
Consideration of Metabolite Efflux in Radiolabelled Choline Kinetics

 , ,
, ,  , ,
, ,
Abstract
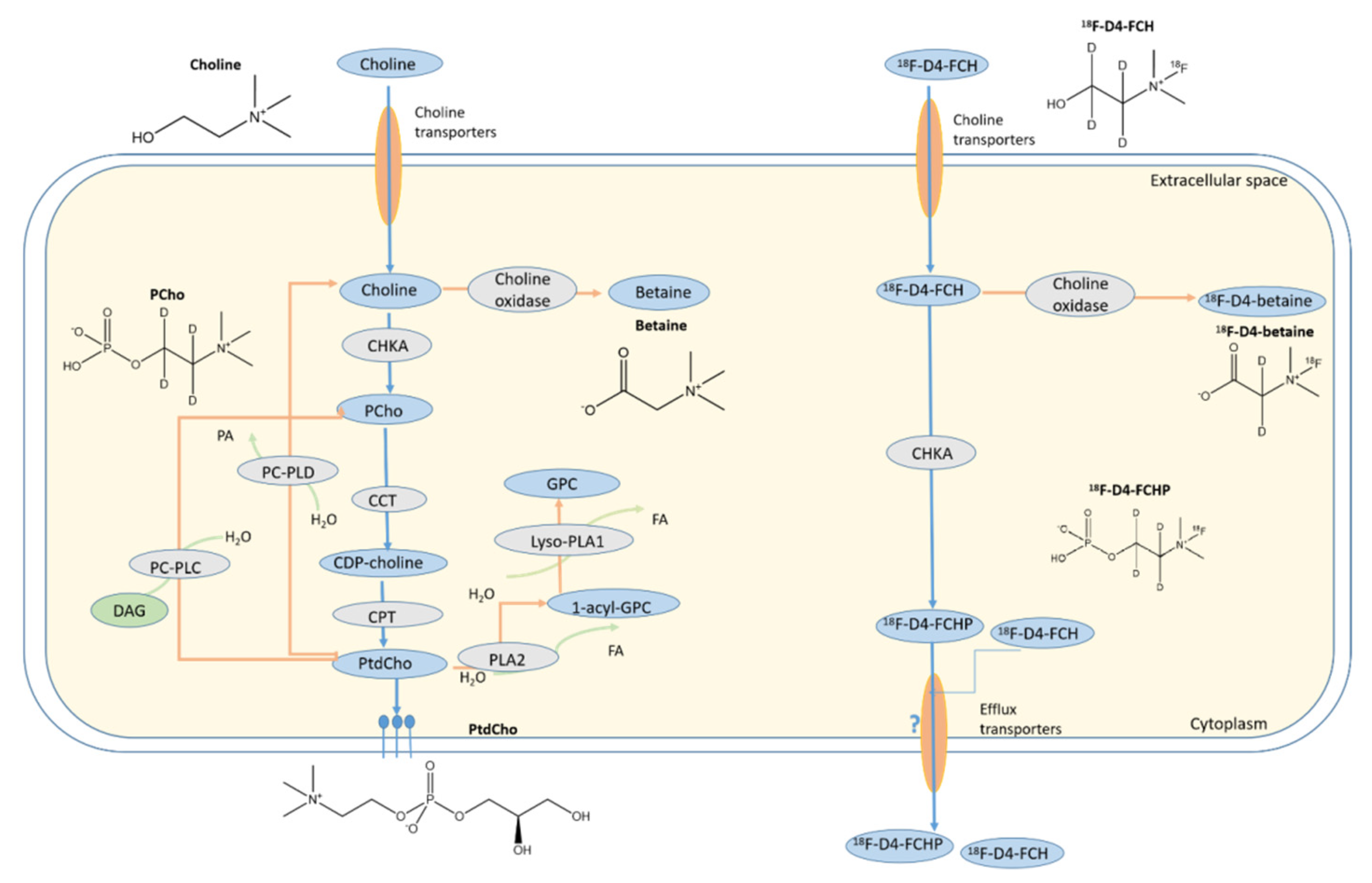
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Drugs
2.2. Hypoxia and CoCl2 Treatment
2.3. Transfection
2.4. pHin Determination
2.5. Immunoblotting
2.6. Immunofluorescence
2.7. Standard Uptake Assay
2.8. Efflux Assay
2.9. Radio-HPLC
2.10. Animal Work
2.11. Patients
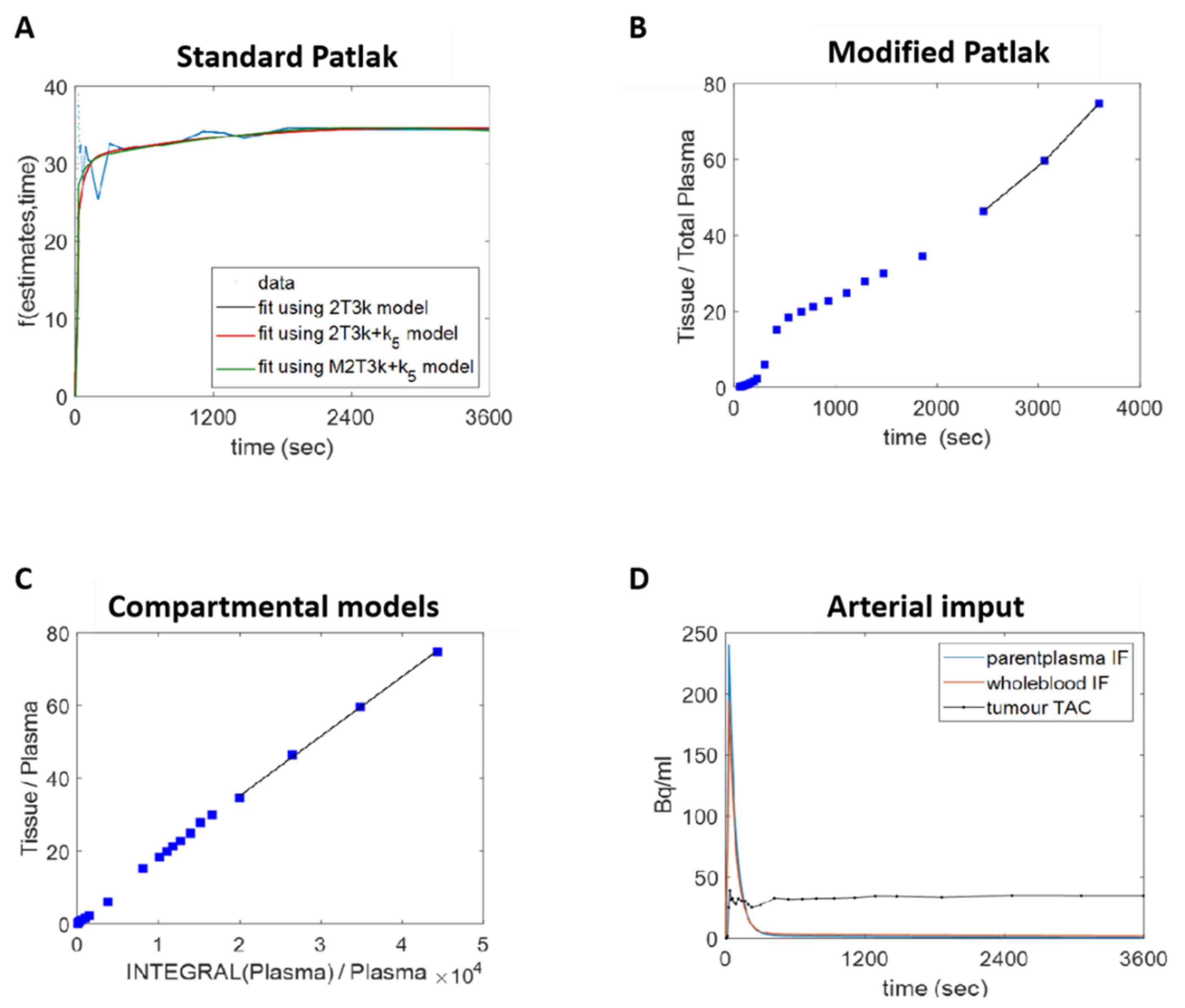
2.12. Graphical Analysis
2.13. Compartmental Analysis
3. Results
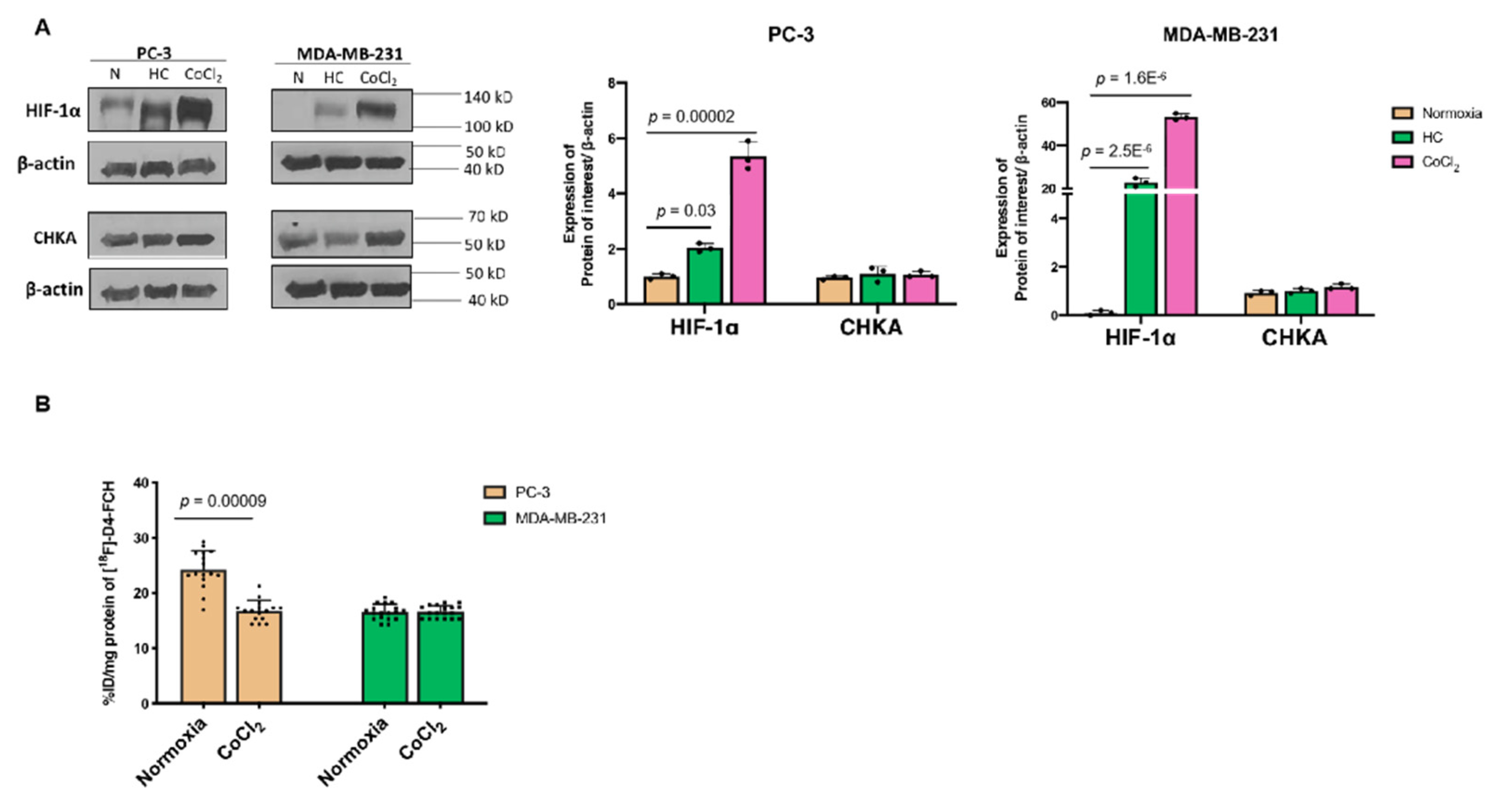
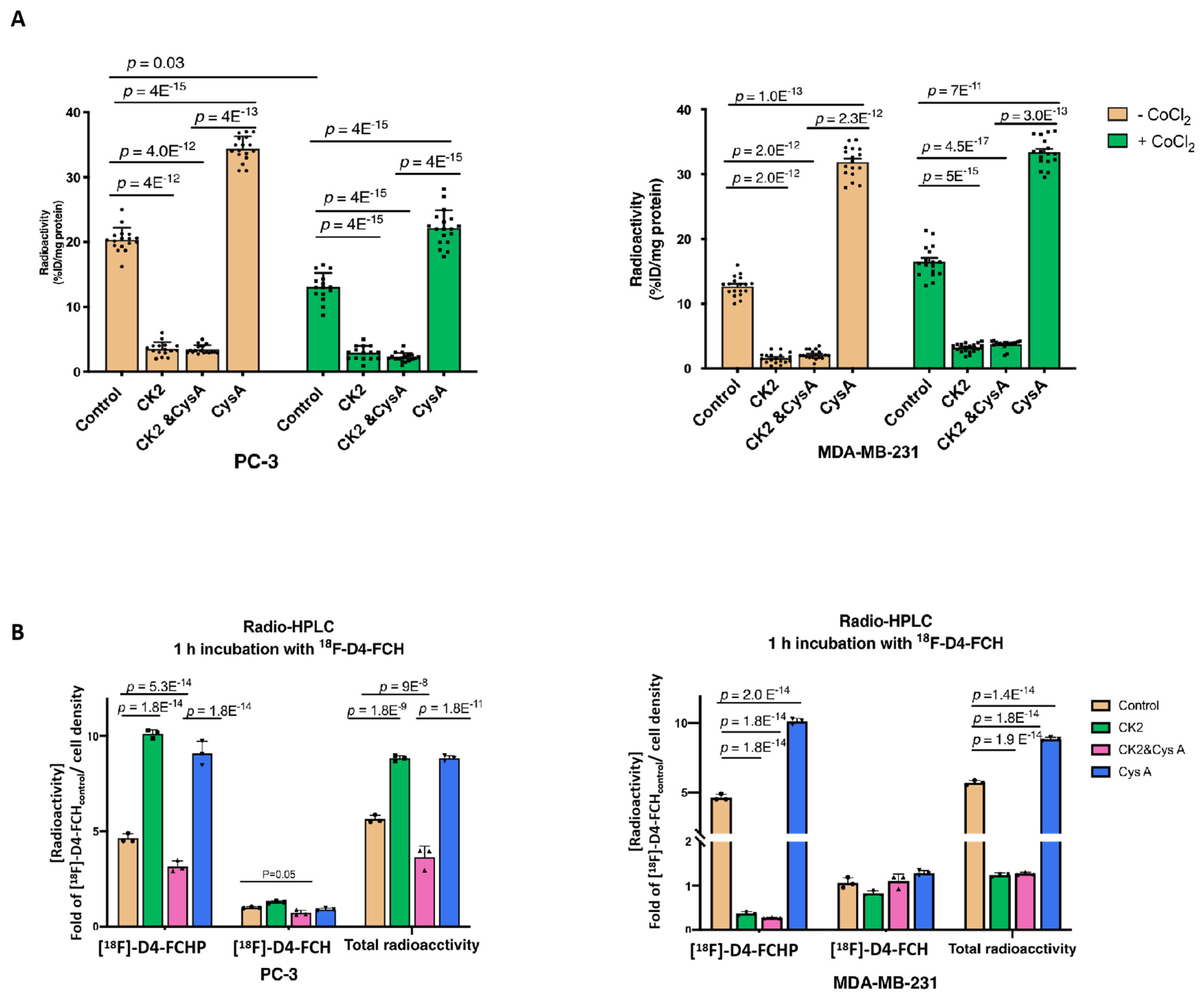
3.1. Hypoxia Does Not Influence Expressions of CHKA and Choline Transporter CTL-1 Proteins
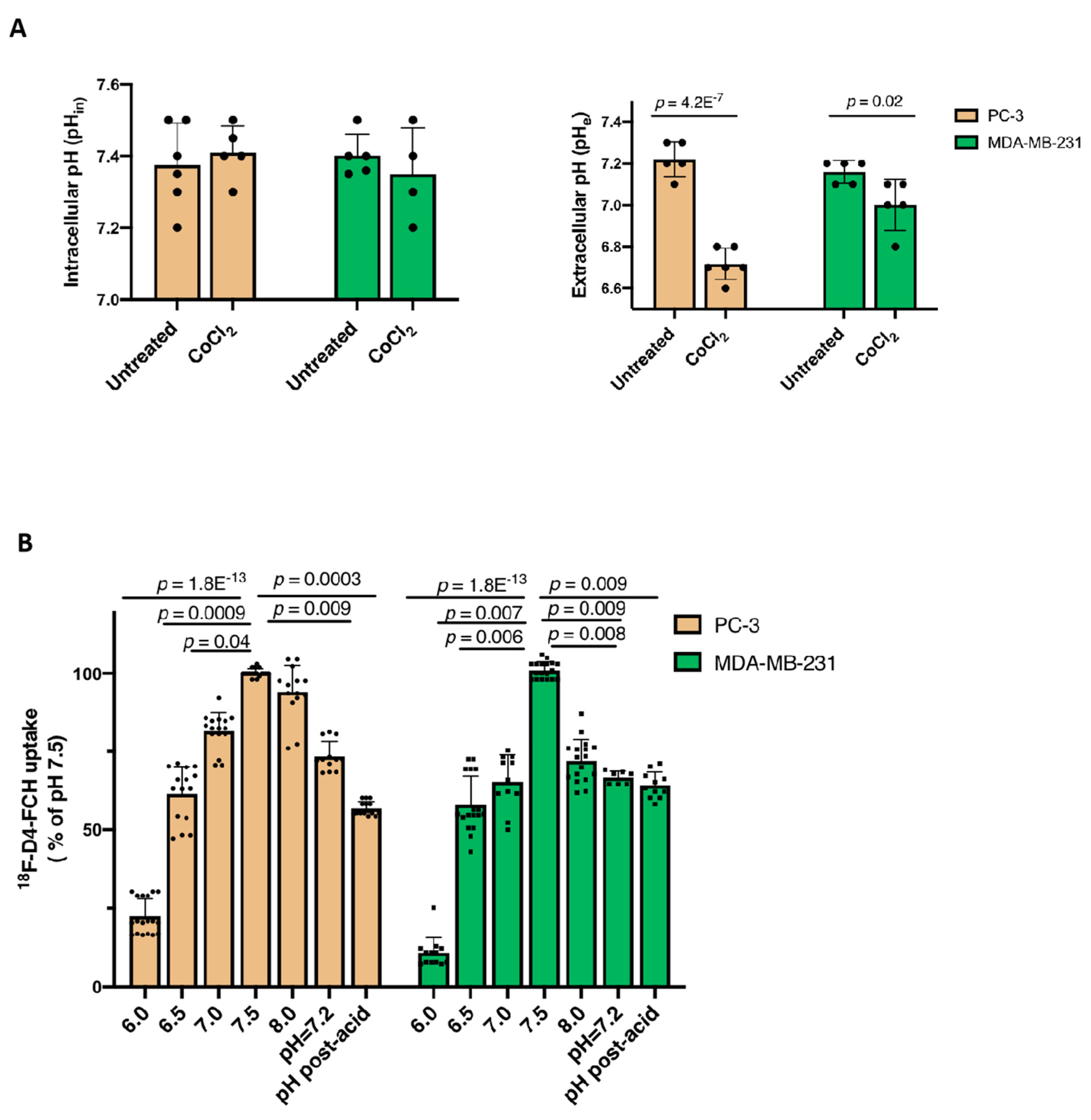
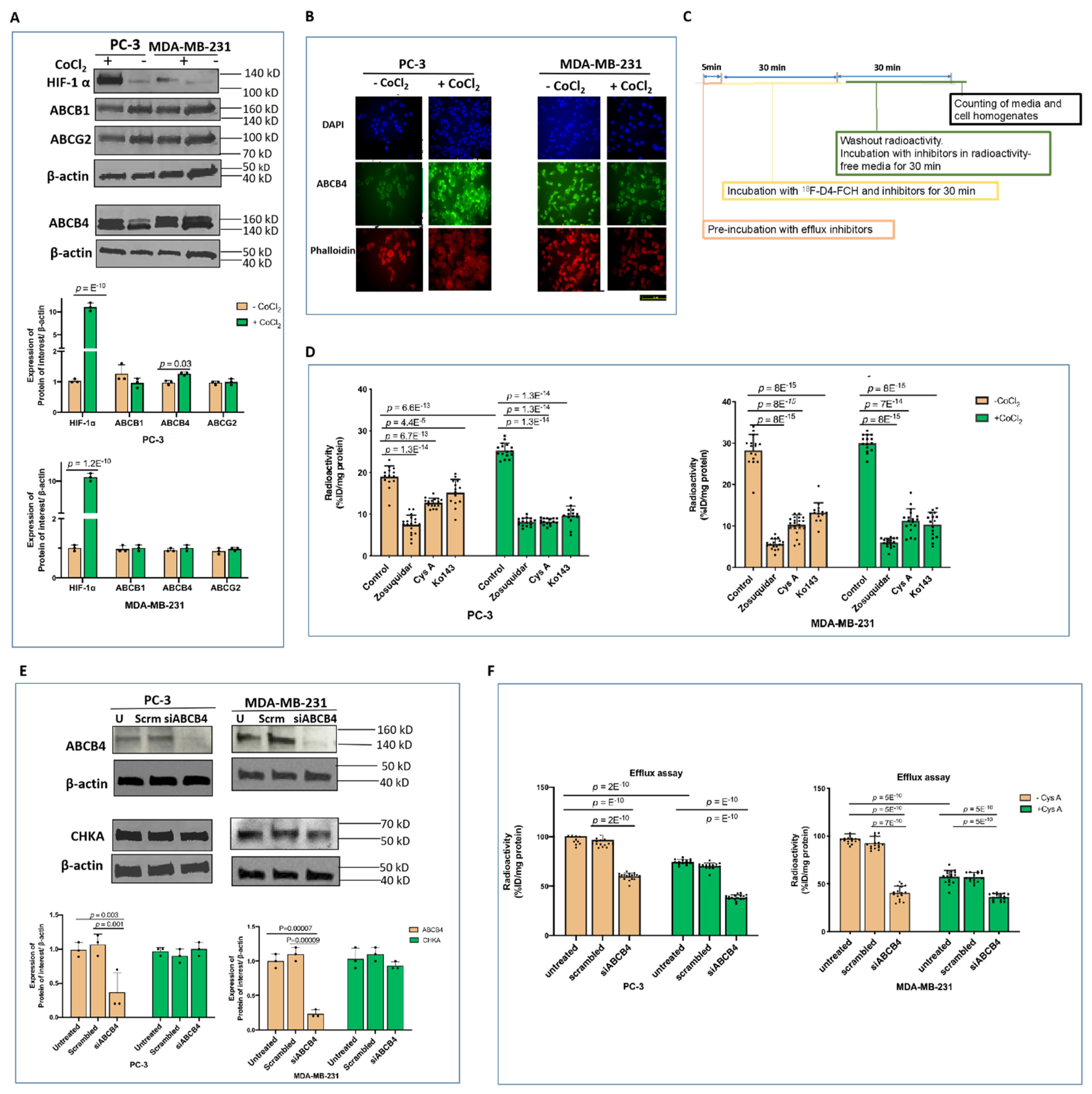
3.2. Other Hypoxia-Related Mechanisms to Explain Differences in Uptake
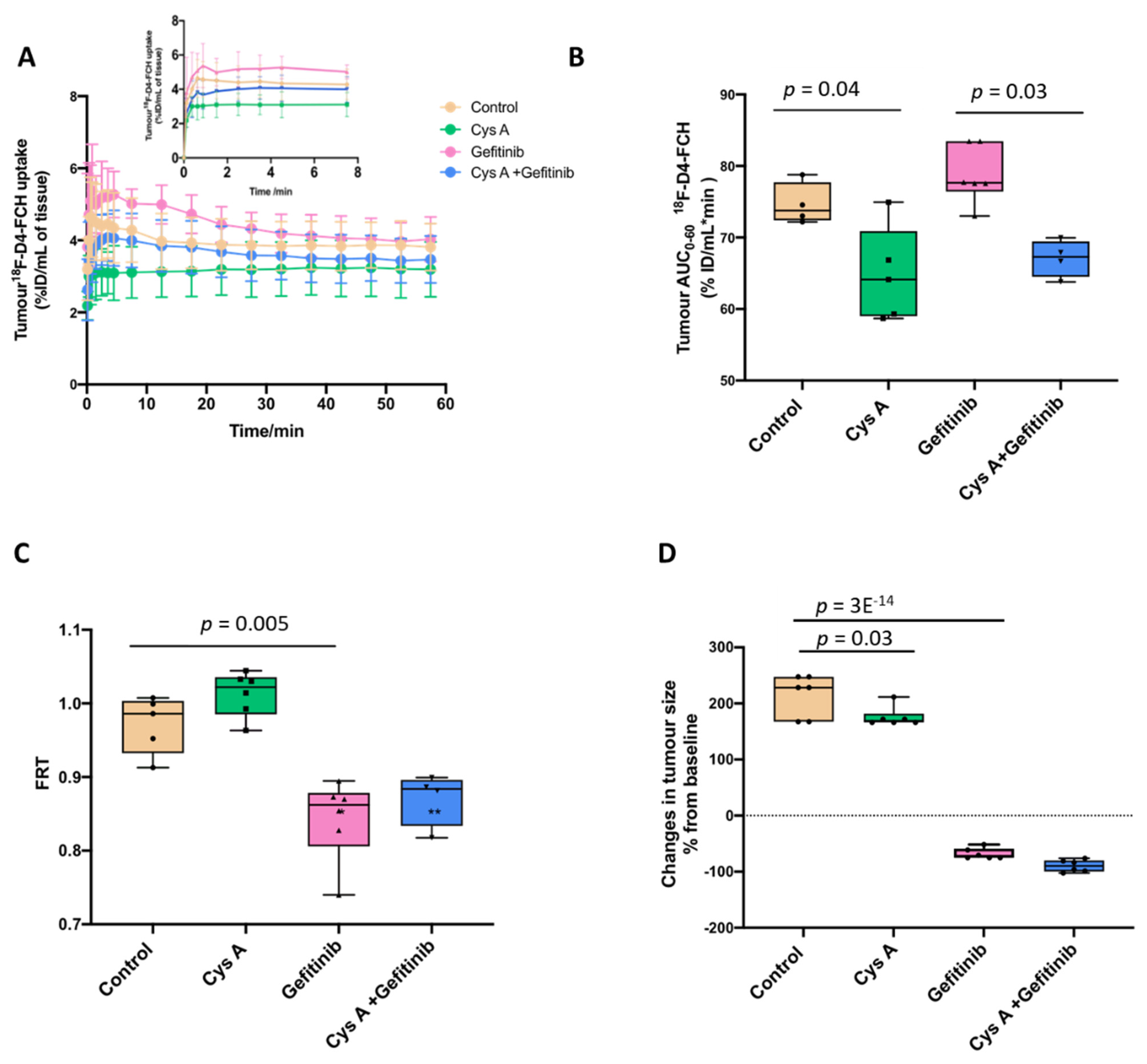
3.3. Dynamic 18F-D4-FCH PET Data Analysis of NSCLC Tumour
3.4. Human Lung Cancer Modelling to Verify Efflux Properties of 18F-D4-FCHP
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kent, C. Regulatory enzymes of phosphatidylcholine biosynthesis: A personal perspective. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2005, 1733, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Glunde, K.; Jie, C.; Bhujwalla, Z.M. Molecalar causes of tile aberrant choline phospholipid metabolism in breast cancer. Cancer Res. 2004, 64, 4270–4276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aboagye, E.O.; Bhujwalla, Z.M. Malignant transformation alters membrane choline phospholipid metabolism of human mammary epithelial cells. Cancer Res. 1999, 59, 80–84. [Google Scholar]
- De Molina, A.R.; Sarmentero-Estrada, J.; Belda-Iniesta, C.; Tarón, M.; de Molina, V.R.; Cejas, P.; Skrzypski, M.; Gallego-Ortega, D.; de Castro, J.; Casado, E.; et al. Expression of choline kinase alpha to predict outcome in patients with early-stage non-small-cell lung cancer. Lancet Oncol. 2007, 8, 889–897. [Google Scholar] [CrossRef]
- Wang, T.; Li, J.; Chen, F.; Zhao, Y.; He, X.; Wan, D.; Gu, J. Choline transporters in human lung adenocarcinoma: Expression and functional implications. Acta Biochim. Biophys. Sin. 2007, 39, 668–674. [Google Scholar] [CrossRef] [Green Version]
