
Comprehensive Evidence of Carrier-Mediated Distribution of Amantadine to the Retina across the Blood–Retinal Barrier in Rats


Abstract
:
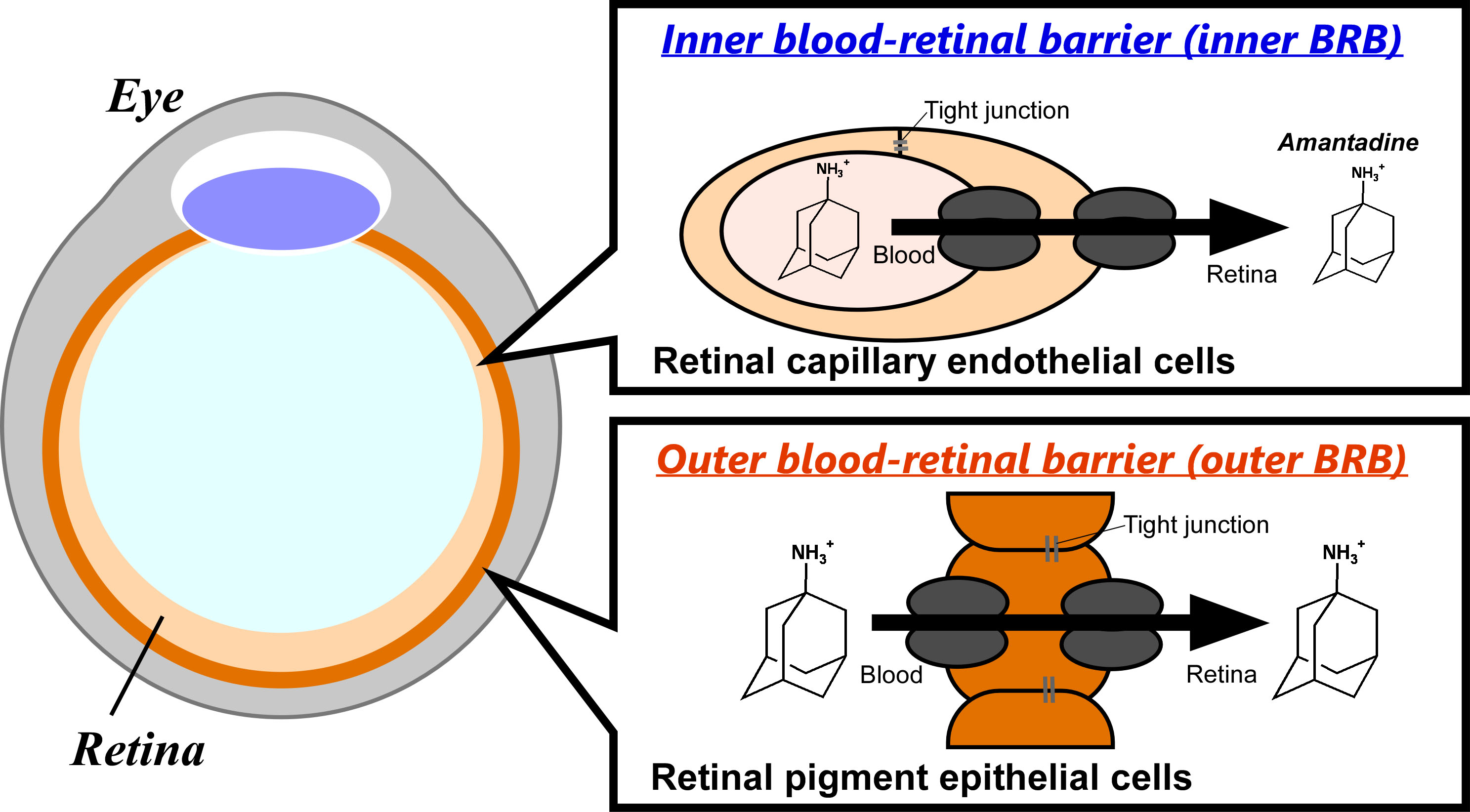
1. Introduction
2. Materials and Methods
2.1. Animals and Reagents
2.2. Assessment of In Vivo Blood-to-Retina Transport
2.3. In Vitro Transport Study
2.4. Data and Statistical Analyses
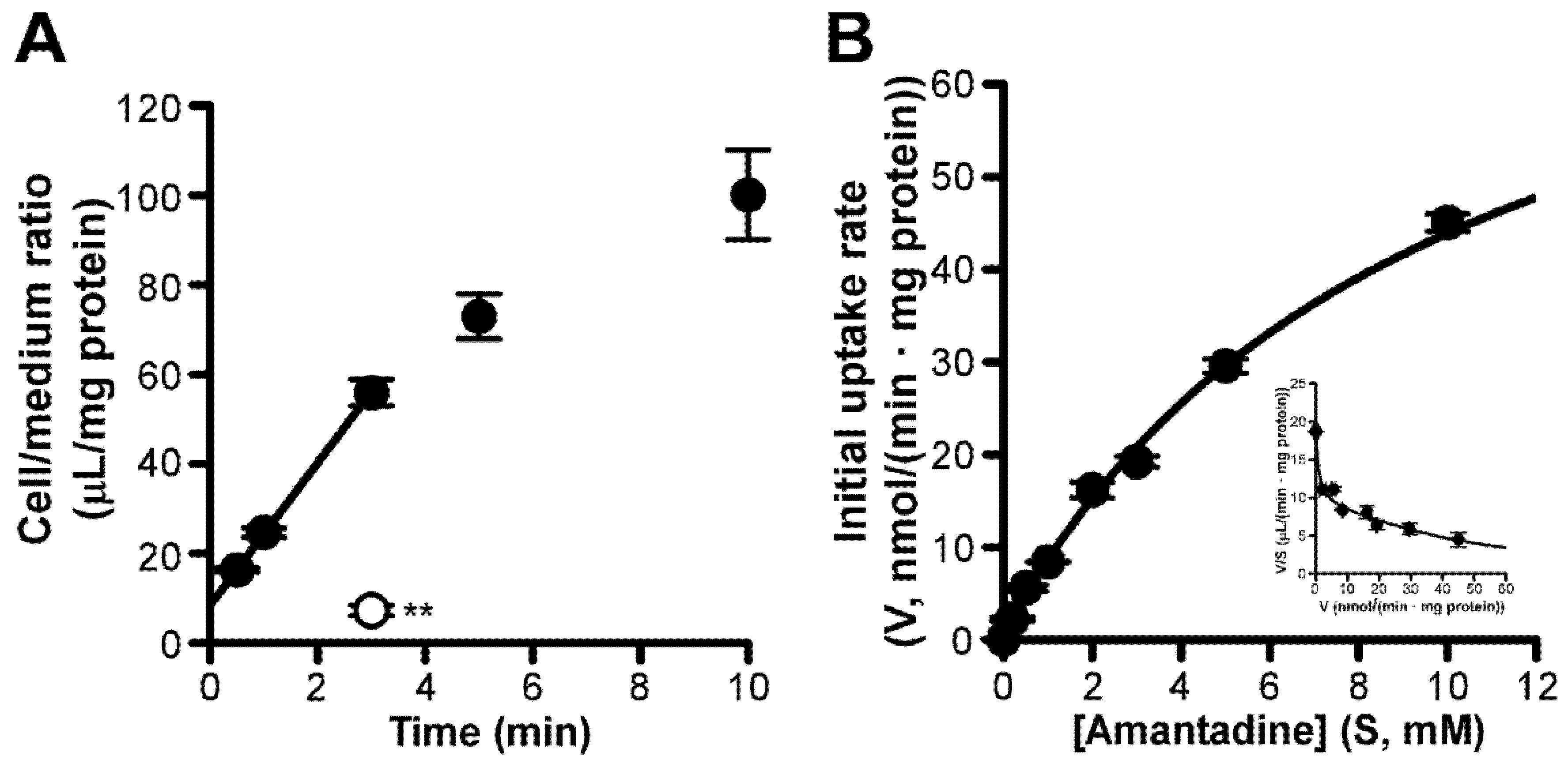
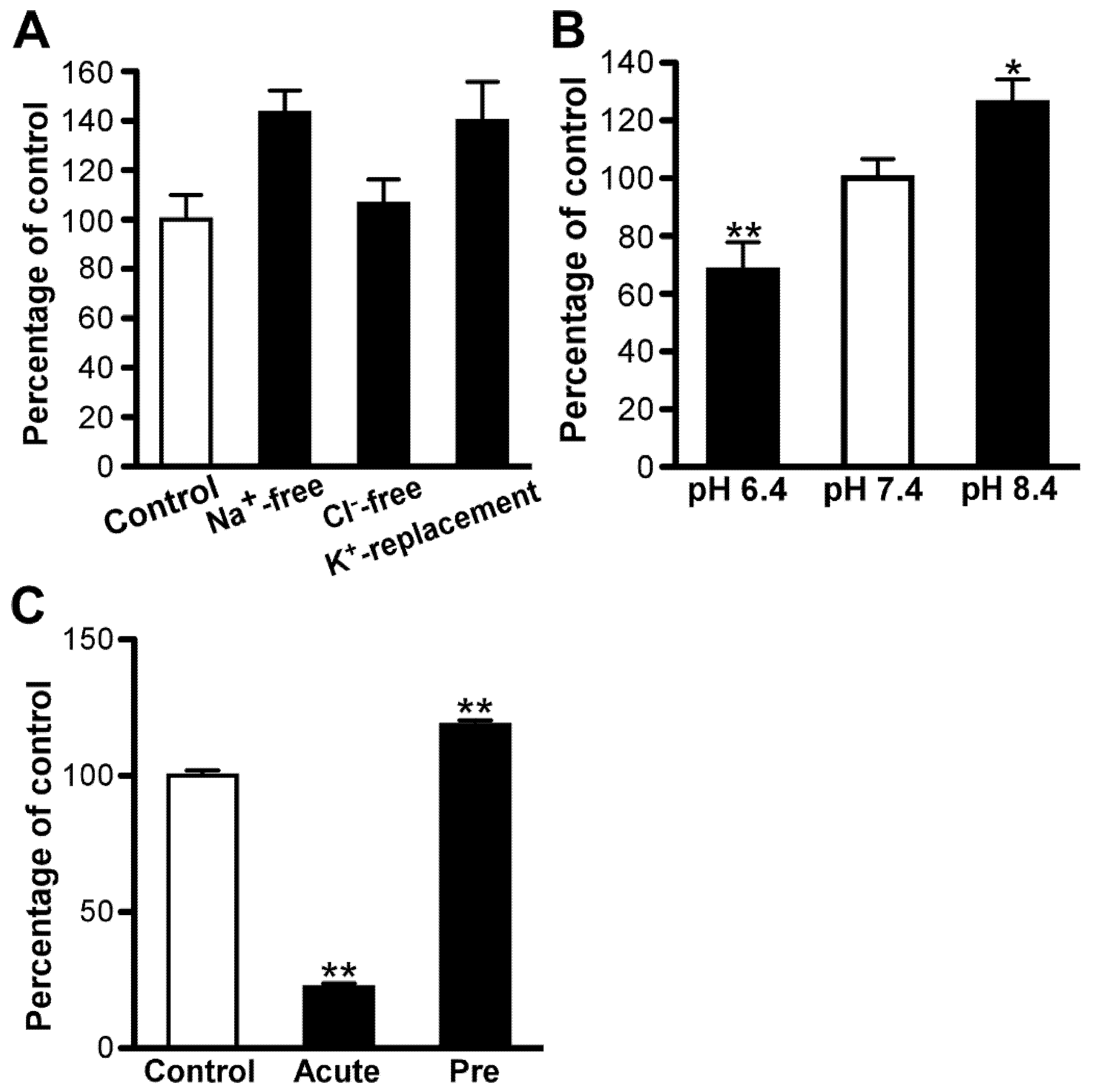
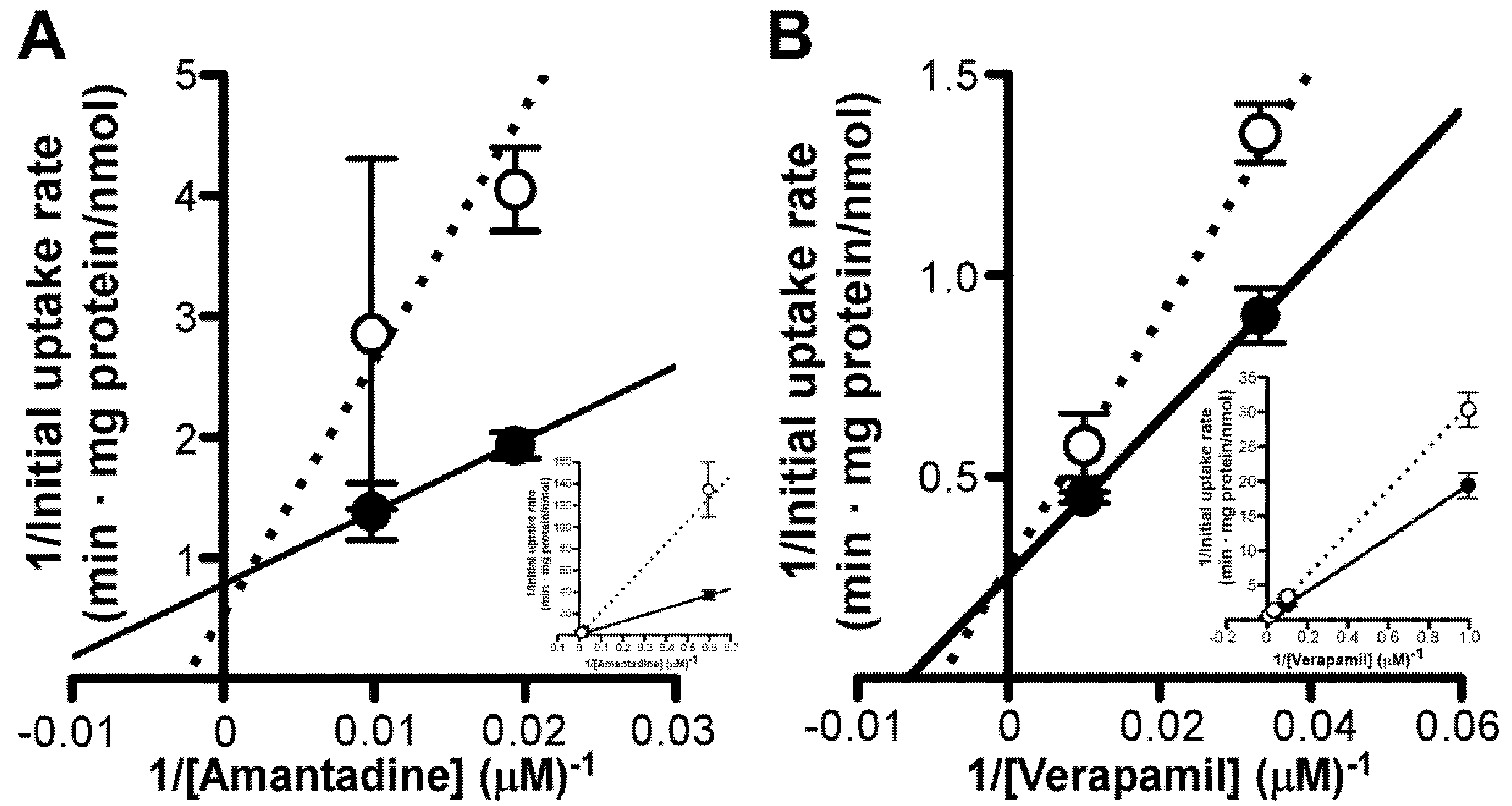
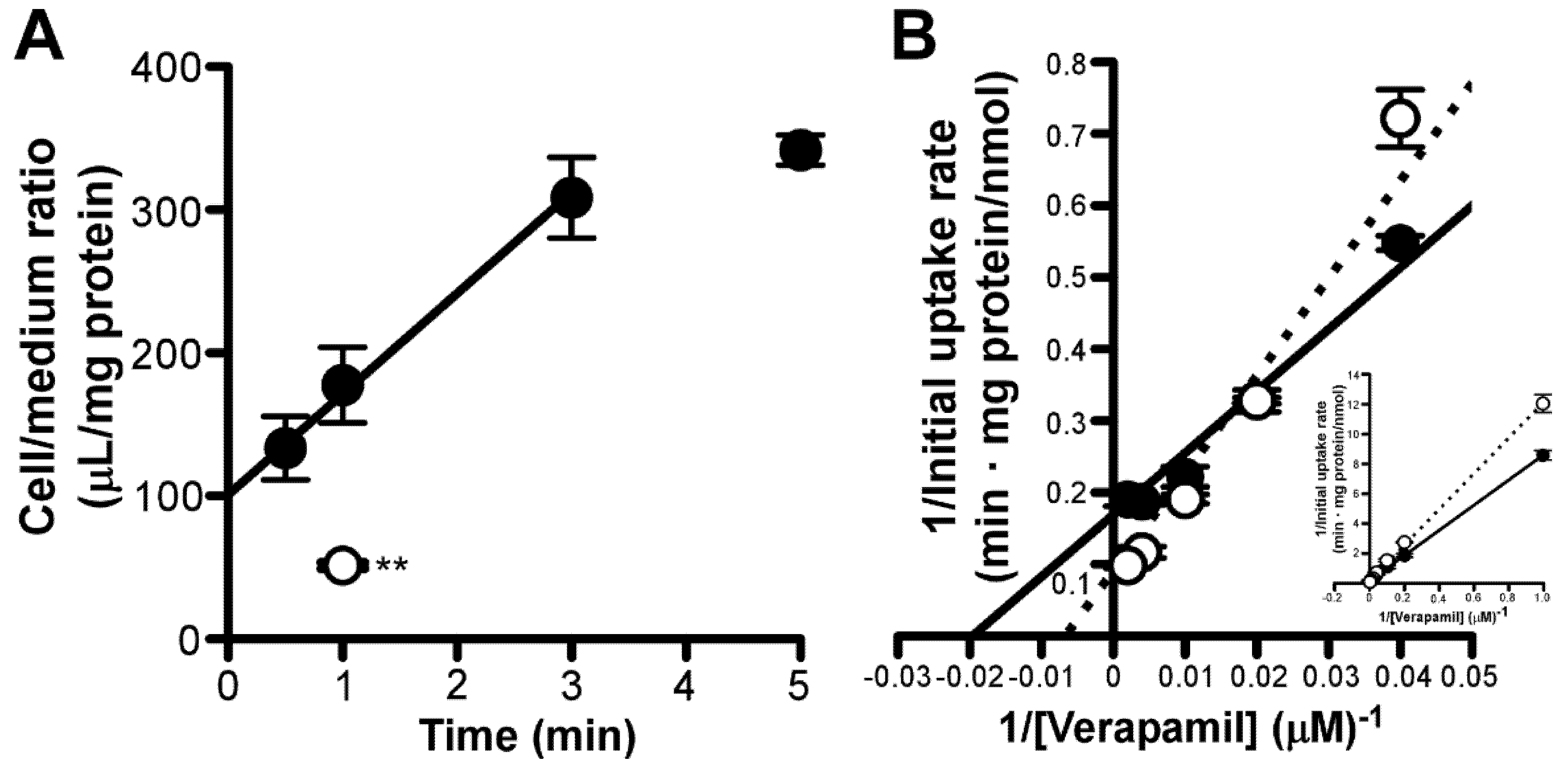
3. Results
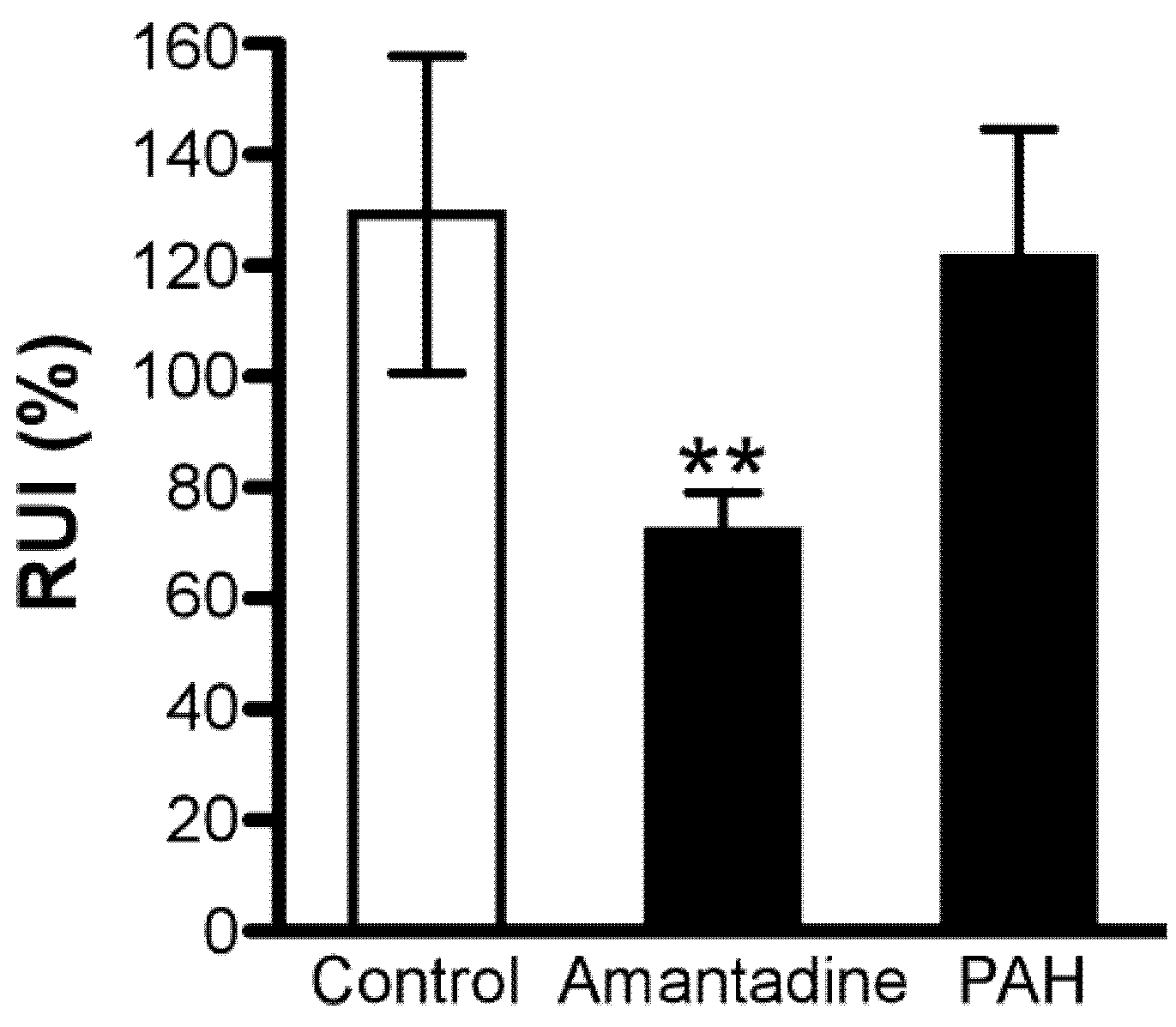
3.1. In Vivo Blood-to-Retina Transport of [3H]Amantadine across the BRB
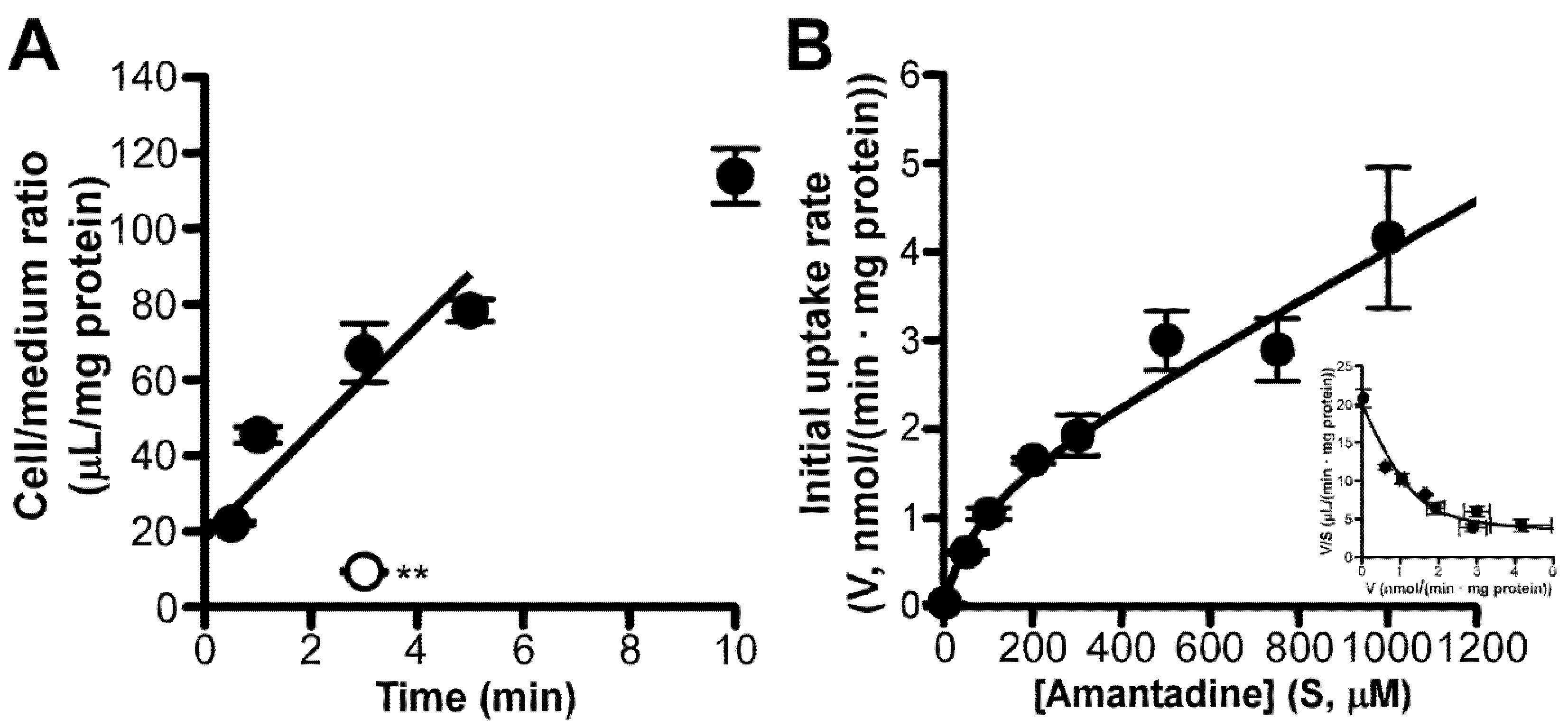
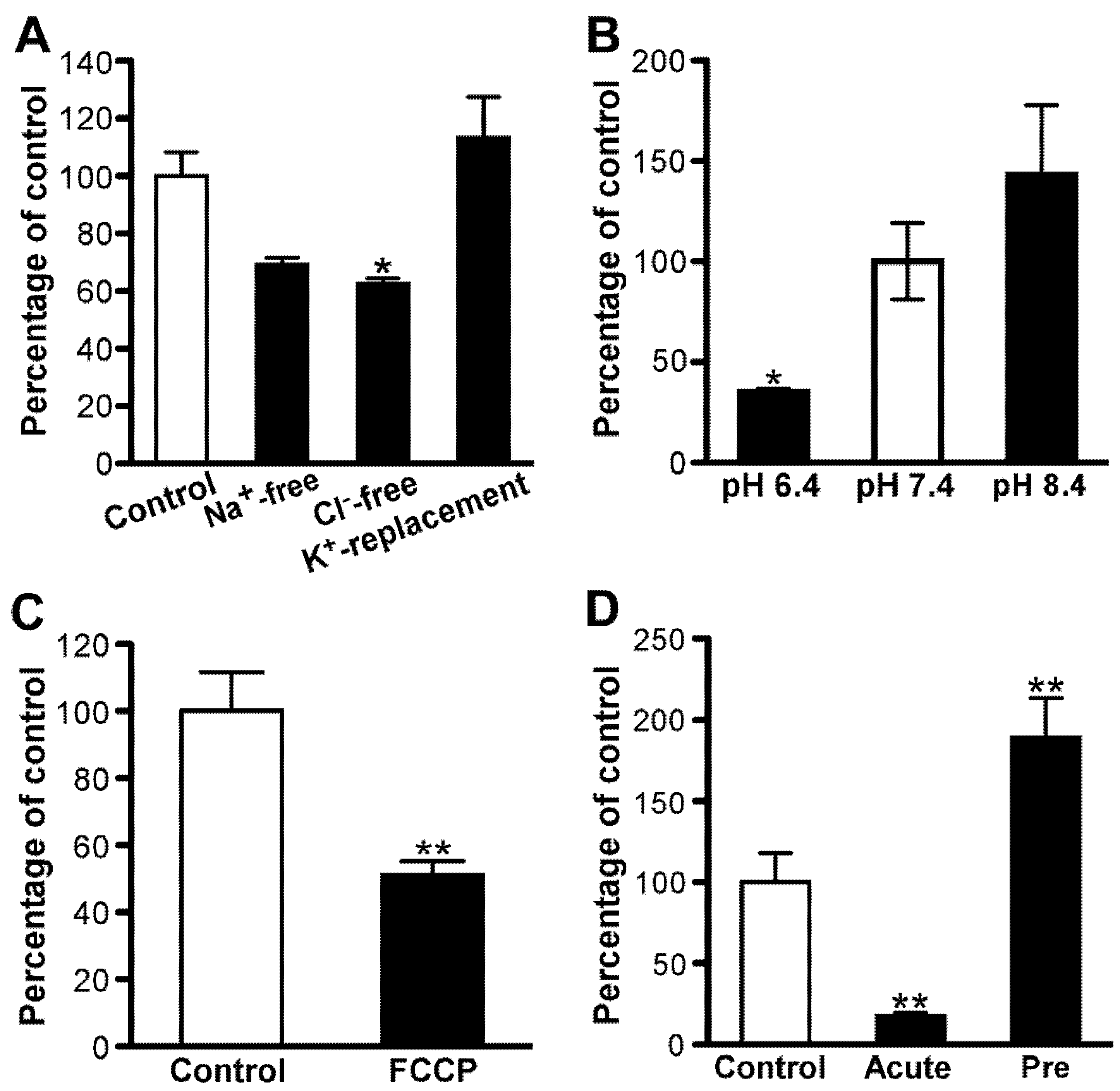
3.2. Amantadine Uptake Properties in TR-iBRB2 Cells
3.3. Amantadine Uptake Properties in RPE-J Cells
3.4. Inhibition of [3H]Amantadine Transport by Drugs/Compounds
3.5. Mutual Effect of Amantadine and Verapamil on Uptake by TR-iBRB2 Cells
3.6. Effect of Amantadine on Concentration-Dependent Verapamil Uptake by RPE-J Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McMonnies, C.W. Glaucoma history and risk factors. J. Optom. 2017, 10, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Lechner, J.; O’Leary, O.E.; Stitt, A.W. The pathology associated with diabetic retinopathy. Vis. Res. 2017, 139, 7–14. [Google Scholar] [CrossRef]
- Fechtner, R.D.; Weinreb, R.N. Mechanisms of optic nerve damage in primary open angle glaucoma. Surv. Ophthalmol. 1994, 39, 23–42. [Google Scholar] [CrossRef]
- Lipton, S.A. Possible role for memantine in protecting retinal ganglion cells from glaucomatous damage. Surv. Ophthalmol. 2003, 48, S38–S46. [Google Scholar] [CrossRef]
- Luo, X.; Heidinger, V.; Picaud, S.; Lambrou, G.; Dreyfus, H.; Sahel, J.; Hicks, D. Selective excitotoxic degeneration of adult pig retinal ganglion cells in vitro. Invest. Ophthalmol. Vis. Sci. 2001, 42, 1096–1106. [Google Scholar]
