1. Introduction
Chagas disease, also known as American trypanosomiasis, is caused by the protozoan
Trypanosoma cruzi. This neglected infectious disease affects 6 to 7 million people worldwide and is endemic in 21 countries of Latin America [
1,
2]. Nowadays, due to migratory flows, Chagas disease has spread to non-endemic countries such as Spain, Canada, Australia, Japan, and North America, becoming a relevant public health challenge [
3,
4].
Chagas disease has two clinical phases. In the acute phase, the parasite disseminates and can be detected by direct bloodstream examination [
5]. In most cases, it courses without symptoms and if there are any symptoms, they are usually mild or nonspecific. The acute phase usually lasts up to 2 months and resolves spontaneously. However, in 5–10% of infected subjects, the disease may become fatal because of heart failure or meningoencephalitis [
6,
7]. After the acute phase, the disease enters the chronic phase, which can adopt two forms. In about 60–70% of cases, the disease remains in the indeterminate form, which may last 10–30 years or for the lifetime of the patient. This form is silent and free of symptoms. In the remaining cases (30–40% of individuals), the patients develop a chronic form of the disease, which mostly involves severe cardiac, digestive, and neurological disorders [
8,
9,
10].
For more than 40 years, only two drugs have been available for the treatment of Chagas disease: nifurtimox and benznidazole. Both have severe side effects that lead to treatment discontinuation in 15–20% of cases [
11,
12,
13,
14]. This, coupled with their limited efficacy in the chronic phase of the disease [
15] and the occurrence of naturally resistant strains [
16], reinforces the need to identify new compounds to fight Chagas disease. Combination therapy has emerged as an alternative in the treatment of Chagas disease. Synergistic drug combinations could improve current regimens, allowing doses to be reduced and consequently diminishing side effects [
17,
18].
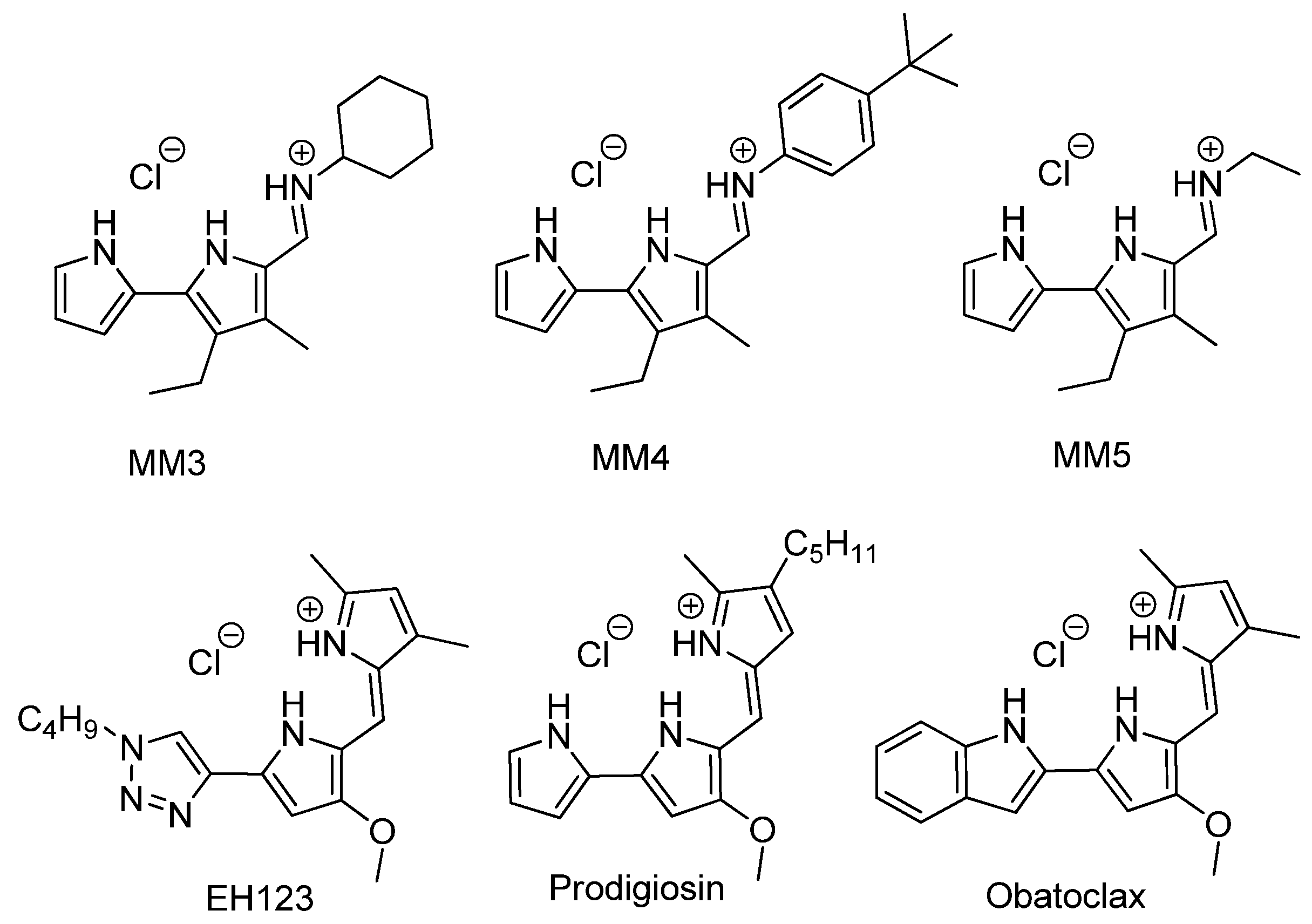
In our search for new trypanocidal drugs, we focused on tambjamines, a family of naturally occurring compounds isolated from bacteria and marine invertebrates such as ascidians, bryozoans, and nudibranchs [
19]. Tambjamines are alkaloids structurally related to prodiginines, sharing with them the 4-methoxy-2,2′-bipyrrole core and also a wide spectrum of useful biological properties that include antimicrobial, antifungal, antimalarial, and antitumor activities [
20,
21,
22].
