
Towards the Development of a Female Animal Model of T1DM Using Hyaluronic Acid Nanocoated Cell Transplantation: Refinements and Considerations for Future Protocols

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
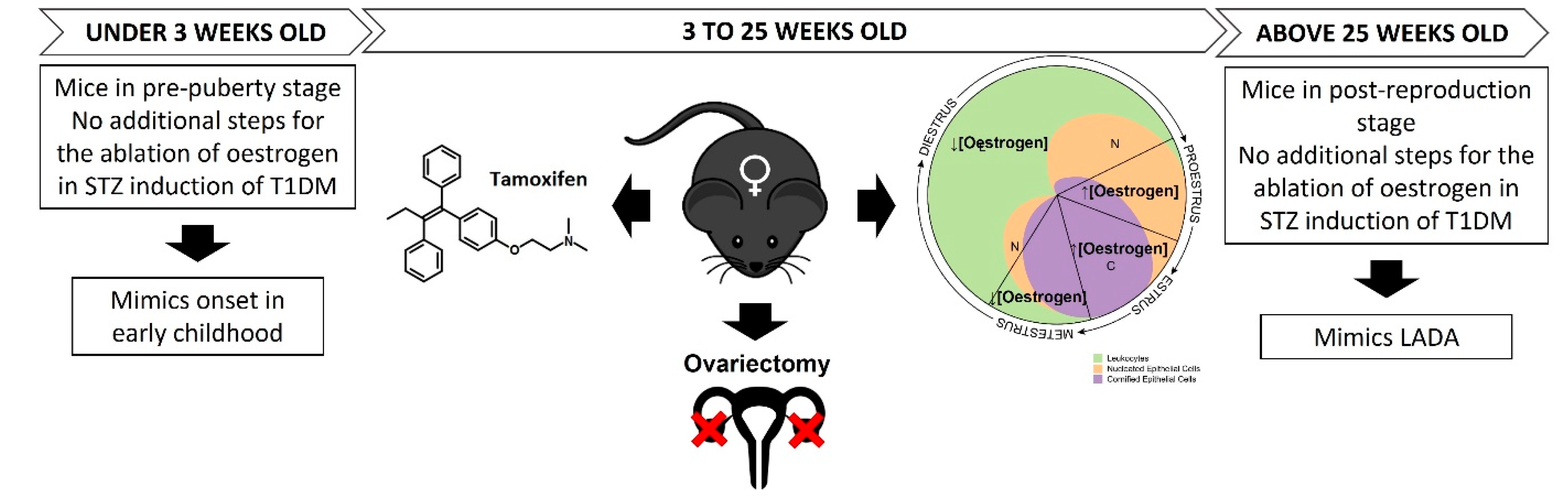
2.3. Model for Diabetes Induction
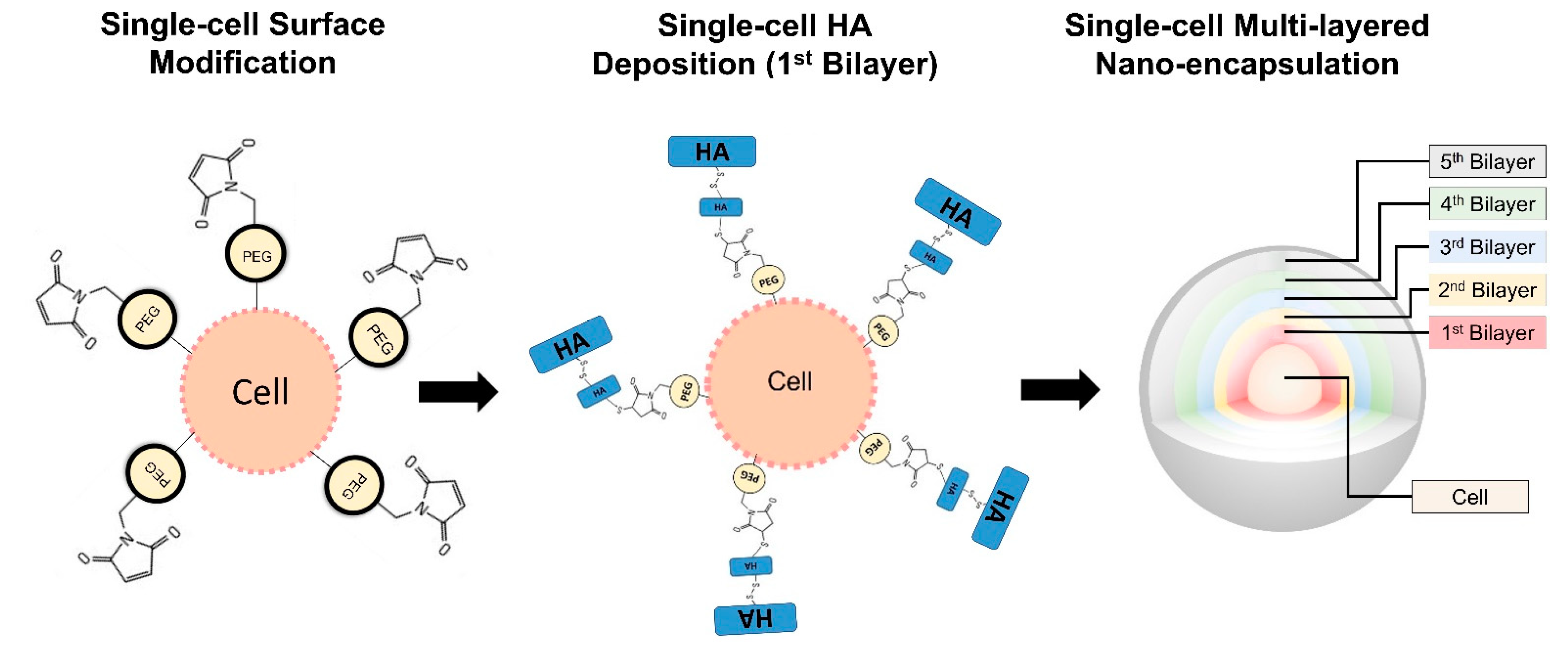
2.4. Conformal Cell Coating
2.5. MIN-6 Graft Transplantation
