
Effects of Lipid Shape and Interactions on the Conformation, Dynamics, and Curvature of Ultrasound-Responsive Liposomes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Simulations of Bilayers at Different Lipid Ratios
2.2. Simulations of a DOXIL Bilayer
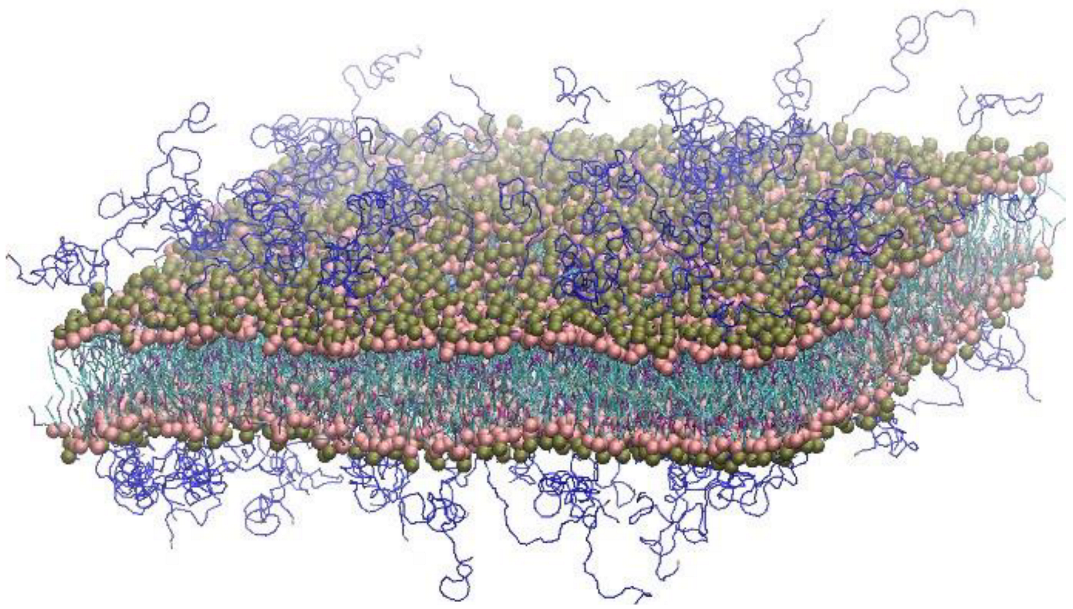
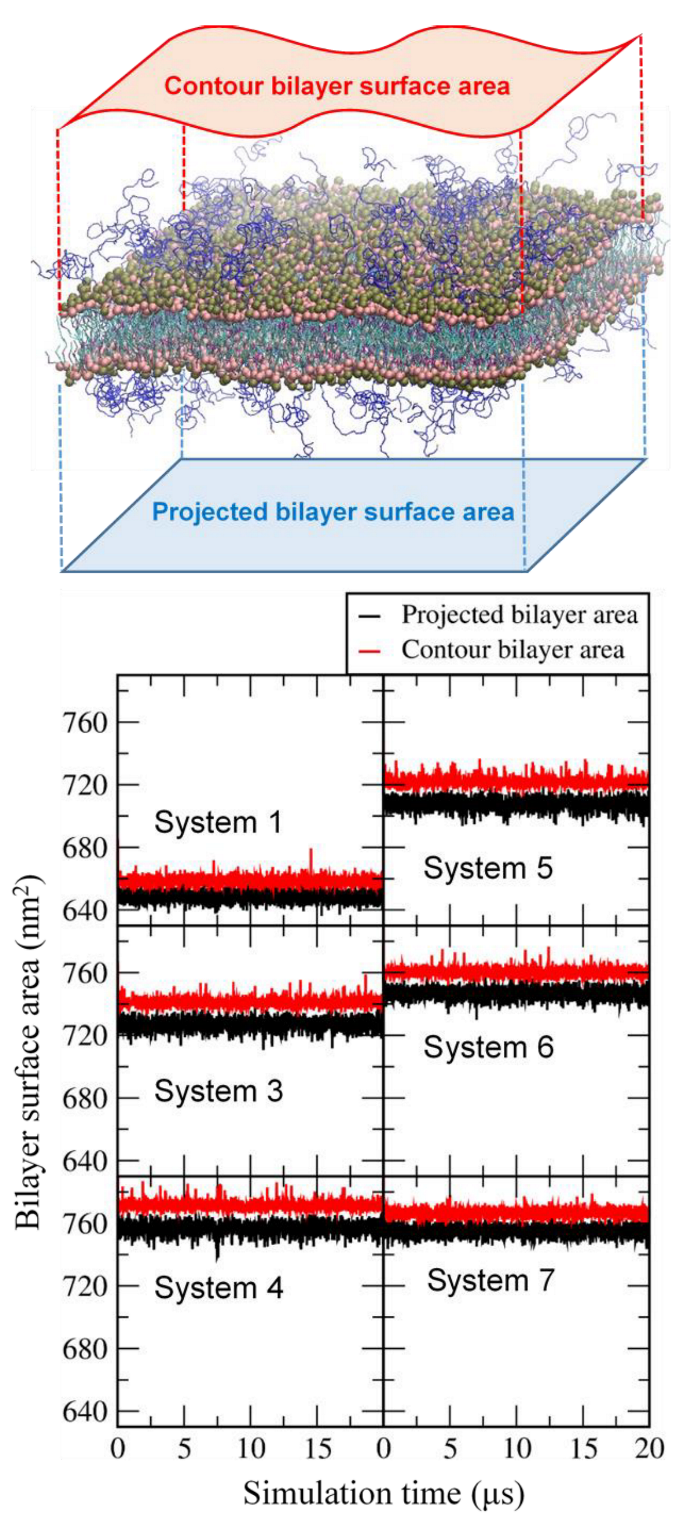
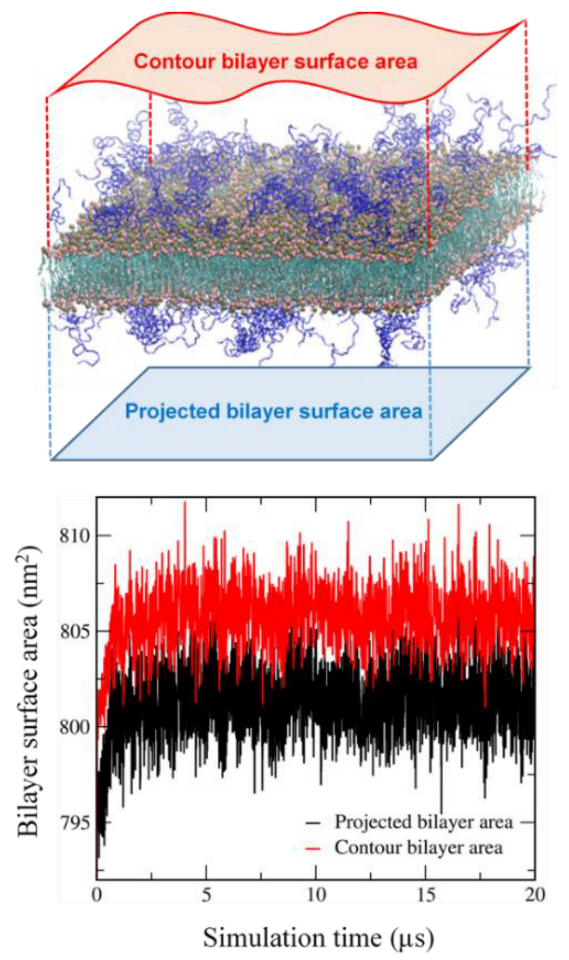
2.3. Calculation of the Contour Bilayer-Surface Area
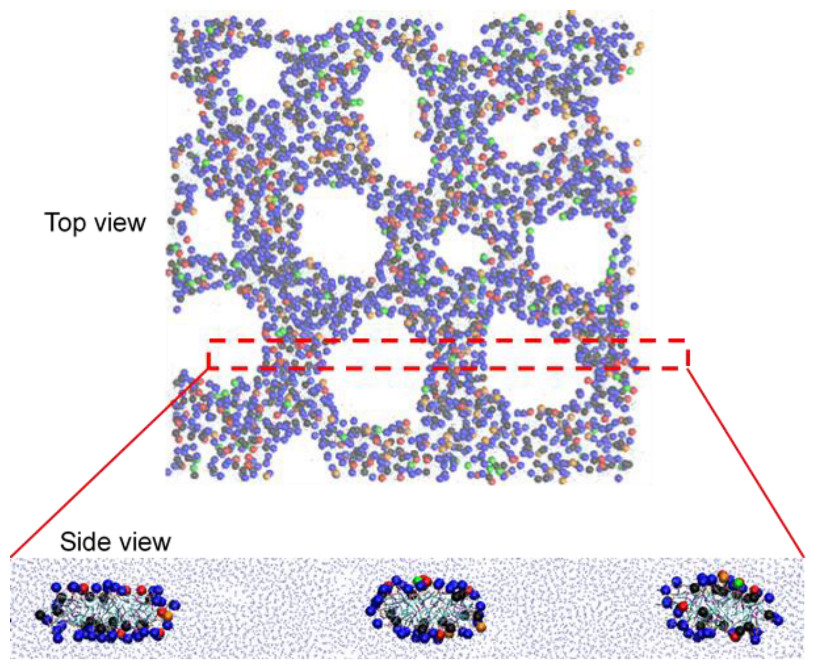
3. Results and Discussion
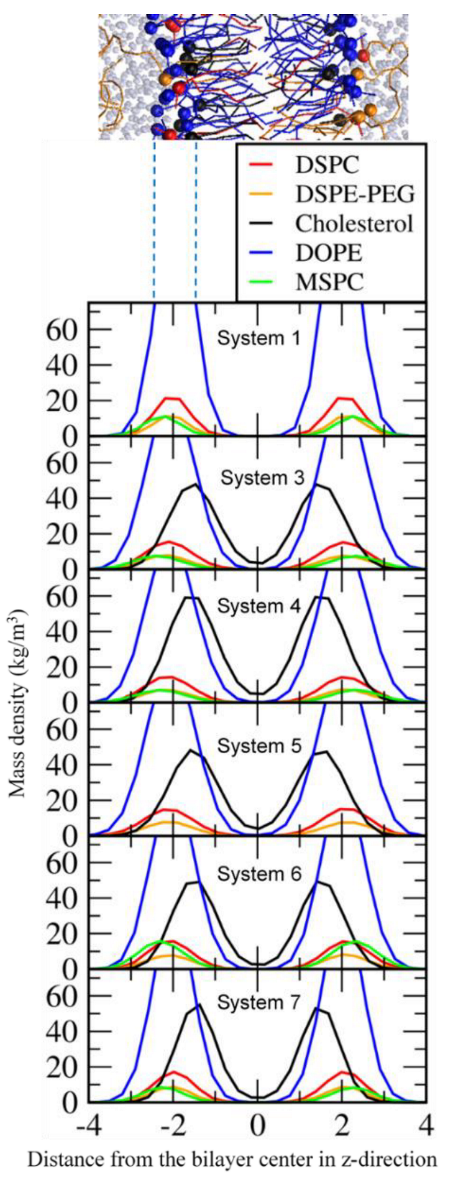
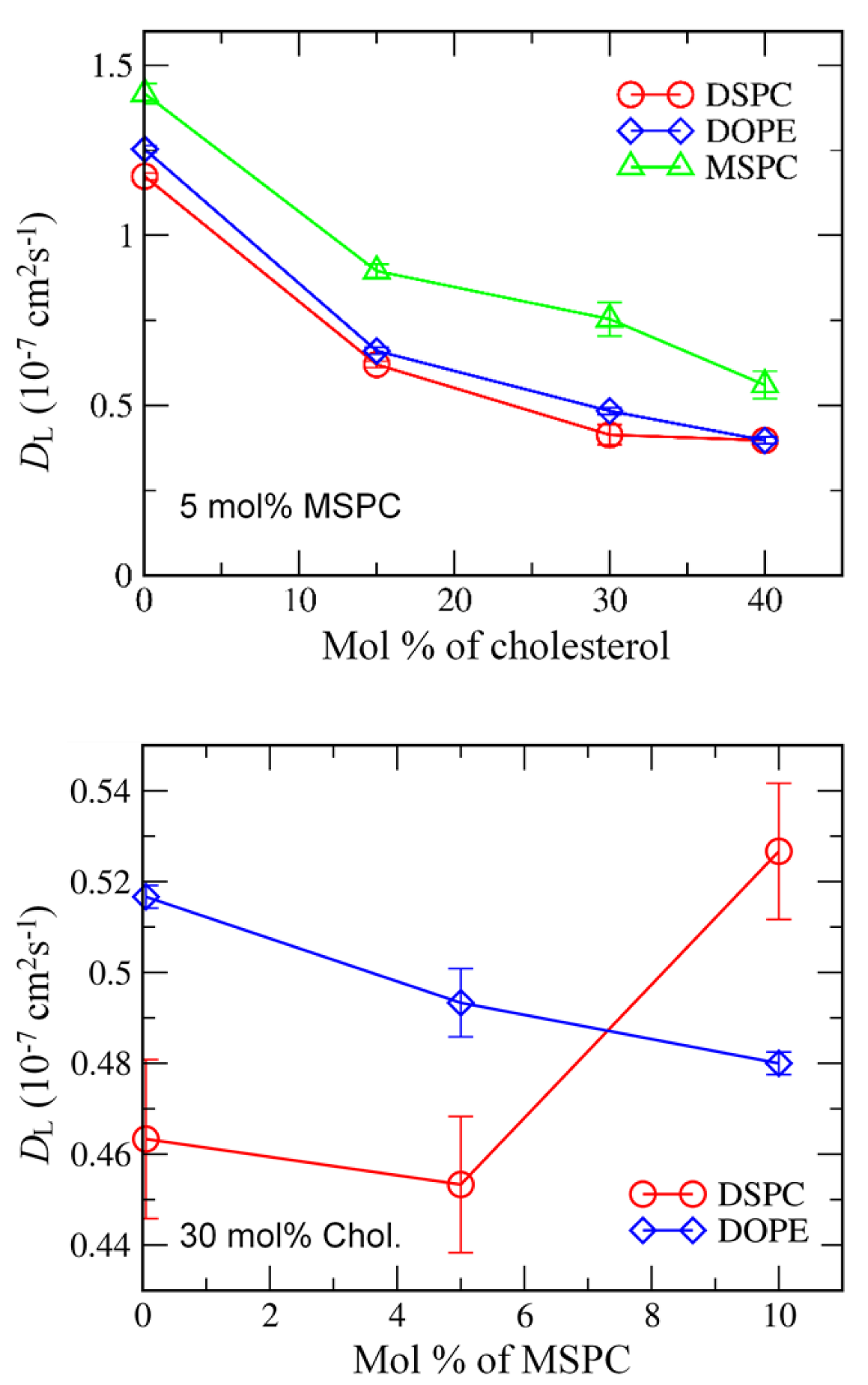
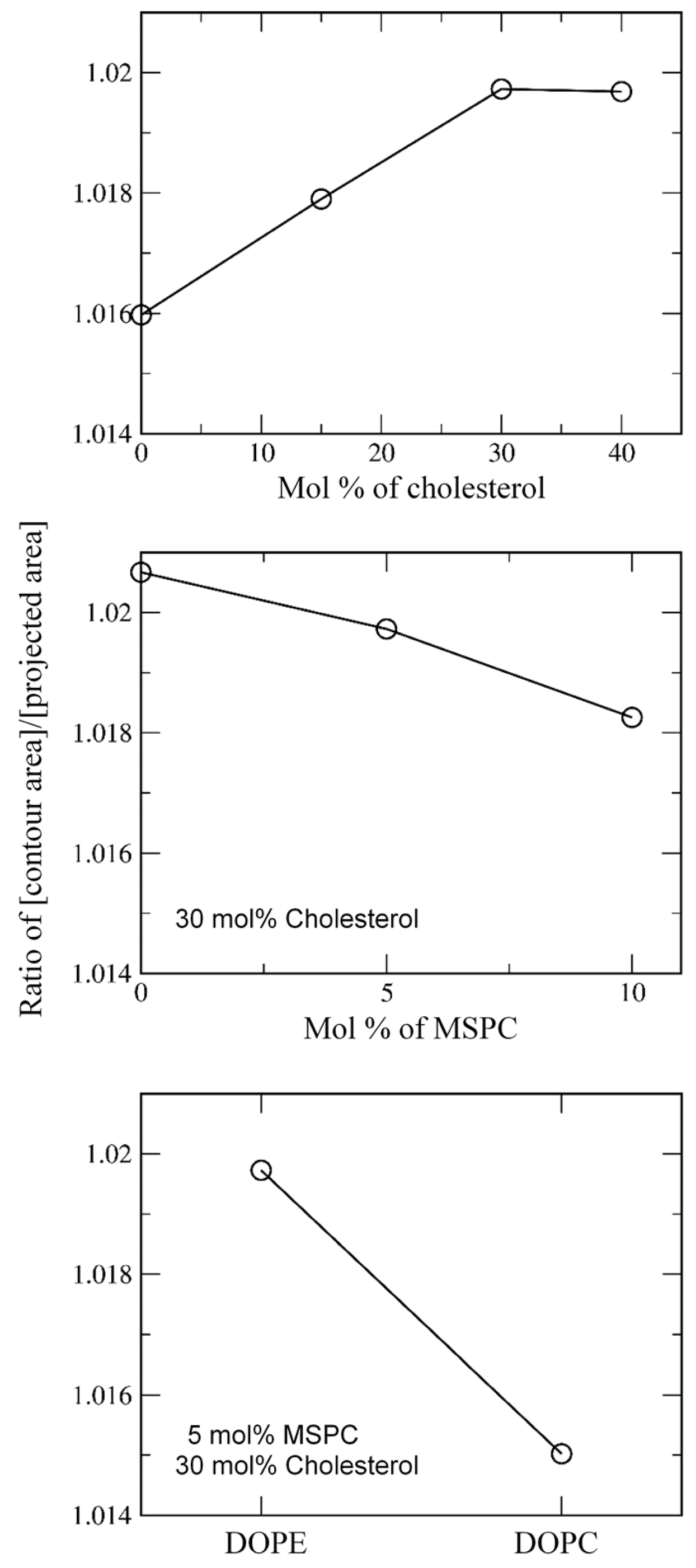
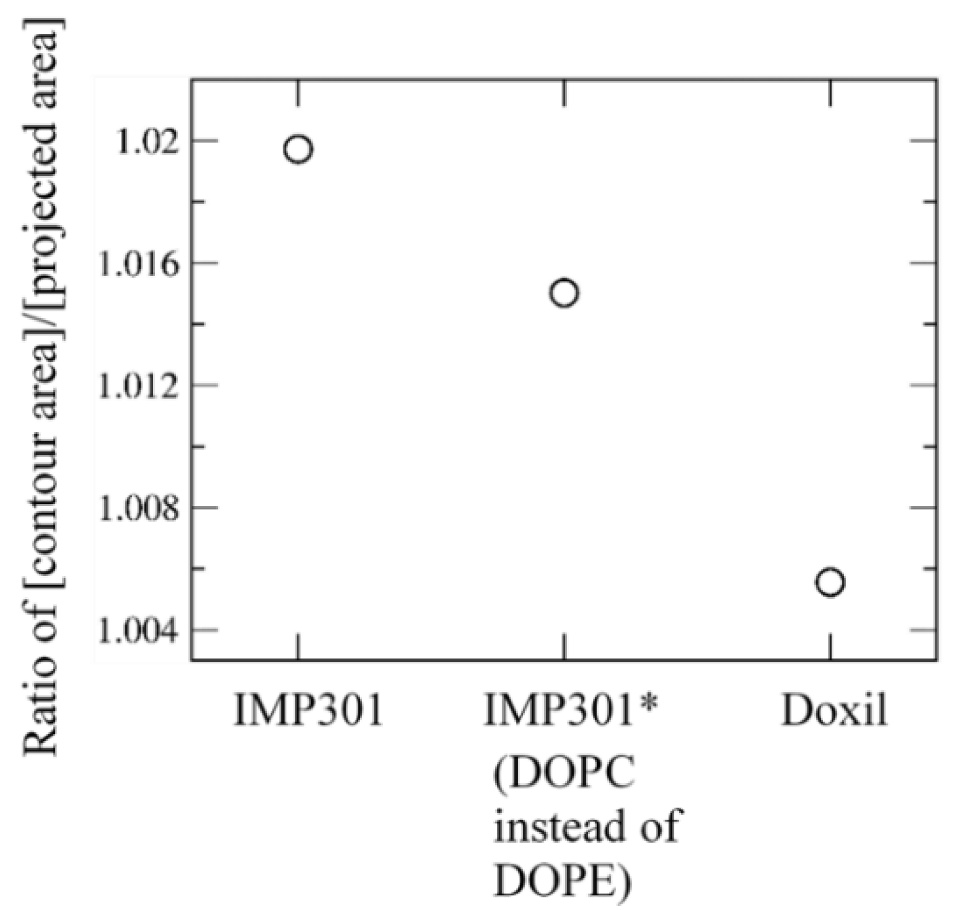
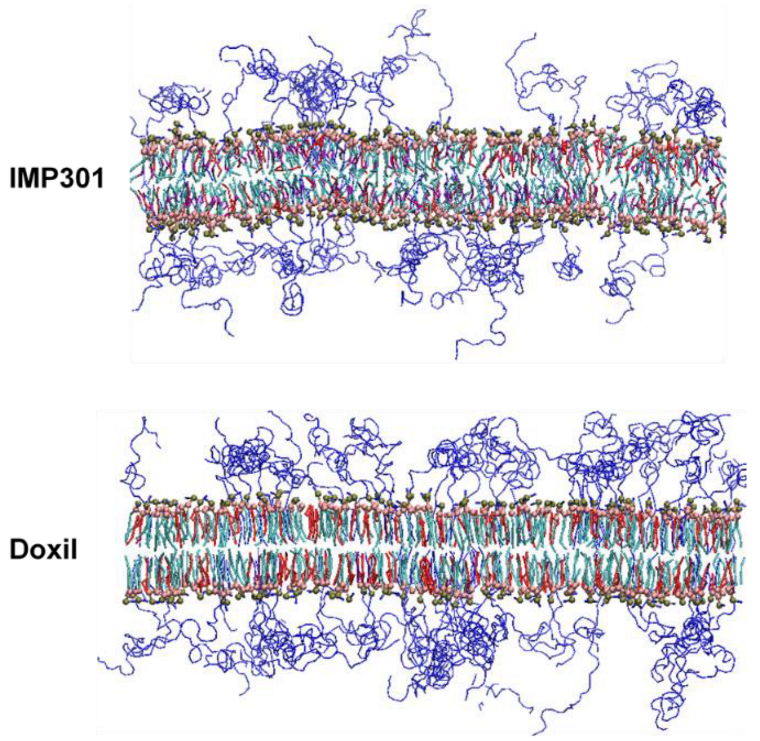
3.1. Effects of Cholesterol and MSPC on the Bilayer Conformation, Dynamics, and Curvature
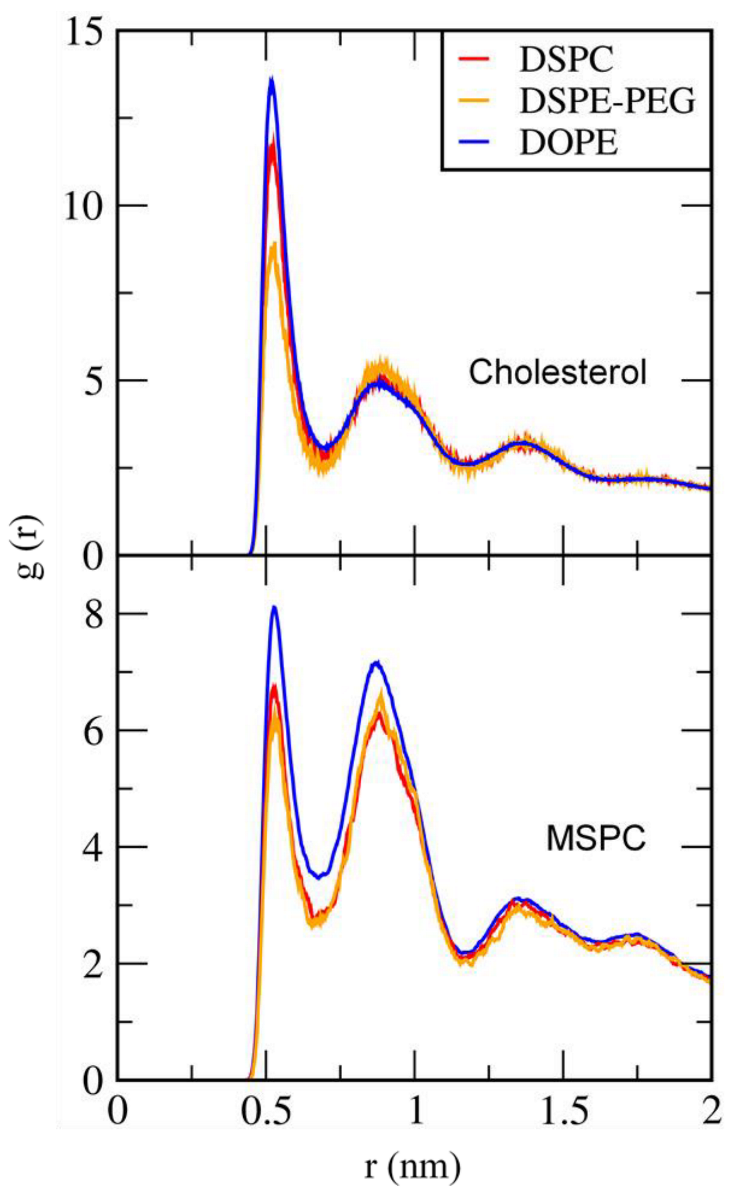
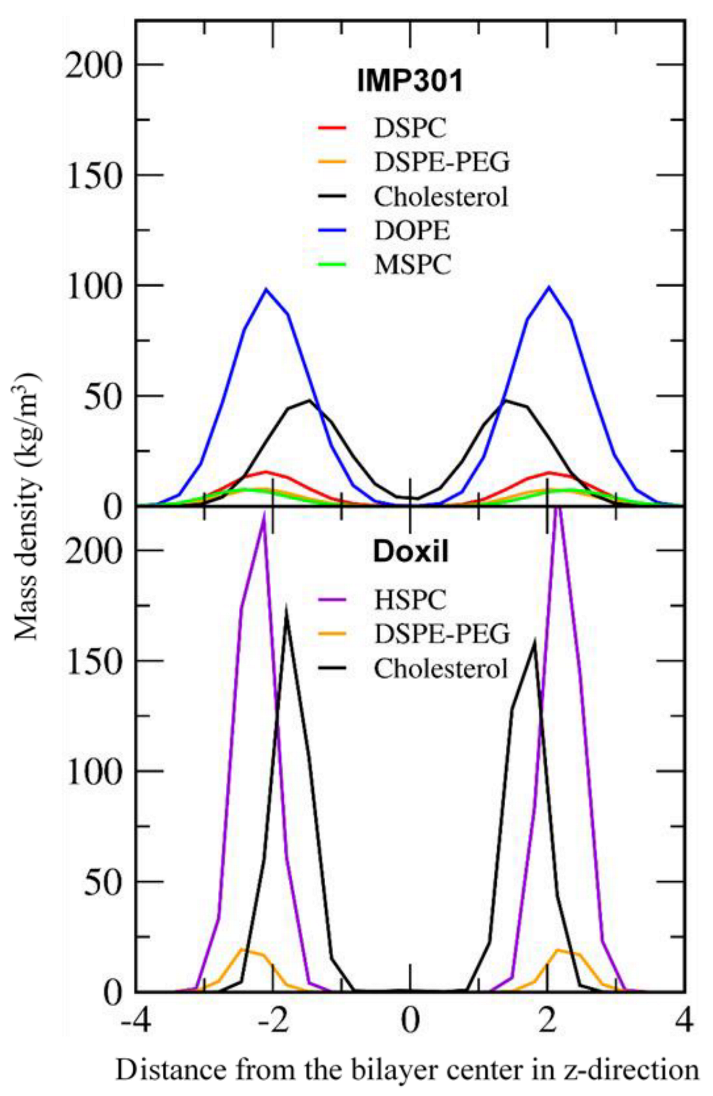
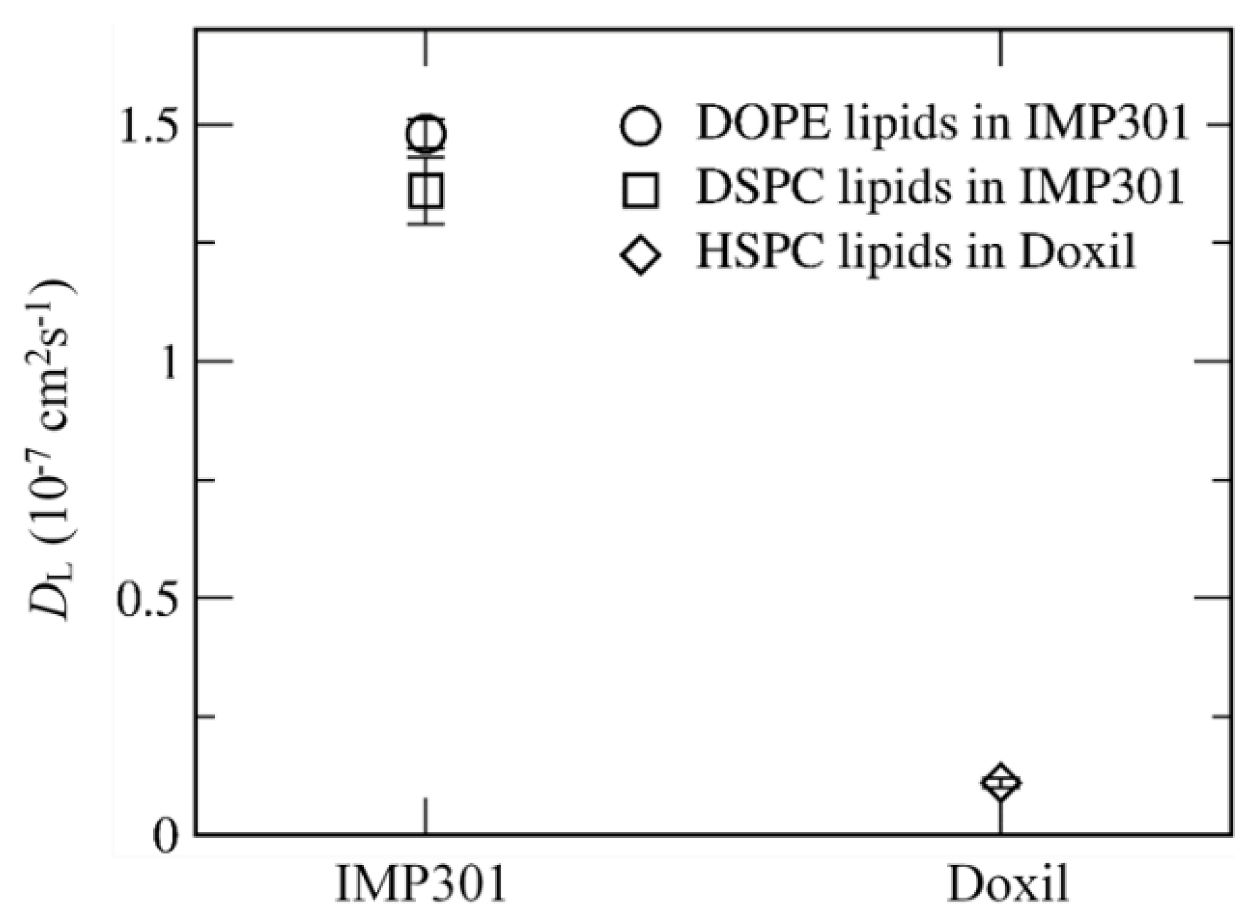
3.2. Effect of Lipid Shape on the Bilayer Conformation, Dynamics, and Curvature
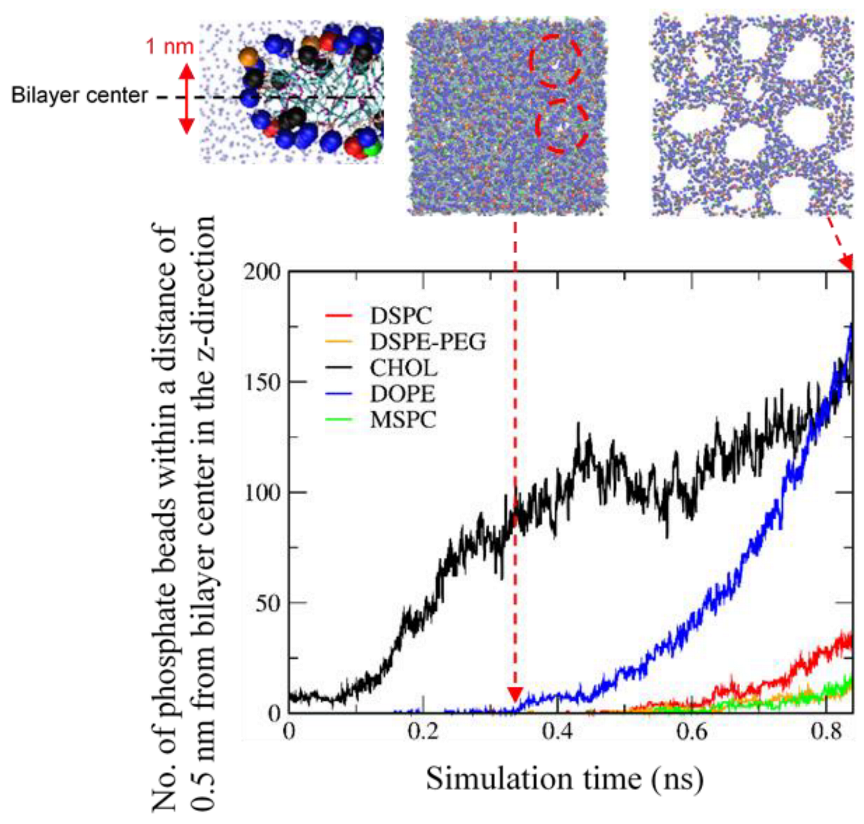
3.3. Pore Formation Induced by an External Pressure
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gregoriadis, G. The Carrier Potential of Liposomes in Biology and Medicine (First of Two Parts). N. Engl. J. Med. 1976, 295, 704–710. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Lee, R.J.; Cauchon, G.; Gorenstein, D.G.; Low, P.S. Delivery of antisense oligodeoxyribonucleotides against the human epidermal growth-factor receptor into cultured kb cells with liposomes conjugated to folate via polyethylene-glycol. Proc. Natl. Acad. Sci. USA 1995, 92, 3318–3322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papahadjopoulos, D.; Allen, T.M.; Gabizon, A.; Mayhew, E.; Matthay, K.; Huang, S.K.; Lee, K.D.; Woodle, M.C.; Lasic, D.D.; Redemann, C.; et al. Sterically stabilized liposomes—Improvements in pharmacokinetics and antitumor therapeutic efficacy. Proc. Natl. Acad. Sci. USA 1991, 88, 11460–11464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrari, M. Cancer Nanotechnology: Opportunities and Challenges. Nat. Rev. Cancer 2005, 5, 161–171. [Google Scholar] [CrossRef]
