
Engineering Therapeutic Strategies in Cancer Immunotherapy via Exogenous Delivery of Toll-like Receptor Agonists


Abstract
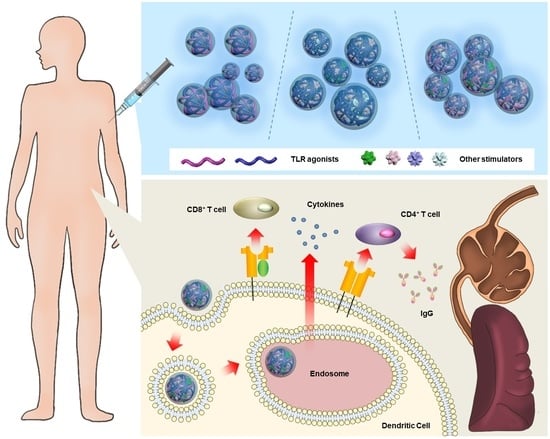
:
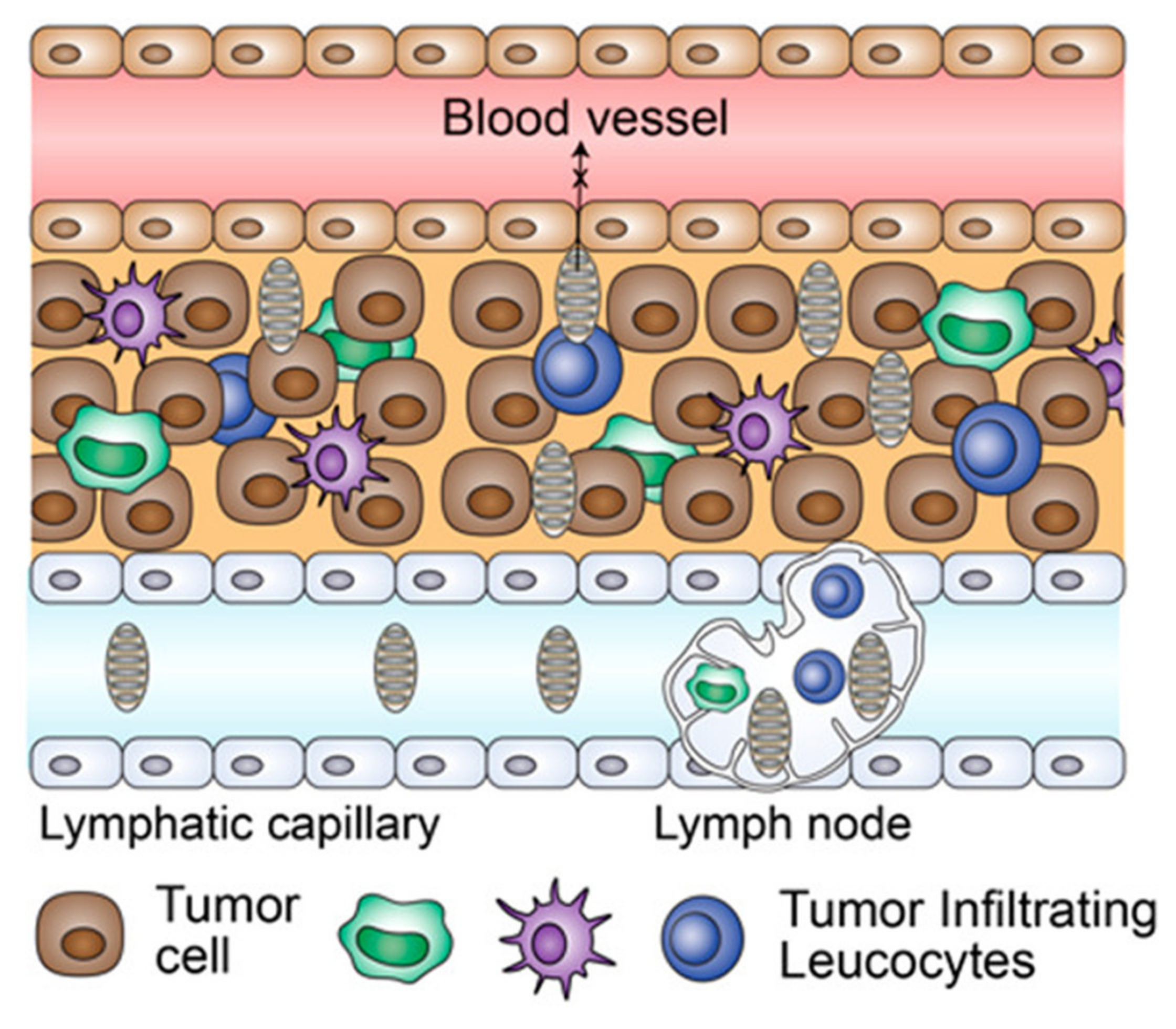
1. Introduction
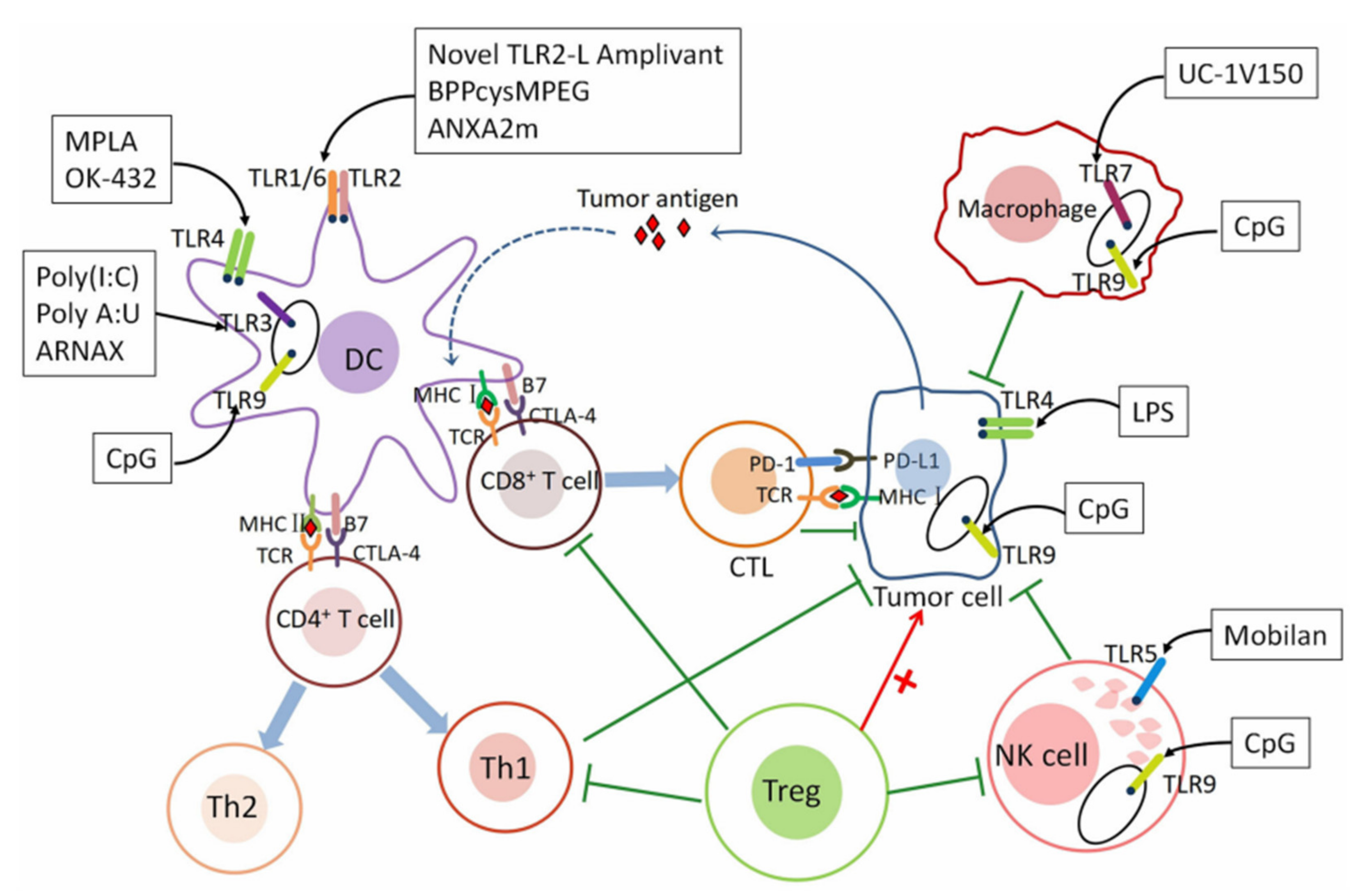
2. Promoting Immune Responses through TLR Agonist Delivery
2.1. Strategies for Immune Cell Stimulation via Exogenous TLR Agonist Delivery
2.2. The Role of TLR Agonists in Immune Check Point Pathways
2.3. TLR Agonists as Adjuvants for Cancer Vaccines
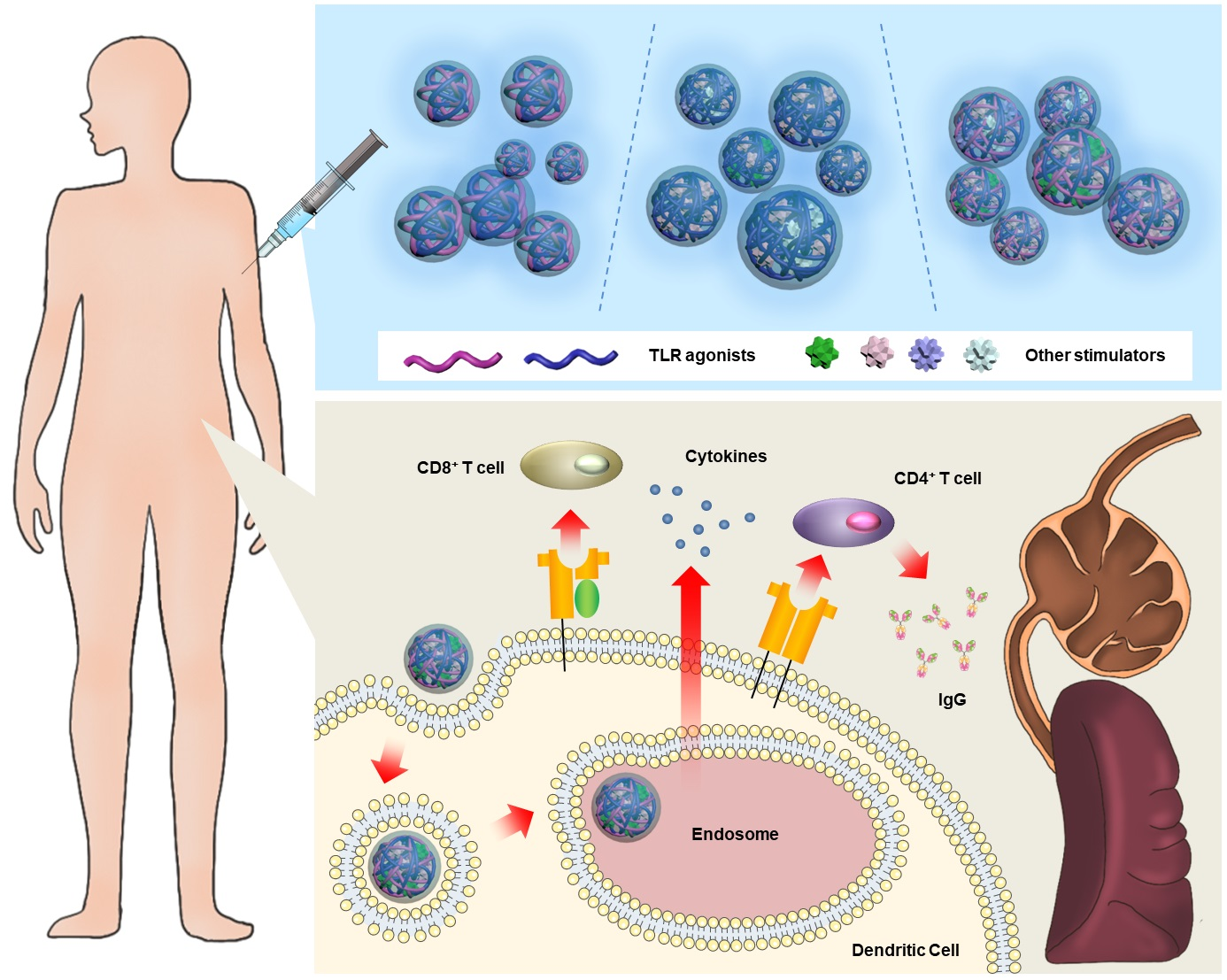
3. Combined Delivery of Multiple TLR Agonists for Enhanced Cancer Immunotherapeutic Efficacy
4. Combinative Delivery of TLR Agonists with Various Biochemical Co-Factors Inducing Immune System
4.1. Co-Delivery Using a Single Immune Substance
4.2. Synergistic Immunogenic Effects via Co-Delivery Using Diverse Co-Factors
4.3. Clinical Relevance of TLR Agonist Delivery for Cancer Treatment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Weber, J.S.; Mule, J.J. Cancer immunotherapy meets biomaterials. Nat. Biotechnol. 2015, 33, 44–45. [Google Scholar] [CrossRef]
- De Vries, J.; Figdor, C. IMMUNOTHERAPY Cancer vaccine triggers antiviral-type defences. Nature 2016, 534, 329–331. [Google Scholar] [CrossRef]
- Wang, R.F.; Wang, H.Y. Immune targets and neoantigens for cancer immunotherapy and precision medicine. Cell Res. 2017, 27, 11–37. [Google Scholar] [CrossRef] [Green Version]
- Mandal, A.; Boopathy, A.V.; Lam, L.K.W.; Moynihan, K.D.; Welch, M.E.; Bennett, N.R.; Turvey, M.E.; Thai, N.; Van, J.H.; Love, J.C.; et al. Cell and fluid sampling microneedle patches for monitoring skin-resident immunity. Sci. Transl. Med. 2018, 10, 467. [Google Scholar] [CrossRef] [PubMed]
- Dellacherie, M.O.; Seo, B.R.; Mooney, D.J. Macroscale biomaterials strategies for local immunomodulation. Nat. Rev. Mater. 2019, 4, 379–397. [Google Scholar] [CrossRef]
- Wang, C.; Ye, Y.; Hu, Q.; Bellotti, A.; Gu, Z. Tailoring Biomaterials for Cancer Immunotherapy: Emerging Trends and Future Outlook. Adv. Mater. 2017, 29, 1606036. [Google Scholar] [CrossRef]
- Schmidt, S.V.; Nino-Castro, A.C.; Schultze, J.L. Regulatory dendritic cells: There is more than just immune activation. Front. Immunol. 2012, 3, 274. [Google Scholar] [CrossRef] [Green Version]
- Wylie, B.; Macri, C.; Mintern, J.D.; Waithman, J. Dendritic Cells and Cancer: From Biology to Therapeutic Intervention. Cancers 2019, 11, 521. [Google Scholar] [CrossRef] [Green Version]
- Escors, D. Tumour immunogenicity, antigen presentation, and immunological barriers in cancer immunotherapy. New J. Sci. 2014, 2014, 1–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawarada, Y.; Ganss, R.; Garbi, N.; Sacher, T.; Arnold, B.; Hammerling, G.J. NK- and CD8(+) T cell-mediated eradication of established tumors by peritumoral injection of CpG-containing oligodeoxynucleotides. J. Immunol. 2001, 167, 5247–5253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fong, L.; Engleman, E.G. Dendritic cells in cancer immunotherapy. Annu. Rev. Immunol. 2000, 18, 245–273. [Google Scholar] [CrossRef]
- Segura, E.; Amigorena, S.; Thery, C. Mature dendritic cells secrete exosomes with strong ability to induce antigen-specific effector immune responses. Blood Cells Mol. Dis. 2005, 35, 89–93. [Google Scholar] [CrossRef]
- Lin, Y.H.; Chen, Z.R.; Lai, C.H.; Hsieh, C.H.; Feng, C.L. Active Targeted Nanoparticles for Oral Administration of Gastric Cancer Therapy. Biomacromolecules 2015, 16, 3021–3032. [Google Scholar] [CrossRef] [PubMed]
