
A Non-Nutritive Feeding Intervention Alters the Expression of Efflux Transporters in the Gastrointestinal Tract

 , and
, and 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Transporter Extraction from the Jejunum
2.4. Preparation of Hormone Blood Samples
2.5. Data Analysis and Visualisation
3. Results
3.1. Efflux Transporter Expression
3.1.1. P-Glycoprotein (P-gp)
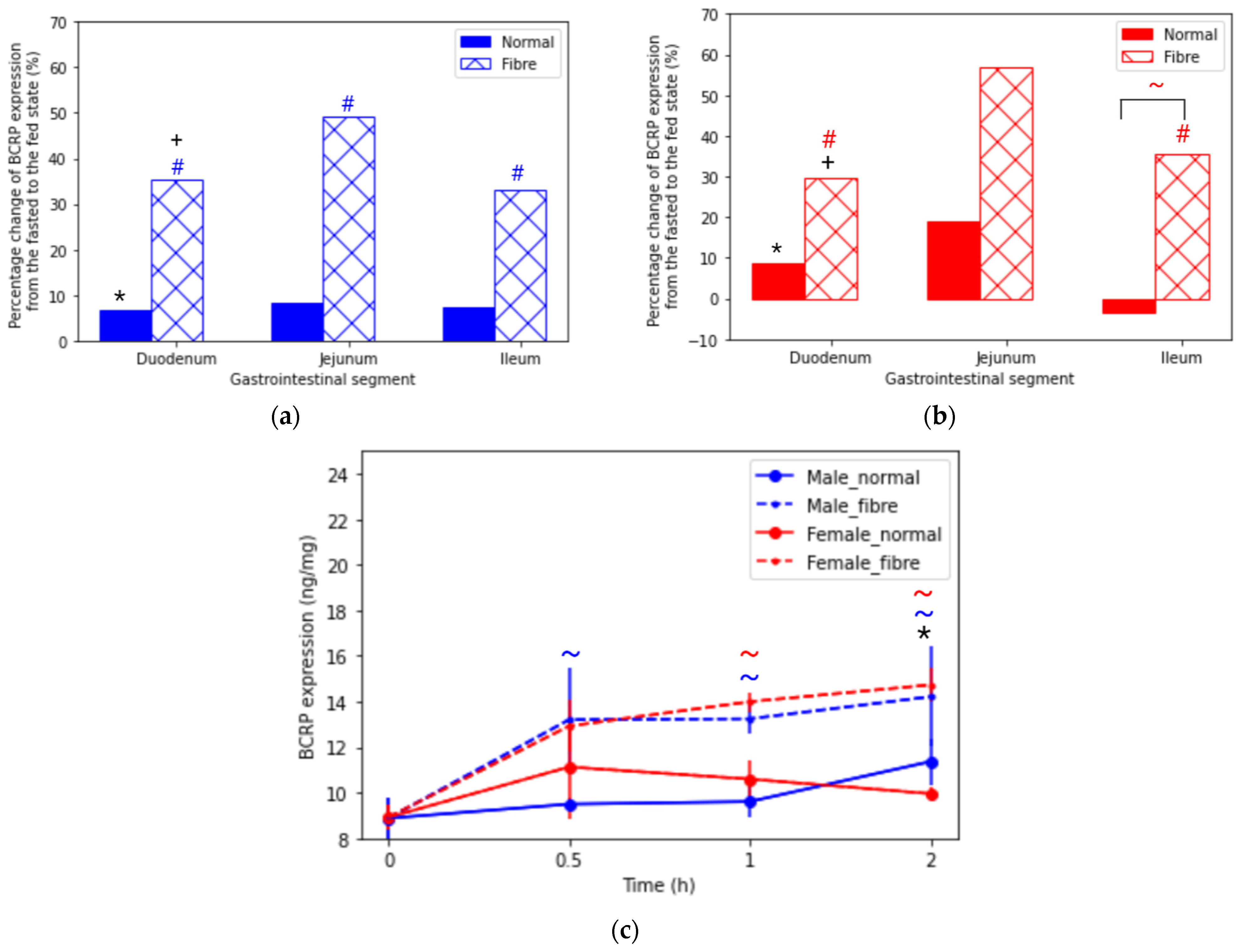
3.1.2. Breast Cancer Resistant Protein (BCRP)
3.1.3. Multidrug-Resistance-Associated Protein 2 (MRP2)
3.2. Hormone Concentration
3.2.1. Gastrointestinal Hormone: Cholecystokinin
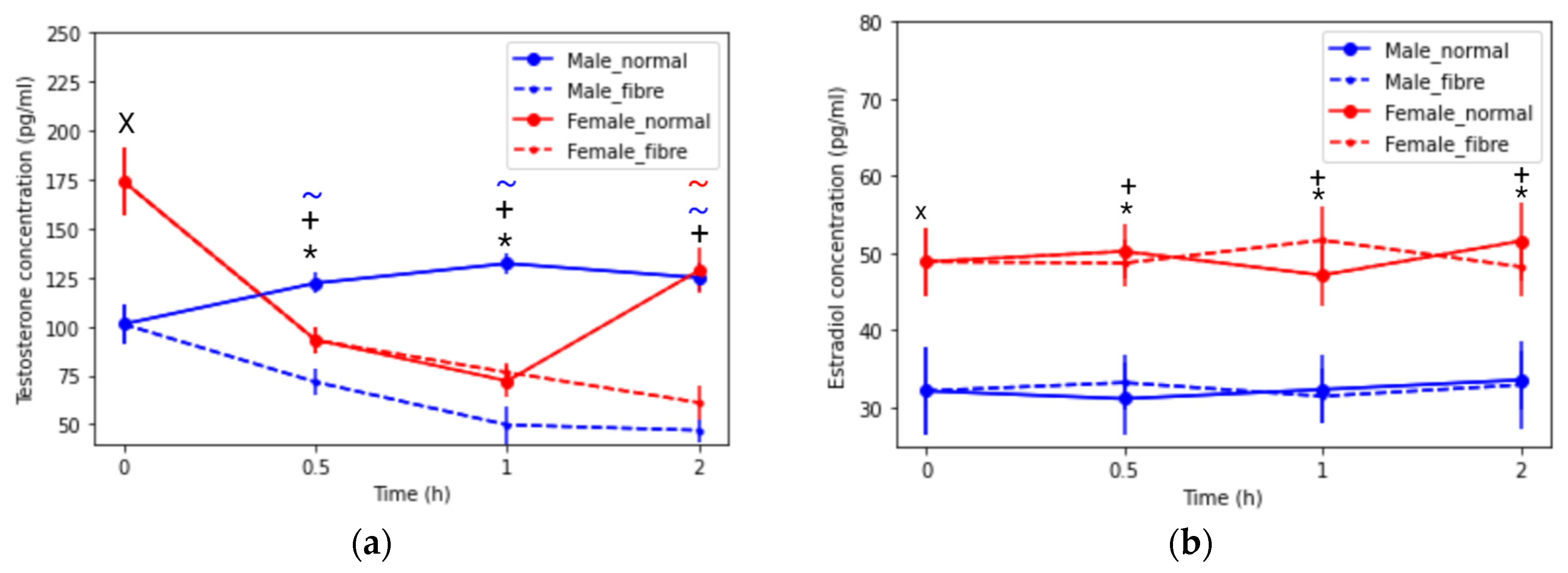
3.2.2. Sex Hormone Concentrations: Testosterone and Oestradiol
3.3. Correlation between Efflux Transporter Expression and Hormone Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Von Erlach, T.; Saxton, S.; Shi, Y.; Minahan, D.; Reker, D.; Javid, F.; Lee, Y.-A.L.; Schoellhammer, C.; Esfandiary, T.; Cleveland, C.; et al. Robotically handled whole-tissue culture system for the screening of oral drug formulations. Nat. Biomed. Eng. 2020, 4, 544–559. [Google Scholar] [CrossRef] [PubMed]
- Vertzoni, M.; Augustijns, P.; Grimm, M.; Koziolek, M.; Lemmens, G.; Parrott, N.; Pentafragka, C.; Reppas, C.; Rubbens, J.; Van Den Alphabeele, J.; et al. Impact of regional differences along the gastrointestinal tract of healthy adults on oral drug absorption: An UNGAP review. Eur. J. Pharm. Sci. 2019, 134, 153–175. [Google Scholar] [CrossRef]
- Stillhart, C.; Vucicevic, K.; Augustijns, P.; Basit, A.W.; Batchelor, H.; Flanagan, T.R.; Gesquiere, I.; Greupink, R.; Keszthelyi, D.; Koskinen, M.; et al. Impact of gastrointestinal physiology on drug absorption in special populations—An UNGAP review. Eur. J. Pharm. Sci. 2020, 147, 105280. [Google Scholar] [CrossRef] [PubMed]
- Elbadawi, M.; McCoubrey, L.E.; Gavins, F.K.H.; Jie Ong, J.; Goyanes, A.; Gaisford, S.; Basit, A.W. Harnessing Artificial Intelligence for the Next Generation of 3D Printed Medicines. Adv. Drug Deliv. Rev. 2021, 175, 113805. [Google Scholar] [CrossRef]
- O’Shea, J.P.; Holm, R.; O’Driscoll, C.M.; Griffin, B.T. Food for thought: Formulating away the food effect—A PEARRL review. J. Pharm. Pharmacol. 2019, 71, 510–535. [Google Scholar] [CrossRef] [Green Version]
