CancerGram: An Effective Classifier for Differentiating Anticancer from Antimicrobial Peptides

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Sets
2.2. Cross-Validation
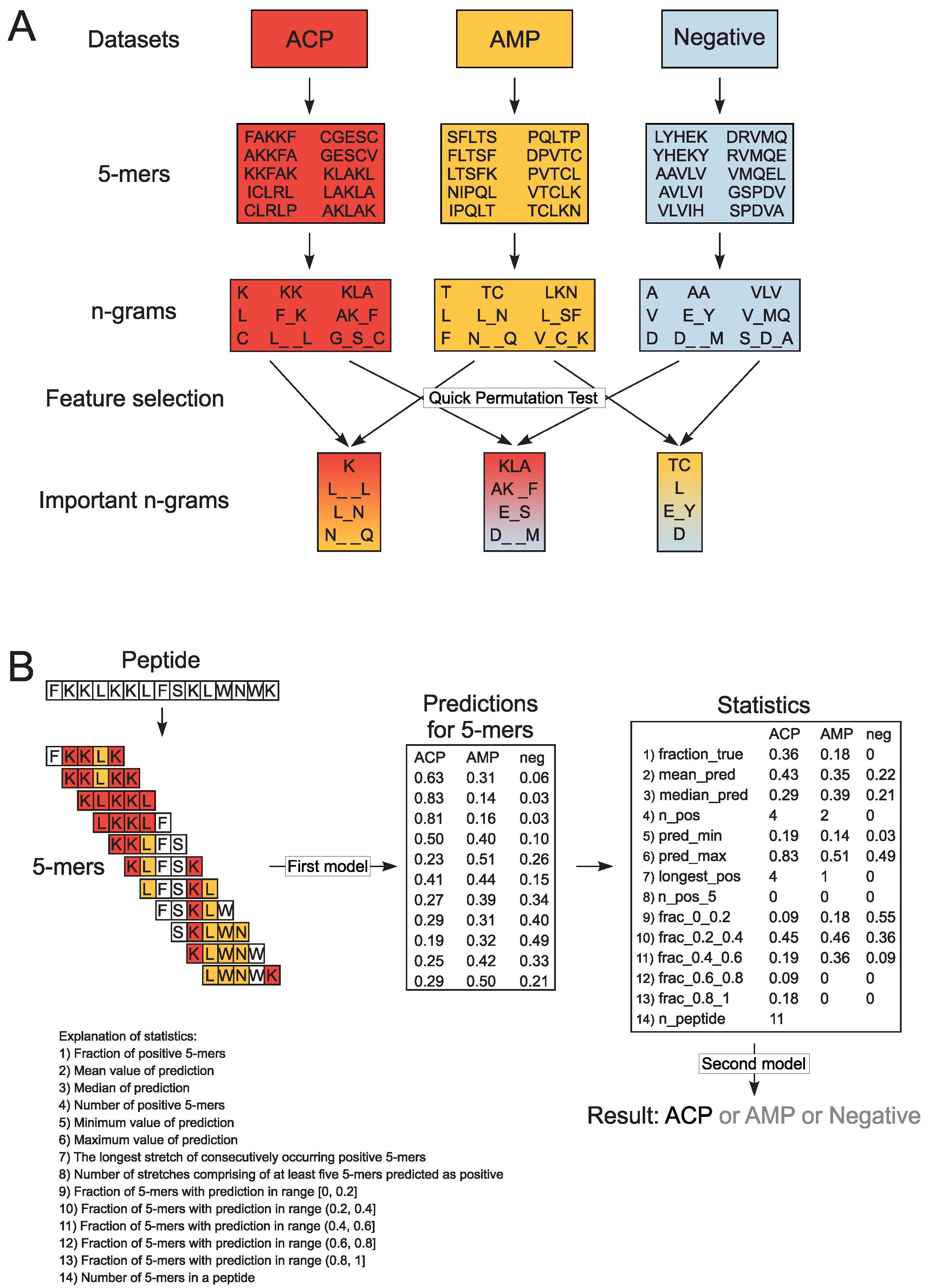
2.3. Extraction of Encoded N-Grams
2.4. Model Training with Random Forests
3. Results and Discussion
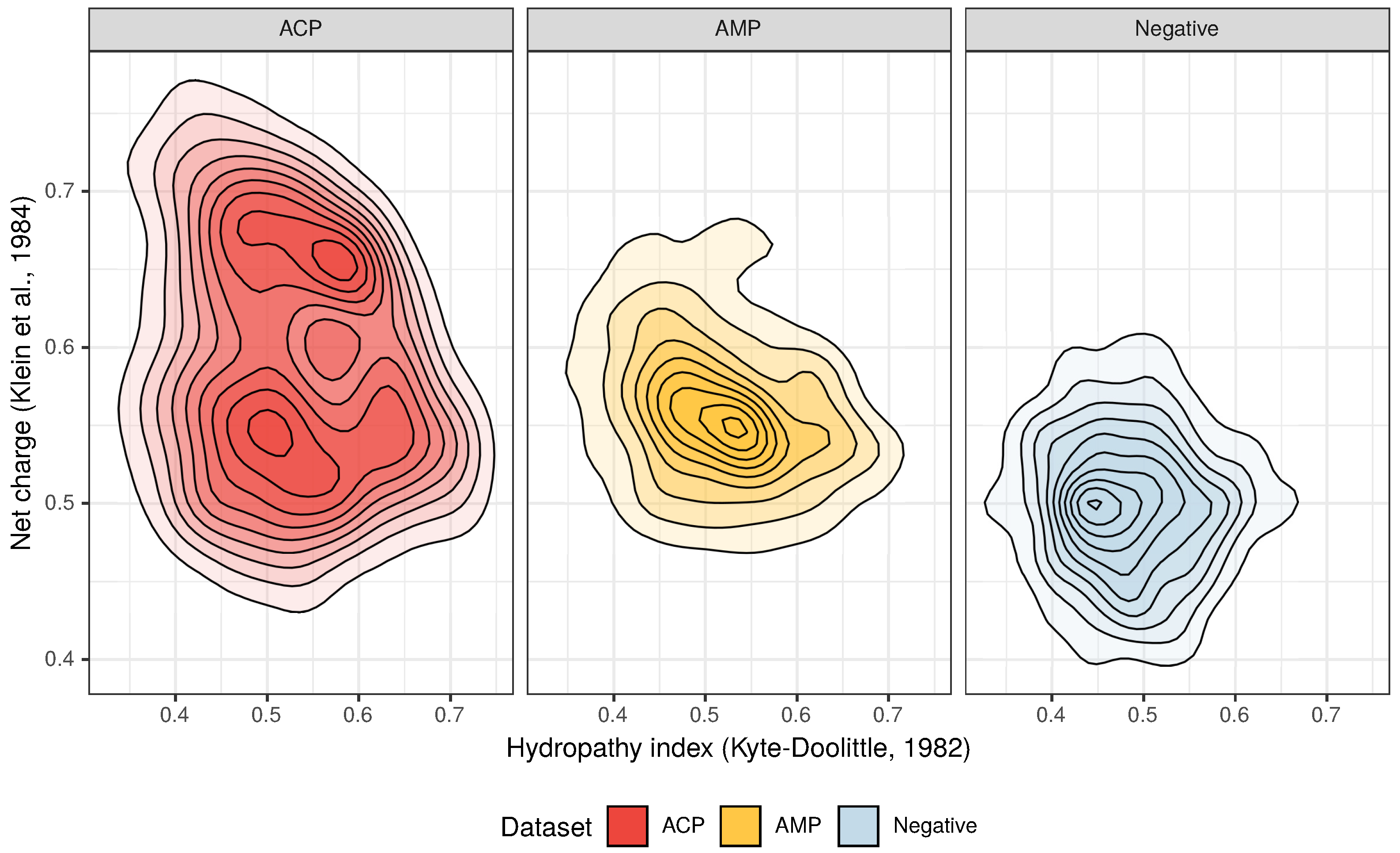
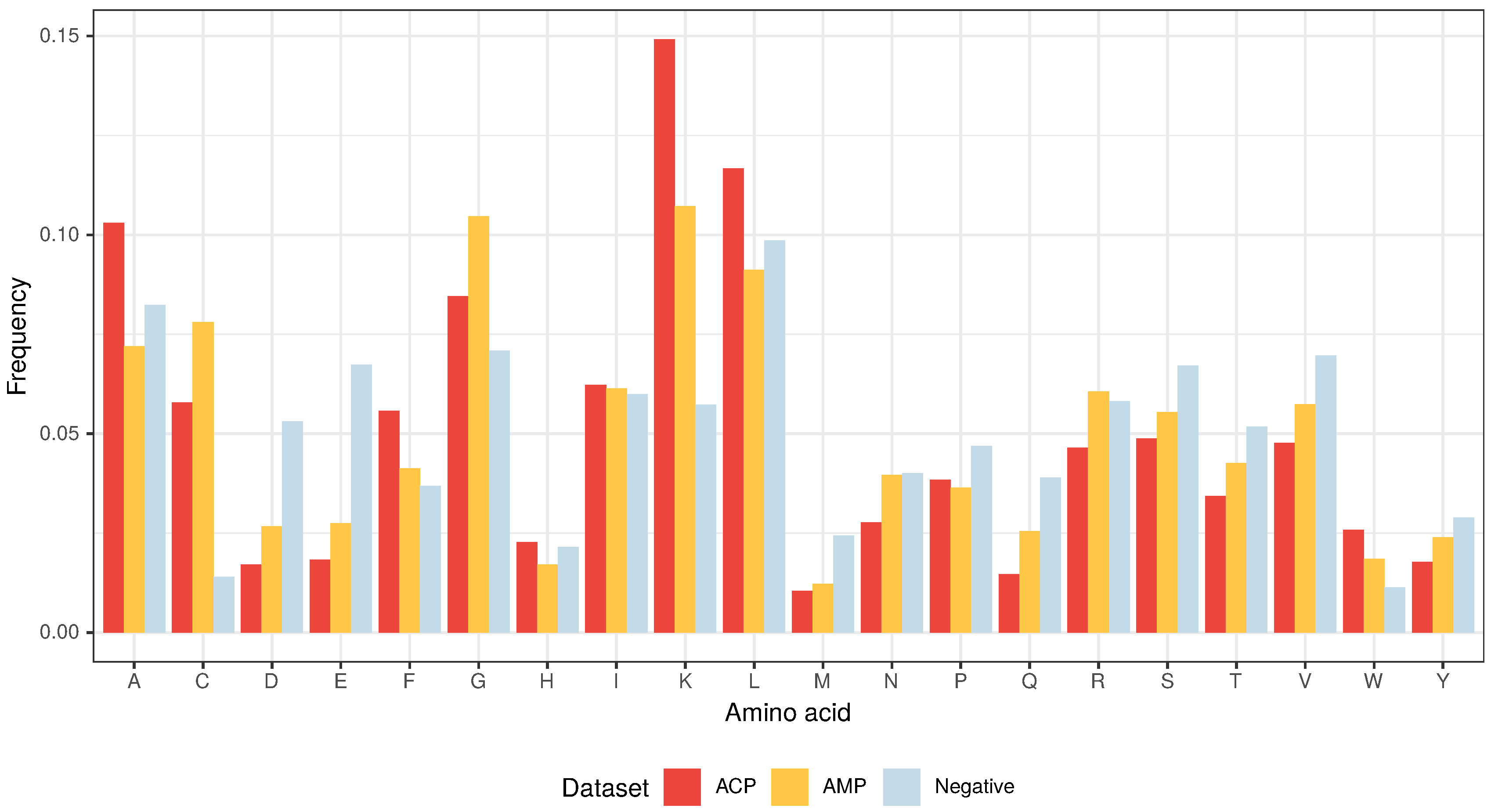
3.1. Composition and Properties of ACPs and AMPs
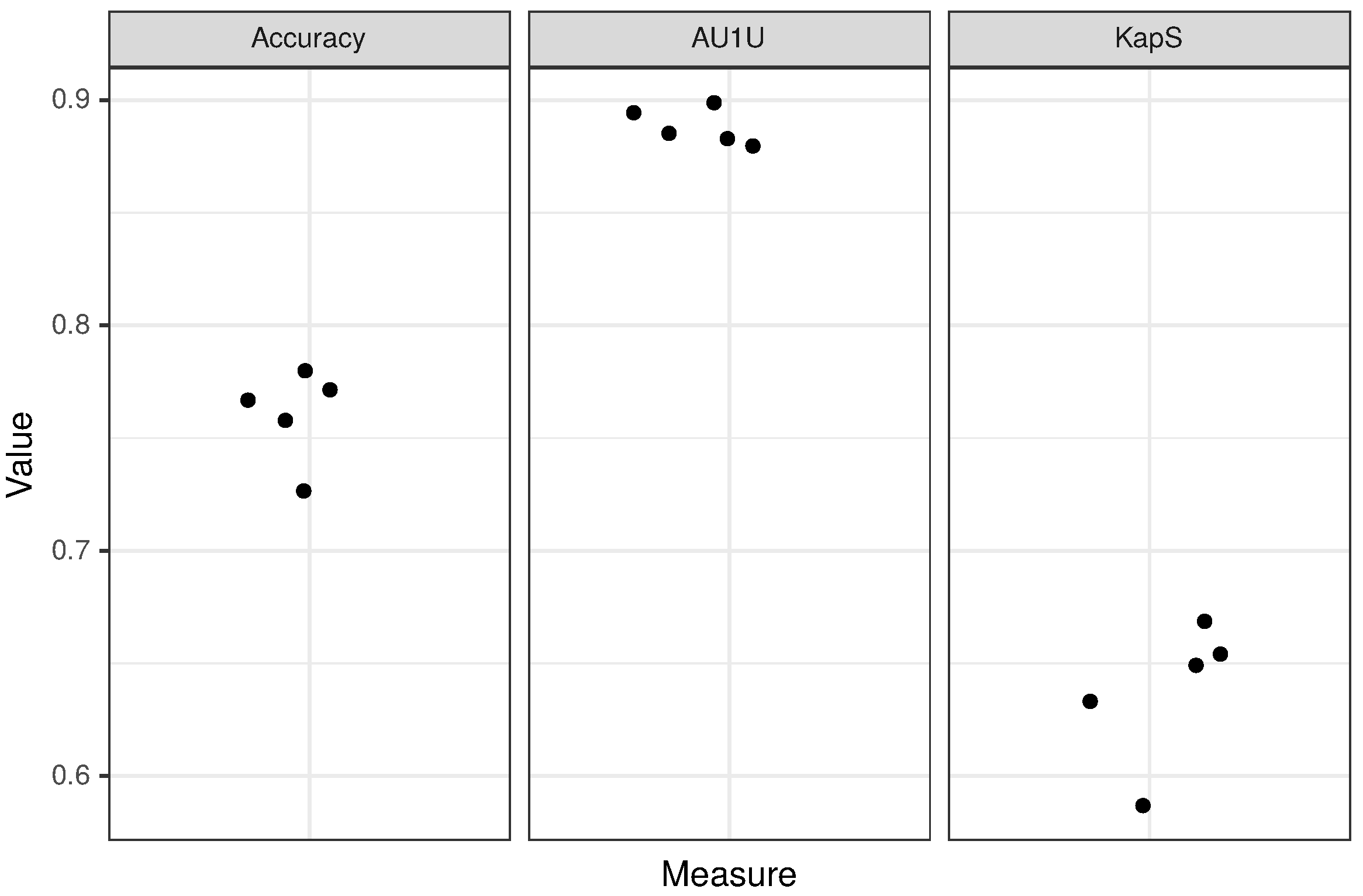
3.2. CancerGram Performance
3.3. Prediction of Mitochondria-Targeted ACPs with CancerGram
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AUC | Area under the ROC curve |
| AU1U | AUC for binary comparisons of each class against each other |
| ACP | Anticancer peptides |
| AMP | Antimicrobial peptides |
| KapS | Kappa statistic |
| MCC | Matthews correlation coefficient |
| A | Alanine |
| R | Arginine |
| N | Asparagine |
| D | Aspartic acid |
| C | Cysteine |
| E | Glutamic acid |
| Q | Glutamine |
| G | Glycine |
| H | Histidine |
| I | Isoleucine |
| L | Leucine |
| K | Lysine |
| M | Methionine |
| F | Phenylalanine |
| P | Proline |
| S | Serine |
| T | Threonine |
| W | Tryptophan |
| Y | Tyrosine |
| V | Valine |
Appendix A. Availability and Implementation
References
- Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 13 October 2020).
- Cassini, A.; Högberg, L.D.; Plachouras, D.; Quattrocchi, A.; Hoxha, A.; Simonsen, G.S.; Colomb-Cotinat, M.; Kretzschmar, M.E.; Devleesschauwer, B.; Cecchini, M.; et al. Attributable deaths and disability-adjusted life-years caused by infections with antibiotic-resistant bacteria in the EU and the European Economic Area in 2015: A population-level modelling analysis. Lancet Infect. Dis. 2019, 19, 56–66. [Google Scholar] [CrossRef] [Green Version]
- CDC. Antibiotic Resistance Threats in the United States, 2019; Centres for Disease Control and Prevention, US Department of Health and Human Services: Washington, DC, USA, 2019.
- Ahmed, A.; Siman-Tov, G.; Hall, G.; Bhalla, N.; Narayanan, A. Human antimicrobial peptides as therapeutics for viral infections. Viruses 2019, 11, 704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mookherjee, N.; Anderson, M.A.; Haagsman, H.P.; Davidson, D.J. Antimicrobial host defence peptides: Functions and clinical potential. Nat. Rev. Drug Discov. 2020, 19, 311–332. [Google Scholar] [CrossRef] [PubMed]