- Michiels, A.; Tuyaerts, S.; Bonehill, A.; Corthals, J.; Breckpot, K.; Heirman, C.; Van Meirvenne, S.; Dullaers, M.; Allard, S.; Brasseur, F.; et al. Electroporation of immature and mature dendritic cells: Implications for dendritic cell-based vaccines. Gene Ther. 2005, 12, 772–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutz, M.B.; Schuler, G. Immature, semi-mature and fully mature dendritic cells: Which signals induce tolerance or immunity? Trends Immunol. 2002, 23, 445–449. [Google Scholar] [CrossRef]
- Kanzler, H.; Barrat, F.J.; Hessel, E.M.; Coffman, R.L. Therapeutic targeting of innate immunity with Toll-like receptor agonists and antagonists. Nat. Med. 2007, 13, 552–559. [Google Scholar] [CrossRef]
- Adams, S. Toll-like receptor agonists in cancer therapy. Immunotherapy 2009, 1, 949–964. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Akira, S. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int. Immunol. 2009, 21, 317–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, G.Y.; Lee, J.W.; Kim, Y.S.; Lee, S.E.; Han, H.D.; Hong, K.J.; Kang, T.H.; Park, Y.M. Interactions between tumor-derived proteins and Toll-like receptors. Exp. Mol. Med. 2020, 52, 1926–1935. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [Green Version]
- Hou, B.; Reizis, B.; DeFranco, A.L. Toll-like receptors activate innate and adaptive immunity by using dendritic cell-intrinsic and -extrinsic mechanisms. Immunity 2008, 29, 272–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, C.; Cao, X. Regulation of Toll-like receptor signaling pathways in innate immune responses. Ann. N. Y. Acad. Sci. 2013, 1283, 67–74. [Google Scholar] [CrossRef]
- Barton, G.M.; Medzhitov, R. Control of adaptive immune responses by Toll-like receptors. Curr. Opin. Immunol. 2002, 14, 380–383. [Google Scholar] [CrossRef]
- Iwasaki, A.; Medzhitov, R. Toll-like receptor control of the adaptive immune responses. Nat. Immunol. 2004, 5, 987–995. [Google Scholar] [CrossRef]
- Van Herck, S.; De Geest, B.G. Nanomedicine-mediated alteration of the pharmacokinetic profile of small molecule cancer immunotherapeutics. Acta Pharmacol. Sin. 2020, 41, 881–894. [Google Scholar] [CrossRef]
- Lynn, G.M.; Laga, R.; Darrah, P.A.; Ishizuka, A.S.; Balaci, A.J.; Dulcey, A.E.; Pechar, M.; Pola, R.; Gerner, M.Y.; Yamamoto, A.; et al. In vivo characterization of the physicochemical properties of polymer-linked TLR agonists that enhance vaccine immunogenicity. Nat. Biotechnol. 2015, 33, 1201–1210. [Google Scholar] [CrossRef]
- Carson, W.E.; Dierksheide, J.E.; Jabbour, S.; Anghelina, M.; Bouchard, P.; Ku, G.; Yu, H.; Baumann, H.; Shah, M.H.; Cooper, M.A. Coadministration of interleukin-18 and interleukin-12 induces a fatal inflammatory response in mice: Critical role of natural killer cell interferon-γ production and STAT-mediated signal transduction. Blood 2000, 96, 1465–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, I.Y.; Yan, X.; Tohme, S.; Ahmed, A.; Cordon-Cardo, C.; Shantha Kumara, H.M.; Kim, S.K.; Whelan, R.L. CpG ODN, Toll like receptor (TLR)-9 agonist, inhibits metastatic colon adenocarcinoma in a murine hepatic tumor model. J. Surg. Res. 2012, 174, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; An, M.; Xi, J.; Liu, H. Targeting CpG Adjuvant to Lymph Node via Dextran Conjugate Enhances Antitumor Immunotherapy. Bioconjug. Chem. 2017, 28, 1993–2000. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, M.F.; Jennings, G.T. Vaccine delivery: A matter of size, geometry, kinetics and molecular patterns. Nat. Rev. Mater. 2010, 10, 787–796. [Google Scholar] [CrossRef]
