Can Implementation of Genetics and Pharmacogenomics Improve Treatment of Chronic Low Back Pain?


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Genes of Interest
3.1. Receptors
3.1.1. OPRM1 (Opioid Receptor Mu 1)
3.1.2. HTR2A (5-Hydroxytryptamine Receptor 2A)
3.1.3. DCC (Deleted in Colorectal Carcinoma)
3.1.4. ESR (Estrogen Receptor 1)
3.1.5. CNR2 (Cannabinoid Receptor 2)
3.1.6. ADRB2 (Adrenoceptor Beta 2)
3.2. Enzymes
3.2.1. COMT (Catechol-O-Methyltransferase)
3.2.2. CASP9 (Caspase-9)
3.2.3. GCH1 (GTP Cyclohydrolase 1)
3.2.4. MMP 1,2,3 (Matrix Metallopeptidases)
3.2.5. FAAH (Fatty Acid Amide Hydrolase)
3.3. Cytokines and Associated Receptors
3.3.1. IL18RAP (Interleukin 18 Receptor Accessory Protein); IL18R1 (Interleukin 18 Receptor 1); IL1A (Interleukin 1 Alpha)
3.3.2. GDF5 (Growth Differentiation Factor 5)
3.3.3. CCL2 (C-C Motif Chemokine Ligand 2)
3.4. Transcription Factors
3.4.1. SOX5 (SRY-Box 5)
3.4.2. CCDC26/GSDMC (CCDC26 Long Non-Coding RNA/Gasdermin C)
3.4.3. PNOC (Prepronociceptin)
3.5. Pharmacogenomics in Management of cLBP
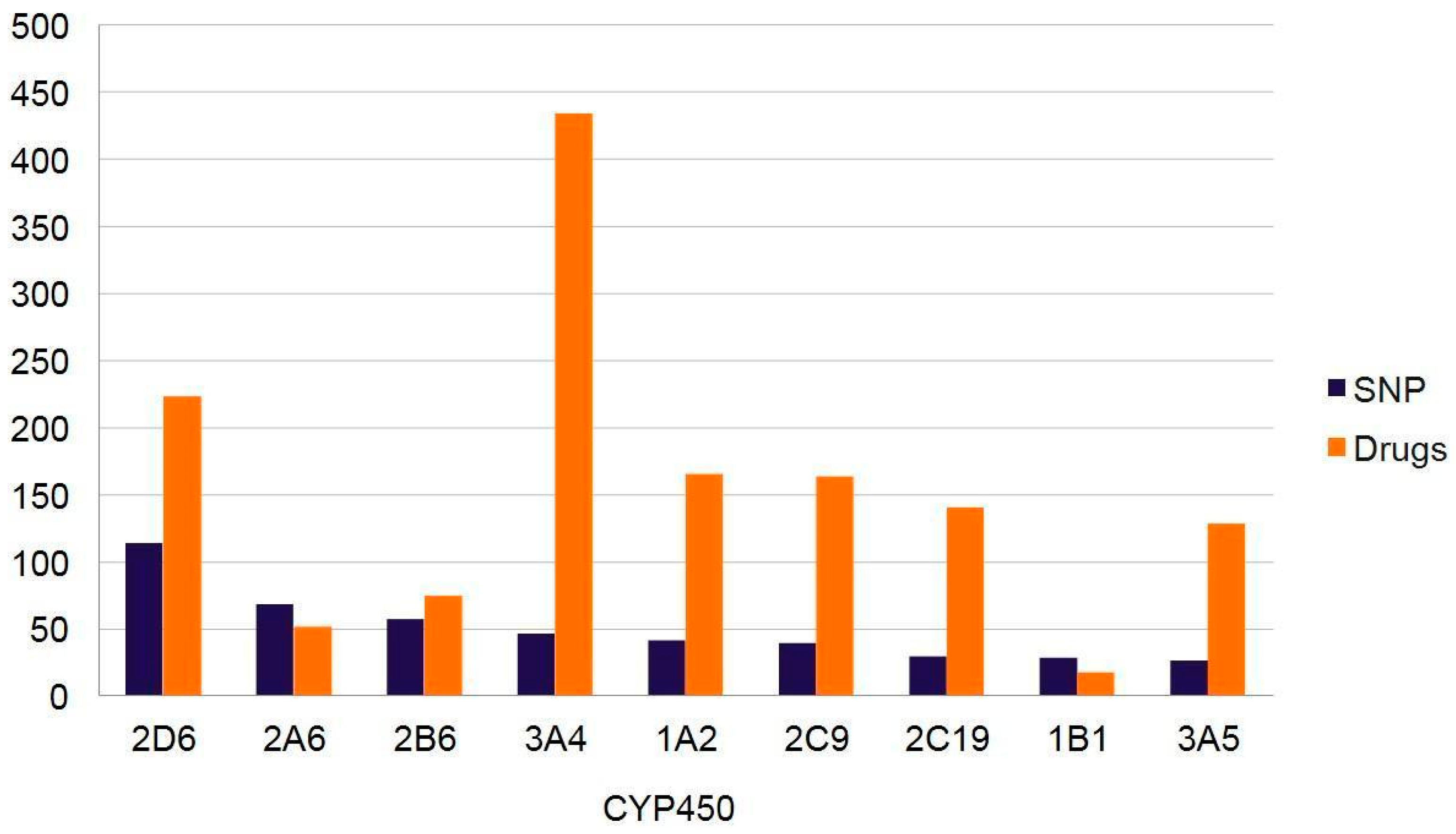
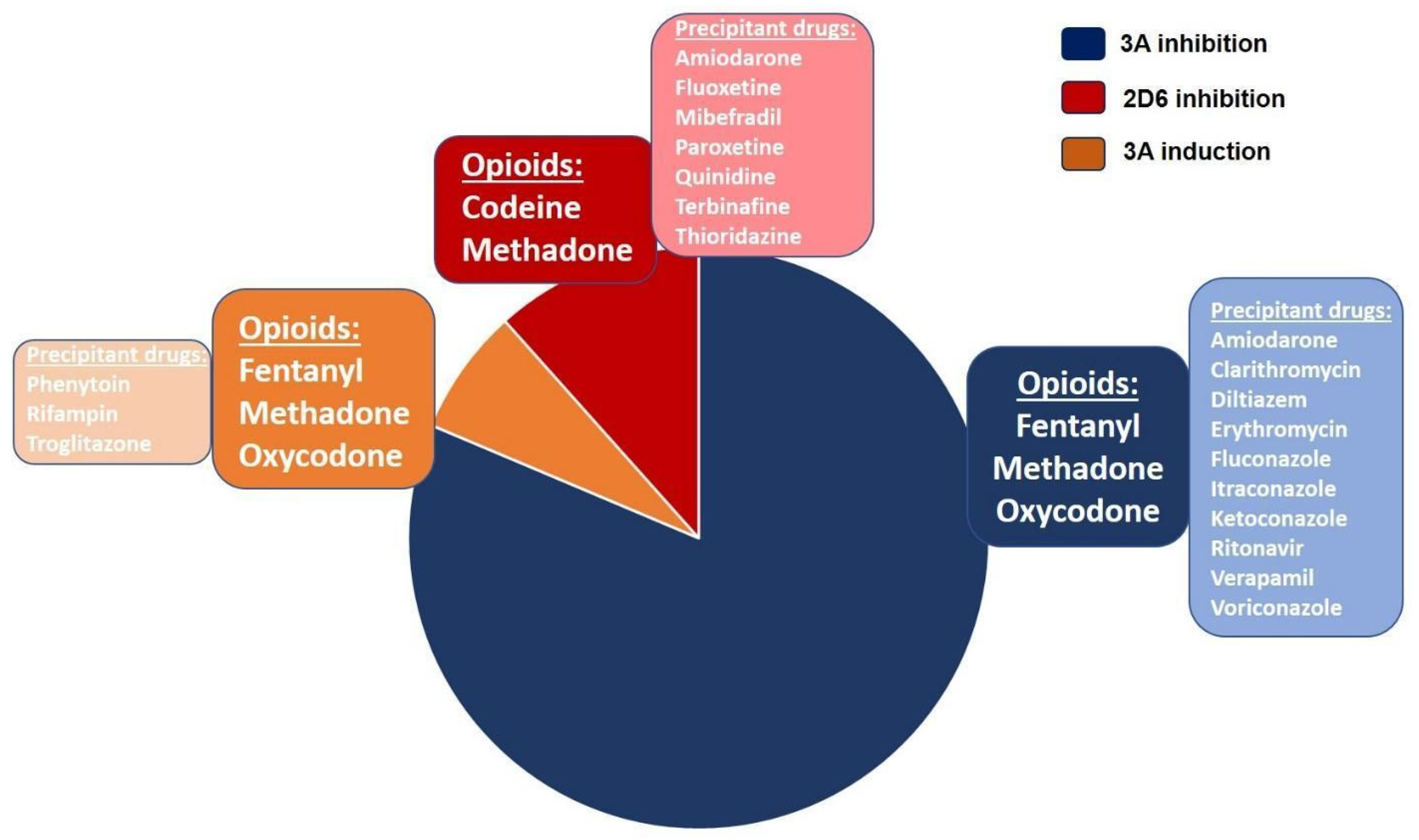
3.5.1. Drug–Drug Interactions
3.5.2. Drug–Drug–Gene Interactions
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Manchikanti, L.; Singh, V.; Falco, F.J.; Benyamin, R.M.; Hirsch, J.A. Epidemiology of low back pain in adults. Neuromodulation 2014, 17 (Suppl. 2), 3–10. [Google Scholar] [CrossRef] [PubMed]
- Shmagel, A.; Foley, R.; Ibrahim, H. Epidemiology of Chronic Low Back Pain in US Adults: Data from the 2009-2010 National Health and Nutrition Examination Survey. Arthritis Care Res. 2016, 68, 1688–1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polderman, T.J.; Benyamin, B.; de Leeuw, C.A.; Sullivan, P.F.; Van Bochoven, A.; Visscher, P.M.; Posthuma, D. Meta-analysis of the heritability of human traits based on fifty years of twin studies. Nat. Genet. 2015, 47, 702–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, P.H.; Beckenkamp, P.; Maher, C.G.; Hopper, J.L.; Ferreira, M.L. Nature or nurture in low back pain? Results of a systematic review of studies based on twin samples. Eur. J. Pain 2013, 17, 957–971. [Google Scholar] [CrossRef]
- Battie, M.C.; Videman, T.; Levalahti, E.; Gill, K.; Kaprio, J. Heritability of low back pain and the role of disc degeneration. Pain 2007, 131, 272–280. [Google Scholar] [CrossRef]
- Rodriguez-Raecke, R.; Niemeier, A.; Ihle, K.; Ruether, W.; May, A. Structural brain changes in chronic pain reflect probably neither damage nor atrophy. PLoS ONE 2013, 8, e54475. [Google Scholar] [CrossRef] [Green Version]
- Baliki, M.N.; Petre, B.; Torbey, S.; Herrmann, K.M.; Huang, L.; Schnitzer, T.J.; Fields, H.L.; Apkarian, A.V. Corticostriatal functional connectivity predicts transition to chronic back pain. Nat. Neurosci. 2012, 15, 1117–1119. [Google Scholar] [CrossRef] [Green Version]
- Seminowicz, D.A.; Wideman, T.H.; Naso, L.; Hatami-Khoroushahi, Z.; Fallatah, S.; Ware, M.A.; Jarzem, P.; Bushnell, M.C.; Shir, Y.; Ouellet, J.A.; et al. Effective treatment of chronic low back pain in humans reverses abnormal brain anatomy and function. J. Neurosci. 2011, 31, 7540–7550. [Google Scholar] [CrossRef] [Green Version]
- Zorina-Lichtenwalter, K.; Meloto, C.B.; Khoury, S.; Diatchenko, L. Genetic predictors of human chronic pain conditions. Neuroscience 2016, 338, 36–62. [Google Scholar] [CrossRef] [Green Version]
- Hasvik, E.; Iordanova Schistad, E.; Grovle, L.; Julsrud Haugen, A.; Roe, C.; Gjerstad, J. Subjective health complaints in patients with lumbar radicular pain and disc herniation are associated with a sex—OPRM1 A118G polymorphism interaction: A prospective 1-year observational study. BMC Musculoskelet. Disord. 2014, 15, 161. [Google Scholar] [CrossRef]
- Grovle, L.; Haugen, A.J.; Ihlebaek, C.M.; Keller, A.; Natvig, B.; Brox, J.I.; Grotle, M. Comorbid subjective health complaints in patients with sciatica: A prospective study including comparison with the general population. J. Psychosom. Res. 2011, 70, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Zubieta, J.K.; Dannals, R.F.; Frost, J.J. Gender and age influences on human brain mu-opioid receptor binding measured by PET. Am. J. Psychiatry 1999, 156, 842–848. [Google Scholar] [CrossRef] [PubMed]
- Ray, R.; Ruparel, K.; Newberg, A.; Wileyto, E.P.; Loughead, J.W.; Divgi, C.; Blendy, J.A.; Logan, J.; Zubieta, J.-K.; Lerman, C. Human Mu Opioid Receptor (OPRM1 A118G) polymorphism is associated with brain mu-opioid receptor binding potential in smokers. Proc. Natl. Acad. Sci. USA 2011, 108, 9268–9273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsunaga, M.; Isowa, T.; Murakami, H.; Kasugai, K.; Yoneda, M.; Kaneko, H.; Ohira, H. Association of polymorphism in the human mu-opioid receptor OPRM1 gene with proinflammatory cytokine levels and health perception. Brain Behav. Immun. 2009, 23, 931–935. [Google Scholar] [CrossRef] [PubMed]
