
Cardiac Sodium/Hydrogen Exchanger (NHE11) as a Novel Potential Target for SGLT2i in Heart Failure: A Preliminary Study
 ,
,  , and
, and
Abstract
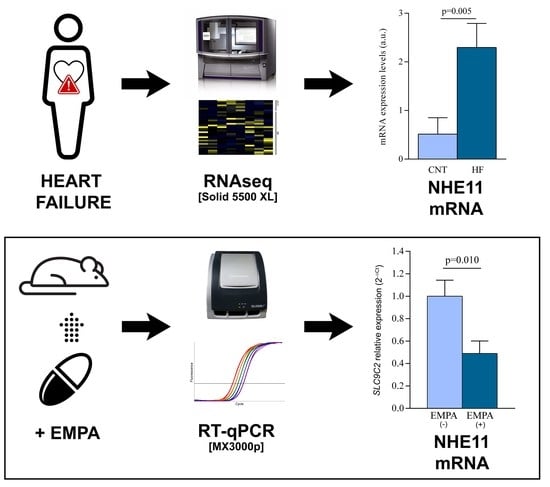
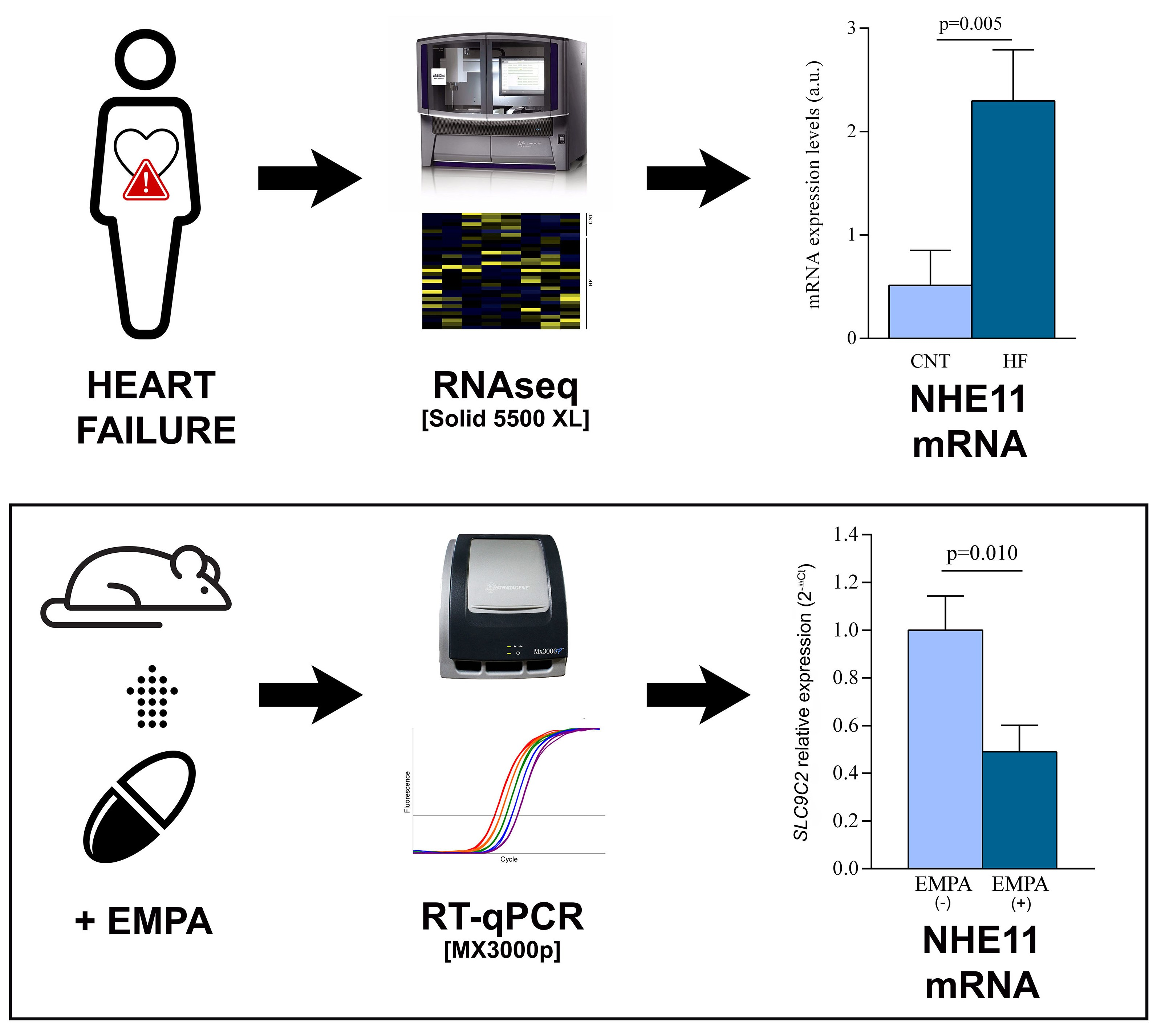
:
1. Introduction
2. Materials and Methods
2.1. Human Sample Collection
2.2. Transcriptomic Analysis
2.3. NHE11 and NHE1 Protein Concentration
2.4. In Vivo Study
2.5. Statistical Analysis
3. Results
3.1. Human Left Ventricle mRNA Expression of the Main Sodium Channels
3.2. Human Protein Expression of NHE11 and NHE1
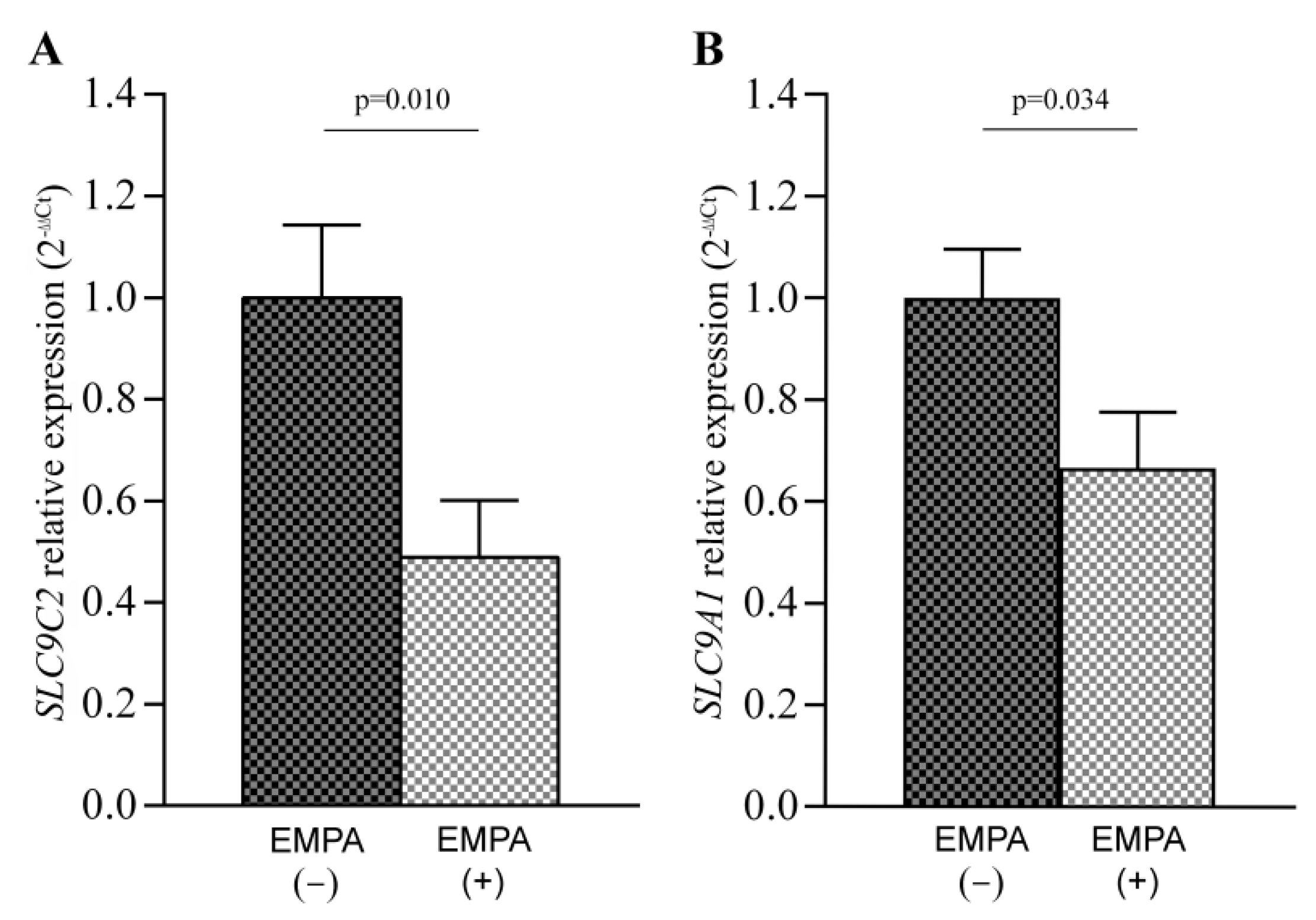
3.3. SLC9C2 (NHE11) and SLC9A1 (NHE1) mRNA Levels in Empagliflozin-Treated Rats
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosello-Lleti, E.; Tarazon, E.; Ortega, A.; Gil-Cayuela, C.; Carnicer, R.; Lago, F.; Gonzalez-Juanatey, J.R.; Portoles, M.; Rivera, M. Protein Inhibitor of NOS1 Plays a Central Role in the Regulation of NOS1 Activity in Human Dilated Hearts. Sci. Rep. 2016, 6, 30902. [Google Scholar] [CrossRef] [PubMed]
- Nabeebaccus, A.; Zheng, S.; Shah, A.M. Heart failure-potential new targets for therapy. Br. Med. Bull. 2016, 119, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Zelniker, T.A.; Wiviott, S.D.; Raz, I.; Im, K.; Goodrich, E.L.; Bonaca, M.P.; Mosenzon, O.; Kato, E.T.; Cahn, A.; Furtado, R.H.M.; et al. SGLT2 inhibitors for primary and secondary prevention of cardiovascular and renal outcomes in type 2 diabetes: A systematic review and meta-analysis of cardiovascular outcome trials. Lancet 2019, 393, 31–39. [Google Scholar] [CrossRef]
- Brito, D.; Bettencourt, P.; Carvalho, D.; Ferreira, J.; Fontes-Carvalho, R.; Franco, F.; Moura, B.; Silva-Cardoso, J.C.; de Melo, R.T.; Fonseca, C. Sodium-Glucose Co-transporter 2 Inhibitors in the Failing Heart: A Growing Potential. Cardiovasc. Drugs Ther. 2020, 34, 419–436. [Google Scholar] [CrossRef]
- Santos-Gallego, C.G.; Garcia-Ropero, A.; Mancini, D.; Pinney, S.P.; Contreras, J.P.; Fergus, I.; Abascal, V.; Moreno, P.; Atallah-Lajam, F.; Tamler, R.; et al. Rationale and Design of the EMPA-TROPISM Trial (ATRU-4): Are the “Cardiac Benefits” of Empagliflozin Independent of its Hypoglycemic Activity? Cardiovasc. Drugs Ther. 2019, 33, 87–95. [Google Scholar] [CrossRef]