- Komura, F.; Okuzumi, K.; Takahashi, Y.; Takakura, Y.; Nishikawa, M. Development of RNA/DNA Hydrogel Targeting Toll-Like Receptor 7/8 for Sustained RNA Release and Potent Immune Activation. Molecules 2020, 25, 728. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.M.; Pan, W.Y.; Wu, C.Y.; Yeh, C.Y.; Korupalli, C.; Luo, P.K.; Chou, C.J.; Chia, W.T.; Sung, H.W. Modulation of tumor microenvironment using a TLR-7/8 agonist-loaded nanoparticle system that exerts low-temperature hyperthermia and immunotherapy for in situ cancer vaccination. Biomaterials 2020, 230, 119629. [Google Scholar] [CrossRef] [PubMed]
- Nam, H.Y.; Kwon, S.M.; Chung, H.; Lee, S.Y.; Kwon, S.H.; Jeon, H.; Kim, Y.; Park, J.H.; Kim, J.; Her, S.; et al. Cellular uptake mechanism and intracellular fate of hydrophobically modified glycol chitosan nanoparticles. J. Control. Release 2009, 135, 259–267. [Google Scholar] [CrossRef]
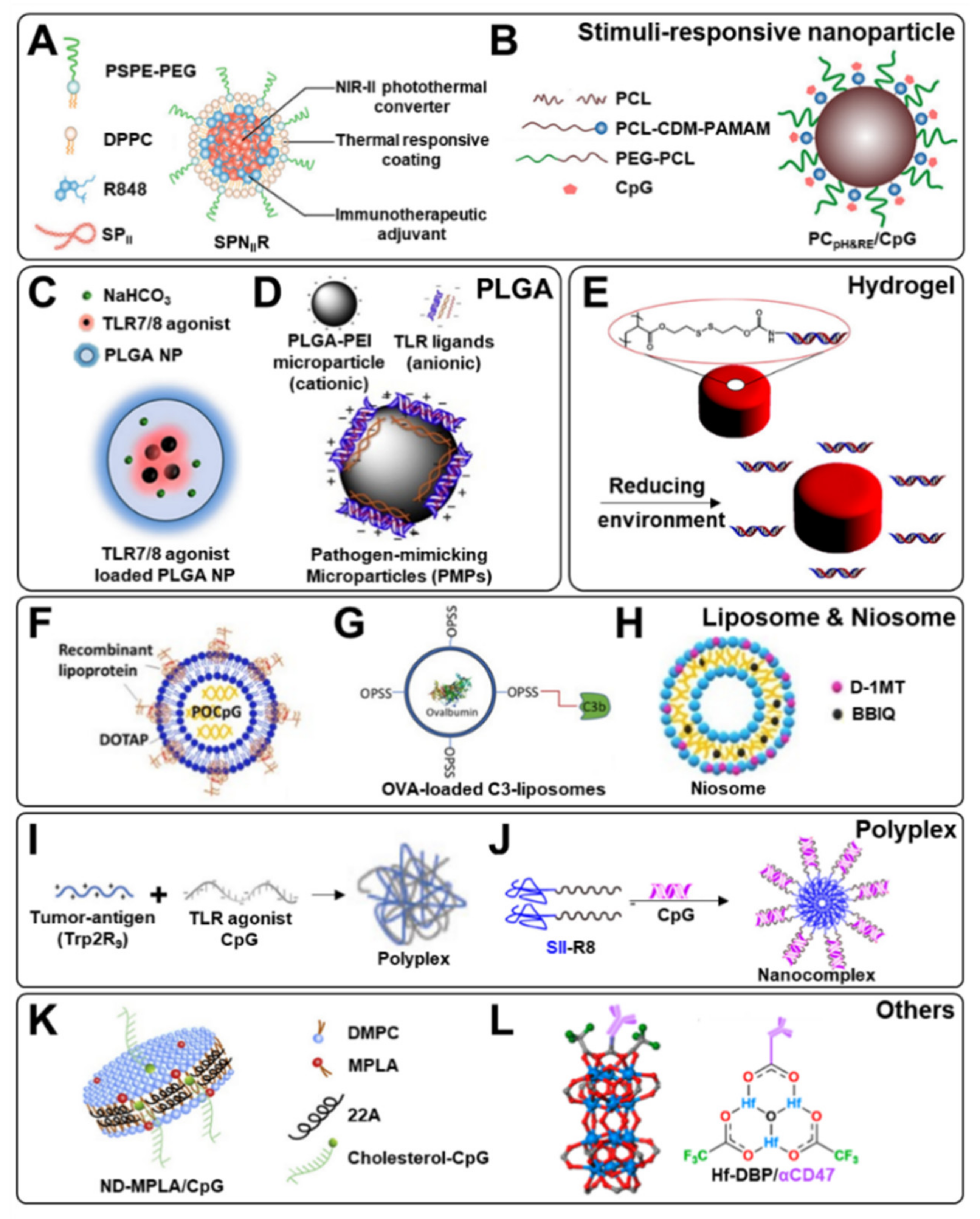
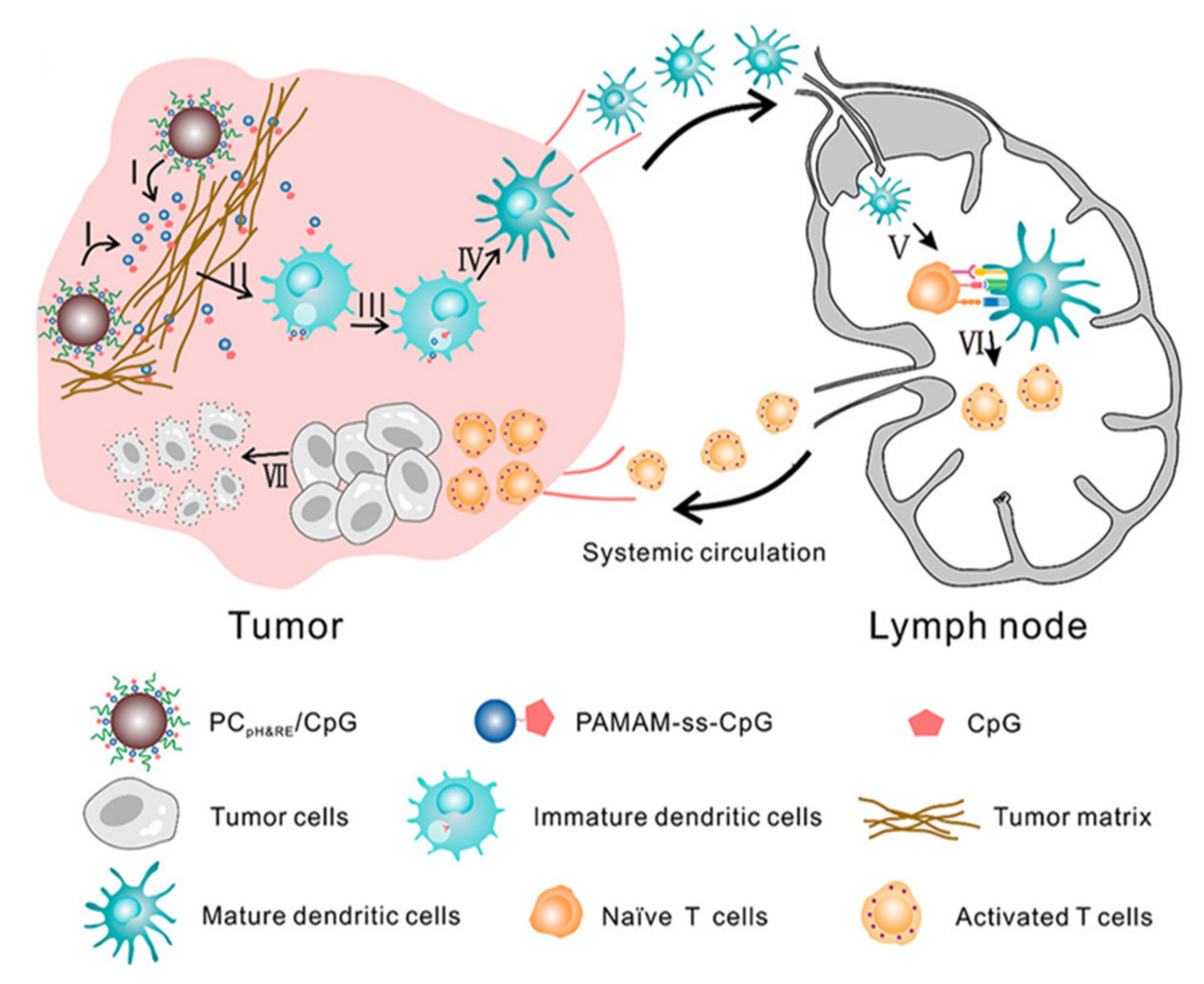
- Li, J.; Yu, X.; Jiang, Y.; He, S.; Zhang, Y.; Luo, Y.; Pu, K. Second Near-Infrared Photothermal Semiconducting Polymer Nanoadjuvant for Enhanced Cancer Immunotherapy. Adv. Mater. 2021, 33, e2003458. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, H.J.; Luo, Y.L.; Chen, Y.F.; Fan, Y.N.; Du, J.Z.; Wang, J. Programmable Delivery of Immune Adjuvant to Tumor-Infiltrating Dendritic Cells for Cancer Immunotherapy. Nano Lett. 2020, 20, 4882–4889. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Khanna, V.; Kucaba, T.A.; Zhang, W.; Sehgal, D.; Ferguson, D.M.; Griffith, T.S.; Panyam, J. TLR7/8 Agonist-Loaded Nanoparticles Augment NK Cell-Mediated Antibody-Based Cancer Immunotherapy. Mol. Pharm. 2020, 17, 2109–2124. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, P.; Qin, H.; Leleux, J.A.; Gwak, D.; Sakamaki, I.; Kwak, L.W.; Roy, K. The effect of combined IL10 siRNA and CpG ODN as pathogen-mimicking microparticles on Th1/Th2 cytokine balance in dendritic cells and protective immunity against B cell lymphoma. Biomaterials 2014, 35, 5491–5504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn, S.S.; Tian, S.; Blake, S.; Wang, J.; Galloway, A.L.; Murphy, A.; Pohlhaus, P.D.; Rolland, J.P.; Napier, M.E.; DeSimone, J.M. Reductively responsive siRNA-conjugated hydrogel nanoparticles for gene silencing. J. Am. Chem. Soc. 2012, 134, 7423–7430. [Google Scholar] [CrossRef] [Green Version]
- Shen, K.Y.; Liu, H.Y.; Yan, W.L.; Wu, C.C.; Lee, M.H.; Leng, C.H.; Liu, S.J. Liposomal TLR9 Agonist Combined with TLR2 Agonist-Fused Antigen Can Modulate Tumor Microenvironment through Dendritic Cells. Cancers 2020, 12, 810. [Google Scholar] [CrossRef] [Green Version]
- Francian, A.; Namen, S.; Stanley, M.; Mann, K.; Martinson, H.; Kullberg, M. Intratumoral delivery of antigen with complement C3-bound liposomes reduces tumor growth in mice. Nanotechnol. Biol. Med. 2019, 18, 326–335. [Google Scholar] [CrossRef]
- Rathee, J.; Kanwar, R.; Kaushik, D.; Salunke, D.B.; Mehta, S.K. Niosomes as efficient drug delivery modules for encapsulation of Toll-like receptor 7 agonists and IDO-inhibitor. Appl. Surf. Sci. 2020, 505, 144078. [Google Scholar] [CrossRef]
- Tsai, S.J.; Amerman, A.; Jewell, C.M. Altering Antigen Charge to Control Self-Assembly and Processing of Immune Signals During Cancer Vaccination. Front. Immunol. 2020, 11, 613830. [Google Scholar] [CrossRef] [PubMed]
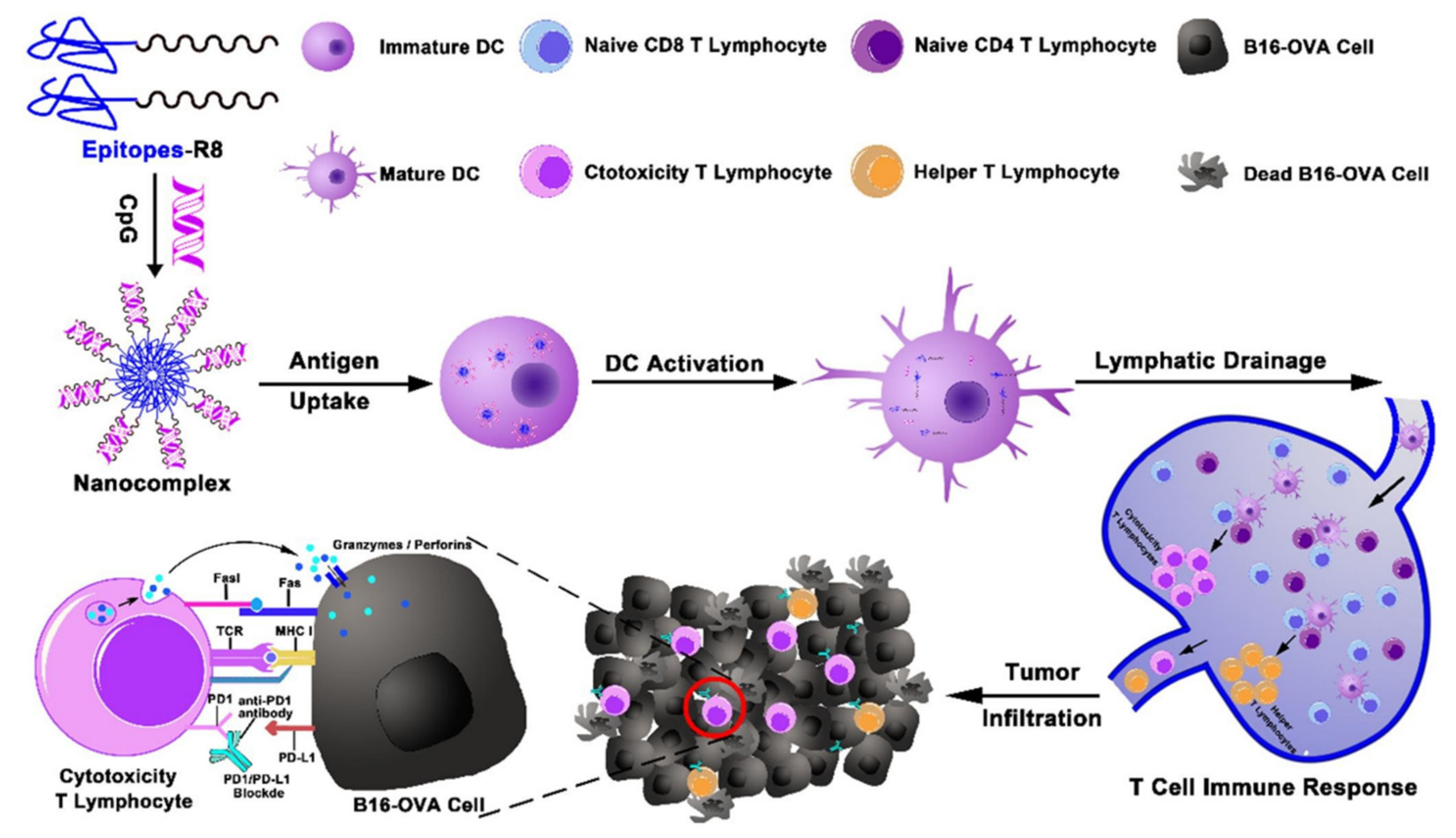