- Chidlow, G.; Osborne, N.N. Rat retinal ganglion cell loss caused by kainate, NMDA and ischemia correlates with a reduction in mRNA and protein of Thy-1 and neurofilament light. Brain Res. 2003, 963, 298–306. [Google Scholar] [CrossRef]
- Hare, W.A.; Wheeler, L. Experimental glutamatergic excitotoxicity in rabbit retinal ganglion cells: Block by memantine. Invest. Ophthalmol. Vis. Sci. 2009, 50, 2940–2948. [Google Scholar] [CrossRef]
- Hare, W.; WoldeMussie, E.; Lai, R.; Ton, H.; Ruiz, G.; Feldmann, B.; Wijono, M.; Chun, T.; Wheeler, L. Efficacy and safety of memantine, an NMDA-type open-channel blocker, for reduction of retinal injury associated with experimental glaucoma in rat and monkey. Surv. Ophthalmol. 2001, 45, S284–S289. [Google Scholar] [CrossRef]
- Weinreb, R.N.; Liebmann, J.M.; Cioffi, G.A.; Goldberg, I.; Brandt, J.D.; Johnson, C.A.; Zangwill, L.M.; Schneider, S.; Badger, H.; Bejanian, M. Oral memantine for the treatment of glaucoma: Design and results of 2 randomized, placebo-controlled, phase 3 studies. Ophthalmology 2018, 125, 1874–1885. [Google Scholar] [CrossRef] [Green Version]
- Bucolo, C.; Platania, C.B.M.; Drago, F.; Bonfiglio, V.; Reibaldi, M.; Avitabile, T.; Uva, M. Novel therapeutics in glaucoma management. Curr. Neuropharmacol. 2018, 16, 978–992. [Google Scholar] [CrossRef]
- Cunha-Vaz, J.G. The blood-retinal barriers system. Basic concepts and clinical evaluation. Exp. Eye. Res. 2004, 78, 715–721. [Google Scholar] [CrossRef]
- Liu, L.; Liu, X. Roles of drug transporters in blood-retinal barrier. Adv. Exp. Med. Biol. 2019, 1141, 467–504. [Google Scholar] [PubMed]
- Aoki, F.Y.; Sitar, D.S.; Ogilvie, R.I. Amantadine kinetics in healthy young subjects after long-term dosing. Clin. Pharmacol. Ther. 1979, 26, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Muller, F.; Weitz, D.; Derdau, V.; Sandvoss, M.; Mertsch, K.; Konig, J.; Fromm, M.F. Contribution of MATE1 to renal secretion of the NMDA receptor antagonist memantine. Mol. Pharm. 2017, 14, 2991–2998. [Google Scholar] [CrossRef] [PubMed]
