An update. J. Control. Release 2020, 323, 253–268. [Google Scholar] [CrossRef] [PubMed]
- Fazal, S.; Lee, R. Biomimetic Bacterial Membrane Vesicles for Drug Delivery Applications. Pharmaceutics 2021, 13, 1430. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, W.; Blanot, D.; De Pedro, M.A. Peptidoglycan structure and architecture. FEMS Microbiol. Rev. 2008, 32, 149–167. [Google Scholar] [CrossRef] [PubMed]
- Farhana, A.; Khan, Y.S. Biochemistry, Lipopolysaccharide; 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK554414/ (accessed on 19 September 2022).
- Lee, E.Y.; Choi, D.S.; Kim, K.P.; Gho, Y.S. Proteomics in gram-negative bacterial outer membrane vesicles. Mass Spectrom. Rev. 2008, 27, 535–555. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Ji, H.; Kong, X.; Lei, P.; Yang, Q.; Wu, W.; Jin, L.; Sun, D. Bacterial Ghosts-Based Vaccine and Drug Delivery Systems. Pharmaceutics 2021, 13, 1892. [Google Scholar] [CrossRef]
- Langemann, T.; Koller, V.J.; Muhammad, A.; Kudela, P.; Mayr, U.B.; Lubitz, W. The bacterial ghost platform system: Production and Applications. Bioeng. Bugs 2010, 1, 326–336. [Google Scholar] [CrossRef]
- Kudela, P.; Koller, V.J.; Lubitz, W. Bacterial ghosts (BGs)—Advanced antigen and drug delivery system. Vaccine 2010, 28, 5760–5767. [Google Scholar] [CrossRef]
- Lubitz, P.; Mayr, U.B.; Lubitz, W. Applications of Bacterial Ghosts in Biomedicine. Pharm. Biotechnol. 2009, 655, 159–170. [Google Scholar] [CrossRef]
- Harisa, G.I.; Sherif, A.Y.; Youssof, A.M.; Alanazi, F.K.; Salem-Bekhit, M.M. Bacteriosomes as a Promising Tool in Biomedical Applications: Immunotherapy and Drug Delivery. AAPS PharmSciTech 2020, 21, 1–13. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Hessvik, N.P.; Llorente, A. Current knowledge on exosome biogenesis and release. Cell. Mol. Life Sci. 2018, 75, 193–208. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Lu, M.; Huang, Y. Bioinspired exosome-like therapeutics and delivery nanoplatforms. Biomaterials 2020, 242, 119925. [Google Scholar] [CrossRef]
- Grangier, A.; Branchu, J.; Volatron, J.; Piffoux, M.; Gazeau, F.; Wilhelm, C.; Silva, A.K. Technological advances towards extracellular vesicles mass production. Adv. Drug Deliv. Rev. 2021, 176, 113843. [Google Scholar] [CrossRef]
- Kaneti, L.; Bronshtein, T.; Dayan, N.M.; Kovregina, I.; Khait, N.L.; Lupu-Haber, Y.; Fliman, M.; Schoen, B.W.; Kaneti, G.; Machluf, M. Nanoghosts as a Novel Natural Nonviral Gene Delivery Platform Safely Targeting Multiple Cancers. Nano Lett. 2016, 16, 1574–1582. [Google Scholar] [CrossRef]
- Furman, N.E.T.; Lupu-Haber, Y.; Bronshtein, T.; Kaneti, L.; Letko, N.; Weinstein, E.; Baruch, L.; Machluf, M. Reconstructed Stem Cell Nanoghosts: A Natural Tumor Targeting Platform. Nano Lett. 2013, 13, 3248–3255. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, N.; Li, Q.; Chen, J.; Wang, Q.; Yang, H.; Tan, H.; Gao, J.; Dong, Z.; Pang, Z.; et al. Biomimetic liposomes hybrid with platelet membranes for targeted therapy of atherosclerosis. Chem. Eng. J. 2020, 408, 127296. [Google Scholar] [CrossRef]