- Shi, X.; Song, H.; Wang, C.; Zhang, C.; Huang, P.; Kong, D.; Zhang, J.; Wang, W. Co-assembled and self-delivered epitope/CpG nanocomplex vaccine augments peptide immunogenicity for cancer immunotherapy. Chem. Eng. J. 2020, 399, 125854. [Google Scholar] [CrossRef]
- Kuai, R.; Sun, X.; Yuan, W.; Ochyl, L.J.; Xu, Y.; Hassani Najafabadi, A.; Scheetz, L.; Yu, M.Z.; Balwani, I.; Schwendeman, A.; et al. Dual TLR agonist nanodiscs as a strong adjuvant system for vaccines and immunotherapy. J. Control. Release 2018, 282, 131–139. [Google Scholar] [CrossRef]
- Ni, K.; Luo, T.; Culbert, A.; Kaufmann, M.; Jiang, X.; Lin, W. Nanoscale Metal-Organic Framework Co-delivers TLR-7 Agonists and Anti-CD47 Antibodies to Modulate Macrophages and Orchestrate Cancer Immunotherapy. J. Am. Chem. Soc. 2020, 142, 12579–12584. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Niu, L.; Larson, P.; Kucaba, T.A.; Murphy, K.A.; James, B.R.; Ferguson, D.M.; Griffith, T.S.; Panyam, J. Polymeric nanoparticles encapsulating novel TLR7/8 agonists as immunostimulatory adjuvants for enhanced cancer immunotherapy. Biomaterials 2018, 164, 38–53. [Google Scholar] [CrossRef]
- Kim, H.; Sehgal, D.; Kucaba, T.A.; Ferguson, D.M.; Griffith, T.S.; Panyam, J. Acidic pH-responsive polymer nanoparticles as a TLR7/8 agonist delivery platform for cancer immunotherapy. Nanoscale 2018, 10, 20851–20862. [Google Scholar] [CrossRef]
- Perry, J.L.; Tian, S.; Sengottuvel, N.; Harrison, E.B.; Gorentla, B.K.; Kapadia, C.H.; Cheng, N.; Luft, J.C.; Ting, J.P.; DeSimone, J.M.; et al. Pulmonary Delivery of Nanoparticle-Bound Toll-like Receptor 9 Agonist for the Treatment of Metastatic Lung Cancer. ACS Nano 2020, 14, 7200–7215. [Google Scholar] [CrossRef] [PubMed]
- Weeratna, R.D.; Makinen, S.R.; McCluskie, M.J.; Davis, H.L. TLR agonists as vaccine adjuvants: Comparison of CpG ODN and Resiquimod (R-848). Vaccine 2005, 23, 5263–5270. [Google Scholar] [CrossRef]
- Cui, L.; Wang, X.; Zhang, D. TLRs as a Promise Target Along With Immune Checkpoint Against Gastric Cancer. Front. Cell Dev. Biol. 2020, 8, 611444. [Google Scholar] [CrossRef]
- Li, K.; Qu, S.; Chen, X.; Wu, Q.; Shi, M. Promising Targets for Cancer Immunotherapy: TLRs, RLRs, and STING-Mediated Innate Immune Pathways. Int. J. Mol. Sci. 2017, 18, 404. [Google Scholar] [CrossRef]
- Ott, P.A.; Hodi, F.S.; Robert, C. CTLA-4 and PD-1/PD-L1 blockade: New immunotherapeutic modalities with durable clinical benefit in melanoma patients. Clin. Cancer Res. 2013, 19, 5300–5309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwai, Y.; Ishida, M.; Tanaka, Y.; Okazaki, T.; Honjo, T.; Minato, N. Involvement of PD-L1 on tumor cells in the escape from host immune system and tumor immunotherapy by PD-L1 blockade. Proc. Natl. Acad. Sci. USA 2002, 99, 12293–12297. [Google Scholar] [CrossRef] [Green Version]
- Urban-Wojciuk, Z.; Khan, M.M.; Oyler, B.L.; Fahraeus, R.; Marek-Trzonkowska, N.; Nita-Lazar, A.; Hupp, T.R.; Goodlett, D.R. The Role of TLRs in Anti-cancer Immunity and Tumor Rejection. Front. Immunol. 2019, 10, 2388. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Kim, S.B.; Lee, Y.; Song, E.C.; Kim, U.; Kim, H.Y.; Suh, J.H.; Goughnour, P.C.; Kim, Y.; Yoon, I.; et al. Endogenous TLR2 ligand embedded in the catalytic region of human cysteinyl-tRNA synthetase 1. J. Immunother. Cancer 2020, 8, e000277. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Vacher, J.; Allison, J.P. TLR1/2 ligand enhances antitumor efficacy of CTLA-4 blockade by increasing intratumoral Treg depletion. Proc. Natl. Acad. Sci. USA 2019, 116, 10453–10462. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Yan, Y.; Liu, J.; Huang, X.; Zhang, X.; Kirschning, C.; Xu, H.C.; Lang, P.A.; Dittmer, U.; Zhang, E.; et al. Toll-Like Receptor 7 Activation Enhances CD8+ T Cell Effector Functions by Promoting Cellular Glycolysis. Front. Immunol. 2019, 10, 2191. [Google Scholar] [CrossRef] [PubMed]