- Lasic, D.D.; Needham, D. The “Stealth” Liposome: A Prototypical Biomaterial. Chem. Rev. 1995, 95, 2601–2628. [Google Scholar] [CrossRef]
- Fahmy, T.M.; Fong, P.M.; Park, J.; Constable, T.; Saltzman, W.M. Nanosystems for Simultaneous Imaging and Drug Delivery to T Cells. Aaps J. 2007, 9, E171–E180. [Google Scholar] [CrossRef]
- Allen, T.M.; Hansen, C. Pharmacokinetics of stealth versus conventional liposomes—Effect of dose. Biochim. Biophys. Acta 1991, 1068, 133–141. [Google Scholar] [CrossRef]
- Allen, T.M.; Hansen, C.; Martin, F.; Redemann, C.; Yauyoung, A. Liposomes containing synthetic lipid derivatives of poly(ethylene glycol) show prolonged circulation half-lives invivo. Biochim. Biophys. Acta 1991, 1066, 29–36. [Google Scholar] [CrossRef]
- Bloom, A. Liposome-Based Patents for Use in the Pharmaceutical Industry. Liposomes Ration. Des. 1999, 397–442. [Google Scholar]
- Maruyama, K.; Yuda, T.; Okamoto, A.; Ishikura, C.; Kojima, S.; Iwatsuru, M. Effect of molecular-weight in amphipathic polyethyleneglycol on prolonging the circulation time of large unilamellar liposomes. Chem. Pharm. Bull. 1991, 39, 1620–1622. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, K.; Yuda, T.; Okamoto, A.; Kojima, S.; Suginaka, A.; Iwatsuru, M. Prolonged circulation time invivo of large unilamellar liposomes composed of distearoyl phosphatidylcholine and cholesterol containing amphipathic poly(ethylene glycol). Biochim. Biophys. Acta 1992, 1128, 44–49. [Google Scholar] [CrossRef]
- Van Ballegooie, C.; Man, A.; Win, M.; Yapp, D.T. Spatially Specific Liposomal Cancer Therapy Triggered by Clinical External Sources of Energy. Pharmaceutics 2019, 11, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yatvin, M.B.; Weinstein, J.N.; Dennis, W.H.; Blumenthal, R. Design of Liposomes for Enhanced Local Release of Drugs by Hyperthermia. Science 1978, 202, 1290–1293. [Google Scholar] [CrossRef]