- Oieni, J.; Levy, L.; Khait, N.L.; Yosef, L.; Schoen, B.; Fliman, M.; Shalom-Luxenburg, H.; Dayan, N.M.; D’Atri, D.; Anavy, N.C.; et al. Nano-Ghosts: Biomimetic membranal vesicles, technology and characterization. Methods 2019, 177, 126–134. [Google Scholar] [CrossRef]
- Shattuck, G.E. Hemolysis of Chicken Blood. J. Gen. Physiol. 1928, 12, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, J.F. Physiological Characteristics of Human Red Blood Cell Ghosts. J. Gen. Physiol. 1958, 42, 9–28. [Google Scholar] [CrossRef]
- He, H.; Guo, C.; Wang, J.; Korzun, W.J.; Wang, X.-Y.; Ghosh, S.; Yang, H. Leutusome: A Biomimetic Nanoplatform Integrating Plasma Membrane Components of Leukocytes and Tumor Cells for Remarkably Enhanced Solid Tumor Homing. Nano Lett. 2018, 18, 6164–6174. [Google Scholar] [CrossRef]
- Wu, H.; Jiang, X.; Li, Y.; Zhou, Y.; Zhang, T.; Zhi, P.; Gao, J. Engineering Stem Cell Derived Biomimetic Vesicles for Versatility and Effective Targeted Delivery. Adv. Funct. Mater. 2020, 30, 2006169. [Google Scholar] [CrossRef]
- Xu, L.; Wu, S.; Wang, J. Cancer cell membrane–coated nanocarriers for homologous target inhibiting the growth of hepatocellular carcinoma. J. Bioact. Compat. Polym. 2018, 34, 58–71. [Google Scholar] [CrossRef]
- Tang, W.; Yang, Y.; Yang, L.; Tang, M.; Chen, Y.; Li, C. Macrophage membrane-mediated targeted drug delivery for treatment of spinal cord injury regardless of the macrophage polarization states. Asian J. Pharm. Sci. 2021, 16, 459–470. [Google Scholar] [CrossRef]
- Yan, X.; Scherphof, G.L.; Kamps, J.A.A.M. Liposome Opsonization. J. Liposome Res. 2005, 15, 109–139. [Google Scholar] [CrossRef]
- Corbo, C.; Molinaro, R.; Tabatabaei, M.; Farokhzad, O.C.; Mahmoudi, M. Personalized protein corona on nanoparticles and its clinical implications. Biomater. Sci. 2017, 5, 378–387. [Google Scholar] [CrossRef]
- Molinaro, R.; Evangelopoulos, M.; Hoffman, J.; Corbo, C.; Taraballi, F.; Martinez, J.O.; Hartman, K.A.; Cosco, D.; Costa, G.; Romeo, I.; et al. Design and Development of Biomimetic Nanovesicles Using a Microfluidic Approach. Adv. Mater. 2018, 30, e1702749. [Google Scholar] [CrossRef]
- Corbo, C.; Molinaro, R.; Taraballi, F.; Furman, N.E.T.; Hartman, K.A.; Sherman, M.B.; De Rosa, E.; Kirui, D.K.; Salvatore, F.; Tasciotti, E. Unveiling the In Vivo Protein Corona of Circulating Leukocyte-like Carriers. ACS Nano 2017, 11, 3262–3273. [Google Scholar] [CrossRef]
- Zinger, A.; Cvetkovic, C.; Sushnitha, M.; Naoi, T.; Baudo, G.; Anderson, M.; Shetty, A.; Basu, N.; Covello, J.; Tasciotti, E.; et al. Humanized Biomimetic Nanovesicles for Neuron Targeting. Adv. Sci. 2021, 8, 2101437. [Google Scholar] [CrossRef]
- Kuerban, K.; Gao, X.; Zhang, H.; Liu, J.; Dong, M.; Wu, L.; Ye, R.; Feng, M.; Ye, L. Doxorubicin-loaded bacterial outer-membrane vesicles exert enhanced anti-tumor efficacy in non-small-cell lung cancer. Acta Pharm. Sin. B 2020, 10, 1534–1548. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, K.; Qin, X.; Li, T.; Qiu, J.; Yin, T.; Huang, J.; McGinty, S.; Pontrelli, G.; Ren, J.; et al. Biomimetic Nanotherapies: Red Blood Cell Based Core–Shell Structured Nanocomplexes for Atherosclerosis Management. Adv. Sci. 2019, 6, 1900172. [Google Scholar] [CrossRef]