- Kooijmans, S.A.; Senyschyn, D.; Mezhiselvam, M.M.; Morizzi, J.; Charman, S.A.; Weksler, B.; Romero, I.A.; Couraud, P.O.; Nicolazzo, J.A. The involvement of a Na(+)- and Cl(-)-dependent transporter in the brain uptake of amantadine and rimantadine. Mol. Pharm. 2012, 9, 883–893. [Google Scholar] [CrossRef] [PubMed]
- Goralski, K.B.; Lou, G.; Prowse, M.T.; Gorboulev, V.; Volk, C.; Koepsell, H.; Sitar, D.S. The cation transporters rOCT1 and rOCT2 interact with bicarbonate but play only a minor role for amantadine uptake into rat renal proximal tubules. J. Pharmacol. Exp. Ther. 2002, 303, 959–968. [Google Scholar] [CrossRef]
- Busch, A.E.; Karbach, U.; Miska, D.; Gorboulev, V.; Akhoundova, A.; Volk, C.; Arndt, P.; Ulzheimer, J.C.; Sonders, M.S.; Baumann, C.; et al. Human neurons express the polyspecific cation transporter hOCT2, which translocates monoamine neurotransmitters, amantadine, and memantine. Mol. Pharmacol. 1998, 54, 342–352. [Google Scholar] [CrossRef] [Green Version]
- Tachikawa, M.; Takeda, Y.; Tomi, M.; Hosoya, K. Involvement of OCTN2 in the transport of acetyl-L-carnitine across the inner blood-retinal barrier. Invest. Ophthalmol. Vis. Sci. 2010, 51, 430–436. [Google Scholar] [CrossRef]
- Kubo, Y.; Yamamoto, M.; Matsunaga, K.; Usui, T.; Akanuma, S.I.; Hosoya, K.I. Retina-to-blood transport of 1-methyl-4-phenylpyridinium involves carrier-mediated process at the blood-retinal barrier. J. Pharm. Sci. 2017, 106, 2583–2591. [Google Scholar] [CrossRef] [Green Version]
- Kubo, Y.; Kusagawa, Y.; Tachikawa, M.; Akanuma, S.; Hosoya, K. Involvement of a novel organic cation transporter in verapamil transport across the inner blood-retinal barrier. Pharm. Res. 2013, 30, 847–856. [Google Scholar] [CrossRef]
- Kubo, Y.; Shimizu, Y.; Kusagawa, Y.; Akanuma, S.; Hosoya, K. Propranolol transport across the inner blood-retinal barrier: Potential involvement of a novel organic cation transporter. J. Pharm. Sci. 2013, 102, 3332–3342. [Google Scholar] [CrossRef] [PubMed]