- Weinstein, J.N.; Magin, R.L.; Cysyk, R.L.; Zaharko, D.S. Treatment of Solid L1210 Murine Tumors with Local Hyperthermia and Temperature-Sensitive Liposomes Containing Methotrexate. Cancer Res. 1980, 40, 1388–1395. [Google Scholar] [PubMed]
- Kiser, P.F.; Wilson, G.; Needham, D. A Synthetic Mimic of the Secretory Granule for Drug Delivery. Nature 1998, 394, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Kiser, P.F.; Wilson, G.; Needham, D. Lipid-Coated Microgels for the Triggered Release of Doxorubicin. J. Control. Release 2000, 68, 9–22. [Google Scholar] [CrossRef]
- Ta, T.; Convertine, A.J.; Reyes, C.R.; Stayton, P.S.; Porter, T.M. Thermosensitive Liposomes Modified with Poly(N-Isopropylacrylamide-Co- Propylacrylic Acid) Copolymers for Triggered Release of Doxorubicin. Biomacromolecules 2010, 11, 1915–1920. [Google Scholar] [CrossRef] [Green Version]
- Na, K.; Lee, S.A.; Jung, S.H.; Hyun, J.; Shin, B.C. Elastin-like Polypeptide Modified Liposomes for Enhancing Cellular Uptake into Tumor Cells. Colloids Surf. B Biointerfaces 2012, 91, 130–136. [Google Scholar] [CrossRef]
- Torchilin, V.P. Multifunctional, Stimuli-Sensitive Nanoparticulate Systems for Drug Delivery. Nat. Rev. Drug Discov. 2014, 13, 813–827. [Google Scholar] [CrossRef] [Green Version]
- Barry, B.W. Novel Mechanisms and Devices to Enable Successful Transdermal Drug Delivery. Eur. J. Pharm. Sci. 2001, 14, 101–114. [Google Scholar] [CrossRef]
- Ferrara, K.; Pollard, R.; Borden, M. Ultrasound Microbubble Contrast Agents: Fundamentals and Application to Gene and Drug Delivery. In Annual Review of Biomedical Engineering; Annual Reviews: San Mateo, CA, USA, 2007; Volume 9, p. 447. [Google Scholar]
- Hernot, S.; Klibanov, A.L. Microbubbles in Ultrasound-Triggered Drug and Gene Delivery. Adv. Drug Deliv. Rev. 2008, 60, 1153–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zasadzinski, J.A. Transmission Electron Microscopy Observations of Sonication-Induced Changes in Liposome Structure. Biophys. J. 1986, 49, 1119–1130. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.-Y.; Thomas, J.L. PEG-Lipids and Oligo (Ethylene Glycol) Surfactants Enhance the Ultrasonic Permeabilizability of Liposomes. Langmuir 2003, 19, 1098–1105. [Google Scholar] [CrossRef]