- Hu, C.-M.J.; Zhang, L.; Aryal, S.; Cheung, C.; Fang, R.H.; Zhang, L. Erythrocyte membrane-camouflaged polymeric nanoparticles as a biomimetic delivery platform. Proc. Natl. Acad. Sci. USA 2011, 108, 10980–10985. [Google Scholar] [CrossRef]
- Liu, Y.; Luo, J.; Chen, X.; Liu, W.; Chen, T. Cell Membrane Coating Technology: A Promising Strategy for Biomedical Applications. Nanomicro Lett. 2019, 11, 100. [Google Scholar] [CrossRef]
- Krishnamurthy, S.; Gnanasammandhan, M.K.; Xie, C.; Huang, K.; Cui, M.Y.; Chan, J.M. Monocyte cell membrane-derived nanoghosts for targeted cancer therapy. Nanoscale 2016, 8, 6981–6985. [Google Scholar] [CrossRef]
- Lu, Y.; Hu, Q.; Jiang, C.; Gu, Z. Platelet for drug delivery. Curr. Opin. Biotechnol. 2018, 58, 81–91. [Google Scholar] [CrossRef]
- Eş, I.; Montebugnoli, L.J.; Filippi, M.F.P.; Malfatti-Gasperini, A.A.; Radaic, A.; de Jesus, M.B.; de la Torre, L.G. High-throughput conventional and stealth cationic liposome synthesis using a chaotic advection-based microfluidic device combined with a centrifugal vacuum concentrator. Chem. Eng. J. 2020, 382, 122821. [Google Scholar] [CrossRef]
- Hu, C.-M.J.; Fang, R.H.; Wang, K.-C.; Luk, B.T.; Thamphiwatana, S.; Dehaini, D.; Nguyen, P.; Angsantikul, P.; Wen, C.H.; Kroll, A.V.; et al. Nanoparticle biointerfacing by platelet membrane cloaking. Nature 2015, 526, 118–121. [Google Scholar] [CrossRef]
- Molinaro, R.; Corbo, C.; Martinez, J.O.; Taraballi, F.; Evangelopoulos, M.; Minardi, S.; Yazdi, I.K.; Zhao, P.; De Rosa, E.; Sherman, M.B.; et al. Biomimetic proteolipid vesicles for targeting inflamed tissues. Nat. Mater. 2016, 15, 1037–1046. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, Y.; Li, Q.; Liu, K.; Hou, J.; Shao, C.; Wang, Y. Immunoregulatory mechanisms of mesenchymal stem and stromal cells in inflammatory diseases. Nat. Rev. Nephrol. 2018, 14, 493–507. [Google Scholar] [CrossRef]
- Piffoux, M.; Silva, A.K.A.; Wilhelm, C.; Gazeau, F.; Tareste, D. Modification of Extracellular Vesicles by Fusion with Liposomes for the Design of Personalized Biogenic Drug Delivery Systems. ACS Nano 2018, 12, 6830–6842. [Google Scholar] [CrossRef]
- Rayamajhi, S.; Nguyen, T.D.T.; Marasini, R.; Aryal, S. Macrophage-derived exosome-mimetic hybrid vesicles for tumor targeted drug delivery. Acta Biomater. 2019, 94, 482–494. [Google Scholar] [CrossRef]
- Lv, Q.; Cheng, L.; Lu, Y.; Zhang, X.; Wang, Y.; Deng, J.; Zhou, J.; Liu, B.; Liu, J. Thermosensitive Exosome–Liposome Hybrid Nanoparticle-Mediated Chemoimmunotherapy for Improved Treatment of Metastatic Peritoneal Cancer. Adv. Sci. 2020, 7, 2000515. [Google Scholar] [CrossRef]
- Dehaini, D.; Wei, X.; Fang, R.H.; Masson, S.; Angsantikul, P.; Luk, B.T.; Zhang, Y.; Ying, M.; Jiang, Y.; Kroll, A.V.; et al. Erythrocyte-Platelet Hybrid Membrane Coating for Enhanced Nanoparticle Functionalization. Adv. Mater. 2017, 29, 1606209. [Google Scholar] [CrossRef]
- Copp, J.A.; Fang, R.H.; Luk, B.T.; Hu, C.-M.J.; Gao, W.; Zhang, K.; Zhang, L. Clearance of pathological antibodies using biomimetic nanoparticles. Proc. Natl. Acad. Sci. USA 2014, 111, 13481–13486. [Google Scholar] [CrossRef]