- Sato-Kaneko, F.; Yao, S.; Ahmadi, A.; Zhang, S.S.; Hosoya, T.; Kaneda, M.M.; Varner, J.A.; Pu, M.; Messer, K.S.; Guiducci, C.; et al. Combination immunotherapy with TLR agonists and checkpoint inhibitors suppresses head and neck cancer. JCI Insight 2017, 2, e93397. [Google Scholar] [CrossRef]
- Khanna, V.; Kim, H.; Zhang, W.; Larson, P.; Shah, M.; Griffith, T.S.; Ferguson, D.; Panyam, J. Novel TLR 7/8 agonists for improving NK cell mediated antibody-dependent cellular cytotoxicity (ADCC). Sci. Rep. 2021, 11, 3346. [Google Scholar] [CrossRef]
- Conroy, H.; Marshall, N.A.; Mills, K.H. TLR ligand suppression or enhancement of Treg cells? A double-edged sword in immunity to tumours. Oncogene 2008, 27, 168–180. [Google Scholar] [CrossRef] [Green Version]
- Farias, A.; Soto, A.; Puttur, F.; Goldin, C.J.; Sosa, S.; Gil, C.; Goldbaum, F.A.; Berguer, P.M. A TLR4 agonist improves immune checkpoint blockade treatment by increasing the ratio of effector to regulatory cells within the tumor microenvironment. Sci. Rep. 2021, 11, 15406. [Google Scholar] [CrossRef]
- Zahm, C.D.; Colluru, V.T.; McIlwain, S.J.; Ong, I.M.; McNeel, D.G. TLR Stimulation during T-cell Activation Lowers PD-1 Expression on CD8(+) T Cells. Cancer Immunol. Res. 2018, 6, 1364–1374. [Google Scholar] [CrossRef] [PubMed]
- Yin, P.; Liu, X.; Mansfield, A.S.; Harrington, S.M.; Li, Y.H.; Yan, Y.Y.; Dong, H.D. CpG-induced antitumor immunity requires IL-12 in expansion of effector cells and down-regulation of PD-1. Oncotarget 2016, 7, 70223–70231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, Y.; Kataoka, K.; Yamagishi, J.; Ogawa, S.; Seya, T.; Matsumoto, M. A TLR3-Specific Adjuvant Relieves Innate Resistance to PD-L1 Blockade without Cytokine Toxicity in Tumor Vaccine Immunotherapy. Cell Rep. 2017, 19, 1874–1887. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Hu, S.; Li, Y.; Jiang, T.T.; Jin, H.; Feng, L. Targeting nuclear acid-mediated immunity in cancer immune checkpoint inhibitor therapies. Signal. Transduct Target. Ther. 2020, 5, 270. [Google Scholar] [CrossRef] [PubMed]
- Chuang, Y.C.; Tseng, J.C.; Huang, L.R.; Huang, C.M.; Huang, C.F.; Chuang, T.H. Adjuvant Effect of Toll-Like Receptor 9 Activation on Cancer Immunotherapy Using Checkpoint Blockade. Front. Immunol. 2020, 11, 1075. [Google Scholar] [CrossRef]
- Vacchelli, E.; Galluzzi, L.; Eggermont, A.; Fridman, W.H.; Galon, J.; Sautes-Fridman, C.; Tartour, E.; Zitvogel, L.; Kroemer, G. Trial watch: FDA-approved Toll-like receptor agonists for cancer therapy. Oncoimmunology 2012, 1, 894–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cen, X.; Liu, S.; Cheng, K. The Role of Toll-Like Receptor in Inflammation and Tumor Immunity. Front. Pharmacol. 2018, 9, 878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordon, Y. Therapeutic cancer vaccine approved. Nat. Rev. Immunol. 2010, 10, 380. [Google Scholar] [CrossRef]
- Li, J.K.; Balic, J.J.; Yu, L.; Jenkins, B. TLR Agonists as Adjuvants for Cancer Vaccines. In Regulation of Inflammatory Signaling in Health and Disease; Xu, D., Ed.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 195–212. [Google Scholar]