- Way, B.M.; Taylor, S.E.; Eisenberger, N.I. Variation in the mu-opioid receptor gene (OPRM1) is associated with dispositional and neural sensitivity to social rejection. Proc. Natl. Acad. Sci. USA 2009, 106, 15079–15084. [Google Scholar] [CrossRef] [Green Version]
- Milner, T.A.; Burstein, S.R.; Marrone, G.F.; Khalid, S.; Gonzalez, A.D.; Williams, T.J.; Schierberl, K.C.; Torres-Reveron, A.; Gonzales, K.L.; McEwen, B.S.; et al. Stress differentially alters mu opioid receptor density and trafficking in parvalbumin-containing interneurons in the female and male rat hippocampus. Synapse 2013, 67, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Nicholl, B.I.; Holliday, K.L.; Macfarlane, G.J.; Thomson, W.; Davies, K.A.; O’Neill, T.; Bartfai, G.; Boonen, S.; Casanueva, F.F.; Finn, J.D.; et al. Association of HTR2A polymorphisms with chronic widespread pain and the extent of musculoskeletal pain: Results from two population-based cohorts. Arthritis Rheum. 2011, 63, 810–818. [Google Scholar] [CrossRef] [Green Version]
- Yildiz, S.H.; Ulasli, A.M.; Ozdemir Erdogan, M.; Dikici, Ö.; Terzi, E.S.A.; Dündar, Ü.; Solak, M. Assessment of Pain Sensitivity in Patients with Chronic Low Back Pain and Association with HTR2A Gene Polymorphism. Arch. Rheumatol. 2017, 32, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Kato, K.; Kikuchi, S.; Konno, S.; Sekiguchi, M. Participation of 5-hydroxytryptamine in pain-related behavior induced by nucleus pulposus applied on the nerve root in rats. Spine 2008, 33, 1330–1336. [Google Scholar] [CrossRef]
- Kanayama, M.; Hashimoto, T.; Shigenobu, K.; Yamane, S. Efficacy of serotonin receptor blocker for symptomatic lumbar disc herniation. Clin. Orthop. Relat. Res. 2003, 411, 159–165. [Google Scholar] [CrossRef]
- Suri, P.; Palmer, M.R.; Tsepilov, Y.A.; Freidin, M.B.; Boer, C.G.; Yau, M.S.; Evans, D.S.; Gelemanović, A.; Bartz, T.M.; Nethander, M.; et al. Genome-wide meta-analysis of 158,000 individuals of European ancestry identifies three loci associated with chronic back pain. PLoS Genet. 2018, 14, e1007601. [Google Scholar] [CrossRef] [Green Version]
- Finci, L.; Zhang, Y.; Meijers, R.; Wang, J.H. Signaling mechanism of the netrin-1 receptor DCC in axon guidance. Prog. Biophys. Mol. Biol. 2015, 118, 153–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.H.; Yuan, X.C.; Gao, F.; Li, H.-P.; Cao, J.; Liu, Y.-S.; Yu, W.; Tian, B.; Meng, X.-F.; Shi, J.; et al. Netrin-1 Contributes to Myelinated Afferent Fiber Sprouting and Neuropathic Pain. Mol. Neurobiol. 2016, 53, 5640–5651. [Google Scholar] [CrossRef] [PubMed]
- Dun, X.P.; Parkinson, D.B. Role of Netrin-1 Signaling in Nerve Regeneration. Int. J. Mol. Sci. 2017, 18, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu, G.; Hou, S.; Ren, D.; Wu, Y.; Shang, W.; Huang, W. Increased expression of netrin-1 and its deleted in colorectal cancer receptor in human diseased lumbar intervertebral disc compared with autopsy control. Spine 2012, 37, 2074–2081. [Google Scholar] [CrossRef]
- Freemont, A.J.; Peacock, T.E.; Goupille, P.; Hoyland, J.A.; O’Brien, J.; Jayson, M.I. Nerve ingrowth into diseased intervertebral disc in chronic back pain. Lancet 1997, 350, 178–181. [Google Scholar] [CrossRef]
- Pinheiro, M.B.; Ferreira, M.L.; Refshauge, K.; Colodro-Conde, L.; Carrillo, E.; Hopper, J.L.; Ordoñana, J.R.; Ferreira, P.H. Genetics and the environment affect the relationship between depression and low back pain: A co-twin control study of Spanish twins. Pain 2015, 156, 496–503. [Google Scholar] [CrossRef] [Green Version]
- Okbay, A.; LifeLines Cohort Study; Baselmans, B.M.; De Neve, J.E.; Turley, P.; Nivard, M.G.; Fontana, M.A.; Meddens, S.F.W.; Linnér, R.K.; Rietveld, C.A.; et al. Genetic variants associated with subjective well-being, depressive symptoms, and neuroticism identified through genome-wide analyses. Nat. Genet. 2016, 48, 624–633. [Google Scholar] [CrossRef] [Green Version]
- Roh, H.L.; Lee, J.S.; Suh, K.T.; Kim, J.I.; Lee, H.S.; Goh, T.S.; Park, S.H. Association between estrogen receptor gene polymorphism and back pain intensity in female patients with degenerative lumbar spondylolisthesis. J. Spinal Disord. Tech. 2013, 26, E53–E57. [Google Scholar] [CrossRef]
- Lindberg, M.K.; Alatalo, S.L.; Halleen, J.M.; Mohan, S.; Gustafsson, J.A.; Ohlsson, C. Estrogen receptor specificity in the regulation of the skeleton in female mice. J. Endocrinol. 2001, 171, 229–236. [Google Scholar] [CrossRef] [Green Version]
- Vidal, O.; Lindberg, M.K.; Hollberg, K.; Baylink, D.J.; Andersson, G.; Lubahn, D.B.; Mohan, S.; Gustafsson, J.-Å.; Ohlsson, C. Estrogen receptor specificity in the regulation of skeletal growth and maturation in male mice. Proc. Natl. Acad. Sci. USA 2000, 97, 5474–5479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ushiyama, T.; Ueyama, H.; Inoue, K.; Nishioka, J.; Ohkubo, I.; Hukuda, S. Estrogen receptor gene polymorphism and generalized osteoarthritis. J. Rheumatol. 1998, 25, 134–137. [Google Scholar] [PubMed]