- Kubo, Y.; Tsuchiyama, A.; Shimizu, Y.; Akanuma, S.; Hosoya, K. Involvement of carrier-mediated transport in the retinal uptake of clonidine at the inner blood-retinal barrier. Mol. Pharm. 2014, 11, 3747–3753. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.H.; Sweet, D.H.; Hu, D.N.; Pritchard, J.B. Characterization of a novel cationic drug transporter in human retinal pigment epithelial cells. J. Pharmacol. Exp. Ther. 2001, 296, 450–457. [Google Scholar]
- Hosoya, K.; Tomi, M.; Ohtsuki, S.; Takanaga, H.; Ueda, M.; Yanai, N.; Obinata, M.; Terasaki, T. Conditionally immortalized retinal capillary endothelial cell lines (TR-iBRB) expressing differentiated endothelial cell functions derived from a transgenic rat. Exp. Eye. Res. 2001, 72, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Nabi, I.R.; Mathews, A.P.; Cohen-Gould, L.; Gundersen, D.; Rodriguez-Boulan, E. Immortalization of polarized rat retinal pigment epithelium. J. Cell. Sci. 1993, 104, 37–49. [Google Scholar] [CrossRef]
- Hosoya, K.; Yamamoto, A.; Akanuma, S.; Tachikawa, M. Lipophilicity and transporter influence on blood-retinal barrier permeability: A comparison with blood-brain barrier permeability. Pharm. Res. 2010, 27, 2715–2724. [Google Scholar] [CrossRef]
- Kubo, Y.; Yamada, M.; Konakawa, S.; Akanuma, S.I.; Hosoya, K.I. Uptake study in lysosome-enriched fraction: Critical involvement of lysosomal trapping in quinacrine uptake but not fluorescence-labeled verapamil transport at blood-retinal barrier. Pharmaceutics 2020, 12, 747. [Google Scholar] [CrossRef] [PubMed]
- Tega, Y.; Tabata, H.; Kurosawa, T.; Kitamura, A.; Itagaki, F.; Oshitari, T.; Deguchi, Y. Structural requirements for uptake of diphenhydramine analogs into hCMEC/D3 cells via the proton-coupled organic cation antiporter. J. Pharm. Sci. 2021, 110, 397–403. [Google Scholar] [CrossRef]
- Lin, H.; Miller, S.S. pHi regulation in frog retinal pigment epithelium: Two apical membrane mechanisms. Am. J. Physiol. 1991, 261, C132–C142. [Google Scholar] [CrossRef]
- Yamaoka, K.; Tanigawara, Y.; Nakagawa, T.; Uno, T. A pharmacokinetic analysis program (multi) for microcomputer. J. Pharmacobiodyn. 1981, 4, 879–885. [Google Scholar] [CrossRef]