- Raffatellu, M. Learning from bacterial competition in the host to develop antimicrobials. Nat. Med. 2018, 24, 1097–1103. [Google Scholar] [CrossRef]
- Suneja, G.; Nain, S.; Sharma, R. Microbiome: A Source of Novel Bioactive Compounds and Antimicrobial Peptides. In Microbial Diversity in Ecosystem Sustainability and Biotechnological Applications; Springer: Berlin, Germany, 2019; pp. 615–630. [Google Scholar]
- Felício, M.R.; Silva, O.N.; Gonçalves, S.; Santos, N.C.; Franco, O.L. Peptides with dual antimicrobial and anticancer activities. Front. Chem. 2017, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Travkova, O.G.; Moehwald, H.; Brezesinski, G. The interaction of antimicrobial peptides with membranes. Adv. Colloid Interface Sci. 2017, 247, 521–532. [Google Scholar] [CrossRef]
- Gaspar, D.; Veiga, A.S.; Castanho, M.A. From antimicrobial to anticancer peptides. A review. Front. Microbiol. 2013, 4, 294. [Google Scholar] [CrossRef] [Green Version]
- Marquette, A.; Bechinger, B. Biophysical investigations elucidating the mechanisms of action of antimicrobial peptides and their synergism. Biomolecules 2018, 8, 18. [Google Scholar] [CrossRef] [Green Version]
- Tornesello, A.L.; Borrelli, A.; Buonaguro, L.; Buonaguro, F.M.; Tornesello, M.L. Antimicrobial peptides as anticancer agents: Functional properties and biological activities. Molecules 2020, 25, 2850. [Google Scholar] [CrossRef]
- Agrawal, P.; Bhagat, D.; Mahalwal, M.; Sharma, N.; Raghava, G.P.S. AntiCP 2.0: An updated model for predicting anticancer peptides. Brief. Bioinf. 2020. [Google Scholar] [CrossRef]
- Martin, W.F.; Neukirchen, S.; Zimorski, V.; Gould, S.B.; Sousa, F.L. Energy for two: New archaeal lineages and the origin of mitochondria. BioEssays 2016, 38, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Wu, D.; Goremykin, V.; Xiao, J.; Xu, Y.; Garg, S.; Zhang, C.; Martin, W.F.; Zhu, R. Phylogenetic analyses with systematic taxon sampling show that mitochondria branch within Alphaproteobacteria. Nat. Ecol. Evol. 2020, 4, 1213–1219. [Google Scholar] [CrossRef] [PubMed]