Although the mode of action of tambjamines has not been elucidated yet, it seems to be closely linked to its ability to act as anion transporters, a feature that is also shared by prodiginines [
20,
23]. Furthermore, the mitochondria of
T. cruzi have been regarded as a potential target, since alterations in the mitochondria have catastrophic effects on respiration and, subsequently, on the survival of the parasite [
24].
Surprisingly, to our knowledge, there are no studies on the use of tambjamines as antitrypanosomal agents. However, it has been postulated that these compounds exert a remarkable effect on other protozoans and have been proposed as antimalarial agents. Kancharla et al. [
25] described the successful resolution of a
Plasmodium yoelii infection in a murine model through treatment with tambjamines. Specifically, the synthetic tambjamine KAR425 had potent in vivo activity against the
P. yoelii infection, showing no signs of cell toxicity.
Our group recently studied prodigiosin of bacterial origin, reporting its remarkable activity against the epimastigotes of
T. cruzi (CL strain, clone B5), with very low IC
50 values (up to 0.8 µM), compared to benznidazole (18 µM). Additionally, we explored the effects of this pigment on the parasite cell surface by atomic force microscopy, which revealed relevant morphological alterations in the size, shape, and roughness [
26]. Based on these encouraging preliminary results, we decided to continue exploring the trypanocidal effect of prodigiosin and related molecules against the intracellular forms of
T. cruzi and investigate the targets presumably involved.
The aims of the present work were to (1) determine the in vitro trypanocidal activity of natural and synthetic prodiginines as well as several tambjamine derivatives against the epimastigotes and trypomastigotes/amastigotes of the parasite, (2) evaluate the toxicity of these compounds in two cultured mammalian cell lines, (3) determine their biological activity in combination with the standard drug benznidazole, and (4) explore their mode of action.