- Dubensky, T.W., Jr.; Reed, S.G. Adjuvants for cancer vaccines. Semin. Immunol. 2010, 22, 155–161. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vacchelli, E.; Eggermont, A.; Fridman, W.H.; Galon, J.; Sautes-Fridman, C.; Tartour, E.; Zitvogel, L.; Kroemer, G. Trial Watch: Experimental Toll-like receptor agonists for cancer therapy. Oncoimmunology 2012, 1, 699–716. [Google Scholar] [CrossRef] [Green Version]
- Fuge, O.; Vasdev, N.; Allchorne, P.; Green, J.S. Immunotherapy for bladder cancer. Res. Rep. Urol. 2015, 7, 65–79. [Google Scholar]
- Ohadian Moghadam, S.; Nowroozi, M.R. Toll-like receptors: The role in bladder cancer development, progression and immunotherapy. Scand. J. Immunol. 2019, 90, e12818. [Google Scholar] [CrossRef] [Green Version]
- Askeland, E.J.; Newton, M.R.; O’Donnell, M.A.; Luo, Y. Bladder Cancer Immunotherapy: BCG and Beyond. Adv. Urol. 2012, 2012, 181987. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, T.F. Clinical update of the AS04-adjuvanted human papillomavirus-16/18 cervical cancer vaccine, Cervarix. Adv. Ther. 2009, 26, 983–998. [Google Scholar] [CrossRef]
- Vasilakos, J.P.; Tomai, M.A. The use of Toll-like receptor 7/8 agonists as vaccine adjuvants. Expert Rev. Vaccines 2013, 12, 809–819. [Google Scholar] [CrossRef]
- Adams, S.; Kozhaya, L.; Martiniuk, F.; Meng, T.C.; Chiriboga, L.; Liebes, L.; Hochman, T.; Shuman, N.; Axelrod, D.; Speyer, J.; et al. Topical TLR7 agonist imiquimod can induce immune-mediated rejection of skin metastases in patients with breast cancer. Clin. Cancer Res. 2012, 18, 6748–6757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagchi, A.; Herrup, E.A.; Warren, H.S.; Trigilio, J.; Shin, H.S.; Valentine, C.; Hellman, J. MyD88-dependent and MyD88-independent pathways in synergy, priming, and tolerance between TLR agonists. J. Immunol. 2007, 178, 1164–1171. [Google Scholar] [CrossRef]
- Warger, T.; Osterloh, P.; Rechtsteiner, G.; Fassbender, M.; Heib, V.; Schmid, B.; Schmitt, E.; Schild, H.; Radsak, M.P. Synergistic activation of dendritic cells by combined Toll-like receptor ligation induces superior CTL responses in vivo. Blood 2006, 108, 544–550. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, D.; Yong, M.; Schroder, W.; Black, M.; Tirrell, M.; Olive, C. Dual stimulation of MyD88-dependent Toll-like receptors induces synergistically enhanced production of inflammatory cytokines in murine bone marrow-derived dendritic cells. J. Infect. Dis. 2010, 202, 318–329. [Google Scholar] [CrossRef] [Green Version]
- Manna, S.; Maiti, S.; Shen, J.; Du, W.; Esser-Kahn, A.P. Pathogen-like Nanoassemblies of Covalently Linked TLR Agonists Enhance CD8 and NK Cell-Mediated Antitumor Immunity. ACS Cent. Sci. 2020, 6, 2071–2078. [Google Scholar] [CrossRef]
- Madan-Lala, R.; Pradhan, P.; Roy, K. Combinatorial Delivery of Dual and Triple TLR Agonists via Polymeric Pathogen-like Particles Synergistically Enhances Innate and Adaptive Immune Responses. Sci. Rep. 2017, 7, 2530. [Google Scholar] [CrossRef] [Green Version]
- Francian, A.; Widmer, A.; Olsson, T.; Ramirez, M.; Heald, D.; Rasic, K.; Adams, L.; Martinson, H.; Kullberg, M. Delivery of toll-like receptor agonists by complement C3-targeted liposomes activates immune cells and reduces tumour growth. J. Drug Target. 2021, 29, 754–760. [Google Scholar] [CrossRef] [PubMed]
- Hess, K.L.; Medintz, I.L.; Jewell, C.M. Designing inorganic nanomaterials for vaccines and immunotherapies. Nano Today 2019, 27, 73–98. [Google Scholar] [CrossRef]