- Ackerstaff, E.; Pflug, B.R.; Nelson, J.B.; Bhujwalla, Z.M. Detection of increased choline compounds with proton nuclear magnetic resonance spectroscopy subsequent to malignant transformation of human prostatic epithelial cells. Cancer Res. 2001, 61, 3599–3603. [Google Scholar]
- Contractor, K.; Challapalli, A.; Barwick, T.; Winkler, M.; Hellawell, G.; Hazell, S.; Tomasi, G.; Nahhas, A.; Mapelli, P.; Kenny, L.M.; et al. Use of [11C]Choline PET-CT as a Noninvasive Method for Detecting Pelvic Lymph Node Status from Prostate Cancer and Relationship with Choline Kinase Expression. Clin. Cancer Res. 2011, 17, 7673–7683. [Google Scholar] [CrossRef] [Green Version]
- Michaud, L.; Touijer, K.A.; Mauguen, A.; Zelefsky, M.J.; Morris, M.J.; Lyashschenko, S.K.; Durack, J.C.; Humm, J.L.; Weber, W.A.; Schöder, H. 11C-Choline PET/CT in Recurrent Prostate Cancer: Retrospective Analysis in a Large, U.S. Patient Series. J. Nucl. Med. 2019, 61, 827–833. [Google Scholar] [CrossRef] [PubMed]
- DeGrado, T.R.; Coleman, R.E.; Wang, S.; Baldwin, S.W.; Orr, M.D.; Robertson, C.N.; Polascik, T.J.; Price, D.T. Synthesis and Evaluation of 18 F-labeled Choline as an Oncologic Tracer for Positron Emission Tomography: Initial Findings in Prostate Cancer. Cancer Res. 2000, 61, 110–117. [Google Scholar]
- Kwee, S.A.; Tiirikainen, M.; Sato, M.M.; Acoba, J.D.; Wei, R.; Jia, W.; Le Marchand, L.; Wong, L.L. Transcriptomics Associates Molecular Features with 18F-Fluorocholine PET/CT Imaging Phenotype and Its Potential Relationship to Survival in Hepatocellular Carcinoma. Cancer Res. 2019, 79, 1696–1704. [Google Scholar] [CrossRef] [Green Version]
- Leyton, J.; Smith, G.; Zhao, Y.; Perumal, M.; Nguyen, Q.-D.; Robins, E.; Arstad, E.; Aboagye, E.O. [18F]Fluoromethyl-[1,2-2H4]-Choline: A Novel Radiotracer for Imaging Choline Metabolism in Tumors by Positron Emission Tomography. Cancer Res. 2009, 69, 7721–7728. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.; Zhao, Y.; Leyton, J.; Shan, B.; Nguyen, Q.-D.; Perumal, M.; Turton, D.; Arstad, E.; Luthra, S.K.; Robins, E.; et al. Radiosynthesis and pre-clinical evaluation of [18F]fluoro-[1,2-2H4]choline. Nucl. Med. Biol. 2011, 38, 39–51. [Google Scholar] [CrossRef]