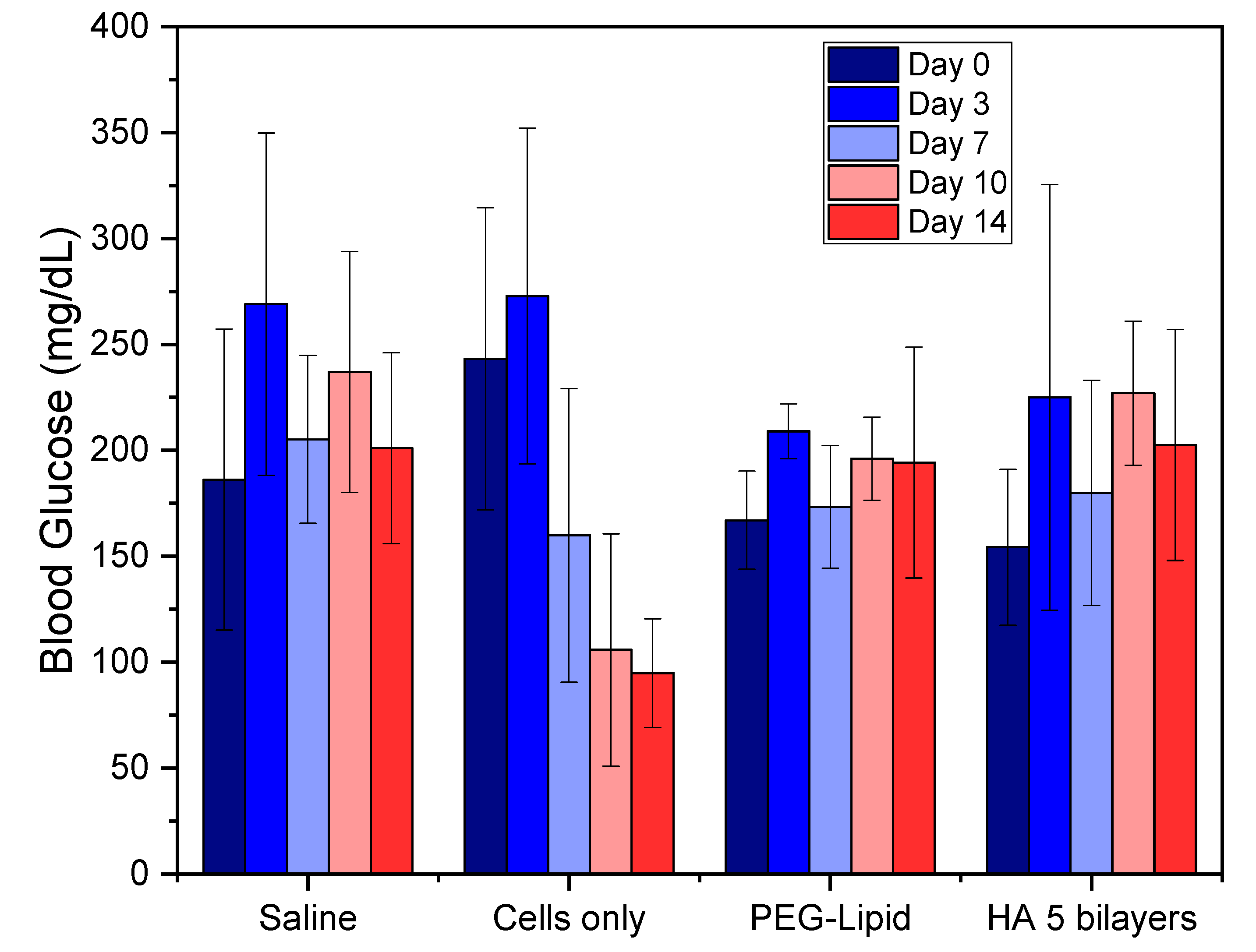
2.6. Blood Glucose Control
2.7. Dissection of Renal and Pancreatic Tissues
2.8. Histology
2.9. Statistical Analysis
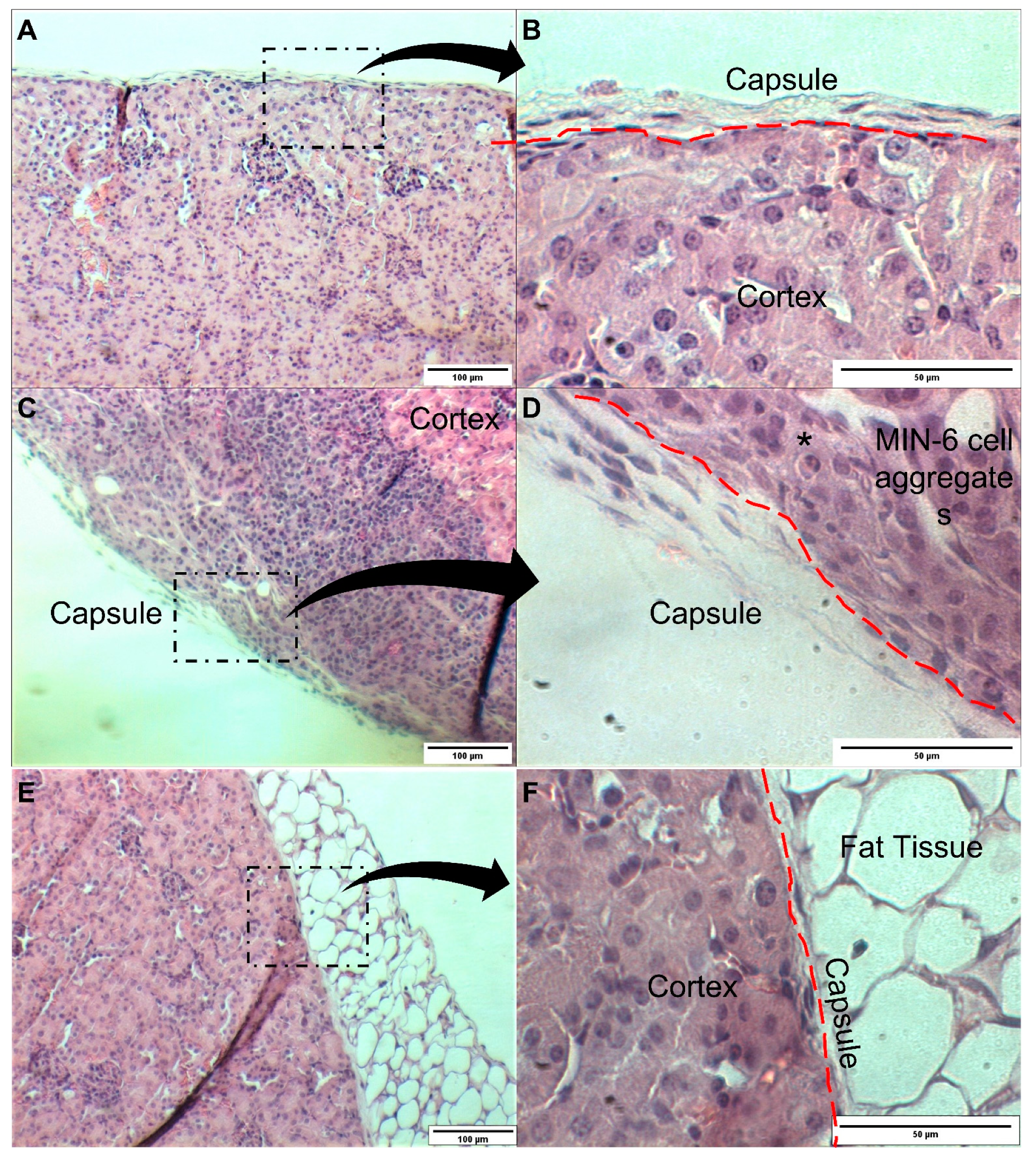
3. Results and Discussion