- Racz, I.; Nadal, X.; Alferink, J.; Banos, J.E.; Rehnelt, J.; Martin, M.; Pintado, B.; Gutierrez-Adan, A.; Sanguino, E.; Manzanares, J.; et al. Crucial role of CB(2) cannabinoid receptor in the regulation of central immune responses during neuropathic pain. J. Neurosci. 2008, 28, 12125–12135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramesh, D.; D’Agata, A.; Starkweather, A.R.; Young, E.E. Contribution of Endocannabinoid Gene Expression and Genotype on Low Back Pain Susceptibility and Chronicity. Clin. J. Pain 2018, 34, 8–14. [Google Scholar] [CrossRef]
- Skouen, J.S.; Smith, A.J.; Warrington, N.M.; O’Sullivan, P.; McKenzie, L.; Pennell, C.E.; Straker, L. Genetic variation in the beta-2 adrenergic receptor is associated with chronic musculoskeletal complaints in adolescents. Eur. J. Pain 2012, 16, 1232–1242. [Google Scholar] [CrossRef]
- Diatchenko, L.; Anderson, A.D.; Slade, G.D.; Fillingim, R.B.; Shabalina, S.A.; Higgins, T.J.; Sama, S.; Belfer, I.; Goldman, D.; Max, M.B.; et al. Three major haplotypes of the beta2 adrenergic receptor define psychological profile, blood pressure, and the risk for development of a common musculoskeletal pain disorder. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2006, 141B, 449–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas-Alarcon, G.; Fragoso, J.M.; Cruz-Robles, D.; Vargas, A.; Martinez, A.; Lao, J.I.; Garcia-Fructuoso, F.; Vallejo, M.; Martínez-Lavín, M. Association of adrenergic receptor gene polymorphisms with different fibromyalgia syndrome domains. Arthritis Rheum. 2009, 60, 2169–2173. [Google Scholar] [CrossRef]
- Starkweather, A.R.; Ramesh, D.; Lyon, D.E.; Siangphoe, U.; Deng, X.; Sturgill, J.; Heineman, A.; Elswick, R.K., Jr.; Dorsey, S.G.; Greenspan, J.; et al. Acute Low Back Pain: Differential Somatosensory Function and Gene Expression Compared with Healthy No-Pain Controls. Clin. J. Pain 2016, 32, 933–939. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Tang, Y.; Xie, M.; Bie, B.; Wu, J.; Yang, H.; Foss, J.; Yang, B.; Rosenquist, R.W.; Naguib, M. Activation of cannabinoid receptor 2 attenuates mechanical allodynia and neuroinflammatory responses in a chronic post-ischemic pain model of complex regional pain syndrome type I in rats. Eur. J. Neurosci. 2016, 44, 3046–3055. [Google Scholar] [CrossRef]
- Omair, A.; Mannion, A.F.; Holden, M.; Fairbank, J.; Lie, B.A.; Hägg, O.; Fritzell, P.; Brox, J.I. Catechol-O-methyltransferase (COMT) gene polymorphisms are associated with baseline disability but not long-term treatment outcome in patients with chronic low back pain. Eur. Spine J. 2015, 24, 2425–2431. [Google Scholar] [CrossRef]
- Nackley, A.G.; Tan, K.S.; Fecho, K.; Flood, P.; Diatchenko, L.; Maixner, W. Catechol-O-methyltransferase inhibition increases pain sensitivity through activation of both beta2- and beta3-adrenergic receptors. Pain 2007, 128, 199–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsen, L.M.; Eriksen, G.S.; Pedersen, L.M.; Gjerstad, J. Catechol-O-methyltransferase (COMT) inhibition reduces spinal nociceptive activity. Neurosci. Lett. 2010, 473, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Zubieta, J.K.; Heitzeg, M.M.; Smith, Y.R.; Bueller, J.A.; Xu, K.; Koeppe, R.A.; Stohler, C.S.; Goldman, D. COMT val158met genotype affects mu-opioid neurotransmitter responses to a pain stressor. Science 2003, 299, 1240–1243. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, L.M.; Schistad, E.I.; Storesund, A.; Pedersen, L.; Rygh, L.; Røe, C.; Gjerstad, J. The COMT rs4680 Met allele contributes to long-lasting low back pain, sciatica and disability after lumbar disc herniation. Eur. J. Pain 2012, 16, 1064–1069. [Google Scholar] [CrossRef] [PubMed]
- Rut, M.; Machoy-Mokrzynska, A.; Reclawowicz, D.; Słoniewski, P.; Kurzawski, M.; Droździk, M.; Safranow, K.; Morawska, M.; Białecka, M. Influence of variation in the catechol-O-methyltransferase gene on the clinical outcome after lumbar spine surgery for one-level symptomatic disc disease: A report on 176 cases. Acta Neurochir. 2014, 156, 245–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, T.M.; Liu, M.; Zhang, Y.G.; Guo, W.T.; Wu, S.X. Association between Caspase-9 promoter region polymorphisms and discogenic low back pain. Connect. Tissue Res. 2011, 52, 133–138. [Google Scholar] [CrossRef]
- Kuida, K.; Haydar, T.F.; Kuan, C.Y.; Gu, Y.; Taya, C.; Karasuyama, H.; Su, M.S.-S.; Rakic, P.; Flavell, R.A. Reduced apoptosis and cytochrome c-mediated caspase activation in mice lacking caspase 9. Cell 1998, 94, 325–337. [Google Scholar] [CrossRef] [Green Version]
- Srinivasula, S.M.; Ahmad, M.; Fernandes-Alnemri, T.; Alnemri, E.S. Autoactivation of procaspase-9 by Apaf-1-mediated oligomerization. Mol. Cell. 1998, 1, 949–957. [Google Scholar] [CrossRef]
- Shivapurkar, N.; Reddy, J.; Chaudhary, P.M.; Gazdar, A.F. Apoptosis and lung cancer: A review. J. Cell Biochem. 2003, 88, 885–898. [Google Scholar] [CrossRef]
