
Seaweed Sulfated Polysaccharides against Respiratory Viral Infections


Abstract
:1. Introduction
2. Symptoms of Respiratory Viral Infections and Approved Therapies
2.1. Symptomatic Treatment
2.2. Antiviral Medication
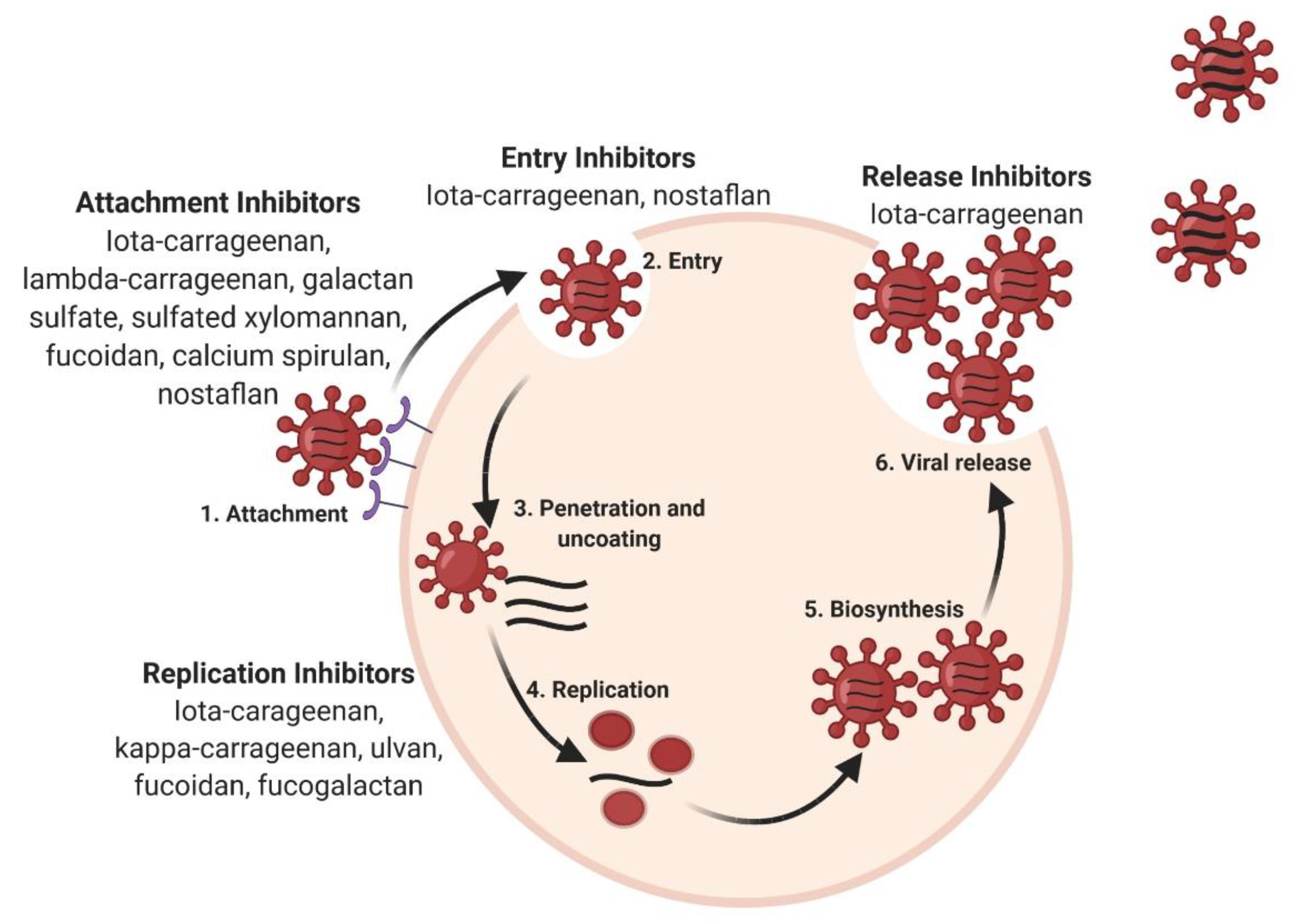
3. Antiviral Activity of Sulfated Polysaccharides and Their Underlying Mechanisms of Action
3.1. Inhibition of Infection by Electrostatic Interaction
3.2. Inhibition of Infection by Allosteric Interaction
3.3. Modulation of Intracellular Antiviral Activity
4. Antiviral Activity of Seaweed Sulfated Polysaccharides against Respiratory Viruses
4.1. Coronaviruses
4.2. Influenza Virus
4.3. Human Parainfluenza Virus
4.4. Respiratory Syncytial Virus
4.5. Human Metapneumovirus
4.6. Human Rhinovirus
4.7. Adenoviruses
5. Limits to be Overcome for the Use of Seaweed Polysaccharides against Viral Respiratory Infections
5.1. Variability in Composition: Physicochemical Characterization
5.2. Complexity of Origin and Sustainability
5.3. Need for an Improved Methodology for Antiviral Properties Assessment
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tang, J.W.; Lam, T.T.; Zaraket, H.; Lipkin, W.I.; Drews, S.J.; Hatchette, T.F.; Heraud, J.M.; Koopmans, M.P.; Abraham, A.M.; Baraket, A.; et al. Global Epidemiology of Non-Influenza RNA Respiratory Viruses: Data Gaps and a Growing Need for Surveillance. Lancet Infect. Dis. 2017, 17, e320–e326. [Google Scholar] [CrossRef]
- Watkins, K. Emerging Infectious Diseases: A Review. Curr. Emerg. Hosp. Med. Rep. 2018, 6, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Patchett, D.; Yang, J.; Northern, J.; Salinas, M.; Bauer, B.A. Viral Respiratory Infections: An Ounce of Prevention Is Worth a Pound of Cure. Mayo Clin. Proc. Innov. Qual. Outcomes 2021, 5, 480–485. [Google Scholar] [CrossRef]
- Nicola, M.; Alsafi, Z.; Sohrabi, C.; Kerwan, A.; Al-Jabir, A.; Iosifidis, C.; Agha, M.; Agha, R. The Socio-Economic Implications of the Coronavirus Pandemic (COVID-19): A Review. Int. J. Surg. 2020, 78, 185–193. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Research Needs for the Battle against Respiratory Viruses (BRaVe). Future Virol. 2013, 1–42. [Google Scholar]
- Van Doorn, H.R.; Yu, H. Viral Respiratory Infections, 2020, 10th ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Al-Tawfik, J.A.; Memish, Z.A. Emerging Respiratory Viral Infections: MERS-CoV and Influenza. Lancet Respir. Med. 2014, 2, 23–25. [Google Scholar] [CrossRef]
- Zhang, N.; Wang, L.; Deng, X.; Liang, R.; Su, M.; He, C.; Hu, L.; Su, Y.; Ren, J.; Yu, F.; et al. Recent Advances in the Detection of Respiratory Virus Infection in Humans. J. Med. Virol. 2020, 92, 408–417. [Google Scholar] [CrossRef]
- Islam, M.T.; Sarkar, C.; El-Kersh, D.M.; Jamaddar, S.; Uddin, S.J.; Shilpi, J.A.; Mubarak, M.S. Natural Products and Their Derivatives against Coronavirus: A Review of the Non-Clinical and Pre-Clinical Data. Phyther. Res. 2020, 34, 2471–2492. [Google Scholar] [CrossRef]
- Moriyama, M.; Hugentobler, W.J.; Iwasaki, A. Seasonality of Respiratory Viral Infections. Annu. Rev. Virol. 2020, 7, 83–101. [Google Scholar] [CrossRef]
- Troeger, C.E.; Blacker, B.F.; Khalil, I.A.; Zimsen, S.R.M.; Albertson, S.B.; Abate, D.; Abdela, J.; Adhikari, T.B.; Aghayan, S.A.; Agrawal, S.; et al. Mortality, Morbidity, and Hospitalisations Due to Influenza Lower Respiratory Tract Infections, 2017: An Analysis for the Global Burden of Disease Study 2017. Lancet Respir. Med. 2019, 7, 69–89. [Google Scholar] [CrossRef] [Green Version]