3.1. Animal Model for Diabetes Induction
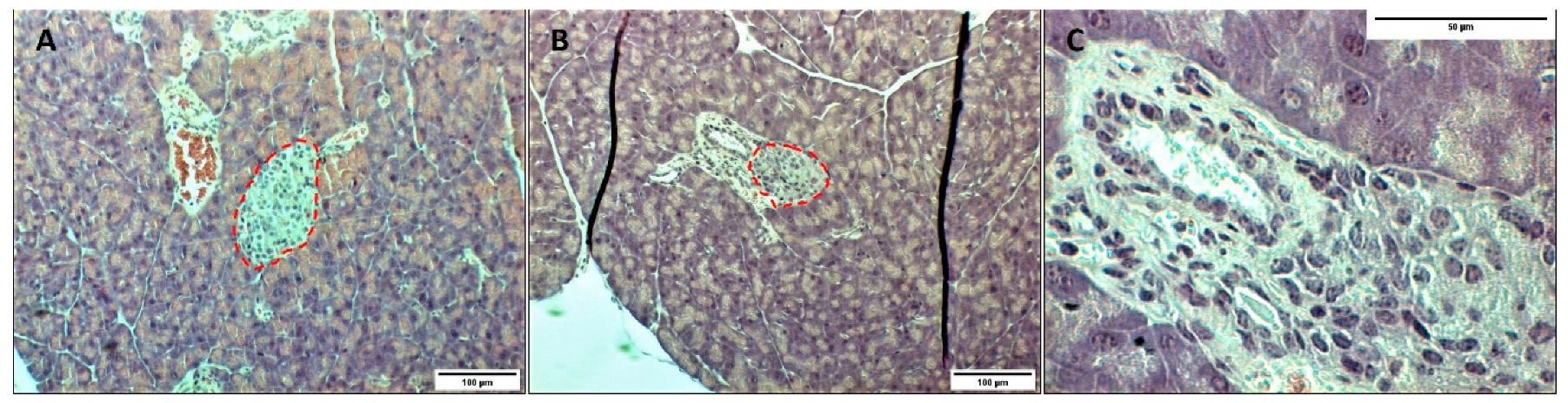
3.2. In Vivo Transplantation in the Renal Subcapsular Space
4. Conclusions
- Increase the daily dose of STZ to above 50 mg/kg.