- Sang, T.K.; Li, C.; Liu, W.; Rodriguez, A.; Abrams, J.M.; Zipursky, S.L.; Jackson, G.R. Inactivation of Drosophila Apaf-1 related killer suppresses formation of polyglutamine aggregates and blocks polyglutamine pathogenesis. Hum. Mol. Genet. 2005, 14, 357–372. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.Q.; Jiang, L.S.; Dai, L.Y. Programmed cell death in intervertebral disc degeneration. Apoptosis 2006, 11, 2079–2088. [Google Scholar] [CrossRef] [PubMed]
- Tegeder, I.; Costigan, M.; Griffin, R.S.; Abele, A.; Belfer, I.; Schmidt, H.; Ehnert, C.; Nejim, J.; Marian, C.; Scholz, J.; et al. GTP cyclohydrolase and tetrahydrobiopterin regulate pain sensitivity and persistence. Nat. Med. 2006, 12, 1269–1277. [Google Scholar] [CrossRef] [PubMed]
- Lotsch, J.; Klepstad, P.; Doehring, A.; Dale, O. A GTP cyclohydrolase 1 genetic variant delays cancer pain. Pain 2010, 148, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Ichinose, H.; Ohye, T.; Takahashi, E.; Seki, N.; Hori, T.-A.; Segawa, M.; Nomura, Y.; Endo, K.; Tanaka, H.; Tsuji, S.; et al. Hereditary progressive dystonia with marked diurnal fluctuation caused by mutations in the GTP cyclohydrolase I gene. Nat. Genet. 1994, 8, 236–242. [Google Scholar] [CrossRef]
- Bonafe, L.; Thony, B.; Penzien, J.M.; Czarnecki, B.; Blau, N. Mutations in the sepiapterin reductase gene cause a novel tetrahydrobiopterin-dependent monoamine-neurotransmitter deficiency without hyperphenylalaninemia. Am. J. Hum. Genet. 2001, 69, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Bisgaard, T.; Klarskov, B.; Rosenberg, J.; Kehlet, H. Characteristics and prediction of early pain after laparoscopic cholecystectomy. Pain 2001, 90, 261–269. [Google Scholar] [CrossRef]
- Bisgaard, T.; Rosenberg, J.; Kehlet, H. From acute to chronic pain after laparoscopic cholecystectomy: A prospective follow-up analysis. Scand. J. Gastroenterol. 2005, 40, 1358–1364. [Google Scholar] [CrossRef]
- Song, Y.Q.; Ho, D.W.; Karppinen, J.; Kao, P.Y.P.; Fan, B.J.; Luk, K.D.K.; Yip, S.P.; Leong, J.C.Y.; Cheah, K.S.E.; Sham, P.C.; et al. Association between promoter -1607 polymorphism of MMP1 and lumbar disc disease in Southern Chinese. BMC Med. Genet. 2008, 9, 38. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, L.M.; Schistad, E.I.; Storesund, A.; Pedersen, L.M.; Espeland, A.; Rygh, L.J.; Røe, C.; Gjerstad, J. The MMP1 rs1799750 2G allele is associated with increased low back pain, sciatica, and disability after lumbar disk herniation. Clin. J. Pain 2013, 29, 967–971. [Google Scholar] [CrossRef]
- Dev, R.; Srivastava, P.K.; Iyer, J.P.; Dastidar, S.G.; Ray, A. Therapeutic potential of matrix metalloprotease inhibitors in neuropathic pain. Expert Opin. Investig. Drugs 2010, 19, 455–468. [Google Scholar] [CrossRef] [PubMed]
- Dong, D.M.; Yao, M.; Liu, B.; Sun, C.Y.; Jiang, Y.Q.; Wang, Y.S. Association between the -1306C/T polymorphism of matrix metalloproteinase-2 gene and lumbar disc disease in Chinese young adults. Eur. Spine J. 2007, 16, 1958–1961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, M.; Haro, H.; Wakabayashi, Y.; Kawa-uchi, T.; Komori, H.; Shinomiya, K. The association of degeneration of the intervertebral disc with 5a/6a polymorphism in the promoter of the human matrix metalloproteinase-3 gene. J. Bone Jt. Surg. Br. 2001, 83, 491–495. [Google Scholar] [CrossRef]
- Omair, A.; Holden, M.; Lie, B.A.; Reikeras, O.; Brox, J.I. Treatment outcome of chronic low back pain and radiographic lumbar disc degeneration are associated with inflammatory and matrix degrading gene variants: A prospective genetic association study. BMC Musculoskelet. Disord. 2013, 14, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlosburg, J.E.; Kinsey, S.G.; Lichtman, A.H. Targeting fatty acid amide hydrolase (FAAH) to treat pain and inflammation. AAPS J. 2009, 11, 39–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piomelli, D.; Sasso, O. Peripheral gating of pain signals by endogenous lipid mediators. Nat. Neurosci. 2014, 17, 164–174. [Google Scholar] [CrossRef] [Green Version]
- Huggins, J.P.; Smart, T.S.; Langman, S.; Taylor, L.; Young, T. An efficient randomised, placebo-controlled clinical trial with the irreversible fatty acid amide hydrolase-1 inhibitor PF-04457845, which modulates endocannabinoids but fails to induce effective analgesia in patients with pain due to osteoarthritis of the knee. Pain 2012, 153, 1837–1846. [Google Scholar]
- Mu, J.; Ge, W.; Zuo, X.; Chen, Y.; Huang, C. A SNP in the 5′UTR of GDF5 is associated with susceptibility to symptomatic lumbar disc herniation in the Chinese Han population. Eur. Spine J. 2014, 23, 498–503. [Google Scholar] [CrossRef] [Green Version]
- Schistad, E.I.; Jacobsen, L.M.; Roe, C.; Gjerstad, J. The interleukin-1alpha gene C > T polymorphism rs1800587 is associated with increased pain intensity and decreased pressure pain thresholds in patients with lumbar radicular pain. Clin. J. Pain 2014, 30, 869–874. [Google Scholar] [CrossRef]