- Reker, D.; Shi, Y.; Kirtane, A.R.; Hess, K.; Zhong, G.J.; Crane, E.; Lin, C.-H.; Langer, R.; Traverso, G. Machine Learning Uncovers Food- and Excipient-Drug Interactions. Cell Rep. 2020, 30, 3710–3716.e14. [Google Scholar] [CrossRef]
- Vinarov, Z.; Abrahamsson, B.; Artursson, P.; Batchelor, H.; Berben, P.; Bernkop-Schnürch, A.; Butler, J.; Ceulemans, J.; Davies, N.; Dupont, D.; et al. Current challenges and future perspectives in oral absorption research: An opinion of the UNGAP network. Adv. Drug Deliv. Rev. 2021, 171, 289–331. [Google Scholar] [CrossRef] [PubMed]
- Fadda, H.M.; McConnell, E.L.; Short, M.D.; Basit, A.W. Meal-Induced Acceleration of Tablet Transit Through the Human Small Intestine. Pharm. Res. 2009, 26, 356–360. [Google Scholar] [CrossRef]
- Varum, F.J.O.; Hatton, G.B.; Basit, A.W. Food, physiology and drug delivery. Int. J. Pharm. 2013, 457, 446–460. [Google Scholar] [CrossRef]
- Madla, C.M.; Gavins, F.K.H.; Merchant, H.; Orlu, M.; Murdan, S.; Basit, A.W. Let’s Talk About Sex: Differences in Drug Therapy in Males and Females. Adv. Drug Deliv. Rev. 2021, 175, 113804. [Google Scholar] [CrossRef]
- Kees, F.; Bucher, M.; Schweda, F.; Gschaidmeier, H.; Faerber, L.; Seifert, R. Neoimmun versus Neoral: A bioequivalence study in healthy volunteers and influence of a fat-rich meal on the bioavailability of Neoimmun. Naunyn Schmiedebergs Arch. Pharmacol. 2007, 375, 393–399. [Google Scholar] [CrossRef] [Green Version]
- Hatton, G.B.; Yadav, V.; Basit, A.W.; Merchant, H.A. Animal Farm: Considerations in Animal Gastrointestinal Physiology and Relevance to Drug Delivery in Humans. J. Pharm. Sci. 2015, 104, 2747–2776. [Google Scholar] [CrossRef]
- Sjögren, E.; Abrahamsson, B.; Augustijns, P.; Becker, D.; Bolger, M.B.; Brewster, M.; Brouwers, J.; Flanagan, T.; Harwood, M.; Heinen, C.; et al. In vivo methods for drug absorption—Comparative physiologies, model selection, correlations with in vitro methods (IVIVC), and applications for formulation/API/excipient characterization including food effects. Eur. J. Pharm. Sci. 2014, 57, 99–151. [Google Scholar] [CrossRef] [PubMed]
- Maerz, L.L.; Sankaran, H.; Scharpf, S.J.; Deveney, C.W. Effect of caloric content and composition of a liquid meal on gastric emptying in the rat. Am. J. Physiol. 1994, 267, R1163–R1167. [Google Scholar] [CrossRef] [PubMed]