- If maintaining the 50 mg/kg dose a day, it may be advisable to increase the number of induction days above the 5 consecutive daily injections;
- In order to guarantee complete diabetes induction without β recovery, the post-induction period should be increased from 14 days to 21 days;
- Assess the interference of the oestrous cycle and the levels of oestrogen (E2) in the efficacy of STZ induction of diabetes in female mice aged >3 weeks (post-puberty stage);
- Assess the feasibility of using female mice aged <3 weeks (pre-puberty stage) to induce diabetes using STZ, as levels of oestrogen are neglectable. At this stage, T1DM may be better represented, as in humans, T1DM onset generally occurs in early childhood.
- Development of pseudo-islet aggregates using MIN-6 cells;
- Encapsulation of pseudo-islet;
- Assessment of glucose-induced insulin secretion;
- Transplantation of encapsulated pseudo-islet;
- Histological assessment of pseudo-islet aggregates under the kidney capsule
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Scow, R.O.; Wagner, E.M.; Cardeza, A.D. Effect of hypophysectomy on the insulin requirement and response to fast-ing of totally pancreatectomized rats. Endocrinology 1957, 61, 380–391. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, Z.Z.; Morris, D.L.; Moss, D.R.; Sims, E.K.; Chiong, Y.; Kono, T.; Evans-Molina, C. Streptozotocin is equally diabetogenic whether administered to fed or fasted mice. Lab. Anim. 2013, 47, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Szkudelski, T. The mechanism of alloxan and streptozotocin action in B cells of the rat pancreas. Physiol. Res. 2001, 50, 537–546. [Google Scholar]
- Kinoshita, N.; Echigo, Y.; Shinohara, S.; Gu, Y.; Miyazaki, J.; Inoue, K.; Imamura, M. Regulation of cell proliferation using tissue engineering in MIN6 cells. Cell Transplant. 2001, 10, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Zamboni, F.; Vieira, S.; Reis, R.L.; Oliveira, J.M.; Collins, M.N. The potential of hyaluronic acid in immunoprotection and immunomodulation: Chemistry, processing and function. Prog. Mater. Sci. 2018, 97, 97–122. [Google Scholar] [CrossRef]
- Zamboni, F.; Collins, M.N. Cell based therapeutics in type 1 diabetes mellitus. Int. J. Pharm. 2017, 521, 346–356. [Google Scholar] [CrossRef]
- Beery, A.K. Inclusion of females does not increase variability in rodent research studies. Curr. Opin. Behav. Sci. 2018, 23, 143–149. [Google Scholar] [CrossRef]
- Clayton, J.A.; Collins, F. Policy: NIH to balance sex in cell and animal studies. Nature 2014, 509, 282. [Google Scholar] [CrossRef]
- Gannon, M.; Kulkarni, R.N.; Tse, H.M.; Mauvais-Jarvis, F. Sex differences underlying pancreatic islet biology and its dysfunction. Mol. Metab. 2018, 15, 82–91. [Google Scholar] [CrossRef]
- Dolensek, J.; Rupnik, M.S.; Stozer, A. Structural similarities and differences between the human and the mouse pan-creas. Islets 2015, 7, e1024405. [Google Scholar] [CrossRef] [Green Version]