- Jeena, M.; Kim, S.; Jin, S.; Ryu, J.H. Recent progress in mitochondria-targeted drug and drug-free agents for cancer therapy. Cancers 2020, 12, 4. [Google Scholar] [CrossRef] [Green Version]
- Newmeyer, D.D.; Ferguson-Miller, S. Mitochondria: Releasing power for life and unleashing the machineries of death. Cell 2003, 112, 481–490. [Google Scholar] [CrossRef] [Green Version]
- Szewczyk, A.; Wojtczak, L. Mitochondria as a pharmacological target. Pharmacol. Rev. 2002, 54, 101–127. [Google Scholar] [CrossRef] [Green Version]
- Zorova, L.D.; Popkov, V.A.; Plotnikov, E.Y.; Silachev, D.N.; Pevzner, I.B.; Jankauskas, S.S.; Babenko, V.A.; Zorov, S.D.; Balakireva, A.V.; Juhaszova, M.; et al. Mitochondrial membrane potential. Anal. Biochem. 2018, 552, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Houston, M.A.; Augenlicht, L.H.; Heerdt, B.G. Stable differences in intrinsic mitochondrial membrane potential of tumor cell subpopulations reflect phenotypic heterogeneity. Int. J. Cell Biol. 2011, 2011. [Google Scholar] [CrossRef] [Green Version]
- Constance, J.E.; Lim, C.S. Targeting malignant mitochondria with therapeutic peptides. Ther. Deliv. 2012, 3, 961–979. [Google Scholar] [CrossRef] [Green Version]
- Bansal, S.; Mittal, A. A statistical anomaly indicates symbiotic origins of eukaryotic membranes. Mol. Biol. Cell 2015, 26, 1238–1248. [Google Scholar] [CrossRef]
- Rappocciolo, E.; Stiban, J. Prokaryotic and mitochondrial lipids: A survey of evolutionary origins. In Bioactive Ceramides in Health and Disease; Springer: Berlin, Germany, 2019; pp. 5–31. [Google Scholar]
- Wollman, F.A. An antimicrobial origin of transit peptides accounts for early endosymbiotic events. Traffic 2016, 17, 1322–1328. [Google Scholar] [CrossRef]
- Garrido, C.O.; Caspari, O.D.; Choquet, Y.; Wollman, F.A.; Lafontaine, I. An antimicrobial origin of targeting peptides to endosymbiotic organelles. Cells 2020, 9, 1795. [Google Scholar] [CrossRef]
- Dudek, J.; Rehling, P.; van der Laan, M. Mitochondrial protein import: Common principles and physiological networks. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Res. 2013, 1833, 274–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Feng, Q.; Yan, Q.; Hao, X.; Chen, Y. Alpha-helical cationic anticancer peptides: A promising candidate for novel anticancer drugs. Mini Rev. Med. Chem. 2015, 15, 73–81. [Google Scholar] [CrossRef]