- Videman, T.; Saarela, J.; Kaprio, J.; Näkki, A.; Levälahti, E.; Gill, K.; Peltonen, L.; Battié, M.C. Associations of 25 structural, degradative, and inflammatory candidate genes with lumbar disc desiccation, bulging, and height narrowing. Arthritis Rheum. 2009, 60, 470–481. [Google Scholar] [CrossRef]
- Puren, A.J.; Fantuzzi, G.; Dinarello, C.A. Gene expression, synthesis, and secretion of interleukin 18 and interleukin 1beta are differentially regulated in human blood mononuclear cells and mouse spleen cells. Proc. Natl. Acad. Sci. USA 1999, 96, 2256–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavanaugh, J.M. Neural mechanisms of lumbar pain. Spine 1995, 20, 1804–1809. [Google Scholar] [CrossRef] [PubMed]
- Rannou, F.; Corvol, M.T.; Hudry, C.; Anract, P.; Dumontier, M.; Tsagris, L.; Revel, M.; Poiraudeau, S.; Serge, M.D. Sensitivity of anulus fibrosus cells to interleukin 1 beta. Comparison with articular chondrocytes. Spine 2000, 25, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Doita, M.; Kanatani, T.; Ozaki, T.; Matsui, N.; Kurosaka, M.; Yoshiya, S. Influence of macrophage infiltration of herniated disc tissue on the production of matrix metalloproteinases leading to disc resorption. Spine 2001, 26, 1522–1527. [Google Scholar] [CrossRef] [PubMed]
- Valdes, A.M.; Spector, T.D.; Doherty, S.; Wheeler, M.; Hart, D.J.; Doherty, M. Association of the DVWA and GDF5 polymorphisms with osteoarthritis in UK populations. Ann. Rheum. Dis. 2009, 68, 1916–1920. [Google Scholar] [CrossRef]
- Chapman, K.; Takahashi, A.; Meulenbelt, I.; Watson, C.; Rodríguez-López, J.; Egli, R.; Tsezou, A.; Malizos, K.N.; Kloppenburg, M.; Shi, D.; et al. A meta-analysis of European and Asian cohorts reveals a global role of a functional SNP in the 5′ UTR of GDF5 with osteoarthritis susceptibility. Hum. Mol. Genet. 2008, 17, 1497–1504. [Google Scholar] [CrossRef] [Green Version]
- Williams, F.M.; Popham, M.; Hart, D.J.; De Schepper, E.; Bierma-Zeinstra, S.; Hofman, A.; Uitterlinden, A.G.; Arden, N.K.; Cooper, C.; Spector, T.D.; et al. GDF5 single-nucleotide polymorphism rs143383 is associated with lumbar disc degeneration in Northern European women. Arthritis Rheum. 2011, 63, 708–712. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Stuber, F.; Stamer, U.M. Inflammatory mediators influence the expression of nociceptin and its receptor in human whole blood cultures. PLoS ONE 2013, 8, e74138. [Google Scholar] [CrossRef] [Green Version]
- Mu, J.; Ge, W.; Zuo, X.; Chen, Y.; Huang, C. Analysis of association between IL-1beta, CASP-9, and GDF5 variants and low-back pain in Chinese male soldier: Clinical article. J. Neurosurg. Spine 2013, 19, 243–247. [Google Scholar] [CrossRef] [Green Version]
- Williams, F.M.; Bansal, A.T.; van Meurs, J.B.; Bell, J.T.; Meulenbelt, I.; Suri, P.; Rivadeneira, F.; Sambrook, P.N.; Hofman, A.; Bierma-Zeinstra, S.; et al. Novel genetic variants associated with lumbar disc degeneration in northern Europeans: A meta-analysis of 4600 subjects. Ann. Rheum. Dis. 2013, 72, 1141–1148. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.F.; Lefebvre, V. The transcription factors SOX9 and SOX5/SOX6 cooperate genome-wide through super-enhancers to drive chondrogenesis. Nucleic Acids Res. 2015, 43, 8183–8203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smits, P.; Li, P.; Mandel, J.; Zhang, Z.; Deng, J.M.; Behringer, R.R.; De Crombrugghe, B.; Lefebvre, V. The transcription factors L-Sox5 and Sox6 are essential for cartilage formation. Dev. Cell 2001, 1, 277–290. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.F.; Samsa, W.E.; Zhou, G.; Lefebvre, V. Transcriptional control of chondrocyte specification and differentiation. Semin. Cell Dev. Biol. 2017, 62, 34–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smits, P.; Lefebvre, V. Sox5 and Sox6 are required for notochord extracellular matrix sheath formation, notochord cell survival and development of the nucleus pulposus of intervertebral discs. Development 2003, 130, 1135–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Fontenla, C.; Calaza, M.; Evangelou, E.; Valdes, A.M.; Arden, N.; Blanco, F.J.; Carr, A.; Chapman, K.; Deloukas, P.; Doherty, M.; et al. Assessment of osteoarthritis candidate genes in a meta-analysis of nine genome-wide association studies. Arthritis Rheumatol. 2014, 66, 940–949. [Google Scholar] [CrossRef] [Green Version]
- Bjornsdottir, G.; Benonisdottir, S.; Sveinbjornsson, G.; Styrkarsdottir, U.; Thorleifsson, G.; Walters, G.B.; Bjornsson, A.; Olafsson, I.H.; Ulfarsson, E.; Vikingsson, A.; et al. Sequence variant at 8q24.21 associates with sciatica caused by lumbar disc herniation. Nat. Commun. 2017, 8, 14265. [Google Scholar] [CrossRef] [Green Version]