- Teramura, Y.; Kaneda, Y.; Iwata, H. Islet-encapsulation in ultra-thin layer-by-layer membranes of poly(vinyl alco-hol) anchored to poly(ethylene glycol)–lipids in the cell membrane. Biomaterials 2007, 28, 4818–4825. [Google Scholar] [CrossRef] [PubMed]
- Zamboni, F.; Okoroafor, C.; Ryan, M.P.; Pembroke, J.T.; Strozyk, M.; Culebras, M.; Collins, M.N. On the bacteriostatic activity of hyaluronic acid composite films. Carbohydr. Polym. 2021, 260, 117803. [Google Scholar] [CrossRef]
- Zamboni, F.; Ryan, E.; Culebras, M.; Collins, M.N. Labile crosslinked hyaluronic acid via urethane formation using bis(β-isocyanatoethyl) disulphide with tuneable physicochemical and immunomodulatory properties. Carbohydr. Polym. 2020, 245, 116501. [Google Scholar] [CrossRef]
- Leiter, E.H. Multiple low-dose streptozotocin-induced hyperglycemia and insulitis in C57BL mice: Influence of inbred background, sex, and thymus. Proc. Natl. Acad. Sci. USA 1982, 79, 630–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandramouli, C.; Reichelt, M.E.; Curl, C.L.; Varma, U.; Bienvenu, L.A.; Koutsifeli, P.; Raaijmakers, A.J.A.; De Blasio, M.; Qin, C.; Jenkins, A.J.; et al. Diastolic dysfunction is more apparent in STZ-induced diabetic female mice, despite less pronounced hyperglycemia. Sci. Rep. 2018, 8, 2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantarelli, E.; Citro, A.; Marzorati, S.; Melzi, R.; Scavini, M.; Piemonti, L. Murine animal models for preclinical islet transplantation: No model fits all (research purposes). Islets 2013, 5, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Paoli, M.; Werstuck, G.H. Role of Estrogen in Type 1 and Type 2 Diabetes Mellitus: A Review of Clinical and Pre-clinical Data. Can. J. Diabetes 2020, 44, 448–452. [Google Scholar] [CrossRef] [PubMed]
- Brust, V.; Schindler, P.M.; Lewejohann, L. Lifetime development of behavioural phenotype in the house mouse (Mus musculus). Front. Zool. 2015, 12, S17. [Google Scholar] [CrossRef] [Green Version]
- Harjutsalo, V.; Sjoberg, L.; Tuomilehto, J. Time trends in the incidence of type 1 diabetes in Finnish children: A co-hort study. Lancet 2008, 371, 1777–1782. [Google Scholar] [CrossRef]
- Souza, V.R.; Mendes, E.; Casaro, M.; Antiorio, A.T.F.B.; Oliveira, F.A.; Ferreira, C.M. Description of Ovariectomy Protocol in Mice. Microb. Environ. Genom. (MEG) 2019, 1916, 303–309. [Google Scholar] [CrossRef]
- Le May, C.; Chu, K.; Hu, M.; Ortega, C.S.; Simpson, E.R.; Korach, K.S.; Tsai, M.-J.; Mauvais-Jatvis, F. Estrogens protect pancreatic β-cells from apoptosis and prevent insulin-deficient diabetes mellitus in mice. Proc. Natl. Acad. Sci. USA 2006, 103, 9232–9237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byers, S.L.; Wiles, M.V.; Dunn, S.L.; Taft, R.A. Mouse Estrous Cycle Identification Tool and Images. PLoS ONE 2012, 7, e35538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caligioni, C.S. Assessing Reproductive Status/Stages in Mice. Curr. Protoc. Neurosci. 2009, 48, A.4I.1–A.4I.8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapustin, J. Latent Autoimmune Diabetes in Adults. J. Nurse Pract. 2008, 4, 681–687. [Google Scholar] [CrossRef]