- Burdukiewicz, M.; Sidorczuk, K.; Rafacz, D.; Pietluch, F.; Chilimoniuk, J.; Rödiger, S.; Gagat, P. Proteomic Screening for Prediction and Design of Antimicrobial Peptides with AmpGram. Int. J. Mol. Sci. 2020, 21, 4310. [Google Scholar] [CrossRef]
- Burdukiewicz, M.; Sobczyk, P.; Rödiger, S.; Duda-Madej, A.; Mackiewicz, P.; Kotulska, M. Amyloidogenic motifs revealed by n-gram analysis. Sci. Rep. 2017, 7, 12961. [Google Scholar] [CrossRef] [Green Version]
- Burdukiewicz, M.; Sobczyk, P.; Chilimoniuk, J.; Gagat, P.; Mackiewicz, P. Prediction of signal peptides in proteins from malaria parasites. Int. J. Mol. Sci. 2018, 19, 3709. [Google Scholar] [CrossRef] [Green Version]
- Burdukiewicz, M.; Gagat, P.; Jabłoński, S.; Chilimoniuk, J.; Gaworski, M.; Mackiewicz, P.; Marcin, Ł. PhyMet2: A database and toolkit for phylogenetic and metabolic analyses of methanogens. Environ. Microbiol. Rep. 2018, 10, 378–382. [Google Scholar]
- Tyagi, A.; Tuknait, A.; Anand, P.; Gupta, S.; Sharma, M.; Mathur, D.; Joshi, A.; Singh, S.; Gautam, A.; Raghava, G.P. CancerPPD: A database of anticancer peptides and proteins. Nucleic Acids Res. 2014, 43, D837–D843. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2015, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, X.; Dong, F.; Shi, C.; Liu, S.; Sun, J.; Chen, J.; Li, H.; Xu, H.; Lao, X.; Zheng, H. DRAMP 2.0, an updated data repository of antimicrobial peptides. Sci. Data 2019, 6, 148. [Google Scholar] [CrossRef] [Green Version]
- Jhong, J.H.; Chi, Y.H.; Li, W.C.; Lin, T.H.; Huang, K.Y.; Lee, T.Y. dbAMP: An integrated resource for exploring antimicrobial peptides with functional activities and physicochemical properties on transcriptome and proteome data. Nucleic Acids Res. 2018, 47, D285–D297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef] [PubMed]
- Burdukiewicz, M.; Sobczyk, P.; Lauber, C. Biogram: N-Gram Analysis of Biological Sequences; GitHub: San Francisco, CA, USA, 2020. [Google Scholar]
