
Impact of Th-17 Cytokines on the Regulation of Transporters in Human Placental Explants

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Placental Tissue Culture
2.2. Quantitative Real-Time PCR Analysis
2.3. Immunodetection
2.4. Viability Analysis
2.5. Functional Transport Assay
2.6. Statistical Analysis
3. Results
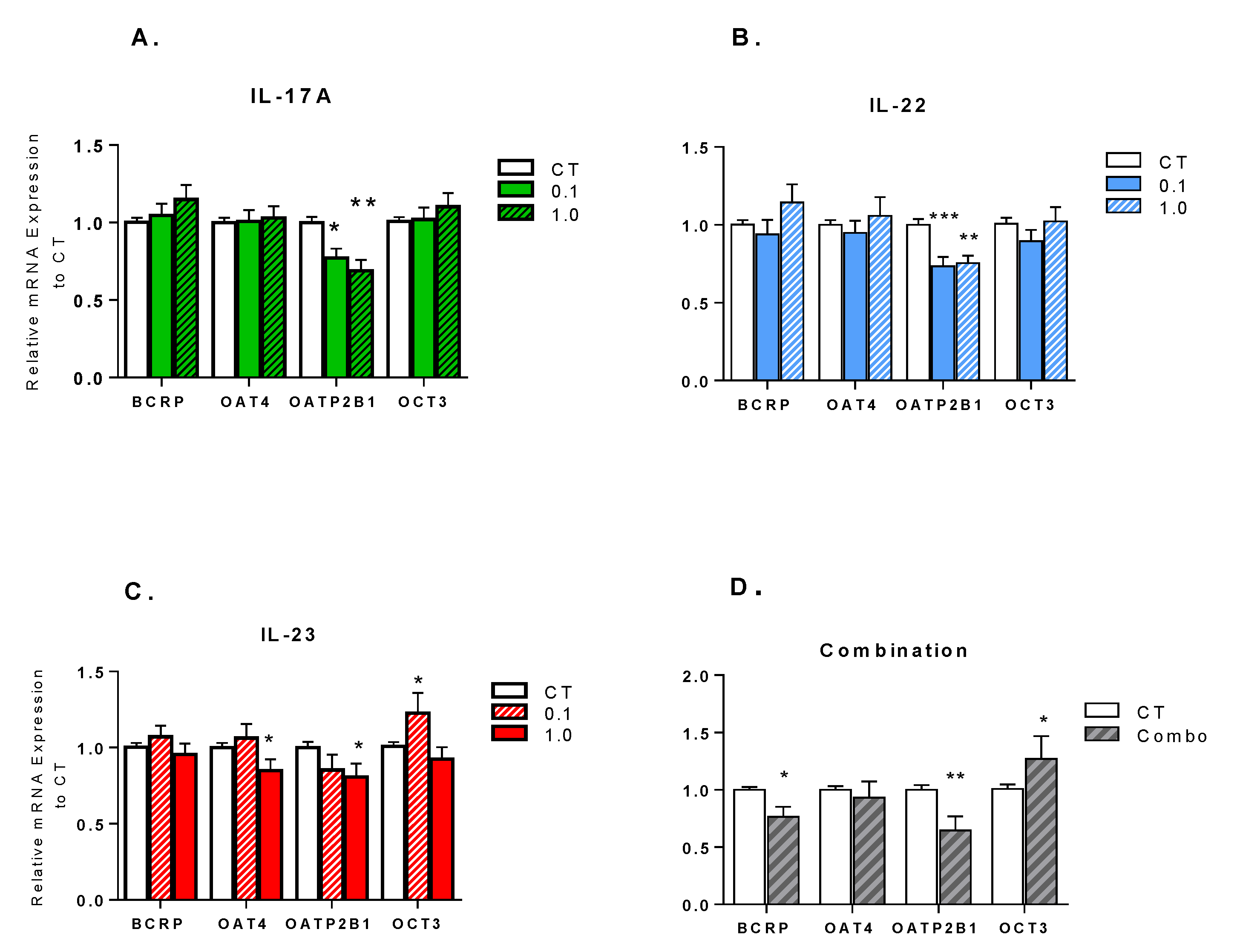
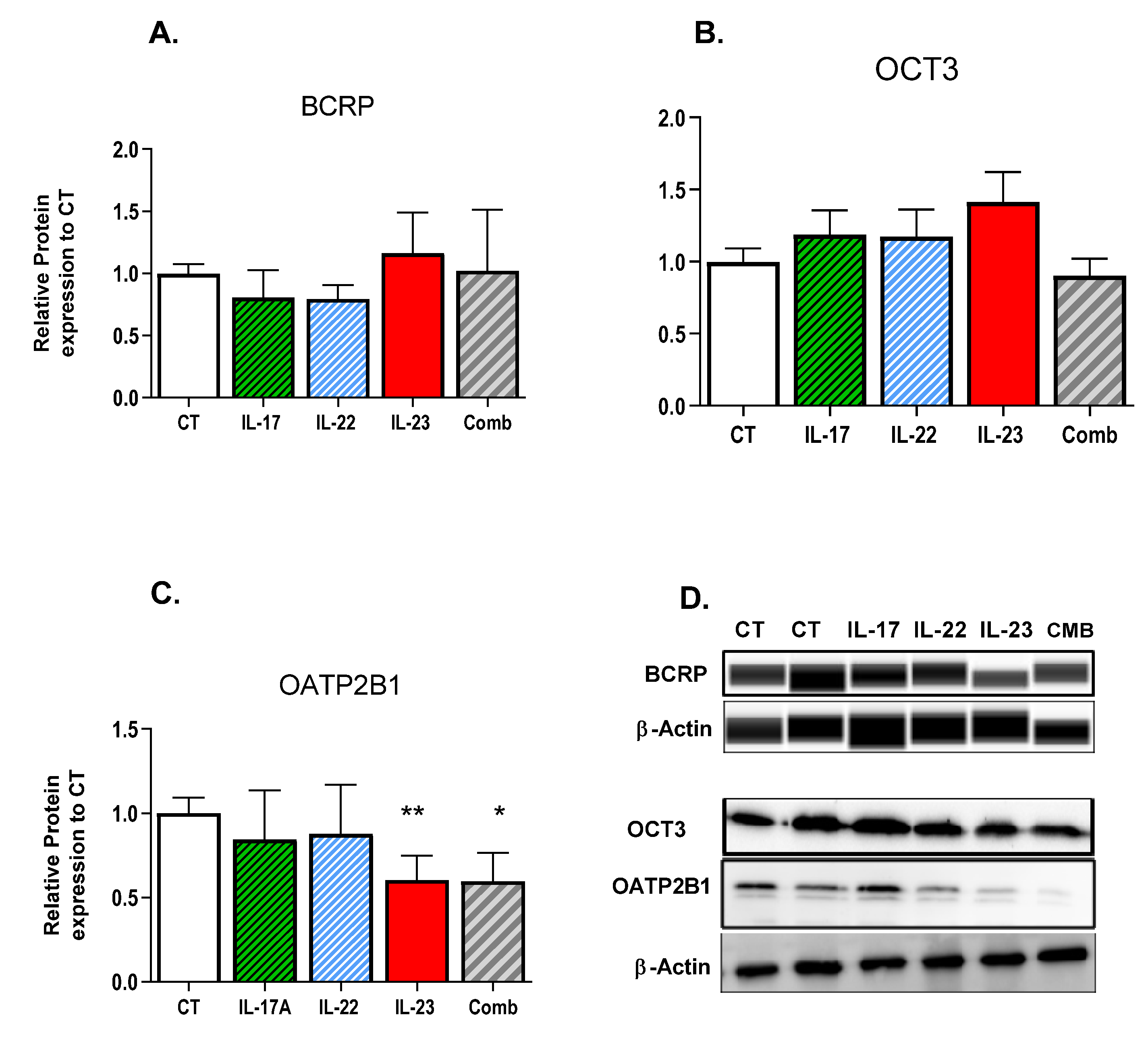
3.1. Effect of Th-17 Cytokines on Transporter Expression
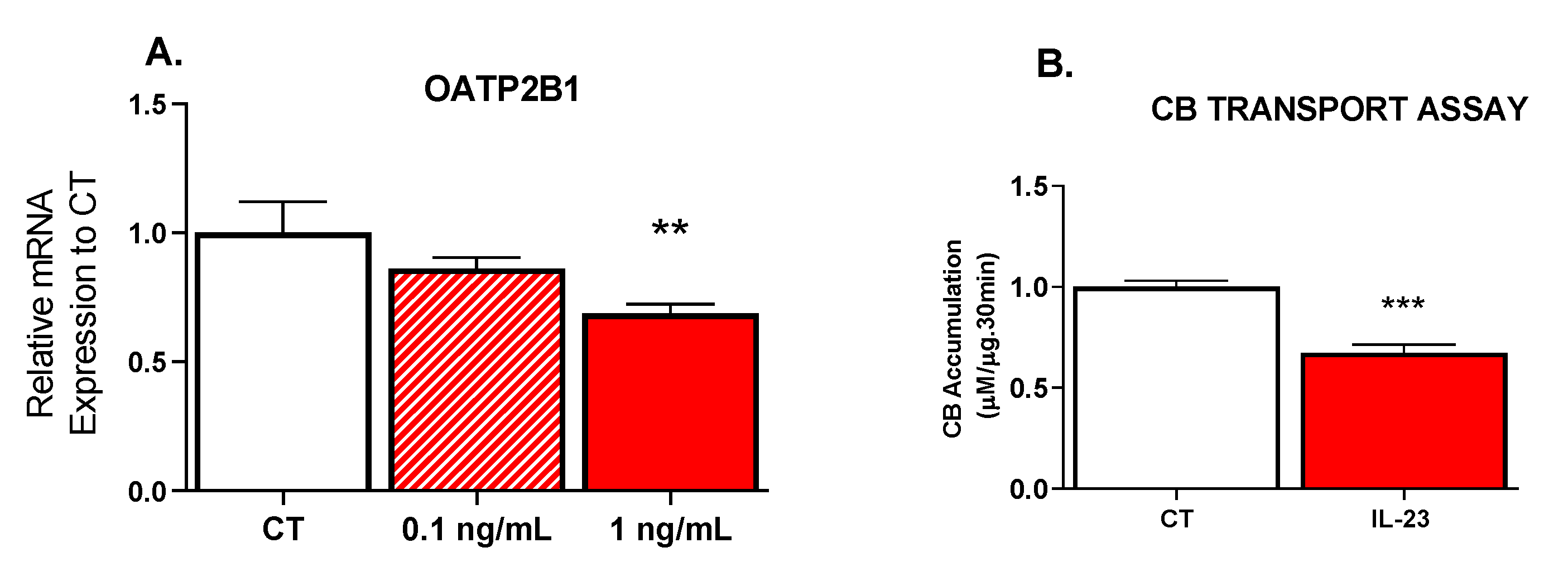
3.2. Effect of IL-23 on OATP Transport Activity
3.3. Effect of Th-17 Cytokines on Tissue Viability and Inflammatory Response
4. Discussion
Strengths and Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Onishi, R.M.; Gaffen, S.L. Interleukin-17 and its target genes: Mechanisms of interleukin-17 function in disease. Immunology 2009, 129, 311–321. [Google Scholar] [CrossRef]
- Tabarkiewicz, J.; Pogoda, K.; Karczmarczyk, A.; Pozarowski, P.; Giannopoulos, K. The Role of IL-17 and Th17 Lymphocytes in Autoimmune Diseases. Arch. Immunol. Ther. Exp. 2015, 63, 435–449. [Google Scholar] [CrossRef] [Green Version]
- Waite, J.C.; Skokos, D. Th17 Response and Inflammatory Autoimmune Diseases. Int. J. Inflam. 2011, 2012, 819467. [Google Scholar] [CrossRef] [Green Version]
- Welsby, I.; Goriely, S. Regulation of Interleukin-23 Expression in Health and Disease. In Regulation of Cytokine Gene Expression in Immunity and Diseases; Ma, X., Ed.; Springer: Dordrecht, The Netherlands, 2016; pp. 167–189. [Google Scholar]
- Leipe, J.; Schramm, M.A.; Grunke, M.; Baeuerle, M.; Dechant, C.; Nigg, A.P.; Witt, M.N.; Vielhauer, V.; Reindl, C.S.; Schulze-Koops, H.; et al. Interleukin 22 serum levels are associated with radiographic progression in rheumatoid arthritis. Ann. Rheum. Dis. 2011, 70, 1453. [Google Scholar] [CrossRef] [Green Version]