- Witney, T.H.; Alam, I.S.; Turton, D.R.; Smith, G.; Carroll, L.; Brickute, D.; Twyman, F.J.; Nguyen, Q.D.; Tomasi, G.; Awais, R.O.; et al. Evaluation of Deuterated 18F- and 11C-Labeled Choline Analogs for Cancer Detection by Positron Emission Tomography. Clin. Cancer Res. 2012, 18, 1063–1072. [Google Scholar] [CrossRef] [Green Version]
- Dubash, S.; Inglese, M.; Mauri, F.; Kozlowski, K.; Trivedi, P.; Arshad, M.; Challapalli, A.; Barwick, T.; Al-Nahhas, A.; Stanbridge, R.; et al. Spatial heterogeneity of radiolabeled choline positron emission tomography in tumors of patients with non-small cell lung cancer: First-in-patient evaluation of [18F]fluoromethyl-(1,2-2H4)-choline. Theranostics 2020, 10, 8677–8690. [Google Scholar] [CrossRef] [PubMed]
- Contractor, K.B.; Kenny, L.M.; Stebbing, J.; Al-Nahhas, A.; Palmieri, C.; Sinnett, D.; Lewis, J.S.; Hogben, K.; Osman, S.; Shousha, S.; et al. [11C]Choline Positron Emission Tomography in Estrogen Receptor–Positive Breast Cancer. Clin. Cancer Res. 2009, 15, 5503–5510. [Google Scholar] [CrossRef] [Green Version]
- Sarri, E.; Garcia-Dorado, D.; Abellan, A.; Soler-Soler, J. Effects of hypoxia, glucose deprivation and acidosis on phosphatidylcholine synthesis in HL-1 cardiomyocytes. CTP:phosphocholine cytidylyltransferase activity correlates with sarcolemmal disruption. Biochem. J. 2006, 394, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Bansal, A.; Harris, R.A.; DeGrado, T.R. Choline phosphorylation and regulation of transcription of choline kinase α in hypoxia. J. Lipid Res. 2012, 53, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Hara, T.; Bansal, A.; DeGrado, T.R. Effect of hypoxia on the uptake of [methyl-3H]choline, [1-14C] acetate and [18F]FDG in cultured prostate cancer cells. Nucl. Med. Biol. 2006, 33, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Glunde, K.; Shah, T.; Winnard, P.T.; Raman, V.; Takagi, T.; Vesuna, F.; Artemov, D.; Bhujwalla, Z.M. Hypoxia regulates choline kinase