
Oral Immunogenicity of Enterotoxigenic Escherichia coli Outer Membrane Vesicles Encapsulated into Zein Nanoparticles Coated with a Gantrez® AN–Mannosamine Polymer Conjugate

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of the OMVs
2.2. Characterization of OMVs
2.2.1. Proteomic Analysis
2.2.2. Immunoblotting
2.3. Preparation of OMV-Loaded Nanoparticles
2.4. Characterization of Nanoparticles
2.5. OMV Quantification
2.6. Biodistribution Studies
2.7. Mice Immunization and Specific Antibody Response
2.8. Statistical Analysis
3. Results
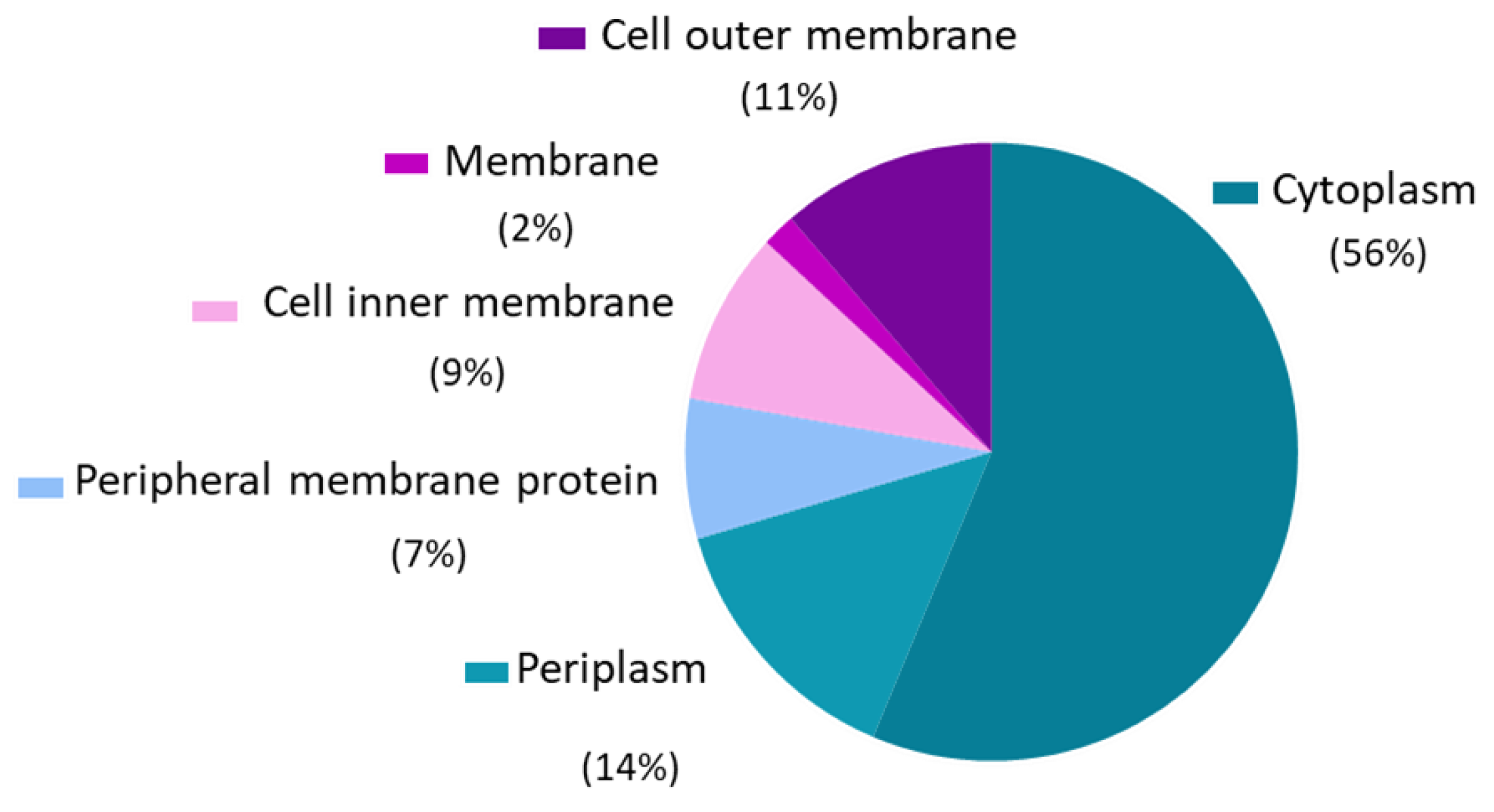
3.1. Outer Membrane Vesicles Proteomic Analysis
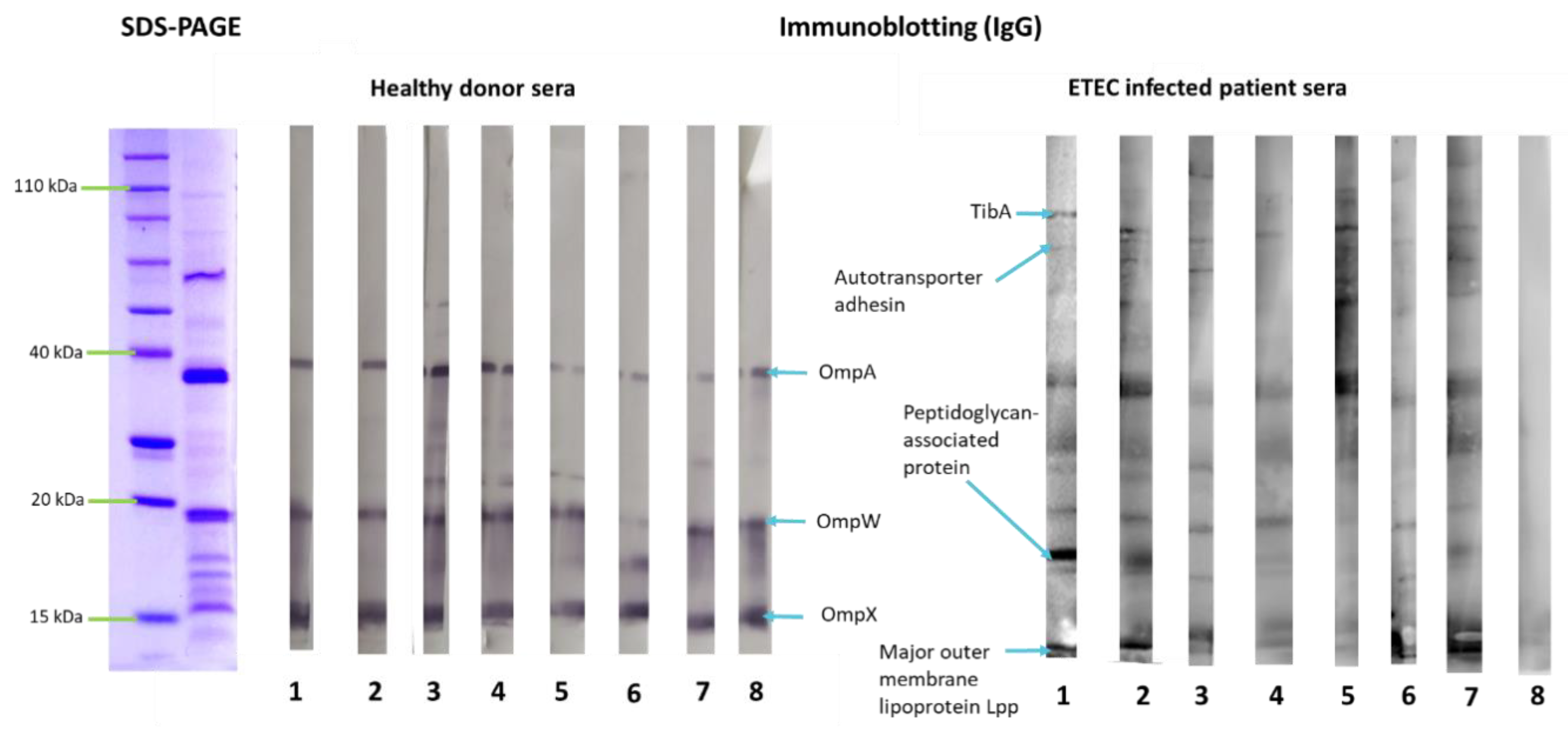
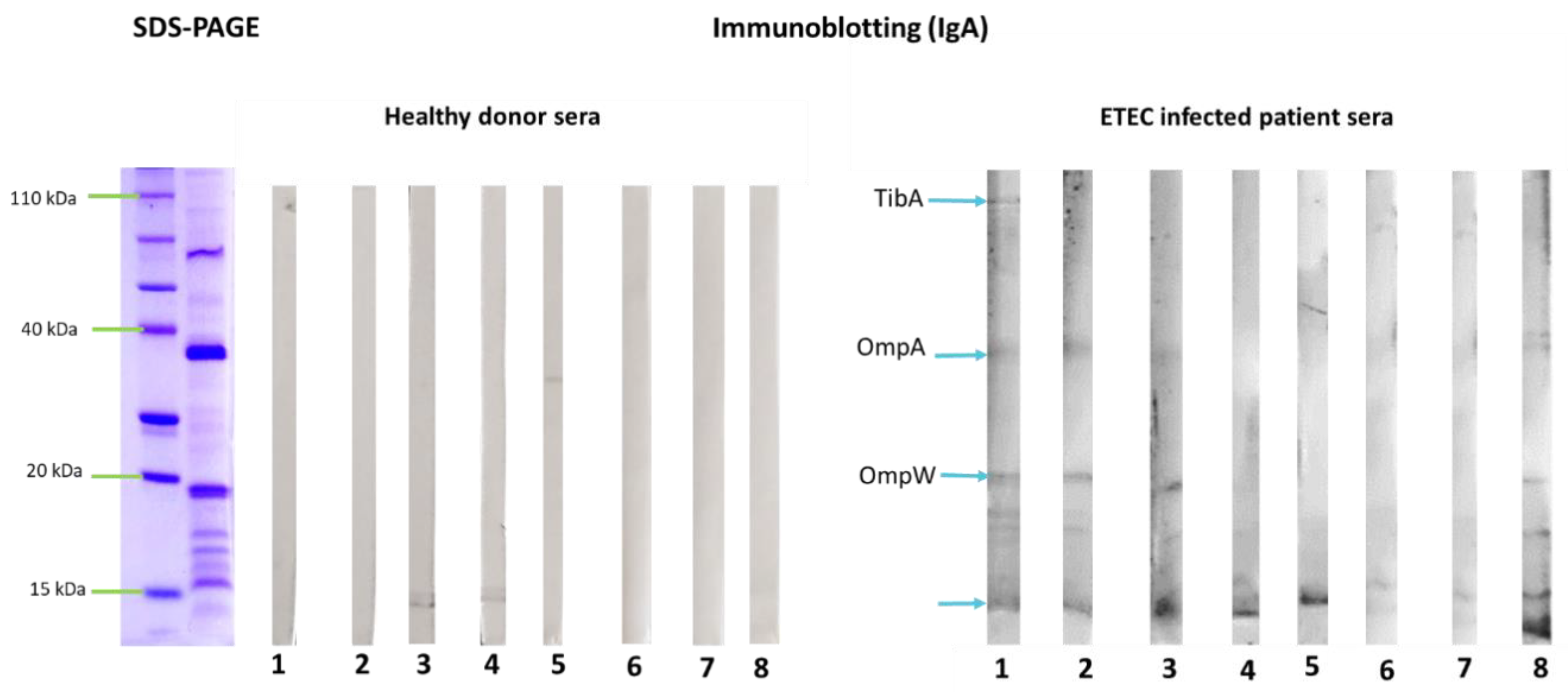
3.2. Outer Membrane Vesicles Antigenicity
3.3. Characterization of OMVs-Containing Nanoparticles
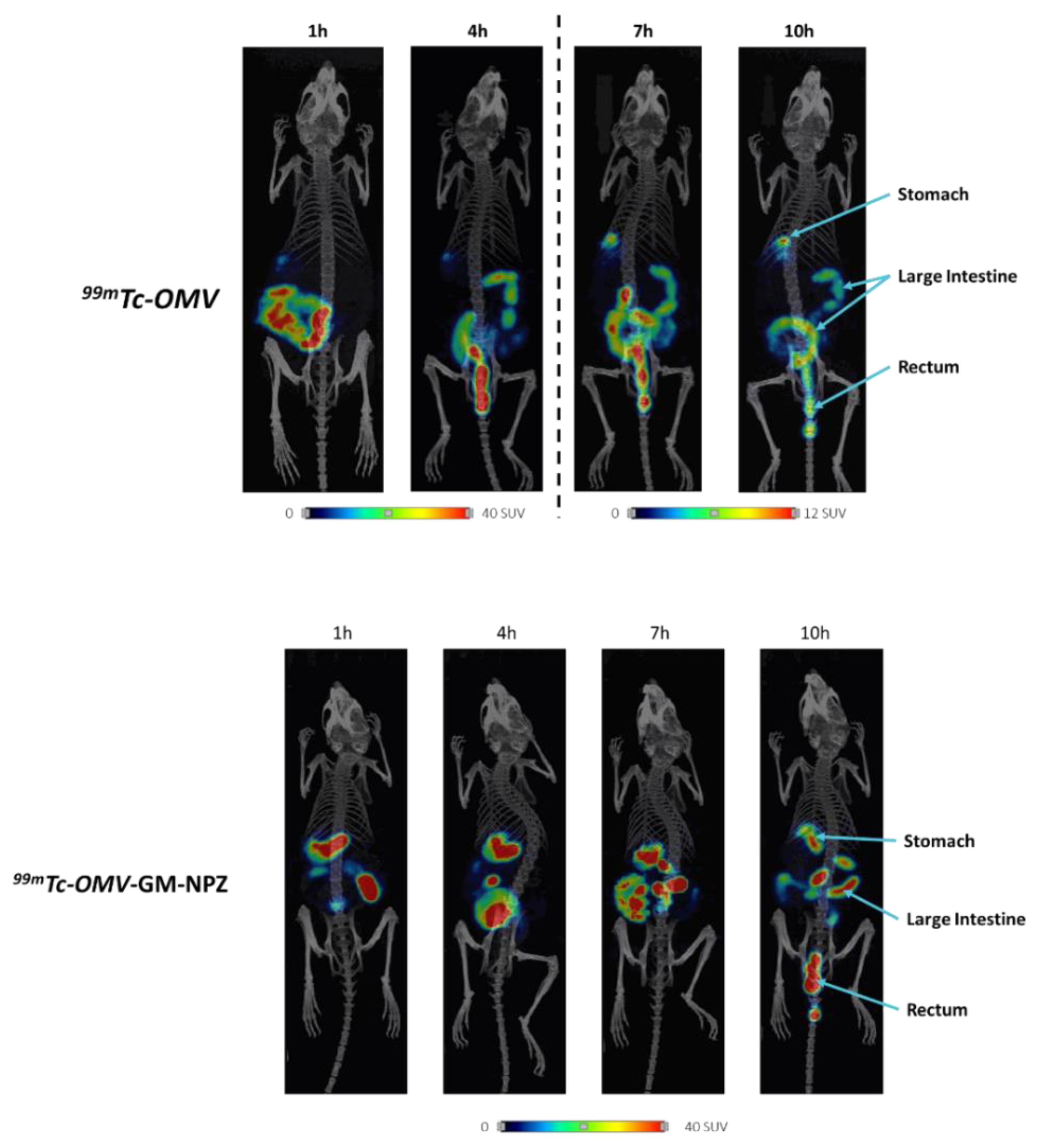
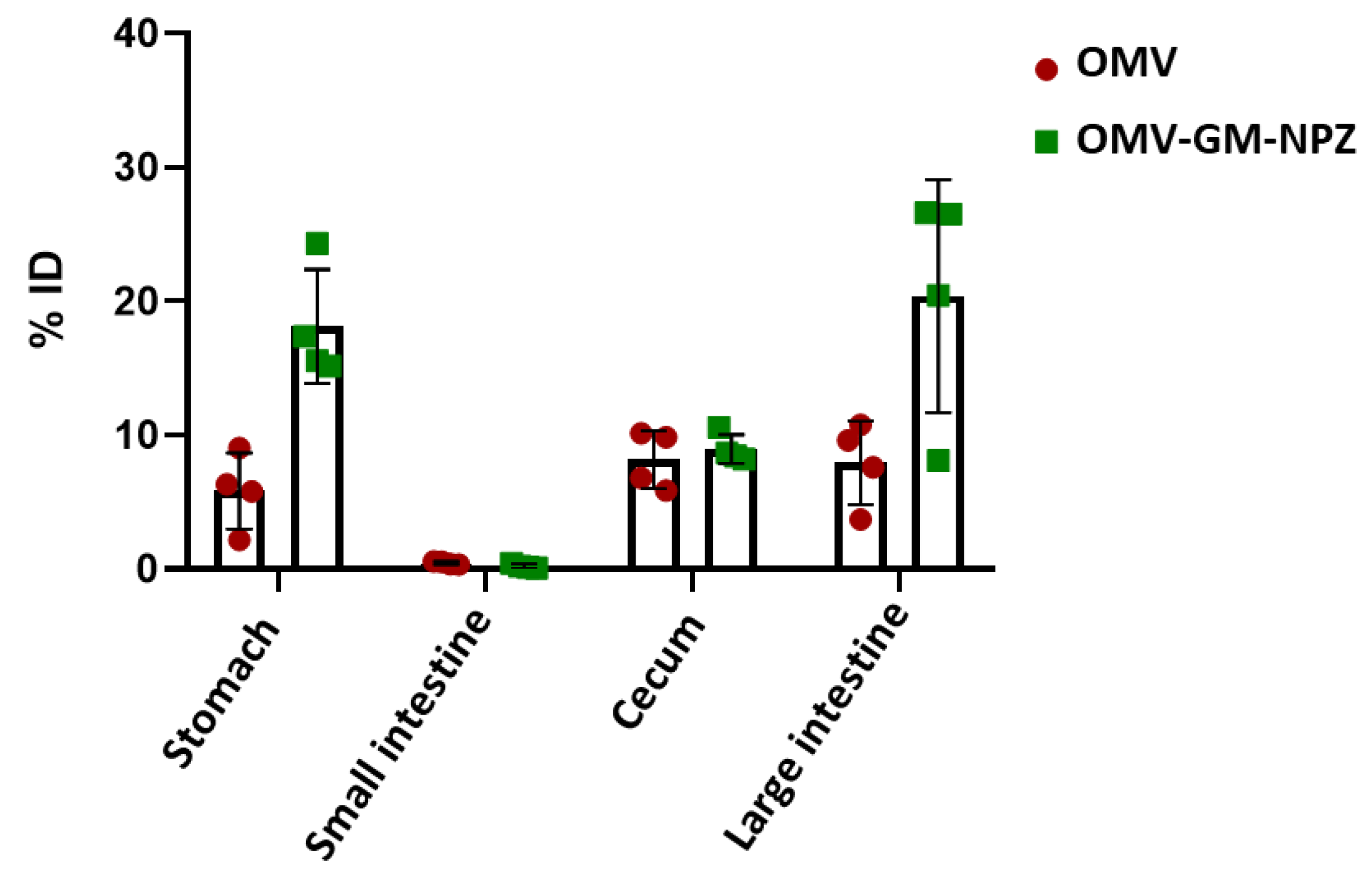
3.4. Biodistribution of OMV-GM-NPZ Compared to Free OMV
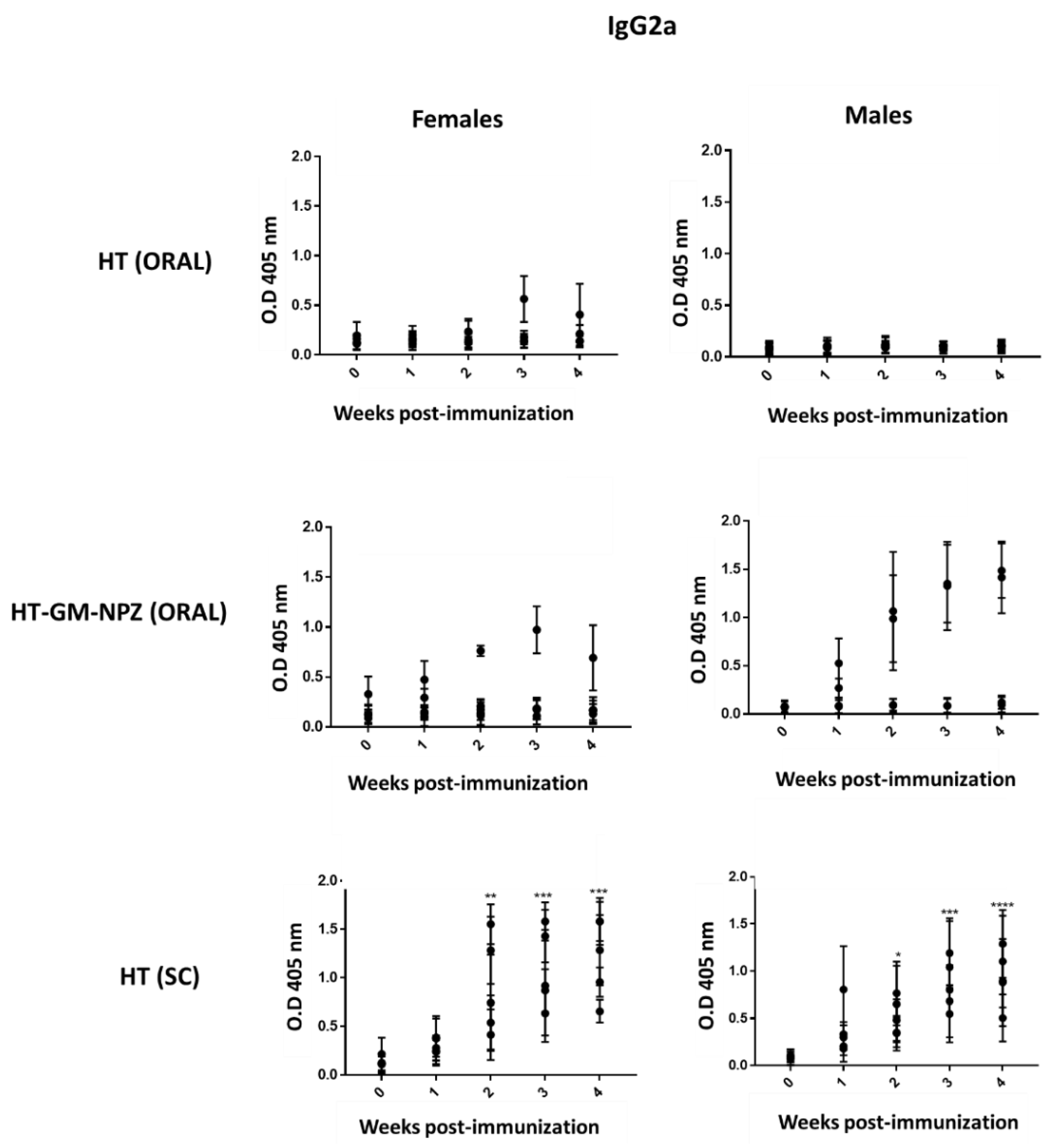
3.5. Evaluation of the Immunogenicity of OMV-GM-NPZ
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef]
- Fleckenstein, J.M.; Hardwidge, P.R.; Munson, G.P.; Rasko, D.A.; Sommerfelt, H.; Steinsland, H. Molecular mechanisms of enterotoxigenic Escherichia coli infection. Microbes Infect. 2010, 12, 89–98. [Google Scholar] [CrossRef]
- Khalil, I.A.; Troeger, C.; Blacker, B.F.; Rao, P.C.; Brown, A.; Atherly, D.E.; Brewer, T.G.; Engmann, C.M.; Houpt, E.R.; Kang, G.; et al. Morbidity and mortality due to shigella and Enterotoxigenic Escherichia coli diarrhoea: The Global Burden of Disease Study 1990–2016. Lancet Infect. Dis. 2018, 18, 1229–1240. [Google Scholar] [CrossRef] [Green Version]
- Sanders, J.W.; Riddle, M.S.; Taylor, D.N.; DuPont, H.L. Epidemiology of Travelers’ Diarrhea. In Travel Medicine; Elsevier Ltd.: Amsterdam, The Netherlands, 2008; pp. 187–198. [Google Scholar]
- Svennerholm, A.-M. From cholera to Enterotoxigenic Escherichia coli (ETEC) vaccine development. Indian J. Med. Res. 2011, 133, 188–196. [Google Scholar]
- Fleckenstein, J.M.; Kuhlmann, F.M. Enterotoxigenic Escherichia coli Infections. Curr. Infect. Dis. Rep. 2019, 21, 9. [Google Scholar] [CrossRef]
- Fleckenstein, J.M. Confronting Challenges to Enterotoxigenic Escherichia coli Vaccine Development. Front. Trop. Dis. 2021, 2, 9907. [Google Scholar] [CrossRef]