- Lennernäs, H.; Regårdh, C.-G. Regional gastrointestinal absorption of the beta-blocker pafenolol in the rat and intestinal transit rate determined by movement of 14 C-polyethylene glycol (PEG) 4000. Pharm. Res. 1993, 10, 130–135. [Google Scholar] [CrossRef]
- Mai, Y.; Dou, L.; Yao, Z.; Madla, C.M.; Gavins, F.K.H.; Taherali, F.; Yin, H.; Orlu, M.; Murdan, S.; Basit, A.W. Quantification of P-Glycoprotein in the Gastrointestinal Tract of Humans and Rodents: Methodology, Gut Region, Sex, and Species Matter. Mol. Pharm. 2021, 18, 1895–1904. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Keiser, M.; Drozdzik, M.; Oswald, S. Expression, regulation and function of intestinal drug transporters: An update. Biol. Chem. 2017, 398, 175–192. [Google Scholar] [CrossRef]
- Glavinas, H.; Krajcsi, P.; Cserepes, J.; Sarkadi, B. The role of ABC transporters in drug resistance, metabolism and toxicity. Curr. Drug Deliv. 2004, 1, 27–42. [Google Scholar] [CrossRef]
- Dou, L.; Gavins, F.K.H.; Mai, Y.; Madla, C.M.; Taherali, F.; Orlu, M.; Murdan, S.; Basit, A.W. Effect of Food and an Animal’s Sex on P-Glycoprotein Expression and Luminal Fluids in the Gastrointestinal Tract of Wistar Rats. Pharmaceutics 2020, 12, 296. [Google Scholar] [CrossRef] [Green Version]
- Dou, L.; Mai, Y.; Madla, C.M.; Orlu, M.; Basit, A.W. P-glycoprotein expression in the gastrointestinal tract of male and female rats is influenced differently by food. Eur. J. Pharm. Sci. 2018, 123, 569–575. [Google Scholar] [CrossRef] [Green Version]
- Deferme, S.; Augustijns, P. The effect of food components on the absorption of P-gp substrates: A review. J. Pharm. Pharmacol. 2003, 55, 153–162. [Google Scholar] [CrossRef]
- Gill, S.K.; Rossi, M.; Bajka, B.; Whelan, K. Dietary fibre in gastrointestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Avila, J.A.; Wall-Medrano, A.; Velderrain-Rodríguez, G.R.; Chen, C.O.; Salazar-López, N.J.; Robles-Sánchez, M.; González-Aguilar, G.A. Gastrointestinal interactions, absorption, splanchnic metabolism and pharmacokinetics of orally ingested phenolic compounds. Food Funct. 2017, 8, 15–38. [Google Scholar] [CrossRef]
- Xie, Q.-S.; Zhang, J.-X.; Liu, M.; Liu, P.-H.; Wang, Z.-J.; Zhu, L.; Jiang, L.; Jin, M.-M.; Liu, X.-N.; Liu, L.; et al. Short-chain fatty acids exert opposite effects on the expression and function of p-glycoprotein and breast cancer resistance protein in rat intestine. Acta Pharmacol. Sin. 2021, 42, 470–481. [Google Scholar] [CrossRef] [PubMed]