- Baartscheer, A.; Schumacher, C.A.; Wust, R.C.; Fiolet, J.W.; Stienen, G.J.; Coronel, R.; Zuurbier, C.J. Empagliflozin decreases myocardial cytoplasmic Na(+) through inhibition of the cardiac Na(+)/H(+) exchanger in rats and rabbits. Diabetologia 2017, 60, 568–573. [Google Scholar] [CrossRef]
- Uthman, L.; Baartscheer, A.; Bleijlevens, B.; Schumacher, C.A.; Fiolet, J.W.T.; Koeman, A.; Jancev, M.; Hollmann, M.W.; Weber, N.C.; Coronel, R.; et al. Class effects of SGLT2 inhibitors in mouse cardiomyocytes and hearts: Inhibition of Na(+)/H(+) exchanger, lowering of cytosolic Na(+) and vasodilation. Diabetologia 2018, 61, 722–726. [Google Scholar] [CrossRef]
- Packer, M. Reconceptualization of the Molecular Mechanism by Which Sodium-Glucose Cotransporter 2 Inhibitors Reduce the Risk of Heart Failure Events. Circulation 2019, 140, 443–445. [Google Scholar] [CrossRef]
- Zuurbier, C.J.; Baartscheer, A.; Schumacher, C.A.; Fiolet, J.W.T.; Coronel, R. Sodium-glucose co-transporter 2 inhibitor empagliflozin inhibits the cardiac Na+/H+ exchanger 1: Persistent inhibition under various experimental conditions. Cardiovasc. Res. 2021, 117, 2699–2701. [Google Scholar] [CrossRef]
- Chung, Y.J.; Park, K.C.; Tokar, S.; Eykyn, T.R.; Fuller, W.; Pavlovic, D.; Swietach, P.; Shattock, M.J. Off-target effects of sodium-glucose co-transporter 2 blockers: Empagliflozin does not inhibit Na+/H+ exchanger-1 or lower [Na+]i in the heart. Cardiovasc. Res. 2021, 117, 2794–2806. [Google Scholar] [CrossRef]
- Donowitz, M.; Ming Tse, C.; Fuster, D. SLC9/NHE gene family, a plasma membrane and organellar family of Na(+)/H(+) exchangers. Mol. Aspects Med. 2013, 34, 236–251. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; King, S.M.; Quill, T.A.; Doolittle, L.K.; Garbers, D.L. A new sperm-specific Na+/H+ exchanger required for sperm motility and fertility. Nat. Cell Biol. 2003, 5, 1117–1122. [Google Scholar] [CrossRef] [PubMed]