expression through hypoxia-inducible factor-1 alpha signaling in a human prostate cancer model. Cancer Res. 2008, 68, 172–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trousil, S.; Carroll, L.; Kalusa, A.; Aberg, O.; Kaliszczak, M.; Aboagye, E. Design of symmetrical and nonsymmetrical N,N-dimethylaminopyridine derivatives as highly potent choline kinase alpha inhibitors. Med. Chem. Comm. 2013, 4, 693–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trousil, S.; Kaliszczak, M.; Schug, Z.; Nguyen, Q.-D.; Tomasi, G.; Favicchio, R.; Brickute, D.; Fortt, R.; Twyman, F.J.; Carroll, L.; et al. The novel choline kinase inhibitor ICL-CCIC-0019 reprograms cellular metabolism and inhibits cancer cell growth. Oncotarget 2016, 7, 37103–37120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du Sert, N.P.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Garner, P.; Karp, N.A.; et al. The arrive guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 40, 1769–1777. [Google Scholar]
- Workman, P.; Aboagye, E.O.; Balkwill, F.; Balmain, A.; Bruder, G.; Chaplin, D.J.; Double, J.A.; Everitt, J.; Farningham, D.A.H.; Glennie, M.J.; et al. Guidelines for the welfare and use of animals in cancer research. Br. J. Cancer 2010, 102, 1555–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patlak, C.S.; Blasberg, R.G. Graphical evaluation of blood-to-brain transfer constants from multiple-time uptake data. Generalizations. J. Cereb. Blood Flow Metab. 1985, 5, 584–590. [Google Scholar] [CrossRef] [Green Version]
- Mankoff, D.A.; Shields, A.F.; Graham, M.M.; Link, J.M.; Krohn, K.A. A graphical analysis method to estimate blood-to-tissue transfer constants for tracers with labeled metabolites. J. Nucl. Med. 1996, 37, 2049–2057. [Google Scholar]
- Akaike, H. A New Look at the Statistical Model Identification. IEEE Trans. Automat. Contr. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Gallego-Ortega, D.; Ramirez de Molina, A.; Ramos, M.A.; Valdes-Mora, F.; Barderas, M.G.; Sarmentero-Estrada, J.; Lacal, J.C. Differential Role of Human Choline Kinase a and b Enzymes in Lipid Metabolism: Implications in Cancer Onset and Treatment. PLoS ONE 2009, 4, e7819. [Google Scholar] [CrossRef] [Green Version]