- Lin, H.-Y.; Thomas, J.L. Factors Affecting Responsivity of Unilamellar Liposomes to 20 KHz Ultrasound. Langmuir 2004, 20, 6100–6106. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, A.; Honen, R.; Turjeman, K.; Gabizon, A.; Kost, J.; Barenholz, Y. Ultrasound Triggered Release of Cisplatin from Liposomes in Murine Tumors. J. Control. Release 2009, 137, 63–68. [Google Scholar] [CrossRef]
- Schroeder, A.; Avnir, Y.; Weisman, S.; Najajreh, Y.; Gabizon, A.; Talmon, Y.; Kost, J.; Barenholz, Y. Controlling Liposomal Drug Release with Low Frequency Ultrasound: Mechanism and Feasibility. Langmuir 2007, 23, 4019–4025. [Google Scholar] [CrossRef]
- Chen, W.; Du, J. Ultrasound and PH Dually Responsive Polymer Vesicles for Anticancer Drug Delivery. Sci. Rep. 2013, 3, srep02162. [Google Scholar] [CrossRef] [Green Version]
- Evjen, T.J.; Nilssen, E.A.; Rögnvaldsson, S.; Brandl, M.; Fossheim, S.L. Distearoylphosphatidylethanolamine-Based Liposomes for Ultrasound-Mediated Drug Delivery. Eur. J. Pharm. Biopharm. 2010, 75, 327–333. [Google Scholar] [CrossRef] [Green Version]
- Kopechek, J.A.; Abruzzo, T.M.; Wang, B.; Chrzanowski, S.M.; Smith, D.A.B.; Kee, P.H.; Huang, S.; Collier, J.H.; McPherson, D.D.; Holland, C.K. Ultrasound-Mediated Release of Hydrophilic and Lipophilic Agents from Echogenic Liposomes. J. Ultrasound Med. 2008, 27, 1597–1606. [Google Scholar] [CrossRef] [Green Version]
- Pong, M.; Umchid, S.; Guarino, A.J.; Lewin, P.A.; Litniewski, J.; Nowicki, A.; Wrenn, S.P. In Vitro Ultrasound-Mediated Leakage from Phospholipid Vesicles. Ultrasonics 2006, 45, 133–145. [Google Scholar] [CrossRef]
- Needham, D.; Hristova, K.; McIntosh, T.J.; Dewhirst, M.; Wu, N.; Lasic, D.D.; Alving, C.R.; Wassef, N.M.; Senior, J.H.; Ghosh, P.C.; et al. Polymer-Grafted Liposomes: Physical Basis for the “Stealth” Property. J. Liposome Res. 1992, 2, 411–430. [Google Scholar] [CrossRef]