- Sayour, A.A.; Olah, A.; Ruppert, M.; Barta, B.A.; Horvath, E.M.; Benke, K.; Polos, M.; Hartyanszky, I.; Merkely, B.; Radovits, T. Characterization of left ventricular myocardial sodium-glucose cotransporter 1 expression in patients with end-stage heart failure. Cardiovasc. Diabetol. 2020, 19, 159. [Google Scholar] [CrossRef] [PubMed]
- Ponikowski, P.; Voors, A.A.; Anker, S.D.; Bueno, H.; Cleland, J.G.; Coats, A.J.; Falk, V.; Gonzalez-Juanatey, J.R.; Harjola, V.P.; Jankowska, E.A.; et al. 2016 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure: The Task Force for the diagnosis and treatment of acute and chronic heart failure of the European Society of Cardiology (ESC). Developed with the special contribution of the Heart Failure Association (HFA) of the ESC. Eur. J. Heart Fail. 2016, 18, 891–975. [Google Scholar]
- Harbo, M.B.; Norden, E.S.; Narula, J.; Sjaastad, I.; Espe, E.K.S. Quantifying left ventricular function in heart failure: What makes a clinically valuable parameter? Prog. Cardiovasc. Dis. 2020, 63, 552–560. [Google Scholar] [CrossRef]
- Macrae, D.J. The Council for International Organizations and Medical Sciences (CIOMS) guidelines on ethics of clinical trials. Proc. Am. Thorac. Soc. 2007, 4, 176–179. [Google Scholar] [CrossRef]
- Rosello-Lleti, E.; Tarazon, E.; Barderas, M.G.; Ortega, A.; Molina-Navarro, M.M.; Martinez, A.; Lago, F.; Martinez-Dolz, L.; Gonzalez-Juanatey, J.R.; Salvador, A.; et al. ATP synthase subunit alpha and LV mass in ischaemic human hearts. J. Cell Mol. Med. 2015, 19, 442–451. [Google Scholar] [CrossRef]
- Rosello-Lleti, E.; Carnicer, R.; Tarazon, E.; Ortega, A.; Gil-Cayuela, C.; Lago, F.; Gonzalez-Juanatey, J.R.; Portoles, M.; Rivera, M. Human Ischemic Cardiomyopathy Shows Cardiac Nos1 Translocation and its Increased Levels are Related to Left Ventricular Performance. Sci. Rep. 2016, 6, 24060. [Google Scholar] [CrossRef]
- Aragon-Herrera, A.; Feijoo-Bandin, S.; Otero Santiago, M.; Barral, L.; Campos-Toimil, M.; Gil-Longo, J.; Costa Pereira, T.M.; Garcia-Caballero, T.; Rodriguez-Segade, S.; Rodriguez, J.; et al. Empagliflozin reduces the levels of CD36 and cardiotoxic lipids while improving autophagy in the hearts of Zucker diabetic fatty rats. Biochem. Pharmacol. 2019, 170, 113677. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Moon, K.H. Screening of Genetic Factor in the Interaction Between Periodontitis and Metabolic Traits Using Candidate Gene Association Study (CGAS). Biochem. Genet. 2019, 57, 466–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Y.; Jia, X.; Bajaj, M.; Birnbaum, Y. Dapagliflozin Attenuates Na(+)/H(+) Exchanger-1 in Cardiofibroblasts via AMPK Activation. Cardiovasc. Drugs Ther. 2018, 32, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Goerg, J.; Sommerfeld, M.; Greiner, B.; Lauer, D.; Seckin, Y.; Kulikov, A.; Ivkin, D.; Kintscher, U.; Okovityi, S.; Kaschina, E. Low-Dose Empagliflozin Improves Systolic Heart Function after Myocardial Infarction in Rats: Regulation of MMP9, NHE1, and SERCA2a. Int. J. Mol. Sci. 2021, 22, 5437. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, H.; Gunasegaram, S.; Harding, S.E.; Avkiran, M. Sarcolemmal Na+/H+ exchanger activity and expression in human ventricular myocardium. J. Am. Coll. Cardiol. 2000, 36, 534–540. [Google Scholar] [CrossRef]
- Santos-Gallego, C.G.; Requena-Ibanez, J.A.; San Antonio, R.; Ishikawa, K.; Watanabe, S.; Picatoste, B.; Flores, E.; Garcia-Ropero, A.; Sanz, J.; Hajjar, R.J.; et al. Empagliflozin Ameliorates Adverse Left Ventricular Remodeling in Nondiabetic Heart Failure by Enhancing Myocardial Energetics. J. Am. Coll. Cardiol. 2019, 73, 1931–1944. [Google Scholar] [CrossRef] [PubMed]
- Feijóo-Bandín, S.; Aragón-Herrera, A.; Otero-Santiago, M.; Anido-Varela, L.; Moraña-Fernández, S.; Tarazón, E.; Roselló-Lletí, E.; Portolés, M.; Gualillo, O.; González-Juanatey, J.R.; et al. Role of Sodium-Glucose Co-Transporter 2 Inhibitors in the Regulation of Inflammatory Processes in Animal Models. Int. J. Mol. Sci. 2022, 23, 5634. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Carrillo, L.; Giménez-Escamilla, I.; Martínez-Dolz, L.; Sánchez-Lázaro, I.J.; Portolés, M.; Roselló-Lletí, E.; Tarazón, E. Implication of Sphingolipid Metabolism Gene Dysregulation and Cardiac Sphingosine-1-Phosphate Accumulation in Heart Failure. Biomedicines 2022, 10, 135. [Google Scholar] [CrossRef]
