
The Fast Track for Intestinal Tumor Cell Differentiation and In Vitro Intestinal Models by Inorganic Topographic Surfaces

Abstract
:1. Introduction
2. Materials and Methods
2.1. Topographic Surfaces
2.2. Cell Culture
Culture on Topographic Surfaces
2.3. Imaging
2.3.1. Trypan Blue Exclusion Assay for Dead Cells
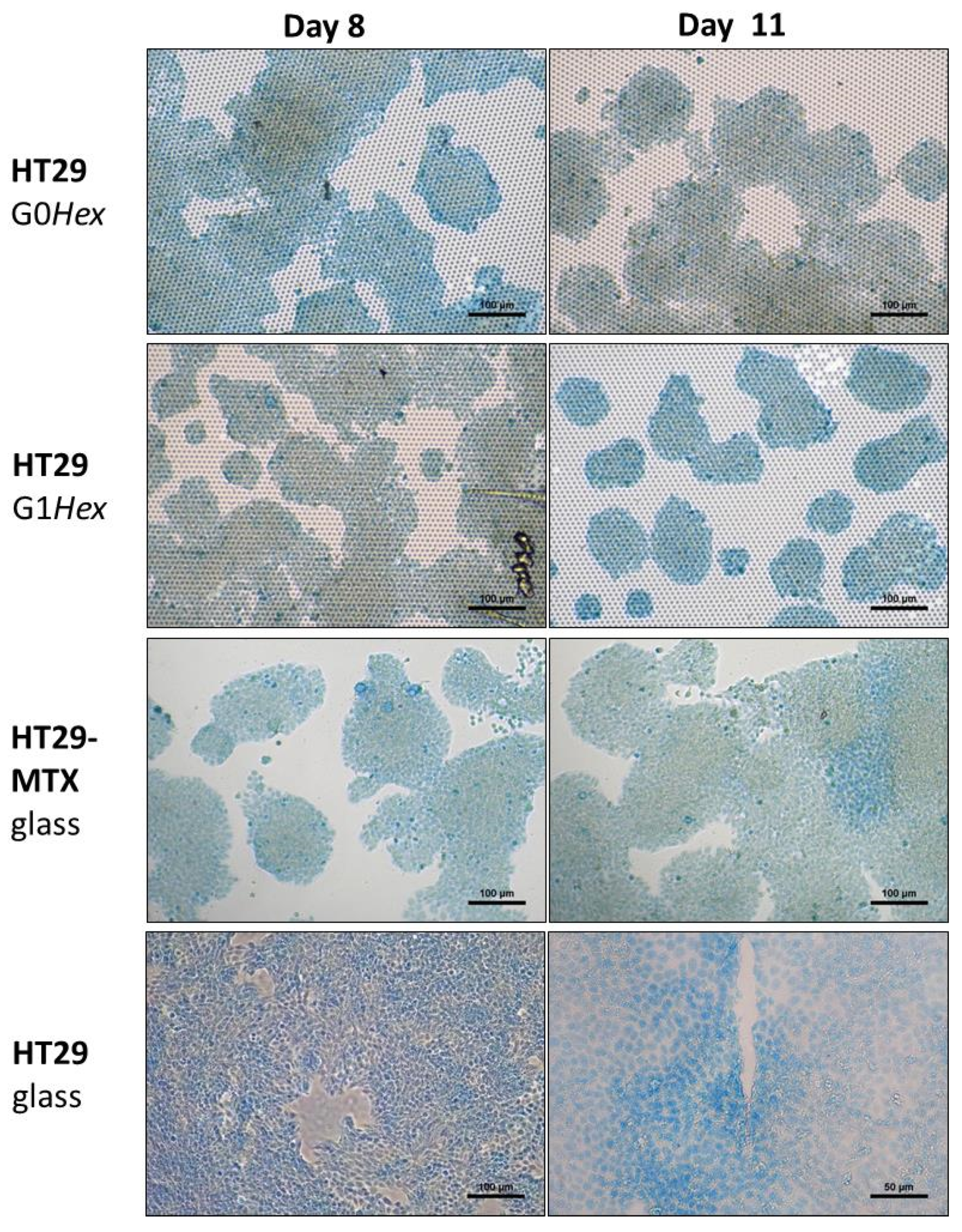
2.3.2. Alcian Blue Staining for Mucus
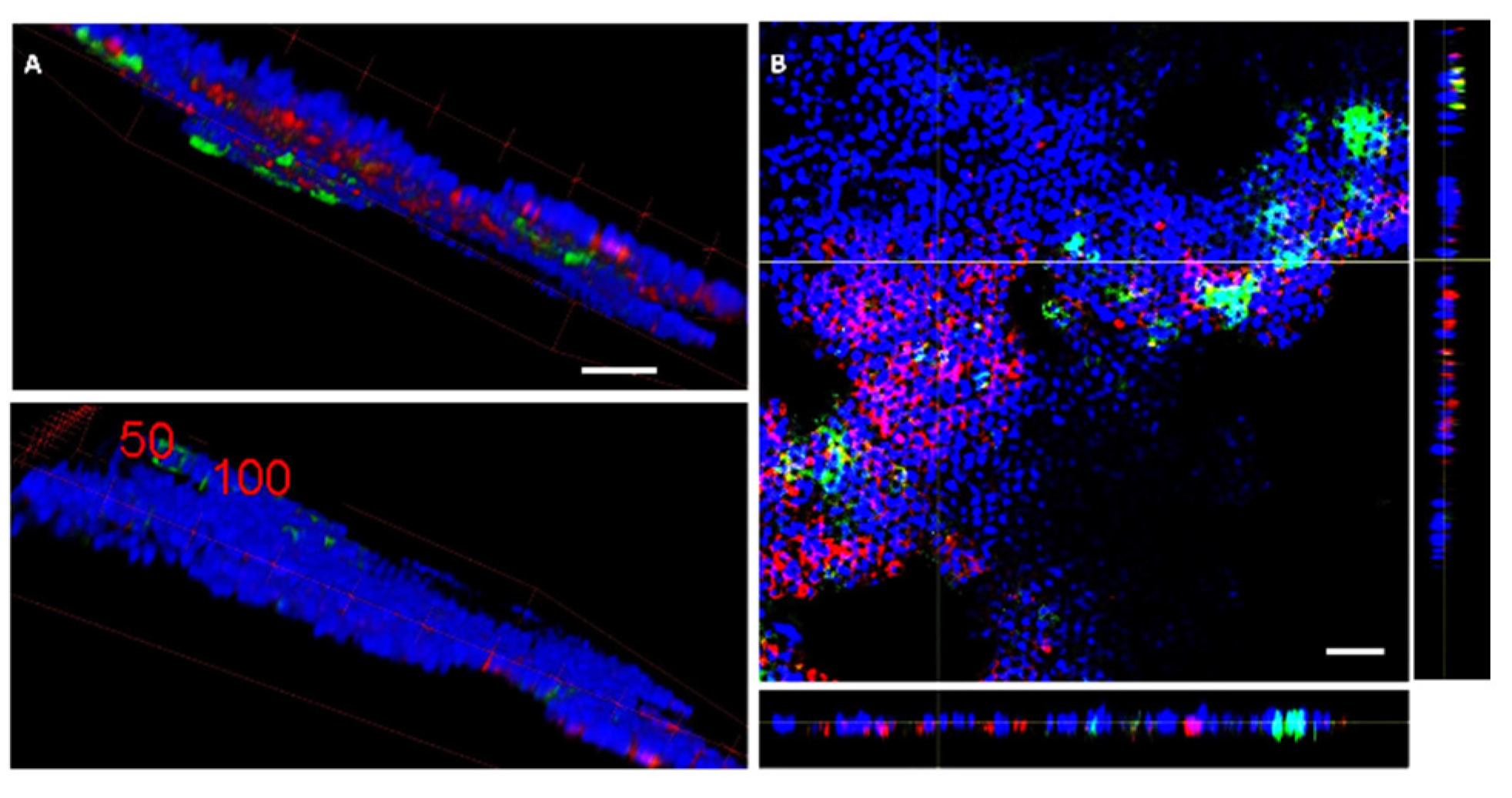
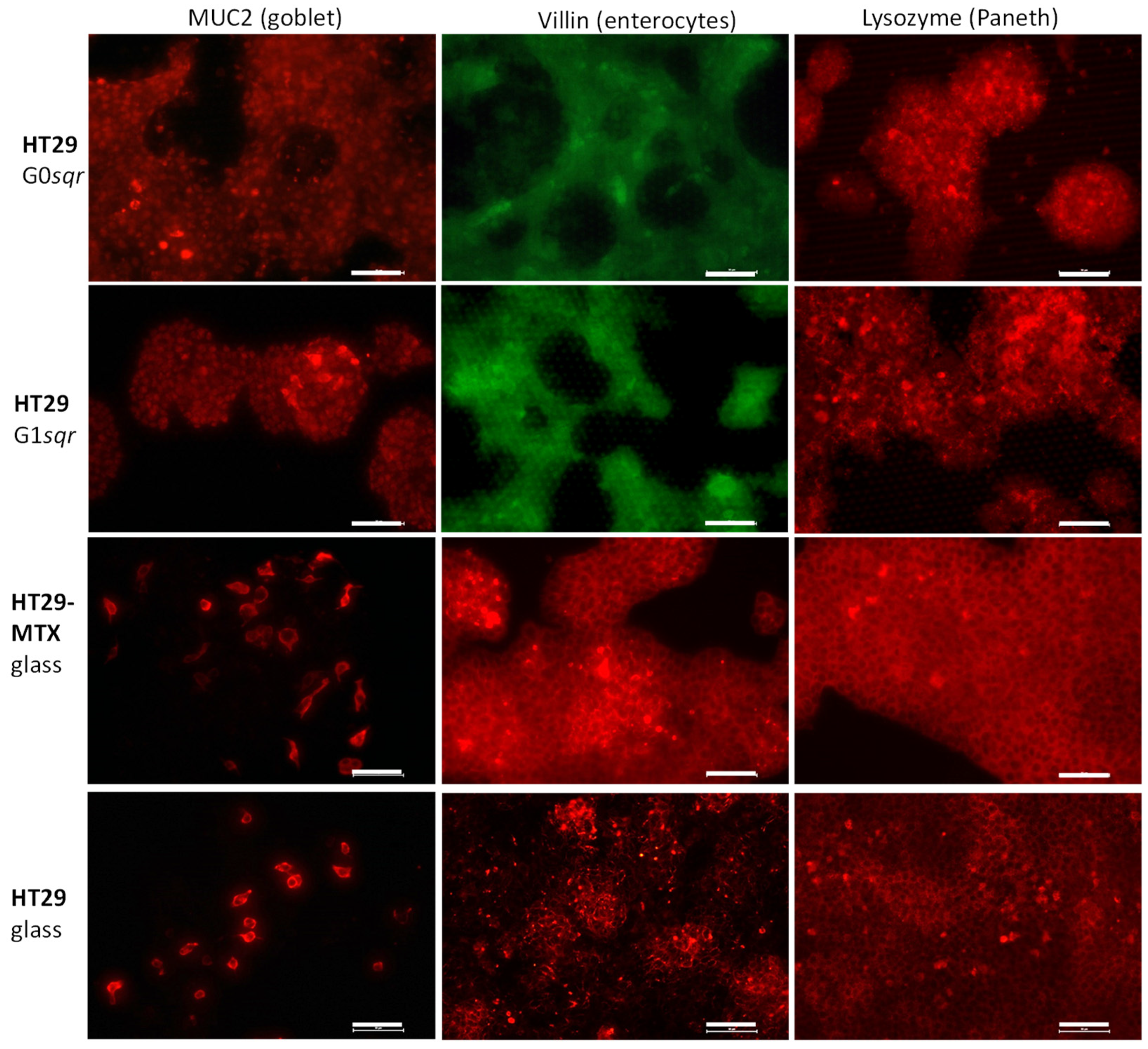
2.3.3. Fluorescence Microscopy
2.3.4. Staining Protocols
2.3.5. Image Analysis
3. Results
Mucus Production
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jensen, C.; Teng, Y. Is It Time to Start Transitioning From 2D to 3D Cell Culture? Front. Mol. Biosci. 2020, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Co, J.Y.; Margalef-Català, M.; Li, X.; Mah, A.T.; Kuo, C.J.; Monack, D.M.; Amieva, M.R. Controlling Epithelial Polarity: A Human Enteroid Model for Host-Pathogen Interactions. Cell Rep. 2019, 26, 2509–2520. [Google Scholar] [CrossRef] [PubMed]
- Finkbeiner, S.R.; Spence, J.R. A gutsy task: Generating intestinal tissue from human pluripotent stem cells. Dig. Dis. Sci. 2013, 58, 1176–1184. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Eglen, R.M. Three-Dimensional Cell Cultures in Drug Discovery and Development. SLAS Discov. Adv. Life Sci. RD 2017, 22, 456–472. [Google Scholar] [CrossRef]
- Dosh, R.H.; Jordan-Mahy, N.; Sammon, C.; Le Maitre, C.L. Tissue Engineering Laboratory Models of the Small Intestine. Tissue Eng. Part B Rev. 2018, 24, 98–111. [Google Scholar] [CrossRef]
- Merker, S.R.; Weitz, J.; Stange, D.E. Gastrointestinal organoids: How they gut it out. Dev. Biol. 2016, 420, 239–250. [Google Scholar] [CrossRef]