- Dewhirst, M.W.; Needham, D. Extravasation of Stealth Liposomes into Tumors: Direct Measurement of Accumulation and Vascular Permeability Using a Skin-Flap Window Chamber. In Stealth Liposomes; Taylor & Francis Group: London, UK, 1995; pp. 127–137. [Google Scholar]
- Anyarambhatla, G.R.; Needham, D. Enhancement of the Phase Transition Permeability of DPPC Liposomes by Incorporation of MPPC: A New Temperature-Sensitive Liposome for Use with Mild Hyperthermia. J. Liposome Res. 1999, 9, 491–506. [Google Scholar] [CrossRef]
- Needham, D.; Anyarambhatla, G.; Kong, G.; Dewhirst, M.W. A New Temperature-Sensitive Liposome for Use with Mild Hyperthermia: Characterization and Testing in a Human Tumor Xenograft Model. Cancer Res. 2000, 60, 1197–1201. [Google Scholar] [PubMed]
- Kong, G.; Anyarambhatla, G.; Petros, W.P.; Braun, R.D.; Colvin, O.M.; Needham, D.; Dewhirst, M.W. Efficacy of Liposomes and Hyperthermia in a Human Tumor Xenograft Model: Importance of Triggered Drug Release. Cancer Res. 2000, 60, 6950–6957. [Google Scholar]
- Mills, J.K.; Needham, D. Lysolipid Incorporation in Dipalmitoylphosphatidylcholine Bilayer Membranes Enhances the Ion Permeability and Drug Release Rates at the Membrane Phase Transition. Biochim. Biophys. Acta Biomembr. 2005, 1716, 77–96. [Google Scholar] [CrossRef] [Green Version]
- Moon, H.; You, W.; Kim, Y.-S.; Kim, H.R.; Jung, E.A. Fabrication of Ultrasound-Responsive Liposomes. Korean Patent No. 10-2336114, 2 December 2021. [Google Scholar]
- Kim, Y.-S.; Ko, M.J.; Moon, H.; Sim, W.; Cho, A.S.; Gil, G.; Kim, H.R. Ultrasound-Responsive Liposomes for Targeted Drug Delivery Combined with Focused Ultrasound. Pharmaceutics 2022, 14, 1314. [Google Scholar] [CrossRef]
- Lindahl, E.; Hess, B.; van der Spoel, D. GROMACS 3.0: A Package for Molecular Simulation and Trajectory Analysis. J. Mol. Model. 2001, 7, 306–317. [Google Scholar] [CrossRef]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J.C. GROMACS: Fast, Flexible, and Free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef]