- Grabowski, T.; Gumulka, S.W.; Borucka, B.; Raszewski, W. Analysis relationships between pharmacokinetic parameters in silico/in vivo of selected antiviral drugs based on structural analysis. Adv. Clin. Exp. Med. 2008, 17, 285–292. [Google Scholar]
- Grundemann, D.; Gorboulev, V.; Gambaryan, S.; Veyhl, M.; Koepsell, H. Drug excretion mediated by a new prototype of polyspecific transporter. Nature 1994, 372, 549–552. [Google Scholar] [CrossRef] [Green Version]
- Urakami, Y.; Okuda, M.; Masuda, S.; Saito, H.; Inui, K.I. Functional characteristics and membrane localization of rat multispecific organic cation transporters, OCT1 and OCT2, mediating tubular secretion of cationic drugs. J. Pharmacol. Exp. Ther. 1998, 287, 800–805. [Google Scholar] [PubMed]
- Grundemann, D.; Babin-Ebell, J.; Martel, F.; Ording, N.; Schmidt, A.; Schomig, E. Primary structure and functional expression of the apical organic cation transporter from kidney epithelial LLC-PK1 cells. J. Biol. Chem. 1997, 272, 10408–10413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kekuda, R.; Prasad, P.D.; Wu, X.; Wang, H.; Fei, Y.J.; Leibach, F.H.; Ganapathy, V. Cloning and functional characterization of a potential-sensitive, polyspecific organic cation transporter (OCT3) most abundantly expressed in placenta. J. Biol. Chem. 1998, 273, 15971–15979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Kekuda, R.; Huang, W.; Fei, Y.J.; Leibach, F.H.; Chen, J.; Conway, S.J.; Ganapathy, V. Identity of the organic cation transporter OCT3 as the extraneuronal monoamine transporter (uptake2) and evidence for the expression of the transporter in the brain. J. Biol. Chem. 1998, 273, 32776–32786. [Google Scholar] [CrossRef] [Green Version]
- Sloan, J.L.; Mager, S. Cloning and functional expression of a human Na(+) and Cl(-)-dependent neutral and cationic amino acid transporter B(0+). J. Biol. Chem. 1999, 274, 23740–23745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, T.; Hatanaka, T.; Huang, W.; Prasad, P.D.; Leibach, F.H.; Ganapathy, M.E.; Ganapathy, V. Na+- and Cl--coupled active transport of carnitine by the amino acid transporter ATB(0,+) from mouse colon expressed in HRPE cells and Xenopus oocytes. J. Physiol. 2001, 532, 297–304. [Google Scholar] [CrossRef]