- Kasendra, M.; Luc, R.; Yin, J.; Manatakis, D.V.; Kulkarni, G.; Lucchesi, C.; Sliz, J.; Apostolou, A.; Sunuwar, L.; Obrigewitch, J.; et al. Duodenum Intestine-Chip for preclinical drug assessment in a human relevant model. eLife 2020, 9, e50135. [Google Scholar] [CrossRef]
- Pleguezuelos-Manzano, C.; Puschhof, J.; den Brink, S.; Geurts, V.; Beumer, J.; Clevers, H. Establishment and Culture of Human Intestinal Organoids Derived from Adult Stem Cells. Curr. Protoc. Immunol. 2020, 130, e106. [Google Scholar] [CrossRef]
- Lu, X.; Walker, T.; MacManus, J.P.; Seligy, V.L. Differentiation of HT-29 human colonic adenocarcinoma cells correlates with increased expression of mitochondrial RNA: Effects of trehalose on cell growth and maturation. Cancer Res. 1992, 52, 3718–3725. [Google Scholar]
- Cohen, E.; Ophir, I.; Shaul, Y.B. Induced differentiation in HT29, a human colon adenocarcinoma cell line. J. Cell Sci. 1999, 112 Pt 1, 2657–2666. [Google Scholar] [CrossRef]
- Lesuffleur, T.; Barbat, A.; Dussaulx, E.; Zweibaum, A. Growth adaptation to methotrexate of HT-29 human colon carcinoma cells is associated with their ability to differentiate into columnar absorptive and mucus-secreting cells. Cancer Res. 1990, 50, 6334–6343. [Google Scholar]
- Lesuffleur, T.; Porchet, N.; Aubert, J.P.; Swallow, D.; Gum, J.R.; Kim, Y.S.; Real, F.X.; Zweibaum, A. Differential expression of the human mucin genes MUC1 to MUC5 in relation to growth and differentiation of different mucus-secreting HT-29 cell subpopulations. J. Cell Sci. 1993, 106 Pt 3, 771–783. [Google Scholar] [CrossRef]
- Huet, C.; Sahuquillo-Merino, C.; Coudrier, E.; Louvard, D. Absorptive and mucus-secreting subclones isolated from a multipotent intestinal cell line (HT-29) provide new models for cell polarity and terminal differentiation. J. Cell Biol. 1987, 105, 345–357. [Google Scholar] [CrossRef]