- Hess, B.; Kutzner, C.; van der Spoel, D.; Lindahl, E. GROMACS 4: Algorithms for Highly Efficient, Load-Balanced, and Scalable Molecular Simulation. J. Chem. Theory Comput. 2008, 4, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Marrink, S.J.; de Vries, A.H.; Mark, A.E. Coarse Grained Model for Semiquantitative Lipid Simulations. J. Phys. Chem. B 2004, 108, 750–760. [Google Scholar] [CrossRef] [Green Version]
- Marrink, S.J.; Risselada, H.J.; Yefimov, S.; Tieleman, D.P.; de Vries, A.H. The MARTINI Force Field: Coarse Grained Model for Biomolecular Simulations. J. Phys. Chem. B 2007, 111, 7812–7824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Venable, R.M.; MacKerell, A.D.; Pastor, R.W. Molecular Dynamics Studies of Polyethylene Oxide and Polyethylene Glycol: Hydrodynamic Radius and Shape Anisotropy. Biophys. J. 2008, 95, 1590–1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; de Vries, A.H.; Marrink, S.J.; Pastor, R.W. A Coarse-Grained Model for Polyethylene Oxide and Polyethylene Glycol: Conformation and Hydrodynamics. J. Phys. Chem. B 2009, 113, 13186–13194. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Pastor, R.W. Coarse-Grained Model for PEGylated Lipids: Effect of PEGylation on the Size and Shape of Self-Assembled Structures. J. Phys. Chem. B 2011, 115, 7830–7837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Larson, R.G. Adsorption of Plasma Proteins onto PEGylated Lipid Bilayers: The Effect of PEG Size and Grafting Density. Biomacromolecules 2016, 17, 1757–1765. [Google Scholar] [CrossRef] [Green Version]
- Lee, H. Molecular Dynamics Studies of PEGylated Single-Walled Carbon Nanotubes: The Effect of PEG Size and Grafting Density. J. Phys. Chem. C 2013, 117, 26334–26341. [Google Scholar] [CrossRef]
- Lee, H.; Larson, R.G. Effects of PEGylation on the Size and Internal Structure of Dendrimers: Self-Penetration of Long PEG Chains into the Dendrimer Core. Macromolecules 2011, 44, 2291–2298. [Google Scholar] [CrossRef]
- Bussi, G.; Donadio, D.; Parrinello, M. Canonical Sampling through Velocity Rescaling. J. Chem. Phys. 2007, 126, 014101. [Google Scholar] [CrossRef] [Green Version]