- Yabuuchi, H.; Tamai, I.; Nezu, J.; Sakamoto, K.; Oku, A.; Shimane, M.; Sai, Y.; Tsuji, A. Novel membrane transporter OCTN1 mediates multispecific, bidirectional, and pH-dependent transport of organic cations. J. Pharmacol. Exp. Ther. 1999, 289, 768–773. [Google Scholar]
- Wu, X.; George, R.L.; Huang, W.; Wang, H.; Conway, S.J.; Leibach, F.H.; Ganapathy, V. Structural and functional characteristics and tissue distribution pattern of rat OCTN1, an organic cation transporter, cloned from placenta. Biochim. Biophys. Acta. 2000, 1466, 315–327. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Huang, W.; Prasad, P.D.; Seth, P.; Rajan, D.P.; Leibach, F.H.; Chen, J.; Conway, S.J.; Ganapathy, V. Functional characteristics and tissue distribution pattern of organic cation transporter 2 (OCTN2), an organic cation/carnitine transporter. J. Pharmacol. Exp. Ther. 1999, 290, 1482–1492. [Google Scholar] [PubMed]
- Tamai, I.; Ohashi, R.; Nezu, J.; Yabuuchi, H.; Oku, A.; Shimane, M.; Sai, Y.; Tsuji, A. Molecular and functional identification of sodium ion-dependent, high affinity human carnitine transporter OCTN2. J. Biol. Chem. 1998, 273, 20378–20382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otsuka, M.; Matsumoto, T.; Morimoto, R.; Arioka, S.; Omote, H.; Moriyama, Y. A human transporter protein that mediates the final excretion step for toxic organic cations. Proc. Natl. Acad. Sci. USA 2005, 102, 17923–17928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohta, K.Y.; Inoue, K.; Hayashi, Y.; Yuasa, H. Molecular identification and functional characterization of rat multidrug and toxin extrusion type transporter 1 as an organic cation/H+ antiporter in the kidney. Drug. Metab. Dispos. 