- Han, H.; Bártolo, R.; Li, J.; Shahbazi, M.-A.; Santos, H.A. Biomimetic platelet membrane-coated nanoparticles for targeted therapy. Eur. J. Pharm. Biopharm. 2022, 172, 1–15. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, A.; Jiang, L.; Gu, Y.; Liu, J. Hybrid Membrane-Coated Biomimetic Nanoparticles (HM@BNPs): A Multifunctional Nanomaterial for Biomedical Applications. Biomacromolecules 2021, 22, 3149–3167. [Google Scholar] [CrossRef]
- Liao, Y.; Zhang, Y.; Blum, N.T.; Lin, J.; Huang, P. Biomimetic hybrid membrane-based nanoplatforms: Synthesis, properties and biomedical applications. Nanoscale Horiz. 2020, 5, 1293–1302. [Google Scholar] [CrossRef]
- Iqbal, P.; Preece, J.A.; Mendes, P.M. Nanotechnology: The “Top-Down” and “Bottom-Up” Approaches. In Supramolecular Chemistry; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2012. [Google Scholar] [CrossRef]
- Deshmukh, S.K.; Khan, M.A.; Singh, S.; Singh, A.P. Extracellular Nanovesicles: From Intercellular Messengers to Efficient Drug Delivery Systems. ACS Omega 2021, 6, 1773–1779. [Google Scholar] [CrossRef]
- Sato, Y.T.; Umezaki, K.; Sawada, S.; Mukai, S.-A.; Sasaki, Y.; Harada, N.; Shiku, H.; Akiyoshi, K. Engineering hybrid exosomes by membrane fusion with liposomes. Sci. Rep. 2016, 6, 21933. [Google Scholar] [CrossRef]
- Tang, J.; Shen, D.; Caranasos, T.G.; Wang, Z.; Vandergriff, A.C.; Allen, T.A.; Hensley, M.T.; Dinh, P.-U.; Cores, J.; Li, T.-S.; et al. Therapeutic microparticles functionalized with biomimetic cardiac stem cell membranes and secretome. Nat. Commun. 2017, 8, 13724. [Google Scholar] [CrossRef]
- Whaley, D.; Damyar, K.; Witek, R.P.; Mendoza, A.; Alexander, M.; Lakey, J.R. Cryopreservation: An Overview of Principles and Cell-Specific Considerations. Cell Transplant. 2021, 30, 096368972199961. [Google Scholar] [CrossRef]
- Steponkus, P.L.; Lynch, D.V. Freeze/thaw-induced destabilization of the plasma membrane and the effects of cold acclimation. J. Bioenerg. Biomembr. 1989, 21, 21–41. [Google Scholar] [CrossRef]
- Sundaram, J.; Mellein, B.R.; Mitragotri, S. An Experimental and Theoretical Analysis of Ultrasound-Induced Permeabilization of Cell Membranes. Biophys. J. 2003, 84, 3087–3101. [Google Scholar] [CrossRef]
- Miller, M.W.; Miller, D.L.; Brayman, A.A. A review of in vitro bioeffects of inertial ultrasonic cavitation from a mechanistic perspective. Ultrasound Med. Biol. 1996, 22, 1131–1154. [Google Scholar] [CrossRef]
- Rivas, D.F.; Kuhn, S. Synergy of Microfluidics and Ultrasound: Process Intensification Challenges and Opportunities. Top Curr. Chem. 2016, 374, 225–254. [Google Scholar] [CrossRef]
- Guo, P.; Huang, J.; Zhao, Y.; Martin, C.R.; Zare, R.N.; Moses, M.A. Nanomaterial Preparation by Extrusion through Nanoporous Membranes. Small 2018, 14, e1703493. [Google Scholar] [CrossRef]
- Lin, Y.; Wu, J.; Gu, W.; Huang, Y.; Tong, Z.; Huang, L.; Tan, J. Exosome-Liposome Hybrid Nanoparticles Deliver CRISPR/Cas9 System in MSCs. Adv. Sci. 2018, 5, 1700611. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Zhao, Y.; Banks, W.A.; Bullock, K.M.; Haney, M.; Batrakova, E.; Kabanov, A.V. Macrophage exosomes as natural nanocarriers for protein delivery to inflamed brain. Biomaterials 2017, 142, 1–12. [Google Scholar] [CrossRef]