- Al-Saadany, H.; Hussein, M.S.; Gaber, R.A.; Zaytoun, H.A. Th-17 cells and serum IL-17 in rheumatoid arthritis patients: Correlation with disease activity and severity. Egypt. Rheumatol. 2016, 38, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Chen, S.; Miao, L.; Liu, Z.; Li, W.; Zhao, Z.; Sun, X.; Jiang, G.; Cheng, Q. Serum levels of interleukin (IL)-18, IL-23 and IL-17 in Chinese patients with multiple sclerosis. J. Neuroimmunol. 2012, 243, 56–60. [Google Scholar] [CrossRef]
- Jia, H.P.; Look, D.C.; Shi, L.; Hickey, M.; Pewe, L.; Netland, J.; Farzan, M.; Wohlford-Lenane, C.; Perlman, S.; McCray, P.B., Jr. ACE2 receptor expression and severe acute respiratory syndrome coronavirus infection depend on differentiation of human airway epithelia. J. Virol. 2005, 79, 14614–14621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cressman, A.M.; Petrovic, V.; Piquette-Miller, M. Inflammation-mediated changes in drug transporter expression/activity: Implications for therapeutic drug response. Expert Rev. Clin. Pharmacol. 2012, 5, 69–89. [Google Scholar] [CrossRef] [PubMed]
- Koren, G.; Pariente, G. Pregnancy-Associated Changes in Pharmacokinetics and their Clinical Implications. Pharm. Res. 2018, 35, 61, 1–7. [Google Scholar] [CrossRef]
- Dos Reis, C.R.P.; Cardoso, G.; Carvalho, C.; Nogueira, I.; Borges, A.; Serrano, F. Prediction of Adverse Pregnancy Outcomes in Women with Systemic Lupus Erythematosus. Clin. Rev. Allergy Immunol. 2020, 59, 287–294. [Google Scholar] [CrossRef]
- Mirdamadi, K.; Salinas, T.; Vali, R.; Papadimitropoulos, M.; Piquette-Miller, M. Meta-Analysis of Pregnancy Outcomes after Exposure to TNF-alpha Inhibitors During Pregnancy for the Treatment of Arthritic Diseases. J. Popul. Ther. Clin. Pharmacol. 2018, 25, e53–e56. [Google Scholar] [CrossRef]