2006, 34, 1868–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, S.; Kusuhara, H.; Yokochi, M.; Toyoshima, J.; Inoue, K.; Yuasa, H.; Sugiyama, Y. Competitive inhibition of the luminal efflux by multidrug and toxin extrusions, but not basolateral uptake by organic cation transporter 2, is the likely mechanism underlying the pharmacokinetic drug-drug interactions caused by cimetidine in the kidney. J. Pharmacol. Exp. Ther. 2012, 340, 393–403. [Google Scholar] [CrossRef] [Green Version]
- Engel, K.; Wang, J. Interaction of organic cations with a newly identified plasma membrane monoamine transporter. Mol. Pharmacol. 2005, 68, 1397–1407. [Google Scholar] [CrossRef] [Green Version]
- Parsons, C.G.; Panchenko, V.A.; Pinchenko, V.O.; Tsyndrenko, A.Y.; Krishtal, O.A. Comparative patch-clamp studies with freshly dissociated rat hippocampal and striatal neurons on the NMDA receptor antagonistic effects of amantadine and memantine. Eur. J. Neurosci. 1996, 8, 446–454. [Google Scholar] [CrossRef]
- Nishikawa, N.; Nagai, M.; Moritoyo, T.; Yabe, H.; Nomoto, M. Plasma amantadine concentrations in patients with Parkinson’s disease. Parkinsonism. Relat. Disord. 2009, 15, 351–353. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Is Amantadine Accepted as a Substrate? | Is It Expressed in the Retina? | |
|---|---|---|---|
| Slc6a14 | ATB0,+ | Yes [15] | N.D. |
| Slc22a1 | OCT1 | Yes [16] | N.D. |
| Slc22a2 | OCT2 | Yes [17] | N.D. |
| Slc22a3 | OCT3 | N.D. | N.D. |
| Slc22a4 | OCTN1 | N.D. | Yes (mRNA) [18] |
| Slc22a5 | OCTN2 | N.D. | Yes (mRNA) [18] |
| Slc29a4 | PMAT | N.D. | Yes (mRNA) [19] |
| Slc47a1 | MATE1 | Yes [14] | Yes (mRNA) [19] |
| Putative transport systems | Verapamil | N.D. | Yes [20,21,22,23] |
| Clonidine | |||
| Propranolol | |||
| Compound | Concentration | % of Control | |
|---|---|---|---|
| (mM) | TR-iBRB2 | RPE-J | |
| Control | 100 ± 12 | 100 ± 12 | |
| Desipramine | 0.2 | 7.05 ± 0.57 ** | 10.7 ± 5.6 ** |
| Imipramine | 0.2 | 8.37 ± 1.43 ** | 16.9 ± 5.2 ** |
| Propranolol | 0.2 | 8.61 ± 0.75 ** | 15.7 ± 5.9 ** |
| Quinidine | 0.2 | 13.7 ± 1.8 ** | 34.1 ± 3.0 ** |
| Memantine | 0.2 | 16.7 ± 2.2 ** | 123 ± 38 ** |
| 1.0 | N.D. | 27.1 ± 2.6 ** | |