- Wright, M.N.; Ziegler, A. Ranger: A Fast Implementation of Random Forests for High Dimensional Data in C++ and R. J. Stat. Softw. 2017, 77, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Bell, J.; Larson, M.; Kutzler, M.; Bionaz, M.; Löhr, C.V.; Hendrix, D. miRWoods: Enhanced Precursor Detection and Stacked Random Forests for the Sensitive Detection of microRNAs. PLoS Comput. Biol. 2019, 15, e1007309. [Google Scholar] [CrossRef]
- Huang, Y.B.; He, L.Y.; Jiang, H.Y.; Chen, Y.X. Role of helicity on the anticancer mechanism of action of cationic-helical peptides. Int. J. Mol. Sci. 2012, 13, 6849–6862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.b.; Wang, X.f.; Wang, H.y.; Liu, Y.; Chen, Y. Studies on mechanism of action of anticancer peptides by modulation of hydrophobicity within a defined structural framework. Mol. Cancer Ther. 2011, 10, 416–426. [Google Scholar] [CrossRef] [Green Version]
- Bhunia, D.; Mondal, P.; Das, G.; Saha, A.; Sengupta, P.; Jana, J.; Mohapatra, S.; Chatterjee, S.; Ghosh, S. Spatial position regulates power of tryptophan: Discovery of a major-groove-specific nuclear-localizing, cell-penetrating tetrapeptide. J. Am. Chem. Soc. 2018, 140, 1697–1714. [Google Scholar] [CrossRef]
- Chiangjong, W.; Chutipongtanate, S.; Hongeng, S. Anticancer peptide: Physicochemical property, functional aspect and trend in clinical application. Int. J. Oncol. 2020, 57, 678–696. [Google Scholar] [CrossRef]
- Ferri, C.; Hernández-Orallo, J.; Modroiu, R. An experimental comparison of performance measures for classification. Pattern Recognit. Lett. 2009, 30, 27–38. [Google Scholar] [CrossRef]
- Hand, D.J.; Till, R.J. A simple generalisation of the area under the ROC curve for multiple class classification problems. Mach. Learn. 2001, 45, 171–186. [Google Scholar] [CrossRef]
- Ben-David, A. About the relationship between ROC curves and Cohen’s kappa. Eng. Appl. Artif. Intell. 2008, 21, 874–882. [Google Scholar] [CrossRef]
- Ranganathan, P.; Pramesh, C.; Aggarwal, R. Common pitfalls in statistical analysis: Measures of agreement. Perspect. Clin. Res. 2017, 8, 187. [Google Scholar] [CrossRef]
- Bischl, B.; Lang, M.; Kotthoff, L.; Schiffner, J.; Richter, J.; Studerus, E.; Casalicchio, G.; Jones, Z.M. mlr: Machine Learning in R. J. Mach. Learn. Res. 2016, 17, 1–5. [Google Scholar]