4. Discussion
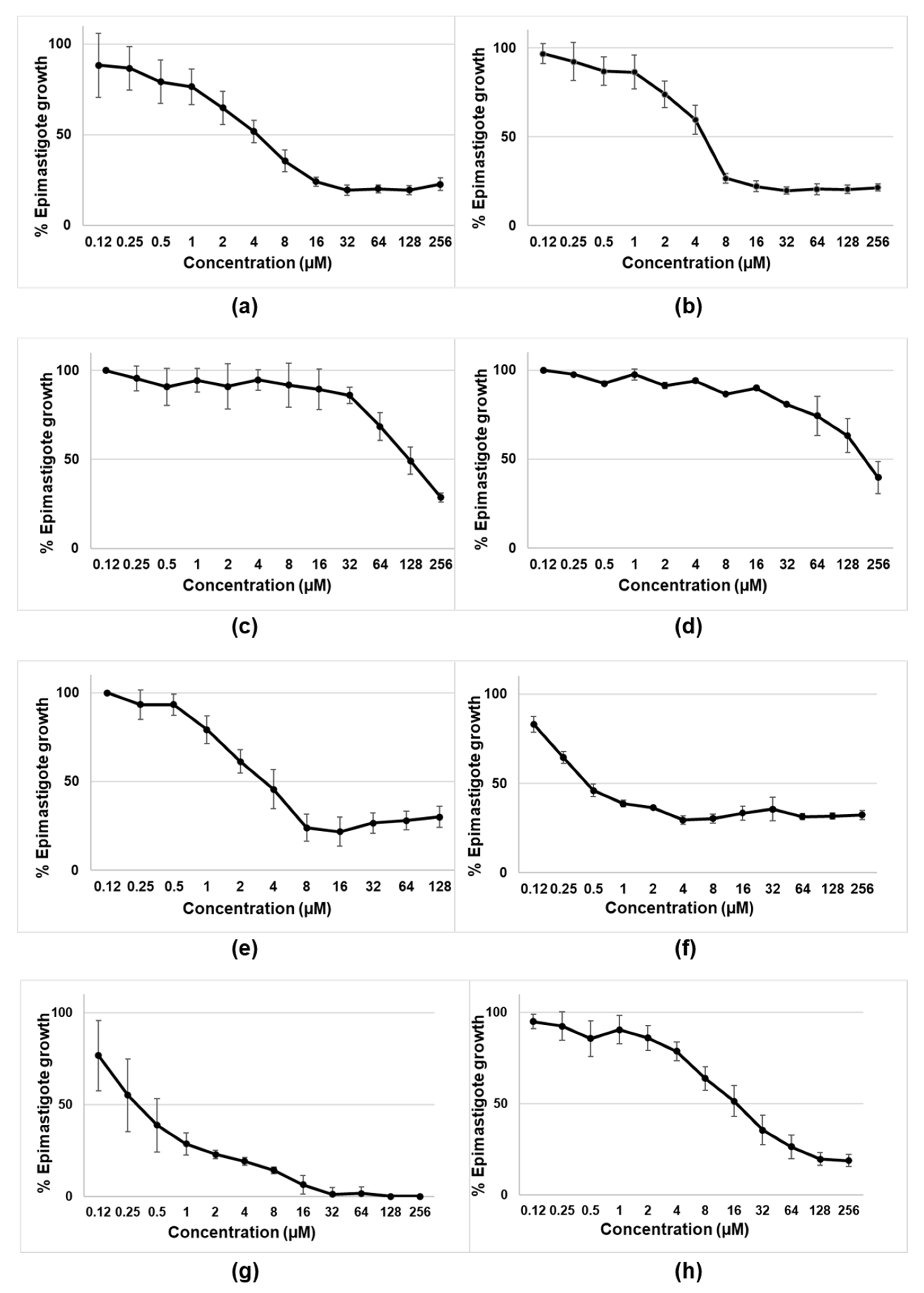
In this work, we evaluated the biological activity of several tambjamines, Obatoclax, and natural and synthetic prodiginines against T. cruzi (CLB strain). The two synthetic tambjamine derivatives (MM3 and MM4) displayed trypanocidal capability against both the extracellular and intracellular forms of T. cruzi.
To our knowledge, this is the first report on the trypanocidal effect of tambjamines, although their usefulness has been explored mainly as antimalarial agents [
25]. By contrast, derivatives MM5 and EH123 did not show detectable levels of trypanocidal activity and were subsequently discarded for the rest of the experiments. Obatoclax displayed remarkable trypanocidal activity against the epimastigotes of
T. cruzi; however, its effectiveness against the intracellular forms of
T. cruzi was slightly lower than that of the other molecules assayed. This is consistent with the results recently reported by Ehrenkaufer et al., who demonstrated that Obatoclax had remarkable activities against
Entamoeba,
Giardia, and
Trypanosoma brucei, with IC
50 values of 0.5, 0.9, and 0.04 µM, respectively [
34]. Here, we provide the effectiveness data of this compound in
T. cruzi. The observed results correlate well with the ability of these compounds to function as transmembrane anion carriers. Compounds MM3, MM4, and MM5 are synthetic tambjamines bearing different substituents in the enamine moiety. The presence of an ethyl group in MM5 resulted in the lack of trypanocidal activity, as opposed to MM3 and MM4. These two compounds are equipped with larger substituents (cyclohexyl and -4-tertbutylphenyl, respectively) resulting in more lipophilic derivatives better suited to facilitate anion transport. The replacement of a pyrrole heterocycle characteristic of both prodigiosin and obatoclax by a 1,2,3-triazole in the case of EH123 also impact negatively in the ability of this latter compound to interact and transport anions, resulting again in a marked decline in the activity of EH123 as a trypanocidal agent.