- Shi, T.; Denouel, A.; Tietjen, A.K.; Campbell, I.; Moran, E.; Li, X.; Campbell, H.; Demont, C.; Nyawanda, B.O.; Chu, H.Y.; et al. Global Disease Burden Estimates of Respiratory Syncytial Virus-Associated Acute Respiratory Infection in Older Adults in 2015: A Systematic Review and Meta-Analysis. J. Infect. Dis. 2020, 222, S577–S583. [Google Scholar] [CrossRef] [Green Version]
- Herfst, S.; Böhringer, M.; Karo, B.; Lawrence, P.; Lewis, N.S.; Mina, M.J.; Russell, C.J.; Steel, J.; de Swart, R.L.; Menge, C. Drivers of Airborne Human-to-Human Pathogen Transmission. Curr. Opin. Virol. 2017, 22, 22–29. [Google Scholar] [CrossRef]
- Shi, T.; McLean, K.; Campbell, H.; Nair, H. Aetiological Role of Common Respiratory Viruses in Acute Lower Respiratory Infections in Children under Five Years: A Systematic Review and Meta–Analysis. J. Glob. Health 2015, 5, 1–10. [Google Scholar] [CrossRef]
- Buetti, N.; Wicky, P.H.; Le Hingrat, Q.; Ruckly, S.; Mazzuchelli, T.; Loiodice, A.; Trimboli, P.; Forni Ogna, V.; de Montmollin, E.; Bernasconi, E.; et al. SARS-CoV-2 Detection in the Lower Respiratory Tract of Invasively Ventilated ARDS Patients. Crit. Care 2020, 24, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K. SARS CoV-2 Detection from Upper and Lower Respiratory Tract Specimens: Diagnostic and Infection Control Implications. Chest 2020, 158, 1804–1805. [Google Scholar] [CrossRef] [PubMed]
- Linden, D.; Guo-Parke, H.; Coyle, P.V.; Fairley, D.; McAuley, D.F.; Taggart, C.C.; Kidney, J. Respiratory Viral Infection: A Potential “Missing Link” in the Pathogenesis of COPD. Eur. Respir. Rev. 2019, 28, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Desforges, M.; Le Coupanec, A.; Dubeau, P.; Bourgouin, A.; Lajoie, L.; Dubé, M.; Talbot, P.J. Human Coronaviruses and Other Respiratory Viruses: Underestimated Opportunistic Pathogens of the Central Nervous System? Viruses 2019, 12, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brendish, N.J.; Clark, T.W. Antiviral Treatment of Severe Non-Influenza Respiratory Virus Infection. Curr. Opin. Infect. Dis. 2017, 30, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Pagarete, A.; Ramos, A.S.; Puntervoll, P.; Allen, M.J.; Verdelho, V. Antiviral Potential of Algal Metabolites—A Comprehensive Review. Mar. Drugs 2021, 19, 94. [Google Scholar] [CrossRef]
- Peghin, M.; Danziger-Isakov, L. Prevention and Treatment of Respiratory Virus Infection. In Infectious Diseases in Solid-Organ Transplant Recipients; Manuel, O., Ison, M., Eds.; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Papadopoulos, N.G.; Megremis, S.; Kitsioulis, N.A.; Vangelatou, O.; West, P.; Xepapadaki, P. Promising Approaches for the Treatment and Prevention of Viral Respiratory Illnesses. J. Allergy Clin. Immunol. 2017, 140, 921–932. [Google Scholar] [CrossRef] [Green Version]
- Shie, J.J.; Fang, J.M. Development of Effective Anti-Influenza Drugs: Congeners and Conjugates—A Review. J. Biomed. Sci. 2019, 26, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Dodge, M.J.; MacNeil, K.M.; Tessier, T.M.; Weinberg, J.B.; Mymryk, J.S. Emerging Antiviral Therapeutics for Human Adenovirus Infection: Recent Developments and Novel Strategies. Antiviral Res. 2021, 188, 105034. [Google Scholar] [CrossRef]
- Boyoglu-Barnum, S.; Tripp, R.A. Up-to-Date Role of Biologics in the Management of Respiratory Syncytial Virus. Expert Opin. Biol. Ther. 2020, 20, 1073–1082. [Google Scholar] [CrossRef]
- Kalergis, A.M.; Soto, J.A.; Gálvez, N.M.S.; Andrade, C.A.; Fernandez, A.; Bohmwald, K.; Bueno, S. Pharmacological Management of Human Respiratory Syncytial Virus Infection. Expert Opin. Pharmacother. 2020, 21, 2293–2303. [Google Scholar] [CrossRef] [PubMed]