- Li, M.; Zhou, H.; Yang, C.; Wu, Y.; Zhou, X.; Liu, H.; Wang, Y. Bacterial outer membrane vesicles as a platform for biomedical applications: An update. J. Control. Release 2020, 323, 253–268. [Google Scholar] [CrossRef]
- Pollard, A.J.; Bijker, E.M. A guide to vaccinology: From basic principles to new developments. Nat. Rev. Immunol. 2021, 21, 83–100. [Google Scholar] [CrossRef]
- Bai, X.; Findlow, J.; Borrow, R. Recombinant protein meningococcal serogroup B vaccine combined with outer membrane vesicles. Expert Opin. Biol. Ther. 2011, 11, 969–985. [Google Scholar] [CrossRef]
- Gorringe, A.R.; Pajón, R. Bexsero: A multicomponent vaccine for prevention of meningococcal disease. Hum. Vaccin. Immunother. 2012, 8, 174–183. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Jin, L.; Chen, T. The Effects of Secretory IgA in the Mucosal Immune System. Biomed Res. Int. 2020, 2020, 2032057. [Google Scholar] [CrossRef]
- Vela Ramirez, J.E.; Sharpe, L.A.; Peppas, N.A. Current state and challenges in developing oral vaccines. Adv. Drug Deliv. Rev. 2017, 114, 116–131. [Google Scholar] [CrossRef]
- Forssell, H. Gastric mucosal defence mechanisms: A brief review. Scand. J. Gastroenterol. Suppl. 1988, 155, 23–28. [Google Scholar] [CrossRef]
- Jazayeri, S.D.; Lim, H.X.; Shameli, K.; Yeap, S.K.; Poh, C.L. Nano and Microparticles as Potential Oral Vaccine Carriers and Adjuvants Against Infectious Diseases. Front. Pharmacol. 2021, 12, 2286. [Google Scholar] [CrossRef]
- Duan, Q.; Xia, P.; Nandre, R.; Zhang, W.; Zhu, G. Review of Newly Identified Functions Associated with the Heat-Labile Toxin of Enterotoxigenic Escherichia coli. Front. Cell. Infect. Microbiol. 2019, 9, 292. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Iqbal, J.; Gómez-Duarte, O.G. Murine immunization with CS21 pili or LngA major subunit of enterotoxigenic Escherichia coli (ETEC) elicits systemic and mucosal immune responses and inhibits ETEC gut colonization. Vet. Microbiol. 2017, 202, 90–100. [Google Scholar] [CrossRef] [Green Version]
- Luo, Q.; Qadri, F.; Kansal, R.; Rasko, D.A.; Sheikh, A.; Fleckenstein, J.M. Conservation and Immunogenicity of Novel Antigens in Diverse Isolates of Enterotoxigenic Escherichia coli. PLoS Negl. Trop. Dis. 2015, 9, e0003446. [Google Scholar] [CrossRef] [Green Version]
- Rao, M.R.; Wierzba, T.F.; Savarino, S.J.; Abu-Elyazeed, R.; El-Ghoreb, N.; Hall, E.R.; Naficy, A.; Abdel-Messih, I.; Frenck, R.W., Jr.; Svennerholm, A.; et al. Serologic Correlates of Protection against Enterotoxigenic Escherichia coli Diarrhea. J. Infect. Dis. 2005, 191, 562–570. [Google Scholar] [CrossRef]