- Brahmkshatriya, P.P.; Mehta, A.A.; Saboo, B.D.; Goyal, R.K. Characteristics and Prevalence of Latent Autoimmune Diabetes in Adults (LADA). ISRN Pharmacol. 2012, 2012, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaney, M.A.; Kowalewska, J.; Treuting, P.M. 16—Urinary System. In Comparative Anatomy and Histology, 2nd ed.; Treuting, P.M., Dintzis, S.M., Mon-Tine, K.S., Eds.; Academic Press: San Diego, CA, USA, 2018. [Google Scholar]
- La Perle, K.M.D.; Dintzis, S.M. 15—Endocrine System. In Comparative Anatomy and Histology, 2nd ed.; Treuting, P.M., Dintzis, S.M., Montine, K.S., Eds.; Academic Press: San Diego, CA, USA, 2018. [Google Scholar]
- Plesner, A.; Liston, P.; Tan, R.; Korneluk, R.G.; Verchere, C.B. The X-Linked Inhibitor of Apoptosis Protein Enhances Survival of Murine Islet Allografts. Diabetes 2005, 54, 2533–2540. [Google Scholar] [CrossRef] [Green Version]
- Cook, M.J. The Anatomy of the Laboratory Mouse; Academic Press: London, UK; New York, NY, USA, 1965. [Google Scholar]
- Micucci, C.; Orciari, S.; Catalano, A. Hyperglycemia Promotes K-Ras-Induced Lung Tumorigenesis through BASCs Amplification. PLoS ONE 2014, 9, e105550. [Google Scholar] [CrossRef]
- Giraldo, J.; Weaver, J.; Stabler, C. Enhancing Clinical Islet Transplantation through Tissue Engineering Strategies. J. Diabetes Sci. Technol. 2010, 4, 1238–1247. [Google Scholar] [CrossRef] [Green Version]
- Bernard, A.B.; Lin, C.C.; Anseth, K.S. XA microwell cell culture platform for the aggregation of pancreatic beta-cells. Tissue Eng. Part. C Methods 2012, 18, 583–592. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blood Glucose Level | SHAM | Cells Only | Cell Encapsulation | Cell with Surface Modification | Total (%) |
|---|---|---|---|---|---|
| 100–149 mg/dL | 2 | 2 | 2 | 2 | 20 |
| 150–199 mg/dL | 4 | 1 | 3 | 5 | 32 |
| 200–299 mg/dL | 2 | 4 | 3 | 1 | 25 |
| 300–399 mg/dL | 1 | 3 | 2 | 1 | 18 |
| ≥400 mg/dL | 1 | 0 | 0 | 1 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zamboni, F.; Cengiz, I.F.; Barbosa, A.M.; Castro, A.G.; Reis, R.L.; Oliveira, J.M.; Collins, M.N. Towards the Development of a Female Animal Model of T1DM Using Hyaluronic Acid Nanocoated Cell Transplantation: Refinements and Considerations for Future Protocols. Pharmaceutics 2021, 13, 1925. https://doi.org/10.3390/pharmaceutics13111925
Zamboni F, Cengiz IF, Barbosa AM, Castro AG, Reis RL, Oliveira JM, Collins MN. Towards the Development of a Female Animal Model of T1DM Using Hyaluronic Acid Nanocoated Cell Transplantation: Refinements and Considerations for Future Protocols. Pharmaceutics. 2021; 13(11):1925. https://doi.org/10.3390/pharmaceutics13111925
Chicago/Turabian StyleZamboni, Fernanda, Ibrahim F. Cengiz, Ana M. Barbosa, Antonio G. Castro, Rui L. Reis, Joaquim M. Oliveira, and Maurice N. Collins. 2021. "Towards the Development of a Female Animal Model of T1DM Using Hyaluronic Acid Nanocoated Cell Transplantation: Refinements and Considerations for Future Protocols" Pharmaceutics 13, no. 11: 1925. https://doi.org/10.3390/pharmaceutics13111925