- Truumees, E. A history of lumbar disc herniation from Hippocrates to the 1990s. Clin. Orthop. Relat. Res. 2015, 473, 1885–1895. [Google Scholar] [CrossRef] [Green Version]
- Chou, D.; Samartzis, D.; Bellabarba, C.; Patel, A.; Luk, K.; Kisser, J.M.S.; Skelly, A.C. Degenerative magnetic resonance imaging changes in patients with chronic low back pain: A systematic review. Spine 2011, 36, S43–S53. [Google Scholar] [CrossRef]
- Endean, A.; Palmer, K.T.; Coggon, D. Potential of magnetic resonance imaging findings to refine case definition for mechanical low back pain in epidemiological studies: A systematic review. Spine 2011, 36, 160–169. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Fukui, N.; Yahata, M.; Katsuragawa, Y.; Tashiro, T.; Ikegawa, S.; Lee, M.T.M. Genome-wide DNA methylation profile implicates potential cartilage regeneration at the late stage of knee osteoarthritis. Osteoarthr. Cartil. 2016, 24, 835–843. [Google Scholar] [CrossRef]
- Zhang, Y.; Fukui, N.; Yahata, M.; Katsuragawa, Y.; Tashiro, T.; Ikegawa, S.; Lee, M.T.M. Identification of DNA methylation changes associated with disease progression in subchondral bone with site-matched cartilage in knee osteoarthritis. Sci. Rep. 2016, 6, 34460. [Google Scholar] [CrossRef] [PubMed]
- Burston, J.J.; Sagar, D.R.; Shao, P.; Bai, M.; King, E.; Brailsford, L.; Turner, J.M.; Hathway, G.; Bennett, A.J.; Walsh, D.A.; et al. Cannabinoid CB2 receptors regulate central sensitization and pain responses associated with osteoarthritis of the knee joint. PLoS ONE 2013, 8, e80440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudd, R.A.; Seth, P.; David, F.; Scholl, L. Increases in Drug and Opioid-Involved Overdose Deaths—United States, 2010–2015. Morb. Mortal. Wkly. Rep. 2016, 65, 1445–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolodny, A.; Courtwright, D.T.; Hwang, C.S.; Kreiner, P.; Eadie, J.L.; Clark, T.W.; Alexander, G.C. The prescription opioid and heroin crisis: A public health approach to an epidemic of addiction. Annu. Rev. Public Health 2015, 36, 559–574. [Google Scholar] [CrossRef]
- Knezevic, N.N.; Tverdohleb, T.; Knezevic, I.; Candido, K.D. The Role of Genetic Polymorphisms in Chronic Pain Patients. Int. J. Mol. Sci. 2018, 19, 1707. [Google Scholar] [CrossRef] [Green Version]
- Meloto, C.B.; Benavides, R.; Lichtenwalter, R.N.; Wen, X.; Tugarinov, N.; Zorina-Lichtenwalter, K.; Chabot-Dore, A.-J.; Piltonen, M.H.; Cattaneo, S.; Verma, V.; et al. Human pain genetics database: A resource dedicated to human pain genetics research. Pain 2018, 159, 749–763. [Google Scholar] [CrossRef]
- Nelson, D.R.; Zeldin, D.C.; Hoffman, S.M.; Maltais, L.J.; Wain, H.M.; Nebert, D.W. Comparison of cytochrome P450 (CYP) genes from the mouse and human genomes, including nomenclature recommendations for genes, pseudogenes and alternative-splice variants. Pharmacogenetics 2004, 14, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Preissner, S.C.; Hoffmann, M.F.; Preissner, R.; Dunkel, M.; Gewiess, A.; Preissner, S. Polymorphic cytochrome P450 enzymes (CYPs) and their role in personalized therapy. PLoS ONE 2013, 8, e82562. [Google Scholar] [CrossRef]
- Yiannakopoulou, E. Pharmacogenomics of acetylsalicylic acid and other nonsteroidal anti-inflammatory agents: Clinical implications. Eur. J. Clin. Pharmacol. 2013, 69, 1369–1373. [Google Scholar] [CrossRef]
- Theken, K.N.; Lee, C.R.; Gong, L.; Caudle, K.E.; Formea, C.M.; Gaedigk, A.; Klein, T.E.; Agúndez, J.A.; Grosser, T. Clinical Pharmacogenetics Implementation Consortium Guideline (CPIC) for CYP2C9 and Nonsteroidal Anti-Inflammatory Drugs. Clin. Pharmacol. Ther. 2020, 108, 191–200. [Google Scholar] [CrossRef]
- Pilotto, A.; Seripa, D.; Franceschi, M.; Scarcelli, C.; Colaizzo, D.; Grandone, E.; Niro, V.; Andriulli, A.; Leandro, G.; Di Mario, F.; et al. Genetic susceptibility to nonsteroidal anti-inflammatory drug-related gastroduodenal bleeding: Role of cytochrome P450 2C9 polymorphisms. Gastroenterology 2007, 133, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Crews, K.R.; Gaedigk, A.; Dunnenberger, H.M.; Klein, T.E.; Shen, D.D.; Callaghan, J.T.; Kharasch, E.D.; Skaar, T.C. Clinical Pharmacogenetics Implementation Consortium (CPIC) guidelines for codeine therapy in the context of cytochrome P450 2D6 (CYP2D6) genotype. Clin. Pharmacol. Ther. 2012, 91, 321–326. [Google Scholar] [CrossRef]
- Dagostino, C.; Allegri, M.; Napolioni, V.; D’Agnelli, S.; Bignami, E.; Mutti, A.; Van Schaik, R.H. CYP2D6 genotype can help to predict effectiveness and safety during opioid treatment for chronic low back pain: Results from a retrospective study in an Italian cohort. Pharmgenomics Pers. Med. 2018, 11, 179–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batistaki, C.; Chrona, E.; Kostroglou, A.; Kostopanagiotou, G.; Gazouli, M. CYP2D6 Basic Genotyping of Patients with Chronic Pain Receiving Tramadol or Codeine. A Study in a Greek Cohort. Pain Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Boswell, M.V.; Stauble, M.E.; Loyd, G.E.; Langman, L.; Ramey-Hartung, B.; Baumgartner, R.N.; Tucker, W.W.; Jortani, S.A. The role of hydromorphone and OPRM1 in postoperative pain relief with hydrocodone. Pain Physician 2013, 16, E227–E235. [Google Scholar] [PubMed]