- Roodsant, T.; Roodsant, T.; Navis, M.; Aknouch, I.; Renes, I.B. A Human 2D Primary Organoid-Derived Epithelial Monolayer Model to Study Host-Pathogen Interaction in the Small Intestine. Front. Cell. Infect. Microbiol. 2020, 1, 272. [Google Scholar] [CrossRef]
- Bartfeld, S.; Clevers, H. Organoids as Model for Infectious Diseases: Culture of Human and Murine Stomach Organoids and Microinjection of Helicobacter Pylori. J. Vis. Exp. 2015, 53359. [Google Scholar] [CrossRef] [PubMed]
- Xavier, M.; García-Hevia, L.; Amado, I.R.; Pastrana, L.; Gonçalves, C. In Vitro Intestinal Uptake and Permeability of Fluorescently-Labelled Hyaluronic Acid Nanogels. Int. J. Nanomed. 2019, 14, 9077–9088. [Google Scholar] [CrossRef]
- Johansson, M.E.V.; Hansson, G.C. Preservation of Mucus in Histological Sections, Immunostaining of Mucins in Fixed Tissue, and Localization of Bacteria with FISH. Methods Mol. Biol. 2012, 842, 229–235. [Google Scholar]
- Fernandez, C.; Mascolo, D.; Monaghan, S.J.; Baily, J.L.; Chalmers, L.; Paladini, G.; Adams, A.; Bron, J.E.; Fridman, S. Methacarn preserves mucus integrity and improves visualization of amoebae in gills of Atlantic salmon (Salmo salar L.). J. Fish Dis. 2019, 42, 883–894. [Google Scholar] [CrossRef]
- The quest for quantitative microscopy. Nat. Methods 2012, 9, 627. [CrossRef] [PubMed]
- Gustafson, A.W.; Gustafson, E.Y.; Douglas, W.H.J. Staining of interphase nuclei and mitotic figures in cultured cells with Alcian blue 8GX. In Vitro 1982, 18, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Chantret, I.; Barbat, A.; Dussaulx, E.; Brattain, M.G.; Zweibaum, A. Epithelial Polarity, Villin Expression, and Enterocytic Differentiation of Cultured Human Colon Carcinoma Cells: A Survey of Twenty Cell Lines. Cancer Res. 1988, 48, 1936–1942. [Google Scholar] [PubMed]