- Gaastra, W.; Svennrholm, A.-M. Colonization factors of human enterotoxigenic Escherichia coli (ETEC). Trends Microbiol. 1996, 4, 444–452. [Google Scholar] [CrossRef]
- Irache, J.M.; González-Navarro, C.J. Zein nanoparticles as vehicles for oral delivery purposes. Nanomedicine 2017, 12, 1209–1211. [Google Scholar] [CrossRef]
- Berzosa, M.; Pastor, Y.; Irache, J.M.; Gamazo, C. Experimental vaccination with nanoparticles containing Escherichia coli virulence factors. In Applications of Nanobiotechnology for Neglected Tropical Diseases; Elsevier Ltd.: Amsterdam, The Netherlands, 2021; pp. 3–27. [Google Scholar]
- Inchaurraga, L.; Martínez-López, A.L.; Abdulkarim, M.; Gumbleton, M.; Quincoces, G.; Peñuelas, I.; Martin-Arbella, N.; Irache, J.M. Modulation of the fate of zein nanoparticles by their coating with a Gantrez® AN-thiamine polymer conjugate. Int. J. Pharm. X 2019, 1, 100006. [Google Scholar] [CrossRef]
- Tamayo, I.; Irache, J.M.; Mansilla, C.; Ochoa-Repáraz, J.; Lasarte, J.J.; Gamazo, C. Poly(Anhydride) Nanoparticles Act as Active Th1 Adjuvants through Toll-Like Receptor Exploitation. Clin. Vaccine Immunol. 2010, 17, 1356–1362. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Shi, G.; Zhang, J.; Song, H.; Niu, J.; Shi, S.; Huang, P.; Wang, Y.; Wang, W.; Li, C.; et al. Targeted antigen delivery to dendritic cell via functionalized alginate nanoparticles for cancer immunotherapy. J. Control. Release 2017, 256, 170–181. [Google Scholar] [CrossRef]
- Lachén-Montes, M.; González-Morales, A.; Zelaya, M.V.; Pérez-Valderrama, E.; Ausín, K.; Ferrer, I.; Fernández-Irigoyen, J.; Santamaría, E. Olfactory bulb neuroproteomics reveals a chronological perturbation of survival routes and a disruption of prohibitin complex during Alzheimer’s disease progression. Sci. Rep. 2017, 7, 9115. [Google Scholar] [CrossRef]
- Lachén-Montes, M.; González-Morales, A.; de Morentin, X.M.; Pérez-Valderrama, E.; Ausín, K.; Zelaya, M.V.; Serna, A.; Aso, E.; Ferrer, I.; Fernández-Irigoyen, J.; et al. An early dysregulation of FAK and MEK/ERK signaling pathways precedes the β-amyloid deposition in the olfactory bulb of APP/PS1 mouse model of Alzheimer’s disease. J. Proteom. 2016, 148, 149–158. [Google Scholar] [CrossRef]