- Sedlakova, O.; Svastova, E.; Takacova, M.; Kopacek, J.; Pastorek, J.; Pastorekova, S. Carbonic anhydrase IX, a hypoxia-induced catalytic component of the pH regulating machinery in tumors. Front. Physiol. 2014, 4, 400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wartenberg, M.; Ling, F.C.; Müschen, M.; Klein, F.; Acker, H.; Gassmann, M.; Petrat, K.; Pütz, V.; Hescheler, J.; Sauer, H. Regulation of the multidrug resistance transporter P-glycoprotein in multicellular tumor spheroids by hypoxia-inducible factor (HIF-1) and reactive oxygen species. FASEB J. 2003, 17, 1–22. [Google Scholar] [CrossRef]
- Adamska, A.; Falasca, M. ATP-binding cassette transporters in progression and clinical outcome of pancreatic cancer: What is the way forward? World J. Gastroenterol. 2018, 24, 3222–3238. [Google Scholar] [CrossRef]
- Szakács, G.; Paterson, J.K.; Ludwig, J.A.; Booth-Genthe, C.; Gottesman, M.M. Targeting multidrug resistance in cancer. Nat. Rev. Drug Discov. 2006, 5, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, A.; Ierano’, C.; Szakacs, G.; Robey, R.; Bates, S.E. The controversial role of ABC transporters in clinical oncology. Essays Biochem. 2011, 50, 209–232. [Google Scholar] [CrossRef]
- Mazarico, J.M.; Lobo, V.J.S.-A.; Favicchio, R.; Greenhalf, W.; Costello, E.; Pau, E.C.D.S.; Marqués, M.; Lacal, J.C.; Aboagye, E.; Real, F.X. Choline Kinase Alpha (CHKα) as a Therapeutic Target in Pancreatic Ductal Adenocarcinoma: Expression, Predictive Value, and Sensitivity to Inhibitors. Mol. Cancer Ther. 2016, 15, 323–333. [Google Scholar] [CrossRef] [Green Version]
- Thews, O.; Gassner, B.; Kelleher, D.K.; Gekle, M. Activity of drug efflux transporters in tumor cells under hypoxic conditions. In Oxygen Transport to Tissue XXIX. Advances in Experimental Medicine and Biology; Springer: Boston, MA, USA, 2008; pp. 157–164. [Google Scholar]
- Martin, C.M.; Ferdous, A.; Gallardo, T.; Humphries, C.; Sadek, H.; Caprioli, A.; Garcia, J.A.; Szweda, L.I.; Garry, M.G.; Garry, D.J. Hypoxia-Inducible Factor-2α Transactivates Abcg2 and Promotes Cytoprotection in Cardiac Side Population Cells. Circ. Res. 2008, 102, 1075–1081. [Google Scholar] [CrossRef] [Green Version]