- Liu, L.; Ma, P.; Wang, H.; Zhang, C.; Sun, H.; Wang, C.; Song, C.; Leng, X.; Kong, D.; Ma, G. Immune responses to vaccines delivered by encapsulation into and/or adsorption onto cationic lipid-PLGA hybrid nanoparticles. J. Control. Release 2016, 225, 230–239. [Google Scholar] [CrossRef]
- Irvine, D.J.; Hanson, M.C.; Rakhra, K.; Tokatlian, T. Synthetic Nanoparticles for Vaccines and Immunotherapy. Chem. Rev. 2015, 115, 11109–11146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gammon, J.M.; Dold, N.M.; Jewell, C.M. Improving the clinical impact of biomaterials in cancer immunotherapy. Oncotarget 2016, 7, 15421. [Google Scholar] [CrossRef] [PubMed]
- Bookstaver, M.L.; Tsai, S.J.; Bromberg, J.S.; Jewell, C.M. Improving Vaccine and Immunotherapy Design Using Biomaterials. Trends Immunol. 2018, 39, 135–150. [Google Scholar] [CrossRef]
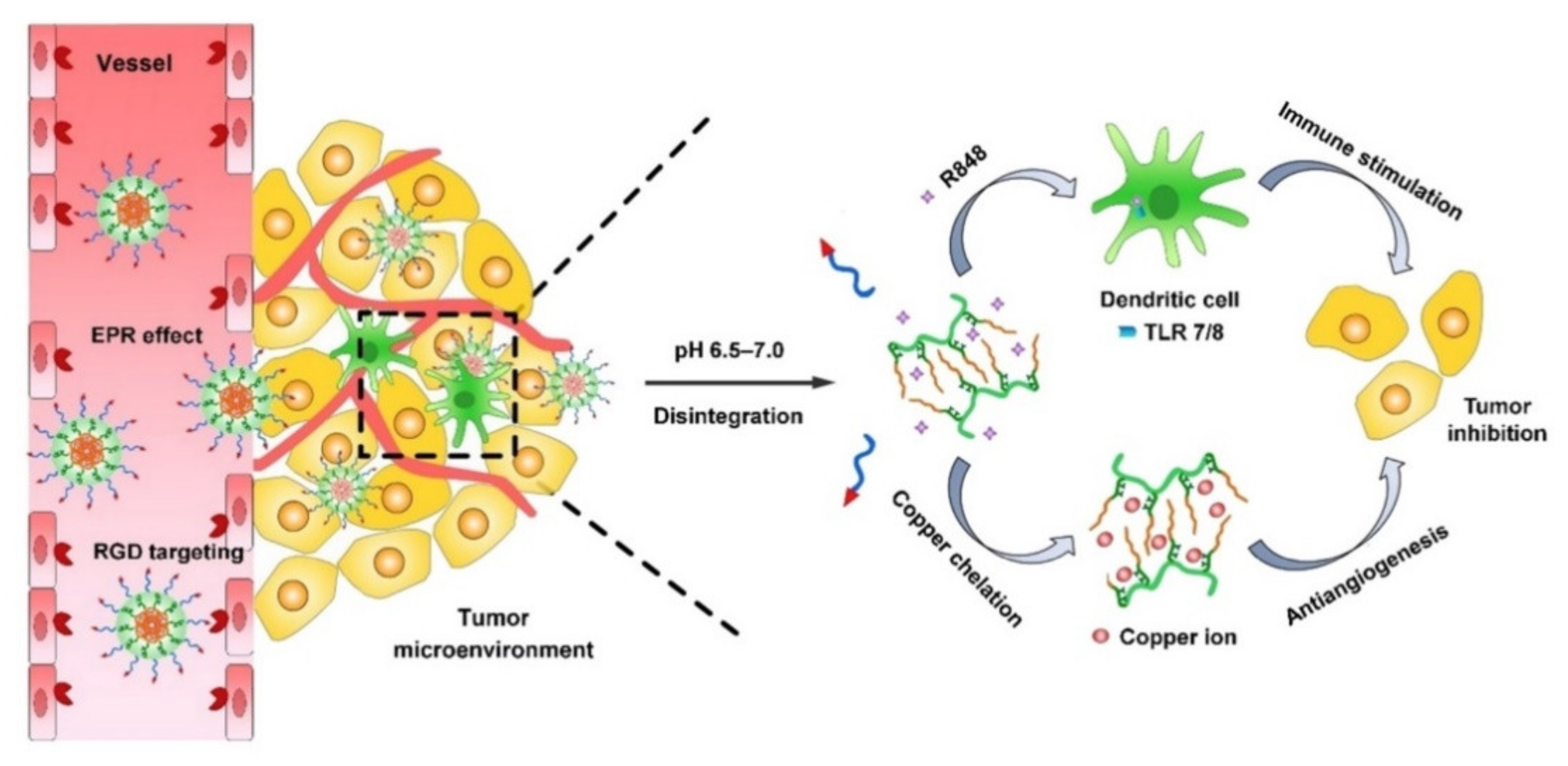
- Zhou, P.; Qin, J.; Zhou, C.; Wan, G.; Liu, Y.; Zhang, M.; Yang, X.; Zhang, N.; Wang, Y. Multifunctional nanoparticles based on a polymeric copper chelator for combination treatment of metastatic breast cancer. Biomaterials 2019, 195, 86–99. [Google Scholar] [CrossRef]
- Jia, X.; Guo, J.; Guo, S.; Zhao, T.; Liu, X.; Cheng, C.; Wang, L.; Zhang, B.; Meng, C.; Jia, H.; et al. Antitumor effects and mechanisms of CpG ODN combined with attenuated Salmonella-delivered siRNAs against PD-1. Int. Immunopharmacol. 2021, 90, 107052. [Google Scholar] [CrossRef]
- Liu, D.; Liu, J.; Ma, B.; Deng, B.; Leng, X.; Kong, D.; Liu, L. A simple self-adjuvanting biomimetic nanovaccine self-assembled with the conjugate of phospholipids and nucleotides can induce a strong cancer immunotherapeutic effect. Biomater. Sci. 2021, 9, 84–92. [Google Scholar] [CrossRef]
- Scheetz, L.M.; Yu, M.; Li, D.; Castro, M.G.; Moon, J.J.; Schwendeman, A. Synthetic HDL Nanoparticles Delivering Docetaxel and CpG for Chemoimmunotherapy of Colon Adenocarcinoma. Int. J. Mol. Sci. 2020, 21, 1777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, M.; Ding, X.; Zhao, R.; Liu, X.; Shen, H.; Cai, C.; Ferrari, M.; Wang, H.Y.; Wang, R.F. Co-delivery of tumor antigen and dual toll-like receptor ligands into dendritic cell by silicon microparticle enables efficient immunotherapy against melanoma. J. Control. Release 2018, 272, 72–82. [Google Scholar] [CrossRef]
- Da Silva, C.G.; Camps, M.G.M.; Li, T.; Chan, A.B.; Ossendorp, F.; Cruz, L.J. Co-delivery of immunomodulators in biodegradable nanoparticles improves therapeutic efficacy of cancer vaccines. Biomaterials 2019, 220, 119417. [Google Scholar] [CrossRef] [PubMed]
- Mohsen, M.O.; Gomes, A.C.; Vogel, M.; Bachmann, M.F. Interaction of Viral Capsid-Derived Virus-Like Particles (VLPs) with the Innate Immune System. Vaccines 2018, 6, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Lemke-Miltner, C.D.; Wongpattaraworakul, W.; Wang, Z.; Chan, C.H.F.; Salem, A.K.; Weiner, G.J.; Simons, A.L. In situ immunization of a TLR9 agonist virus-like particle enhances anti-PD1 therapy. J. Immunother. Cancer 2020, 8, e000940. [Google Scholar] [CrossRef] [PubMed]
- Bogani, G.; Raspagliesi, F.; Ditto, A.; de la Fuente, J. The Adoption of Viral Capsid-Derived Virus-Like Particles (VLPs) for Disease Prevention and Treatments. Vaccines 2020, 8, 432. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TLR | TLR Agonist | Delivery Platform | Target Cancer | Ref |
|---|---|---|---|---|
| Mechanism | Improvements | |||
| 7/8 | GU-rich ORN | RNA/DNA hydrogel | Colorectal cancer | [31] |
|
| |||
| 7/8 | R848 | Hydrophobic PANI with hydrophilic GCS | Colorectal cancer | [32] |
|
| |||
| 7/8 | R848 | SPNIIR | Breast cancer | [34] |
|
| |||