- Shilov, I.V.; Seymour, S.L.; Patel, A.A.; Loboda, A.; Tang, W.H.; Keating, S.P.; Hunter, C.L.; Nuwaysir, L.M.; Schaeffer, D.A. The Paragon Algorithm, a Next Generation Search Engine That Uses Sequence Temperature Values and Feature Probabilities to Identify Peptides from Tandem Mass Spectra. Mol. Cell. Proteom. 2007, 6, 1638–1655. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.H.; Shilov, I.V.; Seymour, S.L. Nonlinear Fitting Method for Determining Local False Discovery Rates from Decoy Database Searches. J. Proteome Res. 2008, 7, 3661–3667. [Google Scholar] [CrossRef]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [Green Version]
- Brotons-Canto, A.; Gamazo, C.; Martín-Arbella, N.; Abdulkarim, M.; Matías, J.; Gumbleton, M.; Irache, J.M. Evaluation of nanoparticles as oral vehicles for immunotherapy against experimental peanut allergy. Int. J. Biol. Macromol. 2018, 110, 328–335. [Google Scholar] [CrossRef]
- Matías, J.; Brotons, A.; Cenoz, S.; Pérez, I.; Abdulkarim, M.; Gumbleton, M.; Irache, J.M.; Gamazo, C. Oral Immunogenicity in Mice and Sows of Enterotoxigenic Escherichia coli Outer-Membrane Vesicles Incorporated into Zein-Based Nanoparticles. Vaccines 2019, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Sack, D.A. Progress and hurdles in the development of vaccines against Enterotoxigenic Escherichia coli in humans. Expert Rev. Vaccines 2012, 11, 677–694. [Google Scholar] [CrossRef]
- Leach, S.; Lundgren, A.; Carlin, N.; Löfstrand, M.; Svennerholm, A.-M. Cross-reactivity and avidity of antibody responses induced in humans by the oral inactivated multivalent Enterotoxigenic Escherichia coli (ETEC) vaccine ETVAX. Vaccine 2017, 35, 3966–3973. [Google Scholar] [CrossRef]
- Seo, H.; Garcia, C.; Ruan, X.; Duan, Q.; Sack, D.A.; Zhang, W. Preclinical Characterization of Immunogenicity and Efficacy against Diarrhea from MecVax, a Multivalent Enterotoxigenic E. coli Vaccine Candidate. Infect. Immun. 2021, 89. [Google Scholar] [CrossRef]
- Qadri, F.; Akhtar, M.; Bhuiyan, T.R.; Chowdhury, M.I.; Ahmed, T.; Rafique, T.A.; Khan, A.; Rahman, S.I.A.; Khanam, F.; Lundgren, A.; et al. Safety and immunogenicity of the oral, inactivated, Enterotoxigenic Escherichia coli vaccine ETVAX in Bangladeshi children and infants: A double-blind, randomised, placebo-controlled phase 1/2 trial. Lancet Infect. Dis. 2020, 20, 208–219. [Google Scholar] [CrossRef] [Green Version]