- Vivekanandhan, K.; Shanmugam, P.; Barabadi, H.; Arumugam, V.; Daniel Raj Daniel Paul Raj, D.; Sivasubramanian, M.; Ramasamy, S.; Anand, K.; Boomi, P.; Chandrasekaran, B.; et al. Emerging Therapeutic Approaches to Combat COVID-19: Present Status and Future Perspectives. Front. Mol. Biosci. 2021, 8, 1–10. [Google Scholar] [CrossRef]
- Beigel, J.H.; Nam, H.H.; Adams, P.L.; Krafft, A.; Ince, W.L.; El-Kamary, S.S.; Sims, A.C. Advances in Respiratory Virus Therapeutics—A Meeting Report from the 6th Isirv Antiviral Group Conference. Antiviral Res. 2019, 167, 45–67. [Google Scholar] [CrossRef] [PubMed]
- Shahani, L.; Ariza-Heredia, E.J.; Chemaly, R.F. Antiviral Therapy for Respiratory Viral Infections in Immunocompromised Patients. Expert Rev. Anti. Infect. Ther. 2017, 15, 401–415. [Google Scholar] [CrossRef] [PubMed]
- Krammer, F.; Smith, G.J.D.; Fouchier, R.A.M.; Peiris, M.; Kedzierska, K.; Doherty, P.C.; Palese, P.; Shaw, M.L.; Treanor, J.; Webster, R.G.; et al. Influenza. Nat. Rev. Dis. Prim. 2018, 4, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Krylova, N.V.; Ermakova, S.P.; Lavrov, V.F.; Leneva, I.A.; Kompanets, G.G.; Iunikhina, O.V.; Nosik, M.N.; Ebralidze, L.K.; Falynskova, I.N.; Silchenko, A.S.; et al. The Comparative Analysis of Antiviral Activity of Native and Modified Fucoidans from Brown Algae Fucus Evanescens in Vitro and in Vivo. Mar. Drugs 2020, 18, 224. [Google Scholar] [CrossRef] [Green Version]
- Alfayate Miguélez, S.; Garcia-Marcos, L. Rational Use of Antimicrobials in the Treatment of Upper Airway Infections. J. Pediatr. 2020, 96, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhou, Y.H.; Ye, F.; Yang, Z.Q. Antivirals for Respiratory Viral Infections: Problems and Prospects. Semin. Respir. Crit. Care Med. 2016, 37, 640–646. [Google Scholar] [CrossRef]
- Johnson, D.C. Antiviral Drugs for Common Respiratory Diseases What’s Here, What’s to Come. Postgrad. Med. 1988, 83, 136–148. [Google Scholar] [CrossRef]
- Bilal, M.; Iqbal, H.M.N. Marine Seaweed Polysaccharides-Based Engineered Cues for the Modern Biomedical Sector. Mar. Drugs 2020, 18, 7. [Google Scholar] [CrossRef] [Green Version]
- Grice, I.D.; Mariottini, G.L. Glycans with Antiviral Activity from Marine Organisms. In Marine Organisms as Model Systems in Biology and Medicine. Results and Problems in Cell Differentiation; Kloc, M., Kubiak, J., Eds.; Springer: Berlin, Germany, 2018; pp. 439–475. [Google Scholar]
- Wopker, P.M.; Schwermer, M.; Sommer, S.; Längler, A.; Fetz, K.; Ostermann, T.; Zuzak, T.J. Complementary and Alternative Medicine in the Treatment of Acute Bronchitis in Children: A Systematic Review. Complement. Ther. Med. 2020, 49, 102217. [Google Scholar] [CrossRef]
- Hans, N.; Malik, A.; Naik, S. Antiviral Activity of Sulfated Polysaccharides from Marine Algae and Its Application in Combating COVID-19: Mini Review. Bioresour. Technol. Rep. 2021, 13, 100623. [Google Scholar] [CrossRef] [PubMed]
- Damonte, E.; Matulewicz, M.; Cerezo, A. Sulfated Seaweed Polysaccharides as Antiviral Agents. Curr. Med. Chem. 2012, 11, 2399–2419. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, A.; Zorofchian Moghadamtousi, S.; Abubakar, S.; Zandi, K. Antiviral Potential of Algae Polysaccharides Isolated from Marine Sources: A Review. Biomed. Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Besednova, N.; Zaporozhets, T.; Kuznetsova, T.; Makarenkova, I.; Fedyanina, L.; Kryzhanovsky, S.; Malyarenko, O.; Ermakova, S. Metabolites of Seaweeds as Potential Agents for the Prevention and Therapy of Influenza Infection. Mar. Drugs 2019, 17, 373. [Google Scholar] [CrossRef] [Green Version]