- Parrinello, M.; Rahman, A. Polymorphic Transitions in Single Crystals: A New Molecular Dynamics Method. J. Appl. Phys. 1981, 52, 7182–7190. [Google Scholar] [CrossRef]
- Marrink, S.J.; Risselada, J.; Mark, A.E. Simulation of Gel Phase Formation and Melting in Lipid Bilayers Using a Coarse Grained Model. Chem. Phys. Lipids 2005, 135, 223–244. [Google Scholar] [CrossRef] [Green Version]
- Koynova, R.; Caffrey, M. Phases and Phase Transitions of the Phosphatidylcholines. Biochim. Biophys. Acta-Rev. Biomembr. 1998, 1376, 91–145. [Google Scholar] [CrossRef]
- Hess, B.; Bekker, H.; Berendsen, H.J.C.; Fraaije, J.G.E.M. LINCS: A Linear Constraint Solver for Molecular Simulations. J. Comput. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]
- Hess, B. P-LINCS: A Parallel Linear Constraint Solver for Molecular Simulation. J. Chem. Theory Comput. 2008, 4, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual Molecular Dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Filippov, A.; Orädd, G.; Lindblom, G. The Effect of Cholesterol on the Lateral Diffusion of Phospholipids in Oriented Bilayers. Biophys. J. 2003, 84, 3079–3086. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Kim, H.R.; Park, J.C. Dynamics and Stability of Lipid Bilayers Modulated by Thermosensitive Polypeptides, Cholesterols, and PEGylated Lipids. Phys. Chem. Chem. Phys. 2014, 16, 3763–3770. [Google Scholar] [CrossRef]
- Park, S.M.; Kim, M.S.; Park, S.; Park, E.S.; Choi, K.; Kim, Y.; Kim, H.R. Novel Temperature-Triggered Liposome with High Stability: Formulation, in Vitro Evaluation, and in Vivo Study Combined with High-Intensity Focused Ultrasound (HIFU). J. Control. Release 2013, 170, 373–379. [Google Scholar] [CrossRef]
- Hofsäß, C.; Lindahl, E.; Edholm, O. Molecular Dynamics Simulations of Phospholipid Bilayers with Cholesterol. Biophys. J. 2003, 84, 2192–2206. [Google Scholar] [CrossRef] [Green Version]
- Israelachvili, J.N.; Mitchell, D.J. A Model for the Packing of Lipids in Bilayer Membranes. Biochim. Biophys. Acta BBA Biomembr. 1975, 389, 13–19. [Google Scholar] [CrossRef]