- Khalil, I.; Walker, R.; Porter, C.K.; Muhib, F.; Chilengi, R.; Cravioto, A.; Guerrant, R.; Svennerholm, A.M.; Qadri, F.; Baqar, S.; et al. Enterotoxigenic Escherichia coli (ETEC) vaccines: Priority activities to enable product development, licensure, and global access. Vaccine 2021, 39, 4266–4277. [Google Scholar] [CrossRef]
- van der Pol, L.; Stork, M.; van der Ley, P. Outer membrane vesicles as platform vaccine technology. Biotechnol. J. 2015, 10, 1689–1706. [Google Scholar] [CrossRef]
- Chen, L.; Valentine, J.L.; Huang, C.-J.; Endicott, C.E.; Moeller, T.D.; Rasmussen, J.A.; Fletcher, J.R.; Boll, J.M.; Rosenthal, J.A.; Dobruchowska, J.; et al. Outer membrane vesicles displaying engineered glycotopes elicit protective antibodies. Proc. Natl. Acad. Sci. USA 2016, 113, E3609–E3618. [Google Scholar] [CrossRef] [Green Version]
- Isidean, S.D.; Riddle, M.S.; Savarino, S.J.; Porter, C.K. A systematic review of ETEC epidemiology focusing on colonization factor and toxin expression. Vaccine 2011, 29, 6167–6178. [Google Scholar] [CrossRef]
- Vidal, R.M.; Muhsen, K.; Tennant, S.M.; Svennerholm, A.M.; Sow, S.O.; Sur, D.; Zaidi, A.K.M.; Faruque, A.S.G.; Saha, D.; Adegbola, R.; et al. Colonization factors among Enterotoxigenic Escherichia coli isolates from children with moderate-to-severe diarrhea and from matched controls in the Global Enteric Multicenter Study (GEMS). PLoS Negl. Trop. Dis. 2019, 13, e0007037. [Google Scholar] [CrossRef] [Green Version]
- Kuhlmann, F.M.; Martin, J.; Hazen, T.H.; Vickers, T.J.; Pashos, M.; Okhuysen, P.C.; Gómez-Duarte, O.G.; Cebelinski, E.; Boxrud, D.; Del Canto, F.; et al. Conservation and global distribution of noncanonical antigens in Enterotoxigenic Escherichia coli. PLoS Negl. Trop. Dis. 2019, 13, e0007825. [Google Scholar] [CrossRef] [Green Version]
- Qadri, F.; Das, S.K.; Faruque, A.S.G.; Fuchs, G.J.; Albert, M.J.; Sack, R.B.; Svennerholm, A.M. Prevalence of toxin types and colonization factors in Enterotoxigenic Escherichia coli isolated during a 2-year period from diarrheal patients in Bangladesh. J. Clin. Microbiol. 2000, 38, 27–31. [Google Scholar] [CrossRef]
- Zhang, S.; Wu, Q.; Zhang, J.; Lai, Z.; Zhu, X. Prevalence, genetic diversity, and antibiotic resistance of Enterotoxigenic Escherichia coli in retail ready-to-eat foods in China. Food Control 2016, 68, 236–243. [Google Scholar] [CrossRef]