- Komaki, F.; Komaki, Y.; Micic, D.; Ido, A.; Sakuraba, A. Outcome of pregnancy and neonatal complications with anti-tumor necrosis factor-alpha use in females with immune mediated diseases; a systematic review and meta-analysis. J. Autoimmun. 2016, 76, 38–52. [Google Scholar] [CrossRef]
- Liu, L.; Liu, X. Contributions of Drug Transporters to Blood-Placental Barrier. Adv. Exp. Med. Biol. 2019, 1141, 505–548. [Google Scholar]
- Ghoneim, R.H.; Kojovic, D.; Piquette-Miller, M. Impact of endotoxin on the expression of drug transporters in the placenta of HIV-1 transgenic rats. Eur. J. Pharm. Sci. 2017, 102, 94–102. [Google Scholar] [CrossRef]
- Petrovic, V.; Wang, J.H.; Piquette-Miller, M. Effect of endotoxin on the expression of placental drug transporters and glyburide disposition in pregnant rats. Drug Metab. Dispos. 2008, 36, 1944–1950. [Google Scholar] [CrossRef]
- Evseenko, D.A.; Paxton, J.W.; Keelan, J.A. ABC drug transporter expression and functional activity in trophoblast-like cell lines and differentiating primary trophoblast. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R1357–R1365. [Google Scholar] [CrossRef]
- Daud, A.N.A.; Bergman, J.E.H.; Oktora, M.P.; Kerstjens-Frederikse, W.; Groen, H.; Bos, J.H.; Hak, E.; Wilffert, B. Maternal use of drug substrates of placental transporters and the effect of transporter-mediated drug interactions on the risk of congenital anomalies. PLoS ONE 2017, 12, e0173530. [Google Scholar] [CrossRef]
- Anoshchenko, O.; Prasad, B.; Neradugomma, N.K.; Wang, J.; Mao, Q.; Unadkat, J.D. Gestational Age-Dependent Abundance of Human Placental Transporters as Determined by Quantitative Targeted Proteomics. Drug Metab. Dispos. 2020, 48, 735–741. [Google Scholar] [CrossRef]