- Margarit, C.; Roca, R.; Inda, M.D.; Muriel, J.; Ballester, P.; Moreu, R.; Conte, A.L.; Nuñez, A.; Morales, M.; Peiró, A.M. Genetic Contribution in Low Back Pain: A Prospective Genetic Association Study. Pain Pract. 2019, 19, 836–847. [Google Scholar] [CrossRef]
- Rodieux, F.; Piguet, V.; Berney, P.; Desmeules, J.; Besson, M. Pharmacogenetics and analgesic effects of antidepressants in chronic pain management. Pers. Med. 2015, 12, 163–175. [Google Scholar] [CrossRef]
- Samer, C.F.; Lorenzini, K.I.; Rollason, V.; Daali, Y.; Desmeules, J.A. Applications of CYP450 testing in the clinical setting. Mol. Diagn. Ther. 2013, 17, 165–184. [Google Scholar] [CrossRef] [Green Version]
- Staddon, S.; Arranz, M.J.; Mancama, D.; Mata, I.; Kerwin, R.W. Clinical applications of pharmacogenetics in psychiatry. Psychopharmacology 2002, 162, 18–23. [Google Scholar] [CrossRef]
- Bertilsson, L.; Dahl, M.L.; Dalen, P.; Al-Shurbaji, A. Molecular genetics of CYP2D6: Clinical relevance with focus on psychotropic drugs. Br. J. Clin. Pharmacol. 2002, 53, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Swen, J.J.; Nijenhuis, M.; de Boer, A.; Grandia, L.; Maitland-van der Zee, A.H.; Mulder, H.; Rongen, G.A.P.J.M.; Van Schaik, R.H.N.; Schalekamp, T.; Touw, D.J.; et al. Pharmacogenetics: From bench to byte—An update of guidelines. Clin. Pharmacol. Ther. 2011, 89, 662–673. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.H.; Chou, C.H.; Yang, Y.K.; Lee, I.H.; Chen, P.S. Association between ABCB1 Polymorphisms and Antidepressant Treatment Response in Taiwanese Major Depressive Patients. Clin. Psychopharmacol. Neurosci. 2015, 13, 250–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.B.; Bousman, C.A.; Ng, C.H.; Byron, K.; Berk, M. ABCB1 polymorphism predicts escitalopram dose needed for remission in major depression. Transl. Psychiatry 2012, 27, e198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suwala, J.; Machowska, M.; Wiela-Hojenska, A. Venlafaxine pharmacogenetics: A comprehensive review. Pharmacogenomics 2019, 20, 829–845. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, C.; Luo, S.; Li, B.; Wager, T.D.; Zhang, W.; Rao, Y.; Han, S. Serotonin transporter polymorphism alters citalopram effects on human pain responses to physical pain. Neuroimage 2016, 135, 186–196. [Google Scholar] [CrossRef]
- Winchell, G.A.; King, J.D.; Chavez-Eng, C.M.; Constanzer, M.L.; Korn, S.H. Cyclobenzaprine pharmacokinetics, including the effects of age, gender, and hepatic insufficiency. J. Clin. Pharmacol. 2002, 42, 61–69. [Google Scholar] [CrossRef]
- Mestres, J.; Seifert, S.A.; Oprea, T.I. Linking pharmacology to clinical reports: Cyclobenzaprine and its possible association with serotonin syndrome. Clin. Pharmacol. Ther. 2011, 90, 662–665. [Google Scholar] [CrossRef]
- Perucca, E. Clinically relevant drug interactions with antiepileptic drugs. Br. J. Clin. Pharmacol. 2006, 61, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Ben-Menachem, E. Pregabalin pharmacology and its relevance to clinical practice. Epilepsia 2004, 45 (Suppl. 6), 13–18. [Google Scholar] [CrossRef]
- Honarmand, A.; Safavi, M.; Zare, M. Gabapentin: An update of its pharmacological properties and therapeutic use in epilepsy. J. Res. Med. Sci. 2011, 16, 1062–1069. [Google Scholar]
- Koepsell, H. The SLC22 family with transporters of organic cations, anions and zwitterions. Mol. Asp. Med. 2013, 34, 413–435. [Google Scholar] [CrossRef] [PubMed]
- Koepsell, H.; Lips, K.; Volk, C. Polyspecific organic cation transporters: Structure, function, physiological roles, and biopharmaceutical implications. Pharm. Res. 2007, 24, 1227–1251. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, P.A.; Benzi, J.R.L.; Azeredo, F.J.; Dach, F.; Ianhez Junior, E.; Zanelli, C.F.; De Moraes, N.V. Pharmacogenetics-based population pharmacokinetic analysis of gabapentin in patients with chronic pain: Effect of OCT2 and OCTN1 gene polymorphisms. Basic Clin. Pharmacol. Toxicol. 2019, 124, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Pergolizzi, J.V., Jr.; Labhsetwar, S.A.; Puenpatom, R.A.; Joo, S.; Ben-Joseph, R.H.; Summers, K.H. Prevalence of exposure to potential CYP450 pharmacokinetic drug-drug interactions among patients with chronic low back pain taking opioids. Pain Pract. 2011, 11, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Pergolizzi, J.V.; Ma, L.; Foster, D.R.; Overholser, B.R.; Sowinski, K.M.; Taylor, R., Jr.; Summers, K.H. The prevalence of opioid-related major potential drug-drug interactions and their impact on health care costs in chronic pain patients. J. Manag. Care Spec. Pharm. 2014, 20, 467–476. [Google Scholar] [CrossRef] [Green Version]