- Guerra, J.A.; Romero-Herazo, Y.C.; Arzuza, O.; Gómez-Duarte, O.G. Phenotypic and genotypic characterization of Enterotoxigenic Escherichia coli clinical isolates from Northern Colombia, South America. BioMed Res. Int. 2014, 2014, 236260. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.A.; Roy, K.; Woo-Rasberry, V.; Hamilton, D.J.; Kansal, R.; Qadri, F.; Fleckenstein, J.M. Directed evaluation of Enterotoxigenic Escherichia coli autotransporter proteins as putative vaccine candidates. PLoS Negl. Trop. Dis. 2011, 5, e1428. [Google Scholar] [CrossRef] [Green Version]
- Rivas, Z.P.; Talbot, K.M.; Merselis, L.C.; McCormack, R.M.; Adkins, B.; Munson, G.P. CexE Is a Coat Protein and Virulence Factor of Diarrheagenic Pathogens. Front. Microbiol. 2020, 11, 626–638. [Google Scholar] [CrossRef]
- Chakraborty, S.; Randall, A.; Vickers, T.J.; Molina, D.; Harro, C.D.; DeNearing, B.; Brubaker, J.; Sack, D.A.; Bourgeois, A.L.; Felgner, P.L.; et al. Human Experimental Challenge with Enterotoxigenic Escherichia coli Elicits Immune Responses to Canonical and Novel Antigens Relevant to Vaccine Development. J. Infect. Dis. 2018, 218, 1436–1446. [Google Scholar] [CrossRef]
- De Smet, R.; Allais, L.; Cuvelier, C.A. Recent advances in oral vaccine development. Hum. Vaccin. Immunother. 2014, 10, 1309–1318. [Google Scholar] [CrossRef] [Green Version]
- Taddio, A.; Ipp, M.; Thivakaran, S.; Jamal, A.; Parikh, C.; Smart, S.; Sovran, J.; Stephens, D.; Katz, J. Survey of the prevalence of immunization non-compliance due to needle fears in children and adults. Vaccine 2012, 30, 4807–4812. [Google Scholar] [CrossRef] [Green Version]
- Davitt, C.J.H.; Lavelle, E.C. Delivery strategies to enhance oral vaccination against enteric infections. Adv. Drug Deliv. Rev. 2015, 91, 52–69. [Google Scholar] [CrossRef] [Green Version]
- Levine, M.M.; Dougan, G. Optimism over vaccines administered via mucosal surfaces. Lancet 1998, 351, 1375–1376. [Google Scholar] [CrossRef]
- Mudie, D.M.; Amidon, G.L.; Amidon, G.E. Physiological Parameters for Oral Delivery and in Vitro Testing. Mol. Pharm. 2010, 7, 1388–1405. [Google Scholar] [CrossRef] [Green Version]
- Wilson-Welder, J.H.; Torres, M.P.; Kipper, M.J.; Mallapragada, S.K.; Wannemuehler, M.J.; Narasimhan, B. Vaccine adjuvants: Current challenges and future approaches. J. Pharm. Sci. 2009, 98, 1278–1316. [Google Scholar] [CrossRef]
- Kunisawa, J.; Kurashima, Y.; Kiyono, H. Gut-associated lymphoid tissues for the development of oral vaccines. Adv. Drug Deliv. Rev. 2012, 64, 523–530. [Google Scholar] [CrossRef]