- Burton, G.J.; Sebire, N.J.; Myatt, L.; Tannetta, D.; Wang, Y.L.; Sadovsky, Y.; Staff, A.C.; Redman, C.W. Optimising sample collection for placental research. Placenta 2014, 35, 9–22. [Google Scholar] [CrossRef]
- Kojovic, D.; Workewych, N.V.; Piquette-Miller, M. Role of Elevated SFLT-1 on the Regulation of Placental Transporters in Women with Pre-Eclampsia. Clin. Transl. Sci. 2020, 13, 580–588. [Google Scholar] [CrossRef]
- Petrovic, V.; Kojovic, D.; Cressman, A.; Piquette-Miller, M. Maternal bacterial infections impact expression of drug transporters in human placenta. Int. Immunopharmacol. 2015, 26, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Marino, G.I.; Castro-Parodi, M.; Dietrich, V.; Damiano, A.E. High levels of human chorionic gonadotropin (hCG) correlate with increased aquaporin-9 (AQP9) expression in explants from human preeclamptic placenta. Reprod. Sci. 2010, 17, 444–453. [Google Scholar] [CrossRef]
- Patik, I.; Szekely, V.; Nemet, O.; Szepesi, A.; Kucsma, N.; Varady, G.; Szakacs, G.; Bakos, E.; Ozvegy-Laczka, C. Identification of novel cell-impermeant fluorescent substrates for testing the function and drug interaction of Organic Anion-Transporting Polypeptides, OATP1B1/1B3 and 2B1. Sci. Rep. 2018, 8, 2630–2642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Windt, T.; Toth, S.; Patik, I.; Sessler, J.; Kucsma, N.; Szepesi, A.; Zdrazil, B.; Ozvegy-Laczka, C.; Szakacs, G. Identification of anticancer OATP2B1 substrates by an in vitro triple-fluorescence-based cytotoxicity screen. Arch. Toxicol. 