| 7/8 | Imidazoquinoline-based agonist 522 | PLGA | Melanoma Bladder cancer | [46] |
|
| |||
| 7/8 | Imidazoquinoline-based agonist 522 | Acidic pH-responsive PLGA | Melanoma | [47] |
|
| |||
| 7/8 | Imidazoquinoline-based agonist 522 | Acidic pH-responsive PLGA | Lung cancer, Mammary carcinoma, Leukemia | [36] |
|
| |||
| 9 | CpG ODN | PAMAM dendrimers | Pancreatic cancer | [35] |
|
| |||
| 9 | CpG ODN | PRINT nanoparticles | Lung cancer | [48] |
|
| |||
| Agent/ Source | Target Agonist | Cancer Type | Induction of Immune Responses | Mediation Step for Antitumor Outcomes | Possible Drawback | Ref |
|---|---|---|---|---|---|---|
| BCG/ Mycobacteri-um bovis | 2/4 | Bladder cancer |
|
|
| [67,73,74,75] |
| MPLA/ Salmonella minnesota | 2/4 | Cervical cancer |
|
| Fever, chills, and rigor, bronchospasm, hypotensive | [67,76] |
| Imiquimod/ Imidazoqui-noline | 7 | Breast cancer |
|
|
| [59,69,70] |
| TLR | TLR Agonist | Delivery Platform | Target Cancer | Ref |
|---|---|---|---|---|
| Mechanism | Improvements | |||
| 2, 9 | rlipoE7m, CpG ODN | DOTAP liposome | Lung cancer | [39] |
|
| |||
| 2/6_7 | Pam2CSK4C-PEG4-DBCO-2Bxy | Amphiphile OL-DSPOE micelles | Melanoma | [82] |
|
| |||
| 1/2, 4, 7/8, 9 | Pam3CSK4, MPLA. R837, CpG ODN | PLGA particles | - | [83] |
|
| |||
| 4, 7/8, 9 | MPLA, R848, CpG 1826 | C3 liposomes | Breast cancer | [84] |
|
| |||
| 4, 9 | MPLA, CpG ODN | sHDL | Melanoma, Lung cancer | [44] |
|
| |||
| TLR | TLR Agonist | Co-Factor | Delivery Platform | Target Cancer | Ref |
|---|---|---|---|---|---|
| Mechanism | Improvements | ||||
| 7 | BBIQ | D-1MT | Niosome | - | [41] |
|
| ||||
| 7 | IMD | Anti-CD47 | Hf-DBP nMOFs | Colorectal cancer | [45] |
|
| ||||
| 7/8 | R848 | RGD-PEG-PGA | RPTDH (Copper chelating polymer) | Breast cancer | [90] |
|
| ||||
| 9 | CpG ODN | SVYDFFVWL peptide antigen | Polyplex-like NPs | Melanoma | [42] |
|
| ||||
| 9 | CpG ODN | Cell-penetrating peptide (R8)-conjugated melanoma specific peptide epitope | Peptide/CpG nanocomplex | Melanoma | [43] |
|
| ||||
| 9 | CpG ODN | PD-1-siRNA | attenuated Salmonella | Melanoma | [91] |
|
| ||||
| 9 | CpG ODN | OVA antigen | DOPE-based amphiphiles | Lymphoma | [92] |
|
| ||||
| 9 | CpG ODN | DTX chemo-drug | sHDL | Colorectal cancer | [93] |
|
| ||||
| 4, 9 | CpG ODN, MPLA | TRP2 peptide | MSV microparticles | Melanoma | [94] |
|
| ||||
| 3, 7/8 | Poly(I:C), R848 | MIP3α | PLGA NPs | Lung cancer, Rhabdomyosarcoma | [95] |
|
| ||||
| TLR | TLR Agonist | Phase | Co-Factors | Conditions | Status | Ref. |
|---|---|---|---|---|---|---|
| 9 | VLP-encapsulated CMP-001 (TLR 9 agonist) | II | Pembrolizumab | Melanoma of Unkown Primary and 5 more | Recruiting | NCT 04708418 |
| I, II | INCAGN01949 | Locally Advanced Malignant Solid Neoplasm and 3 more | Not yet recruiting | NCT 04387071 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, S.; Choi, Y.; Kim, K. Engineering Therapeutic Strategies in Cancer Immunotherapy via Exogenous Delivery of Toll-like Receptor Agonists. Pharmaceutics 2021, 13, 1374. https://doi.org/10.3390/pharmaceutics13091374
Jeong S, Choi Y, Kim K. Engineering Therapeutic Strategies in Cancer Immunotherapy via Exogenous Delivery of Toll-like Receptor Agonists. Pharmaceutics. 2021; 13(9):1374. https://doi.org/10.3390/pharmaceutics13091374
Chicago/Turabian StyleJeong, Sehwan, Yunyoung Choi, and Kyobum Kim. 2021. "Engineering Therapeutic Strategies in Cancer Immunotherapy via Exogenous Delivery of Toll-like Receptor Agonists" Pharmaceutics 13, no. 9: 1374. https://doi.org/10.3390/pharmaceutics13091374