- Synytsya, A.; Bleha, R.; Synytsya, A.; Pohl, R.; Hayashi, K.; Yoshinaga, K.; Nakano, T.; Hayashi, T. Mekabu Fucoidan: Structural Complexity and Defensive Effects against Avian Influenza A Viruses. Carbohydr. Polym. 2014, 111, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Ghosh, K.; Hahn, F.; Wangen, C.; Strojan, H.; Müller, R.; Anand, N.; Ali, I.; Bera, K.; Ray, B.; et al. Chemically Sulfated Polysaccharides from Natural Sources: Assessment of Extraction-Sulfation Efficiencies, Structural Features and Antiviral Activities. Int. J. Biol. Macromol. 2019, 136, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Zeng, K.; Groth, T.; Zhang, K. Recent Advances in Artificially Sulfated Polysaccharides for Applications in Cell Growth and Differentiation, Drug Delivery, and Tissue Engineering. ChemBioChem 2019, 20, 737–746. [Google Scholar] [CrossRef]
- Wang, W.; Wang, S.X.; Guan, H.S. The Antiviral Activities and Mechanisms of Marine Polysaccharides: An Overview. Mar. Drugs 2012, 10, 2795–2816. [Google Scholar] [CrossRef] [PubMed]
- Bobrovnitsky, I.P.; Mikhailov, V.I.; Odinets, A.G.; Neretina, T.V.; Dobrynina, T.V.; Klinov, D.V. Study of the Structure of Fucoidan (Isolated from Laminaria Japonica) and the Mechanism of Its Antiviral Activity by Atomic Force Microscopy. New Med. Technol. 2010, 2, 24–28. [Google Scholar]
- Queiroz, K.C.S.; Medeiros, V.P.; Queiroz, L.S.; Abreu, L.R.D.; Rocha, H.A.O.; Ferreira, C.V.; Jucá, M.B.; Aoyama, H.; Leite, E.L. Inhibition of Reverse Transcriptase Activity of HIV by Polysaccharides of Brown Algae. Biomed. Pharmacother. 2008, 62, 303–307. [Google Scholar] [CrossRef]
- Wang, W.; Wu, J.; Zhang, X.; Hao, C.; Zhao, X.; Jiao, G.; Shan, X.; Tai, W.; Yu, G. Inhibition of Influenza A Virus Infection by Fucoidan Targeting Viral Neuraminidase and Cellular EGFR Pathway. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Makarenkova, I.D.; Logunov, D.Y.; Tukhvatulin, A.I.; Semenova, I.B.; Besednova, N.N.; Zvyagintseva, T.N. Interactions between Sulfated Polysaccharides from Sea Brown Algae and Toll-like Receptors on HEK293 Eukaryotic Cells in Vitro. Bull. Exp. Biol. Med. 2012, 154, 241–244. [Google Scholar] [CrossRef]
- Hayashi, K.; Lee, J.B.; Nakano, T.; Hayashi, T. Anti-Influenza A Virus Characteristics of a Fucoidan from Sporophyll of Undaria Pinnatifida in Mice with Normal and Compromised Immunity. Microbes Infect. 2013, 15, 302–309. [Google Scholar] [CrossRef]
- Bekerman, E.; Einav, S. Combating Emerging Viral Threats. Science 2015, 348, 282–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Han, W.; Wang, G.; Zhao, X. Application Prospect of Polysaccharides in the Development of Anti-Novel Coronavirus Drugs and Vaccines. Int. J. Biol. Macromol. 2020, 164, 331–343. [Google Scholar] [CrossRef]
- Leibbrandt, A.; Meier, C.; König-Schuster, M.; Weinmüllner, R.; Kalthoff, D.; Pflugfelder, B.; Graf, P.; Frank-Gehrke, B.; Beer, M.; Fazekas, T.; et al. Iota-Carrageenan Is a Potent Inhibitor of Influenza a Virus Infection. PLoS ONE 2010, 5, 1–11. [Google Scholar] [CrossRef]
- Sun, T.; Zhang, X.; Miao, Y.; Zhou, Y.; Shi, J.; Yan, M.; Chen, A. Studies on Antiviral and Immuno-Regulation Activity of Low Molecular Weight Fucoidan from Laminaria Japonica. J. Ocean. Univ. China 2018, 17, 705–711. [Google Scholar] [CrossRef]
- Grassauer, A.; Weinmuellner, R.; Meier, C.; Pretsch, A.; Prieschl-Grassauer, E.; Unger, H. Iota-Carrageenan Is a Potent Inhibitor of Rhinovirus Infection. Virol. J. 2008, 5, 5–7. [Google Scholar] [CrossRef] [Green Version]
- Hemilä, H.; Chalker, E. Carrageenan Nasal Spray May Double the Rate of Recovery from Coronavirus and Influenza Virus Infections: Re-Analysis of Randomized Trial Data. Research Square, preprint. Available online: https://assets.researchsquare.com/files/rs-108775/v1/668450ee-3527-4c54-8ea5-da3298c76e84.pdf (accessed on 15 May 2021).