- Nir, S.; Düzgünes, N.; De Lima, M.C.P.; Hoekstra, D. Fusion of enveloped viruses with cells and liposomes. Cell Biophys. 1990, 17, 181–201. [Google Scholar] [CrossRef]
- Aqil, F.; Munagala, R.; Jeyabalan, J.; Agrawal, A.; Gupta, R. Exosomes for the Enhanced Tissue Bioavailability and Efficacy of Curcumin. AAPS J. 2017, 19, 1691–1702. [Google Scholar] [CrossRef] [PubMed]
- Oskouie, M.N.; Moghaddam, N.S.A.; Butler, A.E.; Zamani, P.; Sahebkar, A. Therapeutic use of curcumin-encapsulated and curcumin-primed exosomes. J. Cell. Physiol. 2018, 234, 8182–8191. [Google Scholar] [CrossRef] [PubMed]
- Lentz, B.R.; Lee, J.; Lentz, J.L.B.R. Poly(ethylene glycol) (PEG)-mediated fusion between pure lipid bilayers: A mechanism in common with viral fusion and secretory vesicle release? (Review). Mol. Membr. Biol. 1999, 16, 279–296. [Google Scholar] [CrossRef] [PubMed]
- Ellens, H.; Siegel, D.P.; Alford, D.; Yeagle, P.; Boni, L.; Lis, L.J.; Quinn, P.J.; Bentz, J. Membrane fusion and inverted phases. Biochemistry 1989, 28, 3692–3703. [Google Scholar] [CrossRef]
- Kolašinac, R.; Kleusch, C.; Braun, T.; Merkel, R.; Csiszár, A. Deciphering the Functional Composition of Fusogenic Liposomes. Int. J. Mol. Sci. 2018, 19, 346. [Google Scholar] [CrossRef]
- Suárez-Germà, C.; Montero, M.; Ignés-Mullol, J.; Hernández-Borrell, O. Domènech. Acyl Chain Differences in Phosphatidylethanolamine Determine Domain Formation and LacY Distribution in Biomimetic Model Membranes. J. Phys. Chem. B 2011, 115, 12778–12784. [Google Scholar] [CrossRef]
- Dawaliby, R.; Trubbia, C.; Delporte, C.; Noyon, C.; Ruysschaert, J.-M.; Van Antwerpen, P.; Govaerts, C. Phosphatidylethanolamine Is a Key Regulator of Membrane Fluidity in Eukaryotic Cells. J. Biol. Chem. 2016, 291, 3658–3667. [Google Scholar] [CrossRef]
- Kobierski, J.; Wnętrzak, A.; Chachaj-Brekiesz, A.; Dynarowicz-Latka, P. Predicting the packing parameter for lipids in monolayers with the use of molecular dynamics. Colloids Surf. B Biointerfaces 2021, 211, 112298. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, S.; Kanegae, N.; Nishina, K.; Kamikawa, Y.; Koiwai, K.; Masunaga, H.; Sakurai, K. The role of the helper lipid dioleoylphosphatidylethanolamine (DOPE) for DNA transfection cooperating with a cationic lipid bearing ethylenediamine. Biochim. Biophys. Acta (BBA) -Biomembr. 2013, 1828, 412–418. [Google Scholar] [CrossRef] [Green Version]
- Papahadjopoulos, D.; Nir, S.; Düzgünes, N. Molecular mechanisms of calcium-induced membrane fusion. J. Bioenerg. Biomembr. 1990, 22, 157–179. [Google Scholar] [CrossRef]
- Düzgüne, N.; Nir, S.; Wilschut, J.; Bentz, J.; Newton, C.; Portis, A.; Papahadjopoulos, D. Calcium- and magnesium-induced fusion of mixed phosphatidylserine/phosphatidylcholine vesicles: Effect of ion binding. J. Membr. Biol. 1981, 59, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Wilschut, J.; Holsappel, M.; Jansen, R. Ca2+-induced fusion of cardiolipin/phosphatidylcholine vesicles monitored by mixing of aqueous contents. Biochim. Biophys. Acta (BBA) -Biomembr. 1982, 690, 297–301. [Google Scholar] [CrossRef]