| Pyrilamine | 0.2 | 20.9 ± 1.8 ** | 77.8 ± 38.4 |
| 1.0 | N.D. | 18.2 ± 1.7 ** | |
| Verapamil | 0.2 | 22.1 ± 6.2 ** | 20.5 ± 4.8 ** |
| Amantadine | 0.2 | 34.1 ± 4.1 ** | 89.9 ± 5.6 |
| 1.0 | N.D. | 21.3 ± 3.1 ** | |
| Timolol | 0.2 | 34.6 ± 10.6 ** | 59.1 ± 5.1 |
| Clonidine | 0.2 | 42.1 ± 12.1 ** | 71.6 ± 4.7 |
| 1.0 | N.D. | 30.8 ± 4.0 ** | |
| Pyrimethamine | 0.2 | 44.3 ± 8.8 ** | 88.9 ± 3.3 |
| PAH | 0.2 | 85.8 ± 10.8 | 139 ± 33 |
| Acetazolamide | 0.2 | 90.9 ± 3.8 | N.D. |
| Gluconate | 0.2 | 91.7 ± 21.7 | 86.2 ± 23.4 |
| Cimetidine | 0.2 | 97.6 ± 24.7 | 106 ± 5 |
| Choline | 0.2 | 101 ± 8 | 124 ± 39 |
| MPP+ | 0.2 | 106 ± 31 | 75.5 ± 5.0 |
| 1.0 | N.D. | 91.9 ± 5.6 | |
| Decynium-22 | 0.2 | 106 ± 31 | 86.7 ± 41.9 |
| 0.5 | N.D. | 136 ± 8 | |
| L-Carnitine | 0.2 | 111 ± 35 | 95.1 ± 5.2 |
| 2.5 | 99.5 ± 2.3 | 119 ± 28 | |
| TEA | 0.2 | 117 ± 45 | 87.7 ± 11.8 |
| 1.0 | N.D. | 81.8 ± 10.3 | |
| Serotonin | 0.2 | 126 ± 60 | 158 ± 52 ** |
| L-Glutamic acid | 2.5 | 80.2 ± 10.0 | 84.5 ± 16.4 |
| L-Aspartic acid | 2.5 | 101 ± 23 | 79.5 ± 12.7 |
| Glycine | 2.5 | 102 ± 4 | 116 ± 13 |
| L-Leucine | 2.5 | 112 ± 17 | 113 ± 16 |
| L-Arginine | 2.5 | 127 ± 15 | 150 ± 38 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shinozaki, Y.; Akanuma, S.-i.; Mori, Y.; Kubo, Y.; Hosoya, K.-i. Comprehensive Evidence of Carrier-Mediated Distribution of Amantadine to the Retina across the Blood–Retinal Barrier in Rats. Pharmaceutics 2021, 13, 1339. https://doi.org/10.3390/pharmaceutics13091339
Shinozaki Y, Akanuma S-i, Mori Y, Kubo Y, Hosoya K-i. Comprehensive Evidence of Carrier-Mediated Distribution of Amantadine to the Retina across the Blood–Retinal Barrier in Rats. Pharmaceutics. 2021; 13(9):1339. https://doi.org/10.3390/pharmaceutics13091339
Chicago/Turabian StyleShinozaki, Yusuke, Shin-ichi Akanuma, Yuika Mori, Yoshiyuki Kubo, and Ken-ichi Hosoya. 2021. "Comprehensive Evidence of Carrier-Mediated Distribution of Amantadine to the Retina across the Blood–Retinal Barrier in Rats" Pharmaceutics 13, no. 9: 1339. https://doi.org/10.3390/pharmaceutics13091339