- Ciejka, J.; Botwina Pawełand Nowakowska, M.; Szczubiałka, K.; Pyrc, K. Synthetic Sulfonated Derivatives of Poly (Allylamine Hydrochloride) as Inhibitors of Human Metapneumovirus. PLoS ONE 2019, 14, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matloub, A.A.; Elsouda, S.S.M.; El-Senousy, W.M.; Hamed, M.; Aly, H.; Ali, S.A.; Mohammed, R.S.; Mahmoud, K.; El-Hallouty, S.; Ibrahim, N.A.; et al. In Vitro Antiviral, Cytotoxic, Antioxidant and Hypolipidemic Activites of Polysaccharide Isolated from Marine Algae. Int. J. Pharmacogn. Phytochem. Res. 2015, 7, 1099–1111. [Google Scholar]
- Witvrouw, M.; Este, J.A.; Mateu, M.Q.; Reymen, D.; Andrei, G.; Snoeck, R.; Ikeda, S.; Pauwels, R.; Bianchini, N.V.; Desmyter, J.; et al. Activity of a Sulfated Polysaccharide Extracted from the Red Seaweed Aghardhiella Tenera against Human Immunodeficiency Virus and Other Enveloped Viruses. Antivir. Chem. Chemother. 1994, 5, 297–303. [Google Scholar] [CrossRef] [Green Version]
- Damonte, E.; Neyts, J.; Pujol, C.A.; Snoeck, R.; Andrei, G.; Ikeda, S.; Witvrouw, M.; Reymen, D.; Haines, H.; Matulewicz, M.C.; et al. Antiviral Activity of a Sulphated Polysaccharide from the Red Seaweed Nothogenia Fastigiata. Biochem. Pharmacol. 1994, 47, 2187–2192. [Google Scholar] [CrossRef]
- Tang, F.; Chen, F.; Li, F. Preparation and Potential in Vivo Anti-Influenza Virus Activity of Low Molecular-Weight $κ$-Carrageenans and Their Derivatives. J. Appl. Polym. Sci. 2013, 127, 2110–2115. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, P.; Hao, C.; Zhang, X.E.; Cui, Z.Q.; Guan, H.S. In Vitro Inhibitory Effect of Carrageenan Oligosaccharide on Influenza A H1N1 Virus. Antiviral Res. 2011, 92, 237–246. [Google Scholar] [CrossRef]
- Shao, Q.; Guo, Q.; Xu, W.P.; Li, Z.; Zhao, T.T. Specific Inhibitory Effect of κ-Carrageenan Polysaccharide on Swine Pandemic 2009 H1N1 Influenza Virus. PLoS ONE 2015, 10, 1–17. [Google Scholar] [CrossRef]
- Jang, Y.; Shin, H.; Lee, M.K.; Kwon, O.S.; Shin, J.S.; Kim, Y.; Kim, C.W.; Lee, H.R.; Kim, M. Antiviral Activity of Lambda-Carrageenan against Influenza Viruses and Severe Acute Respiratory Syndrome Coronavirus 2. Sci. Rep. 2021, 11, 1–26. [Google Scholar] [CrossRef]
- Yu, G.; Li, M.; Wang, W.; Liu, X.; Zhao, X.; Lv, Y.; Li, G.; Liao, G.; Zhao, X. Structure and Anti-Influenza A (H1N1) Virus Activity of Three Polysaccharides from Eucheuma Denticulatum. J. Ocean. Univ. China 2012, 11, 527–532. [Google Scholar] [CrossRef]
- Chen, M.Z.; Xie, H.G.; Yang, L.W.; Liao, Z.H.; Yu, J. In Vitro Anti-Influenza Virus Activities of Sulfated Polysaccharide Fractions from Gracilaria Lemaneiformis. Virol. Sin. 2010, 25, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Bansal, S.; Jonsson, C.B.; Taylor, S.L.; Manuel Figueroa, J.; Vanesa, A.; Palacios, C.; César Vega, J. Iota-Carrageenan and Xylitol Inhibit SARS-CoV-2 in Cell Culture. bioRxiv 2020. [Google Scholar] [CrossRef]
- Morokutti-Kurz, M.; Fröba, M.; Graf, P.; Große, M.; Grassauer, A.; Auth, J.; Schubert, U.; Prieschl-Grassauer, E. Iota-Carrageenan Neutralizes SARS-CoV-2 and Inhibits Viral Replication in Vitro. PLoS ONE 2021, 16, 1–13. [Google Scholar] [CrossRef]
- Song, S.; Peng, H.; Wang, Q.; Liu, Z.; Dong, X.; Wen, C.; Ai, C.; Zhang, Y.; Wang, Z.; Zhu, B. Inhibitory Activities of Marine Sulfated Polysaccharides against SARS-CoV-2. Food Funct. 2020, 11, 7415–7420. [Google Scholar] [CrossRef]