- Sauvage, M.; Maziere, P.; Fathallah, H.; Giraud, F. Insulin stimulates NHE1 activity by sequential activation of phosphatidylinositol 3-kinase and protein kinase C zeta in human erythrocytes. Eur. J. Biochem. 2000, 267, 955–962. [Google Scholar] [CrossRef]
- Prasad, V.; Lorenz, J.N.; Miller, M.L.; Vairamani, K.; Nieman, M.L.; Wang, Y.; Shull, G.E. Loss of NHE1 activity leads to reduced oxidative stress in heart and mitigates high-fat diet-induced myocardial stress. J. Mol. Cell Cardiol. 2013, 65, 33–42. [Google Scholar] [CrossRef]
- Vaduganathan, M.; Inzucchi, S.E.; Sattar, N.; Fitchett, D.H.; Ofstad, A.P.; Brueckmann, M.; George, J.T.; Verma, S.; Mattheus, M.; Wanner, C.; et al. Effects of empagliflozin on insulin initiation or intensification in patients with type 2 diabetes and cardiovascular disease: Findings from the EMPA-REG OUTCOME trial. Diabetes Obes. Metab. 2021, 23, 2775–2784. [Google Scholar] [CrossRef]
- Uthman, L.; Li, X.; Baartscheer, A.; Schumacher, C.A.; Baumgart, P.; Hermanides, J.; Preckel, B.; Hollmann, M.W.; Coronel, R.; Zuurbier, C.J.; et al. Empagliflozin reduces oxidative stress through inhibition of the novel inflammation/NHE/[Na(+)]c/ROS-pathway in human endothelial cells. Biomed. Pharmacother. 2022, 146, 112515. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Li, T.; Wang, Y.; Chang, Y.; Cheng, Y.; Lu, Y.; Liu, X.; Xu, L.; Li, X.; Yu, X.; et al. Empagliflozin prevents cardiomyopathy via sGC-cGMP-PKG pathway in type 2 diabetes mice. Clin. Sci. 2019, 133, 1705–1720. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Cryan, E.V.; D’Andrea, M.R.; Belkowski, S.; Conway, B.R.; Demarest, K.T. Human cardiomyocytes express high level of Na+/glucose cotransporter 1 (SGLT1). J. Cell Biochem. 2003, 90, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Williams, S.; Ho, S.; Loraine, H.; Hagan, D.; Whaley, J.M.; Feder, J.N. Quantitative PCR tissue expression profiling of the human SGLT2 gene and related family members. Diabetes Ther. 2010, 1, 57–92. [Google Scholar] [CrossRef]
- Von Lewinski, D.; Rainer, P.P.; Gasser, R.; Huber, M.S.; Khafaga, M.; Wilhelm, B.; Haas, T.; Machler, H.; Rossl, U.; Pieske, B. Glucose-transporter-mediated positive inotropic effects in human myocardium of diabetic and nondiabetic patients. Metabolism 2010, 59, 1020–1028. [Google Scholar] [CrossRef]