- Patnaik, S.; George, S.P.; Pham, E.; Roy, S.; Singh, K.; Mariadason, J.M.; Khurana, S. By moonlighting in the nucleus, Villin regulates epithelial plasticity. Mol. Biol. Cell 2016, 27, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Gouyer, V.; Gottrand, F.; Desseyn, J.-L. The Extraordinarily Complex but Highly Structured Organization of Intestinal Mucus-Gel Unveiled in Multicolor Images. PLoS ONE 2011, 6, e18761. [Google Scholar] [CrossRef]
- Jacobs, L.R.R.; Huber, P.W.W. Regional distribution and alterations of lectin binding to colorectal mucin in mucosal biopsies from controls and subjects with inflammatory bowel diseases. J. Clin. Investig. 1985, 75, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Clark, M.A.; Jepson, M.A.; Simmons, N.L.; Booth, T.A.; Hirst, B.H. Differential expression of lectin-binding sites defines mouse intestinal M-cells. J. Histochem. Cytochem. 1993, 41, 1679–1687. [Google Scholar] [CrossRef] [PubMed]
- Jepson, M.A.; Mason, C.M.; Clark, M.A.; Simmons, N.L.; Hirst, B.H. Variations in Lectin Binding Properties of Intestinal M Cells. J. Drug Target. 1995, 3, 75–77. [Google Scholar] [CrossRef] [PubMed]
- Macedonia, M.C.; Drewes, J.L.; Markham, N.O.; Simmons, A.J.; Roland, J.T.; Vega, P.N.; Scurrah, C.R.; Coffey, R.J.; Shrubsole, M.J.; Sears, C.L.; et al. Clinically adaptable polymer enables simultaneous spatial analysis of colonic tissues and biofilms. NPJ Biofilms Microbiomes 2020, 6, 33. [Google Scholar] [CrossRef]
- Garber, J.M.; Hennet, T.; Szymanski, C.M. Significance of fucose in intestinal health and disease. Mol. Microbiol. 2021, 115, 1086–1093. [Google Scholar] [CrossRef] [PubMed]