- Němcová-Fürstová, V.; Kopperová, D.; Balušíková, K.; Ehrlichová, M.; Brynychová, V.; Václavíková, R.; Petr, D.; Souček, P.; Kovář, J. Characterization of acquired paclitaxel resistance of breast cancer cells and involvement of ABC transporters. Toxicol. Appl. Pharmacol. 2016, 310, 215–228. [Google Scholar] [CrossRef]
- Gautherot, J.; Delautier, D.; Maubert, M.-A.; Aït-Slimane, T.; Bolbach, G.; Delaunay, J.-L.; Durand-Schneider, A.-M.; Firrincieli, D.; Barbu, V.; Chignard, N.; et al. Phosphorylation of ABCB4 impacts its function: Insights from disease-causing mutations. Hepatology 2014, 60, 610–621. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Faussat, A.-M.; Perrot, J.-Y.; Marjanovic, Z.; Cohen, S.; Storme, T.; Morjani, H.; Legrand, O.; Marie, J.-P. Zosuquidar restores drug sensitivity in P-glycoprotein expressing acute myeloid leukemia (AML). BMC Cancer 2008, 8, 51. [Google Scholar] [CrossRef] [Green Version]
- Weidner, L.D.; Zoghbi, S.S.; Lu, S.; Shukla, S.; Ambudkar, S.V.; Pike, V.W.; Mulder, J.; Gottesman, M.M.; Innis, R.B.; Hall, M.D. The Inhibitor Ko143 Is Not Specific for ABCG2. J. Pharmacol. Exp. Ther. 2015, 354, 384–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, S.; Klüver, N.; Burkhardt-Medicke, K.; Pietsch, M.; Schmidt, A.-M.; Wellner, P.; Schirmer, K.; Luckenbach, T. Abcb4 acts as multixenobiotic transporter and active barrier against chemical uptake in zebrafish (Danio rerio) embryos. BMC Biol. 2013, 11, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiedeman, A.M.; Barr, S.I.; Green, T.J.; Xu, Z.; Innis, S.M.; Kitts, D.D. Dietary Choline Intake: Current State of Knowledge Across the Life Cycle. Nutrients 2018, 10, 1513. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Yang, J.; Bai, J.; Ren, J. Reverse of non-small cell lung cancer drug resistance induced by cancer-associated fibroblasts via a paracrine pathway. Cancer Sci. 2018, 109, 944–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyake, T.; Parsons, S.J. Functional interactions between Choline kinase α, epidermal growth factor receptor and c-Src in breast cancer cell proliferation. Oncogene 2012, 31, 1431–1441. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.H.; Huang, C.J.; Yang, C.S.; Chu, Y.C.; Cheng, A.L.; Whang-Peng, J.; Yang, P.C. Gefitinib reverses chemotherapy resistance in gefitinib-insensitive multidrug resistant cancer cells expressing ATP-binding cassette family protein. Cancer Res. 2005, 65, 6943–6949. [Google Scholar] [CrossRef] [Green Version]
- Leggas, M.; Panetta, J.C.; Zhuang, Y.; Schuetz, J.D.; Johnston, B.; Bai, F.; Sorrentino, B.; Zhou, S.; Houghton, P.J.; Stewart, C.F. Gefitinib Modulates the Function of Multiple ATP-Binding Cassette Transporters In vivo. Cancer Res. 2006, 66, 4802–4807. [Google Scholar] [CrossRef] [Green Version]
- Schneider, M.P.; Boesen, E.I.; Pollock, D.M. Contrasting actions of endothelin ETA and ETB receptors in cardiovascular disease. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 731–759. [Google Scholar] [CrossRef] [Green Version]
- Caires, A.; Fernandes, G.; Leme, A.; Castino, B.; Pessoa, E.D.A.; Fernandes, S.; Fonseca, C.; Vattimo, M.; Schor, N.; Borges, F. Endothelin-1 receptor antagonists protect the kidney against the nephrotoxicity induced by cyclosporine-A in normotensive and hypertensive rats. Braz. J. Med. Biol. Res. 2018, 51, e6373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertoldo, A.; Peltoniemi, P.; Oikonen, V.; Knuuti, J.; Nuutila, P.; Cobelli, C. Kinetic modeling of [18F]FDG in skeletal muscle by PET: A four-compartment five-rate-constant model. Am. J. Physiol. Metab. 2001, 281, E524–E536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, R.A. When to use the Bonferroni correction. Ophthalmic Physiol. Opt. 2014, 34, 502–508. [Google Scholar] [CrossRef]
- Ruetz, S.; Gros, P. Phosphatidylcholine translocase: A physiological role for the mdr2 gene. Cell 1994, 77, 1071–1081. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (Mean ± SD) | Standard Patlak | Modified Patlak | ||
|---|---|---|---|---|
| Ki (×10−3) | AIC | Ki (×10−3) | AIC | |
| Primary Tumour | 1.0 ± 0.4 | −231 ± 84 | 29.1 ± 13.7 | −42,259 ± 10,320 |