- Schmid, Y.R.F.; Scheller, L.; Buchmann, S.; Dittrich, P.S. Calcium-Mediated Liposome Fusion to Engineer Giant Lipid Vesicles with Cytosolic Proteins and Reconstituted Mammalian Proteins. Adv. Biosyst. 2020, 4, e2000153. [Google Scholar] [CrossRef]
- Carvalho, B.G.; Ceccato, B.T.; Michelon, M.; Han, S.W.; de la Torre, L.G. Advanced Microfluidic Technologies for Lipid Nano-Microsystems from Synthesis to Biological Application. Pharmaceutics 2022, 14, 141. [Google Scholar] [CrossRef]
- Carugo, D.; Bottaro, E.; Owen, J.; Stride, E.; Nastruzzi, C. Liposome production by microfluidics: Potential and limiting factors. Sci. Rep. 2016, 6, 25876. [Google Scholar] [CrossRef]
- Balbino, T.A.; Aoki, N.T.; Gasperini, A.A.; Oliveira, C.L.; Azzoni, A.R.; Cavalcanti, L.P.; de la Torre, L.G. Continuous flow production of cationic liposomes at high lipid concentration in microfluidic devices for gene delivery applications. Chem. Eng. J. 2013, 226, 423–433. [Google Scholar] [CrossRef]
- Pessoa, A.C.S.N.; Sipoli, C.C.; de la Torre, L.G. Effects of diffusion and mixing pattern on microfluidic-assisted synthesis of chitosan/ATP nanoparticles. Lab Chip 2017, 17, 2281–2293. [Google Scholar] [CrossRef] [PubMed]
- Firmino, P.C.O.S.; Vianna, S.S.V.; da Costa, O.M.M.M.; Malfatti-Gasperini, A.A.; Gobbi, A.L.; Lima, R.S.; de la Torre, L.G. 3D micromixer for nanoliposome synthesis: A promising advance in high mass productivity. Lab Chip 2021, 21, 2971–2985. [Google Scholar] [CrossRef] [PubMed]
- Lang, T.; Yin, Q.; Li, Y. Progress of Cell-Derived Biomimetic Drug Delivery Systems for Cancer Therapy. Adv. Ther. 2018, 1, 1800053. [Google Scholar] [CrossRef]
- Kroll, A.V.; Jiang, Y.; Zhou, J.; Holay, M.; Fang, R.H.; Zhang, L. Biomimetic Nanoparticle Vaccines for Cancer Therapy. Adv. Biosyst. 2018, 3, 1800219. [Google Scholar] [CrossRef]
- Wang, R.; Yan, H.; Yu, A.; Ye, L.; Zhai, G. Cancer targeted biomimetic drug delivery system. J. Drug Deliv. Sci. Technol. 2021, 63, 102530. [Google Scholar] [CrossRef]
- Sun, H.; Su, J.; Meng, Q.; Yin, Q.; Chen, L.; Gu, W.; Zhang, P.; Zhang, Z.; Yu, H.; Wang, S.; et al. Cancer-Cell-Biomimetic Nanoparticles for Targeted Therapy of Homotypic Tumors. Adv. Mater. 2016, 28, 9581–9588. [Google Scholar] [CrossRef] [PubMed]
- Mendanha, D.; de Castro, J.V.; Ferreira, H.; Neves, N. Biomimetic and cell-based nanocarriers—New strategies for brain tumor targeting. J. Control. Release 2021, 337, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Zhao, Y.; Li, Y.; Jiang, L.; Gu, Y.; Liu, J. Cell-derived biomimetic nanocarriers for targeted cancer therapy: Cell membranes and extracellular vesicles. Drug Deliv. 2021, 28, 1237–1255. [Google Scholar] [CrossRef]
- Beh, C.Y.; Prajnamitra, R.P.; Chen, L.-L.; Hsieh, P.C.-H. Advances in Biomimetic Nanoparticles for Targeted Cancer Therapy and Diagnosis. Molecules 2021, 26, 5052. [Google Scholar] [CrossRef]