- Boopathi, V.; Subramaniyam, S.; Malik, A.; Lee, G.; Manavalan, B.; Yang, D.C. mACPpred: A support vector machine-based meta-predictor for identification of anticancer peptides. Int. J. Mol. Sci. 2019, 20, 1964. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Chen, C.X.; Hu, J.; Zhou, P.; Zeng, P.; Cao, C.H.; Lu, J.R. Dual modes of antitumor action of an amphiphilic peptide A9K. Biomaterials 2013, 34, 2731–2737. [Google Scholar] [CrossRef] [PubMed]
- Farsinejad, S.; Gheisary, Z.; Samani, S.E.; Alizadeh, A.M. Mitochondrial targeted peptides for cancer therapy. Tumor Biol. 2015, 36, 5715–5725. [Google Scholar] [CrossRef]
- Yitzchak, H. Disease Treatment Via Antimicrobial Peptides Or Their Inhibitors. U.S. Patent 8202835 B2, 19 June 2012. [Google Scholar]
- Hao, X.; Yan, Q.; Zhao, J.; Wang, W.; Huang, Y.; Chen, Y. TAT modification of alpha-helical anticancer peptides to improve specificity and efficacy. PLoS ONE 2015, 10, e0138911. [Google Scholar] [CrossRef] [Green Version]
- Javadpour, M.M.; Juban, M.M.; Lo, W.C.; Bishop, S.M.; Alberty, J.B.; Cowell, S.M.; Becker, C.L.; McLaughlin, M.L. De novo antimicrobial peptides with low mammalian cell toxicity. J. Med. Chem. 1996, 39, 3107–3113. [Google Scholar] [CrossRef]
- Horton, K.L.; Kelley, S.O. Engineered apoptosis-inducing peptides with enhanced mitochondrial localization and potency. J. Med. Chem. 2009, 52, 3293–3299. [Google Scholar] [CrossRef]
- Ellerby, H.M.; Arap, W.; Ellerby, L.M.; Kain, R.; Andrusiak, R.; Del Rio, G.; Krajewski, S.; Lombardo, C.R.; Rao, R.; Ruoslahti, E.; et al. Anti-cancer activity of targeted pro-apoptotic peptides. Nat. Med. 1999, 5, 1032–1038. [Google Scholar] [CrossRef]
- Bellamy, W.; Takase, M.; Yamauchi, K.; Wakabayashi, H.; Kawase, K.; Tomita, M. Identification of the bactericidal domain of lactoferrin. Biochim. Et Biophys. Acta (BBA) Protein Struct. Mol. Enzymol. 1992, 1121, 130–136. [Google Scholar] [CrossRef]
- Eliassen, L.T.; Berge, G.; Leknessund, A.; Wikman, M.; Lindin, I.; Løkke, C.; Ponthan, F.; Johnsen, J.I.; Sveinbjørnsson, B.; Kogner, P.; et al. The antimicrobial peptide, lactoferricin B, is cytotoxic to neuroblastoma cells in vitro and inhibits xenograft growth in vivo. Int. J. Cancer 2006, 119, 493–500. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Chamorro, L.; Puertollano, M.A.; Puertollano, E.; de Cienfuegos, G.