| Healthy Lung | 0.2 ± 0.2 | −238 ± 52 | 7.4 ± 5.8 | −48,579 ± 10,297 |
| Irreversible Two-Tissue 3k Model | |||||
|---|---|---|---|---|---|
| (Mean ± SD) | K1 (×10−3) | k2 (×10−3) | k3 (×10−3) | Ki (×10−3) | AIC |
| Primary tumour | 150.2 ± 130.6 | 54.4 ± 53.0 | 36.0 ± 19.7 | 49.7 ± 34.5 | −2317 ± 4664 |
| Healthy lung | 51.5 ± 59.3 | 56.7 ± 77.3 | 55.6 ± 76.5 | 17.1 ± 16.6 | −1845 ± 6508 |
| Irreversible Two-Tissue 3k + k5 Model | ||||||
|---|---|---|---|---|---|---|
| (Mean ± SD) | K1 (×10−3) | k2 | k3 | k5 | Ki | AIC |
| Primary tumour | 150.3 ± 130.7 | 48.8 ± 54.4 | 156.0 ± 434.5 | 5.6 ± 7.0 | 68.5 ± 34.1 | −2316 ± 4663 |
| Healthy lung | 51.5 ± 59.4 | 52.9 ± 77.4 | 65.5 ± 57.7 | 4.6 ± 5.6 | 21.2 ± 21.9 | −1842 ± 6509 |
| Modified Irreversible Two-Tissue 6k + k5 Model | ||||||||
|---|---|---|---|---|---|---|---|---|
| (Mean ± SD) | K1 (×10−3) | k2 (×10−3) | k3 (×10−3) | k5 (×10−3) | Ki (×10−3) | K1′ (×10−3) | k2′ (×10−3) | AIC |
| Primary tumour | 153.7 ± 146.9 | 41.7 ± 62.5 | 34.5 ± 32.0 | 13.8 ± 14.5 | 65.5 ± 49.5 | 350.3 ± 1042.5 | 89.1 ± 144.5 | 2750 ± 5052 |
| Healthy lung | 51.0 ± 69.1 | 1629.4 ± 4797.3 | 9784.4 ± 26451.3 | 932.5 ± 2972.4 | 20.8 ± 24.0 | 2124.2 ± 3808.5 | 901.6 ± 1500.3 | 1369 ± 6236 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Inglese, M.; Dubash, S.; Barnes, C.; Brickute, D.; Braga, M.C.; Wang, N.; Beckley, A.; Heinzmann, K.; Allott, L.; et al. Consideration of Metabolite Efflux in Radiolabelled Choline Kinetics. Pharmaceutics 2021, 13, 1246. https://doi.org/10.3390/pharmaceutics13081246
Li Y, Inglese M, Dubash S, Barnes C, Brickute D, Braga MC, Wang N, Beckley A, Heinzmann K, Allott L, et al. Consideration of Metabolite Efflux in Radiolabelled Choline Kinetics. Pharmaceutics. 2021; 13(8):1246. https://doi.org/10.3390/pharmaceutics13081246
Chicago/Turabian StyleLi, Yunqing, Marianna Inglese, Suraiya Dubash, Chris Barnes, Diana Brickute, Marta Costa Braga, Ning Wang, Alice Beckley, Kathrin Heinzmann, Louis Allott, and et al. 2021. "Consideration of Metabolite Efflux in Radiolabelled Choline Kinetics" Pharmaceutics 13, no. 8: 1246. https://doi.org/10.3390/pharmaceutics13081246