2019, 93, 953–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riss, T.L.; Moravec, R.A.; Niles, A.L.; Duellman, S.; Benink, H.A.; Worzella, T.J.; Minor, L. Cell Viability Assays. In Assay Guidance Manual; Markossian, S., Sittampalam, G.S., Grossman, A., Brimacombe, K., Arkin, M., Auld, D., Austin, C.P., Baell, J., Caaveiro, J.M.M., Chung, T.D.Y., et al., Eds.; Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda, MD, USA, 2004. [Google Scholar]
- Lye, P.; Bloise, E.; Javam, M.; Gibb, W.; Lye, S.J.; Matthews, S.G. Impact of Bacterial and Viral Challenge on Multidrug Resistance in First- and Third-Trimester Human Placenta. Am. J. Pathol. 2015, 185, 1666–1675. [Google Scholar] [CrossRef] [PubMed]
- Logiodice, F.; Lombardelli, L.; Kullolli, O.; Haller, H.; Maggi, E.; Rukavina, D.; Piccinni, M. Decidual Interleukin-22-Producing CD4+ T Cells, which also produce IL-4, Are Involved in the Success of Pregnancy. Int. J. Mol. Sci. 2019, 20, 428. [Google Scholar] [CrossRef] [Green Version]
- Kaminski, V.L.; Ellwanger, J.H.; Matte, M.C.C.; Savaris, R.F.; Vianna, P.; Chies, J.A.B. IL-17 blood levels increase in healthy pregnancy but not in spontaneous abortion. Mol. Biol. Rep. 2018, 45, 1565–1568. [Google Scholar] [CrossRef]
- Torricelli, M.; Bellisai, F.; Novembri, R.; Galeazzi, L.R.; Iuliano, A.; Voltolini, C.; Spreafico, A.; Galeazzi, M.; Petraglia, F. High Levels of Maternal Serum IL-17 and Activin A in Pregnant Women Affected by SLE. Am. J. Reprod. Immunol. 2011, 66, 84–89. [Google Scholar] [CrossRef]
- Wong, C.K.; Lit, L.C.W.; Tam, L.S.; Li, E.K.M.; Wong, P.T.Y.; Lam, C.W.K. Hyperproduction of IL-23 and IL-17 in patients with SLE: Implications for Th17-mediated inflammation in auto-immunity. Clin. Immunol. 2008, 127, 385–393. [Google Scholar] [CrossRef]
- Kojovic, D.; Ghoneim, R.H.; Serghides, L.; Piquette-Miller, M. Role of HIV and Antiretroviral Therapy on the Expression of Placental Transporters in Women with HIV. AAPS J. 2020, 22, 1–12. [Google Scholar] [CrossRef]