- Morokutti-Kurz, M.; Graf, C.; Prieschl-Grassauer, E. Amylmetacresol/2,4-Dichlorobenzyl Alcohol, Hexylresorcinol, or Carrageenan Lozenges as Active Treatments for Sore Throat. Int. J. Gen. Med. 2017, 10, 53–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.G.; Hao, Y.; Li, Z.H.; Liu, S.T.; Wang, L.X. Antiviral Activity of Polysaccharide Extract from Laminaria Japonica against Respiratory Syncytial Virus. Biomed. Pharmacother. 2016, 84, 1705–1710. [Google Scholar] [CrossRef]
- Kwon, P.S.; Oh, H.; Kwon, S.J.; Jin, W.; Zhang, F.; Fraser, K.; Hong, J.J.; Linhardt, R.J.; Dordick, J.S. Sulfated Polysaccharides Effectively Inhibit SARS-CoV-2 in Vitro. Cell Discov. 2020, 6, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Makarenkova, I.D.; Deryabin, P.G.; Lvov, D.K.; Zvyagintseva, T.N.; Besednova, N.N. Antiviral Activity of Virus, Sulfated Polysaccharide from the Brown Algae Laminaria Japonica against Avian Influenza A (H5N1) Infection in the Cultured Cells. Probl. Virol. 2010, 1, 41–45. [Google Scholar]
- Komatsu, T.; Kido, N.; Sugiyama, T.; Yokochi, T. Antiviral Activity of Acidic Polysaccharides from Coccomyxa Gloeobotrydiformi, a Green Alga, against an in Vitro Human Influenza A Virus Infection. Immunopharmacol. Immunotoxicol. 2013, 35, 1–7. [Google Scholar] [CrossRef]
- Ivanova, V.; Rouseva, R.; Kolarova, M.; Serkedjieva, J.; Rachev, R.; Manolova, N. Isolation of a Polysaccharide with Antiviral Effect from Ulva Lactuca. Prep. Biochem. 1994, 24, 83–97. [Google Scholar] [CrossRef]
- Mendes, G.d.S.; Soares, A.R.; Martins, F.O.; de Albuquerque, M.C.M.; Costa, S.S.; Yoneshigue-Valentin, Y.; Gestinari, L.M.d.S.; Santos, N.; Romanos, M.T.V. Antiviral Activity of the Green Marine Alga Ulva Fasciata on the Replication of Human Metapneumovirus. Rev. Inst. Med. Trop. Sao Paulo 2010, 52, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, T.; Hayashi, K.; Maeda, M.; Kojima, I. Calcium Spirulan, an Inhibitor of Enveloped Virus Replication, from a Blue-Green Alga Spirulina Platensis. J. Nat. Prod. 1996, 59, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Kanekiyo, K.; Hayashi, K.; Takenaka, H.; Lee, J.B.; Hayashi, T. Anti-Herpes Simplex Virus Target of an Acidic Polysaccharide, Nostoflan, from the Edible Blue-Green Alga Nostoc Flagelliforme. Biol. Pharm. Bull. 2007, 30, 1573–1575. [Google Scholar] [CrossRef] [Green Version]
- Snell, N.J.C. New Treatments for Viral Respiratory Tract Infections—Opportunities and Problems. J. Antimicrob. Chemother. 2001, 47, 251–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Zhang, P.; Yu, G.L.; Li, C.X.; Hao, C.; Qi, X.; Zhang, L.J.; Guan, H.S. Preparation and Anti-Influenza A Virus Activity of $κ$-Carrageenan Oligosaccharide and Its Sulphated Derivatives. Food Chem. 2012, 133, 880–888. [Google Scholar] [CrossRef]
- Song, L.; Chen, X.; Liu, X.; Zhang, F.; Hu, L.; Yue, Y.; Li, K.; Li, P. Characterization and Comparison of the Structural Features, Immune-Modulatory and Anti-Avian Influenza Virus Activities Conferred by Three Algal Sulfated Polysaccharides. Mar. Drugs 2016, 14, 4. [Google Scholar] [CrossRef]
- Akamatsu, E.; Shimanaga, M.; Kamei, Y. Isolation of an Anti-Influenza Virus Substance, MC26 Virus, from a Marine Brown Alga Sargassum Piluliferum and Its Antiviral Activity against Influenza. Coast. Bioenviron. 2003, 1, 29–34. [Google Scholar]