- Yano, K.; Shimizu, S.; Tomono, T.; Ogihara, T. Gastrointestinal Hormone Cholecystokinin Increases P-Glycoprotein Membrane Localization and Transport Activity in Caco-2 Cells. J. Pharm. Sci. 2017, 106, 2650–2656. [Google Scholar] [CrossRef] [Green Version]
- Wessler, J.D.; Grip, L.T.; Mendell, J.; Giugliano, R.P. The P-Glycoprotein Transport System and Cardiovascular Drugs. J. Am. Coll. Cardiol. 2013, 61, 2495–2502. [Google Scholar] [CrossRef] [Green Version]
- Kanado, Y.; Tsurudome, Y.; Omata, Y.; Yasukochi, S.; Kusunose, N.; Akamine, T.; Matsunaga, N.; Koyanagi, S.; Ohdo, S. Estradiol regulation of P-glycoprotein expression in mouse kidney and human tubular epithelial cells, implication for renal clearance of drugs. Biochem. Biophys. Res. Commun. 2019, 519, 613–619. [Google Scholar] [CrossRef]
- Clarke, L.L. A guide to Ussing chamber studies of mouse intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G1151–G1166. [Google Scholar] [CrossRef] [Green Version]
- Hunter, J.D. Matplotlib: A 2D graphics environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- Drozdzik, M.; Gröer, C.; Penski, J.; Lapczuk, J.; Ostrowski, M.; Lai, Y.; Prasad, B.; Unadkat, J.D.; Siegmund, W.; Oswald, S. Protein Abundance of Clinically Relevant Multidrug Transporters along the Entire Length of the Human Intestine. Mol. Pharm. 2014, 11, 3547–3555. [Google Scholar] [CrossRef] [PubMed]
- Giacomini, K.M.; Huang, S.-M.; Tweedie, D.J.; Benet, L.Z.; Brouwer, K.L.R.; Chu, X.; Dahlin, A.; Evers, R.; Fischer, V.; Hillgren, K.M.; et al. Membrane transporters in drug development. Nat. Rev. Drug Discov. 2010, 9, 215–236. [Google Scholar] [CrossRef]
- Austin Doyle, L.; Ross, D.D. Multidrug resistance mediated by the breast cancer resistance protein BCRP (ABCG2). Oncogene 2003, 22, 7340–7358. [Google Scholar] [CrossRef] [Green Version]
- FDA. Guidance for Industry: Food-Effect Bioavailability and Fed Bioequivalence Studies. Available online: https://www.fda.gov/files/drugs/published/Food-Effect-Bioavailability-and-Fed-Bioequivalence-Studies.pdf (accessed on 14 August 2020).
- Henze, L.J.; Koehl, N.J.; O’Shea, J.P.; Holm, R.; Vertzoni, M.; Griffin, B.T. Toward the establishment of a standardized pre-clinical porcine model to predict food effects—Case studies on fenofibrate and paracetamol. Int. J. Pharm. X 2019, 1, 100017. [Google Scholar] [CrossRef]