- McNeela, E.A.; Lavelle, E.C. Recent Advances in Microparticle and Nanoparticle Delivery Vehicles for Mucosal Vaccination. Curr. Top. Microbiol. Immunol. 2012, 354, 75–99. [Google Scholar] [CrossRef]
- Mann, J.F.; Acevedo, R.; del Campo, J.; Pérez, O.; Ferro, V.A. Delivery systems: A vaccine strategy for overcoming mucosal tolerance? Expert Rev. Vaccines 2009, 8, 103–112. [Google Scholar] [CrossRef]
- Camacho, A.I.; Da Costa Martins, R.; Tamayo, I.; de Souza, J.; Lasarte, J.J.; Mansilla, C.; Esparza, I.; Irache, J.M.; Gamazo, C. Poly(methyl vinyl ether-co-maleic anhydride) nanoparticles as innate immune system activators. Vaccine 2011. [Google Scholar] [CrossRef] [Green Version]
- Salman, H.H.; Gamazo, C.; Campanero, M.A.; Irache, J.M. Bioadhesive Mannosylated Nanoparticles for Oral Drug Delivery. J. Nanosci. Nanotechnol. 2006, 6, 3203–3209. [Google Scholar] [CrossRef]
- Matías, J.; Pastor, Y.; Irache, J.M.; Gamazo, C. Protective Passive Immunity in Escherichia coli ETEC-Challenged Neonatal Mice Conferred by Orally Immunized Dams with Nanoparticles Containing Homologous Outer Membrane Vesicles. Vaccines 2020, 8, 286. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ETEC (Clinical Isolate n°1) | |||
|---|---|---|---|
| UniProt Identifier | Gene (Strain) | Protein | iBAQ |
| D7GKK6 | lngA (ETEC1392/75_p557_00053) | Putative pilus biosynthesis protein | 6.50 × 108 |
| D7GKE0 | etpA (ETEC1392/75_p1018_132) | Putative heamagglutinin afimbrial adhesin | 1.91 × 108 |
| D7GK42 | eltB (ETEC1392/75_p1018_007) | Heat-labile enterotoxin B chain | 9.6 × 107 |
| E3PJ90 | yghJ (ETEC_3241) | Putative lipoprotein YghJ (Putative lipoprotein AcfD homolog) | 3.39 × 107 |
| D7GKA6 | cstA-H (ETEC1392/75_p1018_087) | Putative CS3 fimbrial subunit | 3.20 × 107 |
| Q9XD84 | tibA (ETEC_2141) | Adhesin/invasin TibA autotransporter | 6.72 × 106 |
| C8UFQ7 | yghJ (ECO111_3795) | Putative lipoprotein AcfD homolog | 5.49 × 106 |
| D7GK41 | eltA (ETEC1392/75_p1018_006) | Heat-labile enterotoxin A chain | 4.12 × 106 |
| E3PPC4 | cfaB (ETEC_p948_0400) | CFA/I fimbrial subunit B (CFA/I antigen) | 3.53 × 106 |
| B7UI41 | yghJ (E2348C_3253) | Predicted inner membrane lipoprotein | 3.50 × 105 |
| Q84GK0 | eatA (ETEC_p948_0020) | Serine protease EatA | 1.55 × 105 |
| ETEC (Clinical Isolate n°2) | |||
|---|---|---|---|