It has been pointed out that the mode of action of Obatoclax on cancer cells is mainly through its proapoptotic activity. The drug has the ability to inhibit the Bcl-2 family of proteins. However, trypanosomatids, as unicellular protists, lack apparent Bcl-2 homologs. Therefore, they should be, in principle, resistant to Obatoclax unless the mode of action of Obatoclax involves some more targets than the inhibition of this apoptotic regulator. Ehrenkaufer et al. [
34] reported that other Bcl-2 inhibitors such as venetoclax, navitoclax, A-1331852, A-1210477, and S63845 do not have an effect on the viability of different parasites, subsequently suggesting that a different mechanism of action should be investigated. Their results clearly indicated that Obatoclax most likely kills the parasites through a BCL-2-independent mechanism. Our results confirmed the potent trypanocidal activity of prodigiosin, which has been previously demonstrated by our group and others [
26,
35], confirming its effectiveness against the intracellular forms of
T. cruzi.
All the compounds assayed, except for EH123, showed significant toxicity, with all of them being more toxic than benznidazole. EH123 showed low toxicity and low antiparasitic activity. Obatoclax and prodigiosins were particularly toxic, showing nonselective cytotoxic behavior when comparing the cytotoxicity values in both normal and tumor cell lines (mouse L-929 fibroblasts and human hepatocarcinoma cells, respectively) and parasites. On the contrary, MM3 and MM4 appeared to be less toxic than prodigiosins, confirming previous results reported for MM3 by Fiore et al. [
27].
The molecules displaying lower toxicity on mammalian cells were less active against
T. cruzi epimastigotes. Previous observations by Díaz de Greñu et al. demonstrated that the cytotoxicity of these types of molecules was linked to their activity as anionophores [
28]. Our results also suggest that the parasiticidal activity of the compounds is linked, at least partly, to their role as anion carriers. Unlike mammalian cells, which contain multiple mitochondria, the mitochondrion in trypanosomatids appears as a single organelle whose proper function is essential for parasite survival, representing an excellent target for anti-infective chemotherapy [
36]. Mitochondrial disturbances can be indicated by different means, including modifications in cellular respiration.
Our data showed that natural prodigiosin greatly affected cellular oxygen consumption, as demonstrated by the large decrease in the respiration rates of parasites caused by the compound even at low concentrations (73% at nonlethal concentrations (½ IC
50)). Our results are consistent with those obtained by Genes et al. [
37], who reported strong parasiticidal activity of the pigment, as well as an inhibitory effect on mitochondrial respiration. In fact, the inhibition induced by the pigment was similar to that obtained with thenoyltrifluoroacetone (TTFA), a classical inhibitor of respiratory function, indicating that its biological effect involves the mitochondria. Thus, the most potent respiratory inhibitor was also the most effective trypanocidal compound. This may also be the mechanism of antibacterial activity of prodigiosin against bacteria that rely only on respiration, such as
Pseudomonas aeruginosa (minimal inhibitory concentration (MIC) of 8 µM), although not the only mechanism since the pigment also kills microorganisms that obtain their energy strictly through fermentation, such as
Enterococcus faecalis (MIC of 2 µM), as well as facultative bacteria such as
Escherichia coli (MIC of 2 µM too) [
26]. Thus, it seems clear that prodigiosin exerts its antibiotic activity through more than one mechanism.
Furthermore, it is noteworthy that synthetic prodigiosin had a minimal effect on mitochondrial respiration at the concentration equivalent to ½ IC50, but a moderate effect at the highest concentration examined (35.18% of inhibition at ¾ IC50). Thus, it was less powerful than natural prodigiosin. This difference between synthetic and natural pigments could have also been due to limitations in the purification protocol, allowing the persistence of other components in the preparations that might also have had some antiparasitic effects.
Regarding the tambjamines, while MM4 had no inhibitory effect on respiration, MM3 elicited respiratory inhibition close to 50% at the maximum concentration measured (¾ IC
50). These values are similar to those obtained with potassium cyanide (KCN) for respiratory rate inhibition [
37], suggesting that MM3 may also act directly on the electron transport chain.
Although it is clear that benznidazole is the most selective compound, its mechanism of toxicity involves the release of highly reactive agents that have potent cytotoxic effects, leading to dermatological, articular, and digestive hypersensitivity reactions, as well as immune thrombocytopenic purpura and other disturbances mediated by antibodies that lead to treatment discontinuation in as much as 20% of cases [
38,
39,
40,
41].
Hypersensitivity and immune complexes appear to play important roles in adverse reactions beyond in vitro cytotoxicity. This makes mandatory the replacement or the use of reduced doses of benznidazole, as well as the application of other effective but less toxic drugs.
The toxicity of prodigiosin is well recognized, representing the main limitation of its use as an antimicrobial agent. Despite this, prodigiosin displays a large SI, especially over the intracellular forms of
T. cruzi, against which prodigiosin had an SI of 50, suggesting its potential as an antichagasic agent. With other approaches, such as the use of nanoformulated preparations, or other pharmaceutical improvements, a dose reduction could be achieved for prodigiosin and other molecules. This would result in a better therapeutic profile. By the moment the bioapplicability of these compounds has been predicted using the web tool SwissADME (
Figures S2–S7). Moreover, since they are chemically unrelated to benznidazole, it is highly unlikely that resistance to prodigiosin and the rest of the studied molecules can affect susceptibility to Benznidazole and vice versa.






{kind=link}
{kind=link}