- Lentz, K.A.; Quitko, M.; Morgan, D.G.; Grace, J.E., Jr.; Gleason, C.; Marathe, P.H. Development and validation of a preclinical food effect model. J. Pharm. Sci. 2007, 96, 459–472. [Google Scholar] [CrossRef] [PubMed]
- MacLean, C.; Moenning, U.; Reichel, A.; Fricker, G. Closing the Gaps: A Full Scan of the Intestinal Expression of P-Glycoprotein, Breast Cancer Resistance Protein, and Multidrug Resistance-Associated Protein 2 in Male and Female Rats. Drug Metab. Dispos. 2008, 36, 1249. [Google Scholar] [CrossRef] [Green Version]
- Dahan, A.; Amidon, G.L. Segmental Dependent Transport of Low Permeability Compounds along the Small Intestine Due to P-Glycoprotein: The Role of Efflux Transport in the Oral Absorption of BCS Class III Drugs. Mol. Pharm. 2009, 6, 19–28. [Google Scholar] [CrossRef]
- Domínguez, C.J.; Tocchetti, G.N.; Rigalli, J.P.; Mottino, A.D. Acute regulation of apical ABC transporters in the gut. Potential influence on drug bioavailability. Pharmacol. Res. 2020, 163, 105251. [Google Scholar] [CrossRef]
- Schinkel, A.H.; Jonker, J.W. Mammalian drug efflux transporters of the ATP binding cassette (ABC) family: An overview. Adv. Drug Deliv. Rev. 2003, 55, 3–29. [Google Scholar] [CrossRef]
- Nguyen, T.-T.-L.; Duong, V.-A.; Maeng, H.-J. Pharmaceutical Formulations with P-Glycoprotein Inhibitory Effect as Promising Approaches for Enhancing Oral Drug Absorption and Bioavailability. Pharmaceutics 2021, 13, 1103. [Google Scholar] [CrossRef] [PubMed]
- Robey, R.W.; Pluchino, K.M.; Hall, M.D.; Fojo, A.T.; Bates, S.E.; Gottesman, M.M. Revisiting the role of ABC transporters in multidrug-resistant cancer. Nat. Rev. Cancer 2018, 18, 452–464. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Sugiyama, Y. Role of metabolic enzymes and efflux transporters in the absorption of drugs from the small intestine. Eur. J. Pharm. Sci. 2000, 12, 3–12. [Google Scholar] [CrossRef]
- Lemos, C.; Jansen, G.; Peters, G.J. Drug transporters: Recent advances concerning BCRP and tyrosine kinase inhibitors. Br. J. Cancer 2008, 98, 857–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smolinski, M.P.; Urgaonkar, S.; Pitzonka, L.; Cutler, M.; Lee, G.; Suh, K.H.; Lau, J.Y.N. Discovery of Encequidar, First-in-Class Intestine Specific P-glycoprotein Inhibitor. J. Med. Chem. 2021, 64, 3677–3693. [Google Scholar] [CrossRef]