- Lofthouse, E.M.; Cleal, J.K.; O’Kelly, I.M.; Sengers, B.G.; Lewis, R.M. Estrone sulphate uptake by the microvillous membrane of placental syncytiotrophoblast is coupled to glutamate efflux. Biochem. Biophys. Res. Commun. 2018, 506, 237–242. [Google Scholar] [CrossRef]
- Grube, M.; Reuther, S.; Zu Schwabedissen, H.M.; Kock, K.; Draber, K.; Ritter, C.A.; Fusch, C.; Jedlitschky, G.; Kroemer, H.K. Organic Anion Transporting Polypeptide 2B1 and Breast Cancer Resistance Protein Interact in the Transepithelial Transport of Steroid Sulfates in Human Placenta. Drug Metab. Dispos. 2007, 35, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doria, A.; Cutolo, M.; Ghirardello, A.; Zampieri, S.; Vescovi, F.; Sulli, A.; Giusti, M.; Piccoli, A.; Grella, P.; Gambari, P.F. Steroid Hormones and Disease Activity during Pregnancy in SLE. Arthritis Rheum. 2002, 47, 202–209. [Google Scholar] [CrossRef] [Green Version]
- Kaludjerovic, J.; Ward, W.E. The Interplay between Estrogen and Fetal Adrenal Cortex. J. Nutr. Metab. 2012, 2012, 837901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eghbal-Fard, S.; Yousefi, M.; Heydarlou, H.; Ahmadi, M.; Taghavi, S.; Movasaghpour, A.; Jadidi-Niaragh, F.; Yousefi, B.; Dolati, S.; Hojjat-Farsangi, M.; et al. The imbalance of Th17/Treg axis involved in the pathogenesis of preeclampsia. J. Cell. Physiol. 2019, 234, 5106–5116. [Google Scholar] [CrossRef]
- Tomi, M.; Eguchi, H.; Ozaki, M.; Tawara, T.; Nishimura, S.; Higuchi, K.; Maruyama, T.; Nishimura, T.; Nakashima, E. Role of OAT4 in Uptake of Estriol Precursor 16-Hydroxydehydroepiandrosterone Sulfate Into Human Placental Syncytiotrophoblasts From Fetus. Endocrinology 2015, 156, 2704–2712. [Google Scholar] [CrossRef] [Green Version]