Á.; de Pablo, M.A. In vitro biological activities of magainin alone or in combination with nisin. Peptides 2006, 27, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Argiolas, A.; Pisano, J.J. Isolation and characterization of two new peptides, mastoparan C and crabrolin, from the venom of the European hornet, Vespa crabro. J. Biol. Chem. 1984, 259, 10106–10111. [Google Scholar] [PubMed]
- Chen, X.; Zhang, L.; Wu, Y.; Wang, L.; Ma, C.; Xi, X.; Bininda-Emonds, O.R.; Shaw, C.; Chen, T.; Zhou, M. Evaluation of the bioactivity of a mastoparan peptide from wasp venom and of its analogues designed through targeted engineering. Int. J. Biol. Sci. 2018, 14, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.L.S.; Li, J.H.; Yu, C.Y.; Lin, C.J.; Chiu, P.H.; Chen, P.W.; Lin, C.C.; Chen, W.J. Novel cationic antimicrobial peptide GW-H1 induced caspase-dependent apoptosis of hepatocellular carcinoma cell lines. Peptides 2012, 36, 257–265. [Google Scholar] [CrossRef]
- Hilchie, A.; Doucette, C.; DM, P.; Patrzykat, A.; Douglas, S.; Hoskin, D.W. Pleurocidin-family cationic antimicrobial peptides are cytolytic for breast carcinoma cells and prevent growth of tumor xenografts. Breast Cancer Res. 2011, 13, R102. [Google Scholar] [CrossRef] [Green Version]
- Law, B.; Quinti, L.; Choi, Y.; Weissleder, R.; Tung, C.H. A mitochondrial targeted fusion peptide exhibits remarkable cytotoxicity. Mol. Cancer Ther. 2006, 5, 1944–1949. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Set | ACP | AMP | Negative |
|---|---|---|---|
| Training | 686 | 689 | 776 |
| Validation | 171 | 170 | 194 |
| Independent | 57 | 769 | 0 |
| Measure | Mer Layer | Peptide Layer |
|---|---|---|
| Accuracy | 0.64 (+/−0.01) | 0.76 (+/−0.021) |
| AU1U | 0.79 (+/−0.006) | 0.89 (+/−0.008) |
| KapS | 0.44 (+/−0.015) | 0.64 (+/−0.032) |
| Measure | Value |
|---|---|
| Accuracy | 0.77 |
| AU1U | 0.89 |
| KapS | 0.65 |
| Software | MCC | Precision | Sensitivity | Specificity | Accuracy | AUC |
|---|---|---|---|---|---|---|
| CancerGram | 0.15 | 0.17 | 0.30 | 0.89 | 0.85 | 0.60 |
| AntiCP 2.0 | 0.07 | 0.10 | 0.32 | 0.79 | 0.76 | 0.53 |
| Software | MCC | Precision | Sensitivity | Specificity | Accuracy | AUC |