- Banerjee, S.K.; McGaffin, K.R.; Pastor-Soler, N.M.; Ahmad, F. SGLT1 is a novel cardiac glucose transporter that is perturbed in disease states. Cardiovasc. Res. 2009, 84, 111–118. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RNA-Seq Analysis | Protein Analysis | |
|---|---|---|
| HF (n = 26) | HF (n = 70) | |
| Age (years) | 53 ± 10 | 54 ± 10 |
| Gender male (%) | 96 | 84 |
| NYHA class | III–IV | III–IV |
| BMI (kg/m2) | 27 ± 5 | 26 ± 5 |
| Hypercholesterolemia (%) | 13 | 21 |
| Prior hypertension (%) | 25 | 38 |
| Prior type 2 diabetes (%) | 29 | 35 |
| Hemoglobin (mg/mL) | 14 ± 3 | 13 ± 2 |
| Hematocrit (%) | 40 ± 7 | 39 ± 6 |
| LVEF (%) | 21 ± 8 | 23 ± 8 |
| LVESD (mm) | 66 ± 12 | 60 ± 11 |
| LVEDD (mm) | 74 ± 11 | 68 ± 10 |
| LVESD | LVEDD | |
|---|---|---|
| NHE11 | r = 0.334 p = 0.011 | r = 0.290 p = 0.029 |
| NHE1 | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Carrillo, L.; Aragón-Herrera, A.; Giménez-Escamilla, I.; Delgado-Arija, M.; García-Manzanares, M.; Anido-Varela, L.; Lago, F.; Martínez-Dolz, L.; Portolés, M.; Tarazón, E.; et al. Cardiac Sodium/Hydrogen Exchanger (NHE11) as a Novel Potential Target for SGLT2i in Heart Failure: A Preliminary Study. Pharmaceutics 2022, 14, 1996. https://doi.org/10.3390/pharmaceutics14101996
Pérez-Carrillo L, Aragón-Herrera A, Giménez-Escamilla I, Delgado-Arija M, García-Manzanares M, Anido-Varela L, Lago F, Martínez-Dolz L, Portolés M, Tarazón E, et al. Cardiac Sodium/Hydrogen Exchanger (NHE11) as a Novel Potential Target for SGLT2i in Heart Failure: A Preliminary Study. Pharmaceutics. 2022; 14(10):1996. https://doi.org/10.3390/pharmaceutics14101996
Chicago/Turabian StylePérez-Carrillo, Lorena, Alana Aragón-Herrera, Isaac Giménez-Escamilla, Marta Delgado-Arija, María García-Manzanares, Laura Anido-Varela, Francisca Lago, Luis Martínez-Dolz, Manuel Portolés, Estefanía Tarazón, and et al. 2022. "Cardiac Sodium/Hydrogen Exchanger (NHE11) as a Novel Potential Target for SGLT2i in Heart Failure: A Preliminary Study" Pharmaceutics 14, no. 10: 1996. https://doi.org/10.3390/pharmaceutics14101996