- Gebhard, A.; Gebert, A. Brush Cells of the Mouse Intestine Possess a Specialized Glycocalyx as Revealed by Quantitative Lectin Histochemistry: Further Evidence for a Sensory Function. J. Histochem. Cytochem. 1999, 47, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.E.V.; Holmén Larsson, J.M.; Hansson, G.C. The two mucus layers of colon are organized by the MUC2 mucin, whereas the outer layer is a legislator of host-microbial interactions. Proc. Natl. Acad. Sci. USA 2011, 108, 4659–4665. [Google Scholar] [CrossRef] [PubMed]
- Lesuffleur, T.; Barbat, A.; Zweibaum, A.; Kim, Y.S. Expression and Characterization of Mucins Associated with the Resistance to Methotrexate of Human Colonic Adenocarcinoma Cell Line HT29. Cancer Res. 1992, 52, 4655–4662. [Google Scholar]
- Sato, T.; van Es, J.H.; Snippert, H.J.; Stange, D.E.; Vries, R.G.; van den Born, M.; Barker, N.; Shroyer, N.F.; van de Wetering, M.; Clevers, H. Paneth cells constitute the niche for Lgr5 stem cells in intestinal crypts. Nature 2011, 469, 415–418. [Google Scholar] [CrossRef]
- Emde, B.; Heinen, A.; Gödecke, A.; Bottermann, K. Wheat germ agglutinin staining as a suitable method for detection and quantification of fibrosis in cardiac tissue after myocardial infarction. Eur. J. Histochem. 2014, 58, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Ude, V.C.; Brown, D.M.; Stone, V.; Johnston, H.J. Using 3D gastrointestinal tract in vitro models with microfold cells and mucus secreting ability to assess the hazard of copper oxide nanomaterials. J. Nanobiotechnol. 2019, 17, 70. [Google Scholar] [CrossRef]
- Olivier, J. Drug transport to brain with targeted nanoparticles. NeuroRX 2005, 2, 108–119. [Google Scholar] [CrossRef]
- Lueschow, S.R.; McElroy, S.J. The Paneth Cell: The Curator and Defender of the Immature Small Intestine. Front. Immunol. 2020, 11, 587. [Google Scholar] [CrossRef]