- Chibanga, V.P.; Dirr, L.; Guillon, P.; El-Deeb, I.M.; Bailly, B.; Thomson, R.J.; von Itzstein, M. New Antiviral Approaches for Human Parainfluenza: Inhibiting the Haemagglutinin-Neuraminidase. Antiviral Res. 2019, 167, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Graf, C.; Bernkop-Schnürch, A.; Egyed, A.; Koller, C.; Prieschl-Grassauer, E.; Morokutti-Kurz, M. Development of a Nasal Spray Containing Xylometazoline Hydrochloride and Iota-Carrageenan for the Symptomatic Relief of Nasal Congestion Caused by Rhinitis and Sinusitis. Int. J. Gen. Med. 2018, 11, 275–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, M.; Enzenhofer, E.; Schneider, S.; Rauch, M.; Bodenteich, A.; Neumann, K.; Prieschl-Grassauer, E.; Grassauer, A.; Lion, T.; Mueller, C.A. Efficacy of a Carrageenan Nasal Spray in Patients with Common Cold: A Randomized Controlled Trial. Respir. Res. 2013, 14, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eccles, R.; Winther, B.; Johnston, S.L.; Robinson, P.; Trampisch, M.; Koelsch, S. Efficacy and Safety of Iota-Carrageenan Nasal Spray versus Placebo in Early Treatment of the Common Cold in Adults: The ICICC Trial. Respir. Res. 2015, 16, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koenighofer, M.; Lion, T.; Bodenteich, A.; Prieschl-Grassauer, E.; Grassauer, A.; Unger, H.; Mueller, C.A.; Fazekas, T. Carrageenan Nasal Spray in Virus Confirmed Common Cold: Individual Patient Data Analysis of Two Randomized Controlled Trials. Multidiscip. Respir. Med. 2014, 9, 1–12. [Google Scholar] [CrossRef]
- Hebar, A.; Koller, C.; Seifert, J.M.; Chabicovsky, M.; Bodenteich, A.; Bernkop-Schnürch, A.; Grassauer, A.; Prieschl-Grassauer, E. Non-Clinical Safety Evaluation of Intranasal Iota-Carrageenan. PLoS ONE 2015, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Harrach, B.; Tarján, Z.L.; Benkő, M. Adenoviruses across the Animal Kingdom: A Walk in the Zoo. FEBS Lett. 2019, 593, 3660–3673. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.; Hashim, S.N.; Rahman, H.A. Seaweeds: A Sustainable Functional Food for Complementary and Alternative Therapy. Trends Food Sci. Technol. 2012, 23, 83–96. [Google Scholar] [CrossRef]
- Torres, M.D.; Flórez-Fernández, N.; Domínguez, H. Integral Utilization of Red Seaweed for Bioactive Production. Mar. Drugs 2019, 17, 314. [Google Scholar] [CrossRef] [Green Version]
- Jiao, G.; Yu, G.; Zhang, J.; Stephen Ewart, H. Chemical Structures and Bioactivities of Sulfated Polysaccharides from Marine Algae. Mar. Drugs 2011, 9, 196–223. [Google Scholar] [CrossRef] [Green Version]
- Grü, N.; Groth, I.; Alban, S. Evaluation of Seasonal Variations of the Structure and Anti-Inflammatory Activity of Sulfated Polysaccharides Extracted from the Red Alga Delesseria Sanguinea (Hudson) Lamouroux (Ceramiales, Delesseriaceae). Biomacromolecules 2009, 10, 1155–1162. [Google Scholar]
- Stengel, D.B.; Connan, S.; Popper, Z.A. Algal Chemodiversity and Bioactivity: Sources of Natural Variability and Implications for Commercial Application. Biotechnol. Adv. 2011, 29, 483–501. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.; Vieira, H.; Gaspar, H.; Santos, S. Marketed Marine Natural Products in the Pharmaceutical and Cosmeceutical Industries: Tips for Success. Mar. Drugs 2014, 12, 1066–1101. [Google Scholar] [CrossRef] [Green Version]
- Lühn, S.; Grimm, J.C.; Alban, S. Simple and Rapid Quality Control of Sulfated Glycans by a Fluorescence Sensor Assay—Exemplarily Developed for the Sulfated Polysaccharides from Red Algae Delesseria Sanguinea. Mar. Drugs 2014, 12, 2205–2227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koehn, F.E.; Carter, G.T. The Evolving Role of Natural Products in Drug Discovery. Nat. Rev. Drug Discov. 2005, 4, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Molinski, T.F.; Dalisay, D.S.; Lievens, S.L.; Saludes, J.P. Drug Development from Marine Natural Products. Nat. Rev. Drug Discov. 2009, 8, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Bedini, E.; Laezza, A.; Parrilli, M.; Iadonisi, A. A Review of Chemical Methods for the Selective Sulfation and Desulfation of Polysaccharides. Carbohydr. Polym. 2017, 174, 1224–1239. [Google Scholar] [CrossRef]