- Hyslop, P.A.; Morel, B.; Sauerheber, R.D. Organization and Interaction of Cholesterol and Phosphatidylcholine in Model Bilayer Membranes. Biochemistry 1990, 29, 1025–1038. [Google Scholar] [CrossRef]
- Hakobyan, D.; Heuer, A. Key Molecular Requirements for Raft Formation in Lipid/Cholesterol Membranes. PLoS ONE 2014, 9, e87369. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Huang, G.; Leal, C. Role of Lipid Polymorphism in Acoustically Sensitive Liposomes. Soft Matter 2014, 10, 8846–8854. [Google Scholar] [CrossRef] [PubMed]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Simulation System No. | No. of Membrane Components | Ratio | |||||
|---|---|---|---|---|---|---|---|
| DSPC | DSPE-PEG | Chol. | DOPE | DOPC | MSPC | ||
| 1 | 256 | 128 | - | 1664 | - | 128 | 10:5:0:65:5 |
| 2 | 256 | 128 | 384 | 1664 | - | 128 | 10:5:15:65:5 |
| 3 (IMP301) | 256 | 128 | 768 | 1664 | - | 128 | 10:5:30:65:5 |
| 4 | 256 | 128 | 1024 | 1664 | - | 128 | 10:5:40:65:5 |
| 5 | 256 | 128 | 768 | 1664 | - | - | 10:5:30:65:0 |
| 6 | 256 | 128 | 768 | 1664 | - | 256 | 10:5:30:65:10 |
| 7 | 256 | 128 | 768 | - | 1664 | 128 | 10:5:30:65:5 |
| 8 | 1920 | 128 | 768 | - | - | 128 | 75:5:30:0:5 |
| 9 | 1024 | 512 | 3072 | 6656 | - | 512 | 10:5:30:65:5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.; Moon, H.; Kim, H.-R. Effects of Lipid Shape and Interactions on the Conformation, Dynamics, and Curvature of Ultrasound-Responsive Liposomes. Pharmaceutics 2022, 14, 1512. https://doi.org/10.3390/pharmaceutics14071512
Lee H, Moon H, Kim H-R. Effects of Lipid Shape and Interactions on the Conformation, Dynamics, and Curvature of Ultrasound-Responsive Liposomes. Pharmaceutics. 2022; 14(7):1512. https://doi.org/10.3390/pharmaceutics14071512
Chicago/Turabian StyleLee, Hwankyu, Hyungwon Moon, and Hyun-Ryoung Kim. 2022. "Effects of Lipid Shape and Interactions on the Conformation, Dynamics, and Curvature of Ultrasound-Responsive Liposomes" Pharmaceutics 14, no. 7: 1512. https://doi.org/10.3390/pharmaceutics14071512