- Hanisch, F.G.; Baldus, S.E. The Thomsen-Friedenreich (TF) antigen: A critical review on the structural, biosynthetic and histochemical aspects of a pancarcinoma-associated antigen. Histol. Histopathol. 1997, 12, 263–281. [Google Scholar] [PubMed]
- Rhodes, J.M.; Campbell, B.J.; Yu, L.-G. Lectin–epithelial interactions in the human colon. Biochem. Soc. Trans. 2008, 36, 1482–1486. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Vijayan, M.; Surolia, A. Imparting exquisite specificity to peanut agglutinin for the tumor- associated Thomsen-Friedenreich antigen by redesign of its combining site. J. Biol. Chem. 1996, 271, 21209–21213. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Jenkins, G.J.S.; Asadi, R.; Doak, S.H. Potential toxicity of superparamagnetic iron oxide nanoparticles (SPION). Nano Rev. 2010, 1, 5358. [Google Scholar] [CrossRef] [PubMed]
- Velcich, A.; Palumbo, L.; Jarry, A.; Laboisse, C.; Racevskis, J.; Augenlicht, L. Patterns of expression of lineage-specific markers during the in vitro-induced differentiation of HT29 colon carcinoma cells. Cell Growth Differ. 1995, 6, 749–757. [Google Scholar]
- Jang, Y.; Jin, X.; Shankar, P.; Lee, J.H.; Jo, K.; Lim, K. Molecular-Level Interactions between Engineered Materials and Cells. Int. J. Mol. Sci. 2019, 20, 4142. [Google Scholar] [CrossRef]
- Mydin, R.B.S.M.N.; Sreekantan, S.; Hazan, R.; Qazem, E.Q.; Wajidi, M.F.F. Mechanosensitivity response of epithelial HT29 cells on titanium dioxide nanotube array surface via CK8 protein expression. Int. J. Nano Biomater. 2019, 8, 118. [Google Scholar] [CrossRef]
- Available online: www.phe-culturecollections.org.uk/media/55154/12040401-ht29-mtx-e12-images.pdf (accessed on 20 November 2021).