- Verbeurgt, P.; Mamiya, T.; Oesterheld, J. How common are drug and gene interactions? Prevalence in a sample of 1143 patients with CYP2C9, CYP2C19 and CYP2D6 genotyping. Pharmacogenomics 2014, 15, 655–665. [Google Scholar] [CrossRef]
- Malki, M.A.; Pearson, E.R. Drug-drug-gene interactions and adverse drug reactions. Pharm. J. 2020, 20, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Storelli, F.; Desmeules, J.; Daali, Y. Physiologically-Based Pharmacokinetic Modeling for the Prediction of CYP2D6-Mediated Gene-Drug-Drug Interactions. CPT Pharmacomet. Syst. Pharmacol. 2019, 8, 567–576. [Google Scholar] [CrossRef]
- Trescot, A.M.; Faynboym, S. A review of the role of genetic testing in pain medicine. Pain Physician 2014, 17, 425–445. [Google Scholar]



{kind=link}
{kind=link}
| Gene | Function/Pathway | Condition(s) | Citation | Number of Subjects/Geographic Region |
|---|---|---|---|---|
| DCC | Receptor for Netrin-1, as an axonal guidance molecule | LBP | Suri et al., 2018 [21] | n = 168,000 |
| ESR1 | Other/Estrogen receptor 1 | LBP | Roh et al., 2013 [29] | n = 192, South Korea |
| ADRB2 | Neurotransmission/beta-2 adrenergic receptor | TMD/LBP/Fibromyalgia LBP comorbid with neck pain | Diatchenko et al., 2006 [36]/Skouen et al. [35]/Vargas-Alarcon et al., 2009 [37] | n = 1004; Western Australian Pregnancy (Raine) Cohort |
| CNR2 | Peripheral cannabinoid receptor; nociceptive transmission, inflammatory response, bone homeostasis | LBP/mechanical allodynia, neuroinflammation in CRPS1/Joint pain | Starkweather et al., 2017 [38]; Ramesh et al., 2018 [34]/Xu et al., 2016 [39] | n = 62 USA; n = 84 USA/animal model/animal model |
| OPRM1 | Neurotransmission/Mu opioid receptor | LBP | Hasvik et al., 2014 [10], Omair et al. have not replicated the above (2015) [40] | n = 118 Caucasians, Norway |
| Gene | Function/Pathway | Condition(s) | Citation | Number of Subjects/Geographic Region |
|---|---|---|---|---|
| FAAH | hydrolyzes many primary and secondary fatty acid amides, including anandamide and oleamide as neuromodulators | cLBP | Ramesh et al., 2018 [34] | n = 84, USA |
| COMT | Neurotransmission/Catechol-O-methyltransferase | LBP | Rut et al., 2014 [45], Jacobsen et al., 2012 [44], Omair et al., 2013, 2015 [40,64] | n = 176, Poland/n = 258, Norway/n = 93, Norway (West Eur), n = 371, Europe |
| GCH | Guanosine triphosphate cyclohydrolase | LBP | Tegeder et al., 2006 [53] | Animal studies |
| MMP1 | Protein degradation/matrix metalloproteinase | LBP | Song et al., 2008 [59] Jacobsen et al., 2013 [60] | n = 691, southern China; n = 260, Norway Eur White |
| MMP2 | Protein degradation | LBP | Dong et al., 2007 [62] | n = 162, China |
| MMP3 | Protein degradation | LBP | Takahashi et al., 2001 [63] | n = 103, Japan |
| CASP9 | Apoptosis-mediating caspase | LBP | Guo et al., 2011 [46], Mu et al., 2013 [68] | n = 154/216 controls in China; n = 305/587 controls Chinese soldiers |
| Gene | Function/Pathway | Condition(s) | Citation | Number of Subjects/Geographic Region |
|---|---|---|---|---|
| CCL2 | Chemotactic factor for monocytes and basophils | LBP | Starkweather et al., 2017 [38] | n = 62, USA |
| IL18R1 IL18RAP IL1A | Immune response/Interleukin receptors | LBP | Omair et al., 2013 [64] Schistad et al., 2014 [69] | n = 93, Norway; n = 121, Norway |
| GDF5 | Part of TGF-beta family, Cellular growth/Skeletal tissue differentiation | LBP | Mu et al., 2013 [79] | n = 305/587 controls Chinese soldiers |
| Gene | Function/Pathway | Condition(s) | Citation | Number of Subjects/Geographic Location |
|---|---|---|---|---|
| SOX5 | Transcription factor, embryonic development | LBP | Suri et al., 2018 [21] | n = 168,000; worldwide |
| CCDC26/GSDMC | Non-coding/Codes gasdermin C; the N-terminal moiety promotes pyroptosis with unknown physiologic significance | LBP | Suri et al., 2018 [21] | n = 168,000; worldwide |
| PNOC | Codes prepronociceptin; nociceptin is a ligand of the opioid receptor-like receptor OPRL1; may modulate nociceptive and locomotor behavior | LBP | Starkweather et al., 2016 [38] | n = 62, CT USA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suntsov, V.; Jovanovic, F.; Knezevic, E.; Candido, K.D.; Knezevic, N.N. Can Implementation of Genetics and Pharmacogenomics Improve Treatment of Chronic Low Back Pain? Pharmaceutics 2020, 12, 894. https://doi.org/10.3390/pharmaceutics12090894
Suntsov V, Jovanovic F, Knezevic E, Candido KD, Knezevic NN. Can Implementation of Genetics and Pharmacogenomics Improve Treatment of Chronic Low Back Pain? Pharmaceutics. 2020; 12(9):894. https://doi.org/10.3390/pharmaceutics12090894
Chicago/Turabian StyleSuntsov, Vladislav, Filip Jovanovic, Emilija Knezevic, Kenneth D. Candido, and Nebojsa Nick Knezevic. 2020. "Can Implementation of Genetics and Pharmacogenomics Improve Treatment of Chronic Low Back Pain?" Pharmaceutics 12, no. 9: 894. https://doi.org/10.3390/pharmaceutics12090894