- Kliman, H.J.; Quaratella, S.B.; Setaro, A.C.; Siegman, E.C.; Subha, Z.T.; Tal, R.; Milano, K.M.; Steck, T.L. Pathway of Maternal Serotonin to the Human Embryo and Fetus. Endocrinology 2018, 159, 1609–1629. [Google Scholar] [CrossRef]
- Sata, R.; Ohtani, H.; Tsujimoto, M.; Murakami, H.; Koyabu, N.; Nakamura, T.; Uchiumi, T.; Kuwano, M.; Nagata, H.; Tsukimori, K.; et al. Functional analysis of organic cation transporter 3 expressed in human placenta. J. Pharmacol. Exp. Ther. 2005, 315, 888–895. [Google Scholar] [CrossRef] [Green Version]
- Ahmadimoghaddam, D.; Cerveny, L.; Ceckova, M.; Neumanova, Z.; Staud, F.; Dolezelova, E.; Zemankova, L.; Nachtigal, P.; Kacerovsk, M.; Micuda, S. Organic Cation Transporter 3 and Multidrug and Toxin Extrusion 1 Transporter in the Placenta and Fetal Tissues: Expression Profile and Fetus Protective Role at Different Stages of Gestation. Biol. Reprod. 2013, 88, 1–10. [Google Scholar] [CrossRef]
- Lee, N.; Hebert, M.F.; Wagner, D.J.; Easterling, T.R.; Liang, C.J.; Rice, K.; Wang, J. Organic Cation Transporter 3 Facilitates Fetal Exposure to Metformin during Pregnancy. Mol. Pharmacol. 2018, 94, 1125–1131. [Google Scholar] [CrossRef]
- Martinez, G.J.; Nurieva, R.I.; Yang, X.O.; Dong, C. Regulation and function of proinflammatory TH17 cells. Ann. N. Y. Acad. Sci. 2008, 1143, 188–211. [Google Scholar] [CrossRef] [Green Version]
- Valeri, M.; Raffatellu, M. Cytokines IL-17 and IL-22 in the host response to infection. Pathog. Dis. 2016, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, J.A.; Hueber, A.J.; Wilson, S.; Galm, M.; Baum, W.; Kitson, C.; Auer, J.; Lorenz, S.H.; Moelleken, J.; Bader, M.; et al. Combined inhibition of tumor necrosis factor alpha and IL-17 as a therapeutic opportunity in rheumatoid arthritis: Development and characterization of a novel bispecific antibody. Arthritis Rheumatol. 