- Wang, Z.; Xie, J.; Shen, M.; Nie, S.; Xie, M. Sulfated Modification of Polysaccharides: Synthesis, Characterization and Bioactivities. Trends Food Sci. Technol. 2018, 74, 147–157. [Google Scholar] [CrossRef]
- Dias, D.A.; Urban, S.; Roessner, U. A Historical Overview of Natural Products in Drug Discovery. Metabolites 2012, 2, 303–336. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L.; Critchley, A.T. The COVID 19 Novel Coronavirus Pandemic 2020: Seaweeds to the Rescue? Why Does Substantial, Supporting Research about the Antiviral Properties of Seaweed Polysaccharides Seem to Go Unrecognized by the Pharmaceutical Community in These Desperate Times? J. Appl. Phycol. 2020, 32, 1875–1877. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| * FDA/EMA Approved Antivirals | ||||
|---|---|---|---|---|
| Respiratory Virus | Synthetic Antivirals | Natural Seaweed Polysaccharides | Vaccine | Reference |
| Human rhinovirus | No | Iota-carrageenan | No | |
| Adenovirus | Cidofovir, ganciclovir, ribavirin | No | Yes (not accessible to all) | [24] |
| Human metapneumovirus | No | No | No | |
| Parainfluenza virus | No | No | No | |
| Influenza virus | Ribavirin, amantadine | No | Yes | [23] |
| Rimantadine, zanamivir | ||||
| Oseltamivir, peramivir | ||||
| Laninamivir | ||||
| Respiratory syncytial virus | Ribavarin, palivizumab | No | No | [25,26] |
| SARS CoV-2 | Remdesivir | No | Yes | [27] |
| HCoV-OC43 | No | No | No | |
| Marine Algae | Polysaccharide | Respiratory Virus | Entry Inhibition | Replication Inhibition | Reference |
|---|---|---|---|---|---|
| Red Algae | Iota-carrageenan | HRV | × | [55,56] | |
| Iota-carrageenan | HMPV | × | [57] | ||
| Iota-carrageenan | Adenovirus | × | [58] | ||
| Galactan sulfate | HPIV | × | [59] | ||
| Galactan sulfate | Influenza | × | [59] | ||
| Sulfated xylomannan | × | [60] | |||
| kappa-carrageenan | × | [45,61,62,63] | |||
| Iota-carrageenan | × | [53,56] | |||
| Lambda-carrageenan | × | [64] | |||
| hybrid carrageenan (ı/κ/ν-carrageenan) | Not mentioned | [65] | |||
| Polysaccharide fractions | × | × | [66] | ||
| Galactan sulfate | RSV | × | [39,59] | ||
| Sulfated xylomannan | × | [60] | |||
| Iota-carrageenan | SARS CoV-2 | × | [67,68] | ||
| Iota-carrageenan | × | [68,69] | |||
| Lambda-carrageenan | × | [64] | |||
| Iota-carrageenan | HCoV-OC43 | × | [56,70] | ||
| Brown Algae | Fucoidan | Adenovirus | × | [54] | |
| Fucoidan | HPIV | × | [54] | ||
| Extract | RSV | × | [71] | ||
| Fucoidan | SARS CoV-2 | × | [69,72] | ||
| Fucoidan | Influenza | × | [41,54,73] | ||
| × | [48] | ||||
| Fucogalactan | × | [42,50] | |||
| Green Algae | Acidic polysaccharide | Influenza | × | [74] | |
| Ulvan polysaccharide | × | [75] | |||
| Extracts | HMPV | × | × | [76] | |
| Blue-GreenAlgae | Calcium spirulan | Influenza | × | [77] | |
| Nostaflan | × | [78] | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jabeen, M.; Dutot, M.; Fagon, R.; Verrier, B.; Monge, C. Seaweed Sulfated Polysaccharides against Respiratory Viral Infections. Pharmaceutics 2021, 13, 733. https://doi.org/10.3390/pharmaceutics13050733
Jabeen M, Dutot M, Fagon R, Verrier B, Monge C. Seaweed Sulfated Polysaccharides against Respiratory Viral Infections. Pharmaceutics. 2021; 13(5):733. https://doi.org/10.3390/pharmaceutics13050733
Chicago/Turabian StyleJabeen, Mehwish, Mélody Dutot, Roxane Fagon, Bernard Verrier, and Claire Monge. 2021. "Seaweed Sulfated Polysaccharides against Respiratory Viral Infections" Pharmaceutics 13, no. 5: 733. https://doi.org/10.3390/pharmaceutics13050733