| UniProt Identifier | Gene (Strain) | Protein | iBAQ |
| Q9XD84 | tibA (ETEC_2141) | Adhesin/invasin TibA autotransporter | 1.84 × 108 |
| D7GK42 | eltB (ETEC1392/75_p1018_007) | Heat-labile enterotoxin B chain | 5.97 × 107 |
| E3PPC4 | cfaB (ETEC_p948_0400) | CFA/I fimbrial subunit B (CFA/I antigen) | 3.84 × 107 |
| E3PJ90 | yghJ (ETEC_3241) | Putative lipoprotein YghJ (Putative lipoprotein AcfD homolog) | 2.68 × 107 |
| E3PPC6 | cfaE (ETEC_p948_0420) | Cfa/I fimbrial subunit E | 1.89 × 107 |
| D7GKK6 | lngA (ETEC1392/75_p557_00053) | Putative pilus biosynthesis protein | 1.70 × 107 |
| D7GK41 | eltA (ETEC1392/75_p1018_006) | Heat-labile enterotoxin A chain | 1.49 × 107 |
| C8UFQ7 | yghJ (ECO111_3795) | Putative lipoprotein AcfD homolog | 1.06 × 107 |
| E3PPC3 | cfaA (ETEC_p948_0390) | CfA/I fimbrial subunit A | 5.31 × 106 |
| Q84GK0 | eatA (ETEC_p948_0020) | Serine protease EatA | 2.55 × 106 |
| D7GKE0 | etpA (ETEC1392/75_p1018_132) | EtpA adhesin | 2.52 × 106 |
| Formulation | Size (nm) | Polydispersity Index (PDI) | Zeta Potential (mV) | OMV Loading (μg/mg) | EE (%) |
|---|---|---|---|---|---|
| GM-NPZ | 245 ± 6.00 | 0.07 ± 0.01 | −51 ± 1.60 | N.D | N.D |
| HT-GM-NPZ | 211 ± 5.00 | 0.14 ± 0.04 | −48.71 ± 0.68 | 5.80 | ~70% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berzosa, M.; Nemeskalova, A.; Calvo, A.; Quincoces, G.; Collantes, M.; Pareja, F.; Gamazo, C.; Irache, J.M. Oral Immunogenicity of Enterotoxigenic Escherichia coli Outer Membrane Vesicles Encapsulated into Zein Nanoparticles Coated with a Gantrez® AN–Mannosamine Polymer Conjugate. Pharmaceutics 2022, 14, 123. https://doi.org/10.3390/pharmaceutics14010123
Berzosa M, Nemeskalova A, Calvo A, Quincoces G, Collantes M, Pareja F, Gamazo C, Irache JM. Oral Immunogenicity of Enterotoxigenic Escherichia coli Outer Membrane Vesicles Encapsulated into Zein Nanoparticles Coated with a Gantrez® AN–Mannosamine Polymer Conjugate. Pharmaceutics. 2022; 14(1):123. https://doi.org/10.3390/pharmaceutics14010123
Chicago/Turabian StyleBerzosa, Melibea, Alzbeta Nemeskalova, Alba Calvo, Gemma Quincoces, María Collantes, Felix Pareja, Carlos Gamazo, and Juan Manuel Irache. 2022. "Oral Immunogenicity of Enterotoxigenic Escherichia coli Outer Membrane Vesicles Encapsulated into Zein Nanoparticles Coated with a Gantrez® AN–Mannosamine Polymer Conjugate" Pharmaceutics 14, no. 1: 123. https://doi.org/10.3390/pharmaceutics14010123