- Kantoff, P.W.; Higano, C.S.; Shore, N.D.; Berger, E.R.; Small, E.J.; Penson, D.F.; Redfern, C.H.; Ferrari, A.C.; Dreicer, R.; Sims, R.B.; et al. Sipuleucel-T Immunotherapy for Castration-Resistant Prostate Cancer. N. Engl. J. Med. 2010, 363, 411–422. [Google Scholar] [CrossRef]
- Chen, Q.; Huang, G.; Wu, W.; Wang, J.; Hu, J.; Mao, J.; Chu, P.; Bai, H.; Tang, G. A Hybrid Eukaryotic–Prokaryotic Nanoplatform with Photothermal Modality for Enhanced Antitumor Vaccination. Adv. Mater. 2020, 32, e1908185. [Google Scholar] [CrossRef]
- Kines, R.C.; Thompson, C.D.; Spring, S.; Li, Z.; Pinos, E.D.L.; Monks, S.; Schiller, J.T. Virus-Like Particle–Drug Conjugates Induce Protective, Long-lasting Adaptive Antitumor Immunity in the Absence of Specifically Targeted Tumor Antigens. Cancer Immunol. Res. 2021, 9, 693–706. [Google Scholar] [CrossRef]
- Mizrak, A.; Bolukbasi, M.F.; Ozdener, G.B.; Brenner, G.J.; Madlener, S.; Erkan, E.P.; Ströbel, T.; Breakefield, X.O.; Saydam, O. Genetically Engineered Microvesicles Carrying Suicide mRNA/Protein Inhibit Schwannoma Tumor Growth. Mol. Ther. 2013, 21, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, L.; Yu, J.; Zhu, D.; Zhang, Y.; Li, X.; Gu, H.; Zhang, C.-Y.; Zen, K. Microvesicle-mediated delivery of transforming growth factor β1 siRNA for the suppression of tumor growth in mice. Biomaterials 2014, 35, 4390–4400. [Google Scholar] [CrossRef]
- MacDiarmid, J.A.; Amaro-Mugridge, N.B.; Madrid-Weiss, J.; Sedliarou, I.; Wetzel, S.; Kochar, K.; Brahmbhatt, V.N.; Phillips, L.; Pattison, S.T.; Petti, C.; et al. Sequential treatment of drug-resistant tumors with targeted minicells containing siRNA or a cytotoxic drug. Nat. Biotechnol. 2009, 27, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, H.; Gorain, B.; Kesharwani, P. Physiology of the biological barriers. In Theory and Applications of Nonparenteral Nanomedicines; Elsevier: Amsterdam, The Netherlands, 2021; pp. 79–95. [Google Scholar] [CrossRef]
- Elliott, R.; He, M. Unlocking the Power of Exosomes for Crossing Biological Barriers in Drug Delivery. Pharmaceutics 2021, 13, 122. [Google Scholar] [CrossRef]
- Torres-Vanegas, J.; Cruz, J.; Reyes, L. Delivery Systems for Nucleic Acids and Proteins: Barriers, Cell Capture Pathways and Nanocarriers. Pharmaceutics 2021, 13, 428. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Song, H.; Xu, J.; Huang, J.; Hu, M.; Gu, X.; Chen, J.; Zheng, G.; Chen, H.; Gao, X. Biomimetic ApoE-Reconstituted High Density Lipoprotein Nanocarrier for Blood–Brain Barrier Penetration and Amyloid Beta-Targeting Drug Delivery. Mol. Pharm. 2016, 13, 3976–3987. [Google Scholar] [CrossRef] [PubMed]
- Gray, S.J.; Woodard, K.T.; Samulski, R.J. Viral vectors and delivery strategies for CNS gene therapy. Ther. Deliv. 2010, 1, 517–534. [Google Scholar] [CrossRef]
- Piret, J.; Boivin, G. Pandemics Throughout History. Front. Microbiol. 2021, 11, 631736. [Google Scholar] [CrossRef]
- Depelsenaire, A.; Kendall, M.; Young, P.; Muller, D. Introduction to Vaccines and Vaccination. In Micro and Nanotechnology in Vaccine Development; Elsevier: Amsterdam, The Netherlands, 2017; pp. 47–62. [Google Scholar] [CrossRef]