- Dituri, F.; Centonze, M.; Berenschot, E.J.W.; Tas, N.R.; Susarrey-Arce, A.; Krol, S. Complex Tumor Spheroid Formation and One-Step Cancer-Associated Fibroblasts Purification from Hepatocellular Carcinoma Tissue Promoted by Inorganic Surface Topography. Nanomaterials 2021, 11, 3233. [Google Scholar] [CrossRef]
- Goodwin, T.J.; Jessup, J.M.; Wolf, D.A. Morphologic differentiation of colon carcinoma cell lines HT-29 and HT-29KM in rotating-wall vessels. In Vitro Cell. Dev. Biol. 1992, 28A, 47–60. [Google Scholar] [CrossRef]
- Singh, R.; Subramanian, S.; Rhodes, J.M.; Campbell, B.J. Peanut lectin stimulates proliferation of colon cancer cells by interaction with glycosylated CD44v6 isoforms and consequential activation of c-Met and MAPK: Functional implications for disease-associated glycosylation changes. Glycobiology 2006, 16, 594–601. [Google Scholar] [CrossRef]
- Yu, S.; Balasubramanian, I.; Laubitz, D.; Tong, K.; Bandyopadhyay, S.; Lin, X.; Flores, J.; Singh, R.; Liu, Y.; Macazana, C.; et al. Paneth Cell-Derived Lysozyme Defines the Composition of Mucolytic Microbiota and the Inflammatory Tone of the Intestine. Immunity 2020, 53, 398–416.e8. [Google Scholar] [CrossRef]
- Wehkamp, J.; Stange, E.F. An Update Review on the Paneth Cell as Key to Ileal Crohn’s Disease. Front. Immunol. 2020, 11, 646. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fixative | 1st Step | 2nd Step | 3rd Step | 4th Step |
|---|---|---|---|---|
| PFA | Fixation | TRITC-Phalloidin | FITC-UEA | DAPI |
| Propidium iodide | Fixation | FITC-UEA | DAPI | |
| Fixation | Alcian blue | |||
| Fixation | Antibody (MUC2; lysozyme; villin) | |||
| Methacarn | Trypan blue | Imaging | Fixation | Alcian blue |
| Fixation | FITC-WGA/-PNA/-UEA | Imaging | Alcian blue | |
| Fixation | FITC-PNA | DAPI | ||
| Fixation | FITC-lectin | DAPI | Antibody (MUC2/lysozyme/villin) |
| Cell Culture (Days) | MUC2 % | Villin % | Lysozyme % |
|---|---|---|---|
| HT29 on 24-well plate * | 5.84 ± 3.50 | 4.67 ± 0.21 | 1.12 ± 0.69 |
| HT29-MTX on 24-well plate * | 8.67 ± 4.76 | 4.04 ± 2.78 | 0.18 ± 0.13 |
| HT29 on G0 (4 days) | 0 | 0.35 ± 0.47 | 0.63 ± 1.10 |
| HT29 on G0 (8 days) | 1.19 ± 1.45 | 0.38 ± 0.34 | 11.17 ± 7.65 |
| HT29 on G0 (11 days) | 2.14 ± 1.40 | 6.26 ± 2.00 | 12.34 ± 9.25 |
| HT29 on G1 (4 days) | 0.01 ± 0.02 | 0.14 ± 0.09 | 0 |
| HT29 on G1 (8 days) | 2.42 ± 1.43 | 0.27 ± 0.32 | 6.99 ± 4.60 |
| HT29 on G1 (11 days) | 0.57 ± 0.67 | 1.92 ± 1.58 | 10.97 ± 2.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Centonze, M.; Berenschot, E.J.W.; Serrati, S.; Susarrey-Arce, A.; Krol, S. The Fast Track for Intestinal Tumor Cell Differentiation and In Vitro Intestinal Models by Inorganic Topographic Surfaces. Pharmaceutics 2022, 14, 218. https://doi.org/10.3390/pharmaceutics14010218
Centonze M, Berenschot EJW, Serrati S, Susarrey-Arce A, Krol S. The Fast Track for Intestinal Tumor Cell Differentiation and In Vitro Intestinal Models by Inorganic Topographic Surfaces. Pharmaceutics. 2022; 14(1):218. https://doi.org/10.3390/pharmaceutics14010218
Chicago/Turabian StyleCentonze, Matteo, Erwin J. W. Berenschot, Simona Serrati, Arturo Susarrey-Arce, and Silke Krol. 2022. "The Fast Track for Intestinal Tumor Cell Differentiation and In Vitro Intestinal Models by Inorganic Topographic Surfaces" Pharmaceutics 14, no. 1: 218. https://doi.org/10.3390/pharmaceutics14010218