|---|---|---|---|---|---|---|
| CancerGram | 0.57 | 0.78 | 0.71 | 0.85 | 0.79 | 0.83 |
| mACPpred | 0.21 | 0.48 | 0.90 | 0.27 | 0.54 | 0.68 |
| Peptide | Sequence | Reference |
|---|---|---|
| AK | AAAAAAAAAK | [50] |
| hCAP-18 | FRKSKEKIGKEFKRIVQRIKDFLRNLVPRTES | [51,52] |
| HPRP-A1-TAT | FKKLKKLFSKLWNWKRKKRRQRRR | [53] |
| KLA | KLAKLAKKLAKLAK | [54,55,56] |
| Lactoferricin B | FKCRRWQWRMKKLGAPSITCVRRAF | [57,58] |
| Magainin 1 | GIGKFLHSAGKFGKAFVGEIMKS | [59] |
| Mastoparan-C | LNLKALLAVAKKIL | [60,61] |
| NGR Peptide 1 | CNGRCGGKLAKLAKKLAKLAK | [56] |
| GW-H1 | GYNYAKKLANLAKKFANALW | [62] |
| Pleurocidin NRC-03 | GRRKRKWLRRIGKGVKIIGGAALDHL | [63] |
| R7-kla | RRRRRRRKLAKLAKKLAKLAK | [64] |
| RGD-4C-GG-(KLAKLAK) | ACDCRGDCFCGGKLAKLAKKLAKLAK | [56] |
| Peptide | ACP | AMP | Negative | Decision |
|---|---|---|---|---|
| AK | 0.10 | 0.32 | 0.58 | Negative |
| GW-H1 | 0.31 | 0.64 | 0.06 | AMP |
| hCAP-18 | 0.96 | 0.04 | 0.00 | ACP |
| HPRP-A1-TAT | 0.66 | 0.33 | 0.01 | ACP |
| KLA | 1.00 | 0.00 | 0.00 | ACP |
| Lactoferricin B | 0.10 | 0.90 | 0.00 | AMP |
| Magainin 1 | 0.63 | 0.32 | 0.05 | ACP |
| Mastoparan-C | 0.96 | 0.04 | 0.00 | ACP |
| NGR Peptide 1 | 0.65 | 0.35 | 0.00 | ACP |
| Pleurocidin 03 | 0.00 | 1.00 | 0.00 | AMP |
| R7-kla | 0.96 | 0.04 | 0.00 | ACP |
| RGD-4C-GG-(KLAKLAK) | 0.98 | 0.02 | 0.00 | ACP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burdukiewicz, M.; Sidorczuk, K.; Rafacz, D.; Pietluch, F.; Bąkała, M.; Słowik, J.; Gagat, P. CancerGram: An Effective Classifier for Differentiating Anticancer from Antimicrobial Peptides. Pharmaceutics 2020, 12, 1045. https://doi.org/10.3390/pharmaceutics12111045
Burdukiewicz M, Sidorczuk K, Rafacz D, Pietluch F, Bąkała M, Słowik J, Gagat P. CancerGram: An Effective Classifier for Differentiating Anticancer from Antimicrobial Peptides. Pharmaceutics. 2020; 12(11):1045. https://doi.org/10.3390/pharmaceutics12111045
Chicago/Turabian StyleBurdukiewicz, Michał, Katarzyna Sidorczuk, Dominik Rafacz, Filip Pietluch, Mateusz Bąkała, Jadwiga Słowik, and Przemysław Gagat. 2020. "CancerGram: An Effective Classifier for Differentiating Anticancer from Antimicrobial Peptides" Pharmaceutics 12, no. 11: 1045. https://doi.org/10.3390/pharmaceutics12111045