- Morris, A.; Wongrakpanich, A.; Geary, S.; Salem, A. Microparticles and Nanoparticles for Cancer-Targeting Vaccines. In Micro and Nanotechnology in Vaccine Development; Elsevier: Amsterdam, The Netherlands, 2017; pp. 171–183. [Google Scholar] [CrossRef]
- de Bruijn, I.; Nauta, J.; Gerez, L.; Palache, A. The virosomal influenza vaccine Invivac®: Immunogenicity and tolerability compared to an adjuvanted influenza vaccine (Fluad®) in elderly subjects. Vaccine 2006, 24, 6629–6631. [Google Scholar] [CrossRef]
- Herzog, C.; Hartmann, K.; Künzi, V.; Kürsteiner, O.; Mischler, R.; Lazar, H.; Glück, R. Eleven years of Inflexal® V—a virosomal adjuvanted influenza vaccine. Vaccine 2009, 27, 4381–4387. [Google Scholar] [CrossRef]
- Bovier, P.A. Epaxal®: A virosomal vaccine to prevent hepatitis A infection. Expert Rev. Vaccines 2008, 7, 1141–1150. [Google Scholar] [CrossRef] [PubMed]
- Van Der Pol, L.; Stork, M.; Van Der Ley, P. Outer membrane vesicles as platform vaccine technology. Biotechnol. J. 2015, 10, 1689–1706. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A. Nanomaterials Based on Lipids for Vaccine Development. In Micro and Nanotechnology in Vaccine Development; Elsevier: Amsterdam, The Netherlands, 2017; pp. 241–257. [Google Scholar] [CrossRef]
- Chu, J.S.; Villarreal, D.O.; Weiner, D.B. DNA Vaccines: A Strategy for Developing Novel Multivalent TB Vaccines. Breast Cancer 2016, 1403, 355–361. [Google Scholar] [CrossRef]
- Takamura, S.; Niikura, M.; Li, T.-C.; Takeda, N.; Kusagawa, S.; Takebe, Y.; Miyamura, T.; Yasutomi, Y. DNA vaccine-encapsulated virus-like particles derived from an orally transmissible virus stimulate mucosal and systemic immune responses by oral administration. Gene Ther. 2004, 11, 628–635. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, P.-Q.; Guo, S.; Chen, G.; Zhao, Z.; Wang, G.-X.; Zhu, B. Application of Biomimetic Cell-Derived Nanoparticles with Mannose Modification as a Novel Vaccine Delivery Platform against Teleost Fish Viral Disease. ACS Biomater. Sci. Eng. 2020, 6, 6770–6777. [Google Scholar] [CrossRef] [PubMed]
- Ebensen, T.; Paukner, S.; Link, C.; Kudela, P.; de Domenico, C.; Lubitz, W.; Guzmán, C.A. Bacterial Ghosts Are an Efficient Delivery System for DNA Vaccines. J. Immunol. 2004, 172, 6858–6865. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mougenot, M.F.; Pereira, V.S.; Costa, A.L.R.; Lancellotti, M.; Porcionatto, M.A.; da Silveira, J.C.; de la Torre, L.G. Biomimetic Nanovesicles—Sources, Design, Production Methods, and Applications. Pharmaceutics 2022, 14, 2008. https://doi.org/10.3390/pharmaceutics14102008
Mougenot MF, Pereira VS, Costa ALR, Lancellotti M, Porcionatto MA, da Silveira JC, de la Torre LG. Biomimetic Nanovesicles—Sources, Design, Production Methods, and Applications. Pharmaceutics. 2022; 14(10):2008. https://doi.org/10.3390/pharmaceutics14102008
Chicago/Turabian StyleMougenot, Marcel Franco, Vanessa Sousa Pereira, Ana Letícia Rodrigues Costa, Marcelo Lancellotti, Marimelia Aparecida Porcionatto, Juliano Coelho da Silveira, and Lucimara Gaziola de la Torre. 2022. "Biomimetic Nanovesicles—Sources, Design, Production Methods, and Applications" Pharmaceutics 14, no. 10: 2008. https://doi.org/10.3390/pharmaceutics14102008