2015, 67, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Evseenko, D.A.; Paxton, J.W.; Keelan, J.A. Independent regulation of apical and basolateral drug transporter expression and function in placental trophoblasts by cytokines, steroids, and growth factors. Drug Metab. Dispos. 2007, 35, 595–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
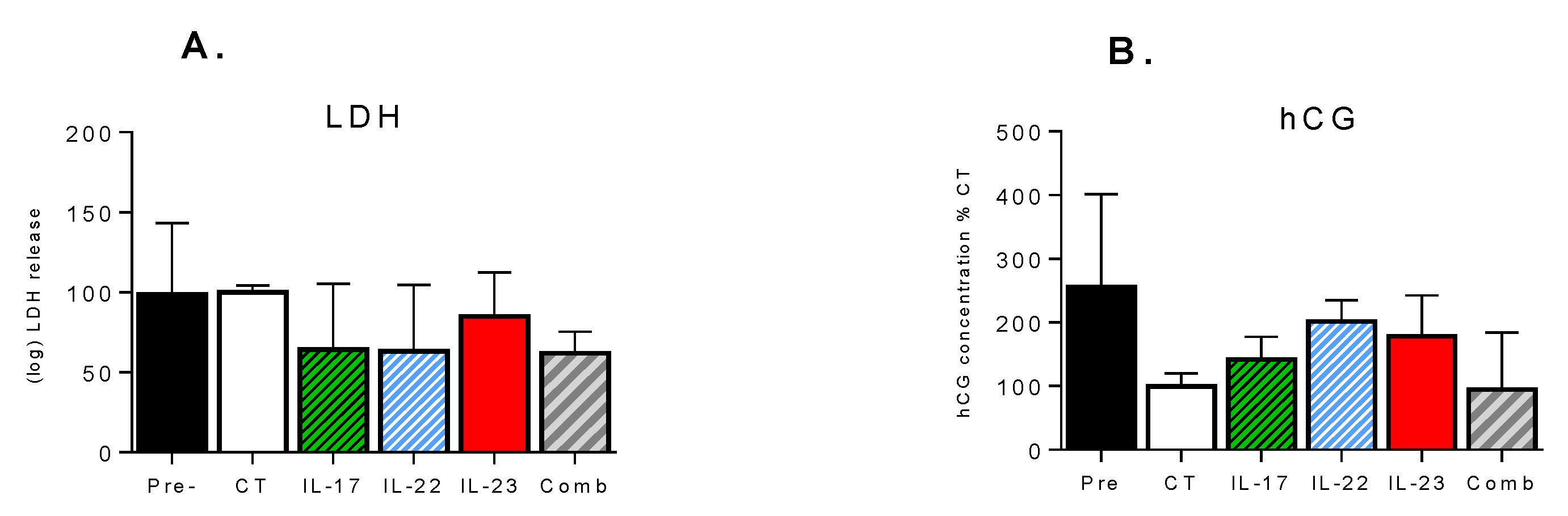{kind=link}
| Gene | Sequence |
|---|---|
| ABCG2 (BCRP) | Fw: 5′GGCCTTGCGATACTTTGAATC3′ Rv: 5′GAATCTCCATTAATGATGTCCA3′ |
| SLC22A11 (OAT4) | Fw: 5′ GGACCTGGAGAGCCAGAAATC3′ Rv: 5′GAGCGAGGTACTTTCCACAGTGA3′ |
| SLCO2B1 (OATP2B1) | Fw: 5′ACCCTCCTTCATGCTCATCCT3′ Rv: 5′ATGCCCACAGCCAAAGTCTT3′ |
| SLC22A3 (OCT3) | Fw: 5′TGCCAGAGACAGTGGATGATG3′ Rv: 5′TTTTCTTATTCCTGCCACATTTACAG3′ |
| Cyclophilin A (PPIA) | Fw: 5′GCCCGTAGTGCTTCAGCTT3′ Rv: 5′GGAGATGGCACAGGAGGAA3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirdamadi, K.; Kwok, J.; Nevo, O.; Berger, H.; Piquette-Miller, M. Impact of Th-17 Cytokines on the Regulation of Transporters in Human Placental Explants. Pharmaceutics 2021, 13, 881. https://doi.org/10.3390/pharmaceutics13060881
Mirdamadi K, Kwok J, Nevo O, Berger H, Piquette-Miller M. Impact of Th-17 Cytokines on the Regulation of Transporters in Human Placental Explants. Pharmaceutics. 2021; 13(6):881. https://doi.org/10.3390/pharmaceutics13060881
Chicago/Turabian StyleMirdamadi, Kamelia, Jacinda Kwok, Ori Nevo, Howard Berger, and Micheline Piquette-Miller. 2021. "Impact of Th-17 Cytokines on the Regulation of Transporters in Human Placental Explants" Pharmaceutics 13, no. 6: 881. https://doi.org/10.3390/pharmaceutics13060881