- Kissileff, H.R.; Carretta, J.C.; Geliebter, A.; Pi-Sunyer, F.X. Cholecystokinin and stomach distension combine to reduce food intake in humans. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2003, 285, R992–R998. [Google Scholar] [CrossRef]
- Karhunen, L.J.; Juvonen, K.R.; Huotari, A.; Purhonen, A.K.; Herzig, K.H. Effect of protein, fat, carbohydrate and fibre on gastrointestinal peptide release in humans. Regul. Pept. 2008, 149, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Arias, A.; Rigalli, J.P.; Villanueva, S.S.; Ruiz, M.L.; Luquita, M.G.; Perdomo, V.G.; Vore, M.; Catania, V.A.; Mottino, A.D. Regulation of expression and activity of multidrug resistance proteins MRP2 and MDR1 by estrogenic compounds in Caco-2 cells. Role in prevention of xenobiotic-induced cytotoxicity. Toxicology 2014, 320, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Shchul’kin, A.V.; Yakusheva, E.N.; Chernykh, I.V.; Nikiforov, A.A.; Popova, N.P. Effects of Testosterone on the Functional Activity of P-Glycoprotein. Pharm. Chem. J. 2017, 51, 743–747. [Google Scholar] [CrossRef]
- Janssen, S.; Depoortere, I. Nutrient sensing in the gut: New roads to therapeutics? Trends Endocrinol. Metab. 2013, 24, 92–100. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mai, Y.; Gavins, F.K.H.; Dou, L.; Liu, J.; Taherali, F.; Alkahtani, M.E.; Murdan, S.; Basit, A.W.; Orlu, M. A Non-Nutritive Feeding Intervention Alters the Expression of Efflux Transporters in the Gastrointestinal Tract. Pharmaceutics 2021, 13, 1789. https://doi.org/10.3390/pharmaceutics13111789
Mai Y, Gavins FKH, Dou L, Liu J, Taherali F, Alkahtani ME, Murdan S, Basit AW, Orlu M. A Non-Nutritive Feeding Intervention Alters the Expression of Efflux Transporters in the Gastrointestinal Tract. Pharmaceutics. 2021; 13(11):1789. https://doi.org/10.3390/pharmaceutics13111789
Chicago/Turabian StyleMai, Yang, Francesca K. H. Gavins, Liu Dou, Jing Liu, Farhan Taherali, Manal E. Alkahtani, Sudaxshina Murdan, Abdul W. Basit, and Mine Orlu. 2021. "A Non-Nutritive Feeding Intervention Alters the Expression of Efflux Transporters in the Gastrointestinal Tract" Pharmaceutics 13, no. 11: 1789. https://doi.org/10.3390/pharmaceutics13111789