
Presence of Poly(A) Tails at the 3'-Termini of Some mRNAs of a Double-Stranded RNA Virus, Southern Rice Black-Streaked Dwarf Virus


Abstract
:1. Introduction
2. Materials and Methods
2.1 Virus and RNA Extraction
2.2 Rapid Amplification of cDNA End (RACE) PCR

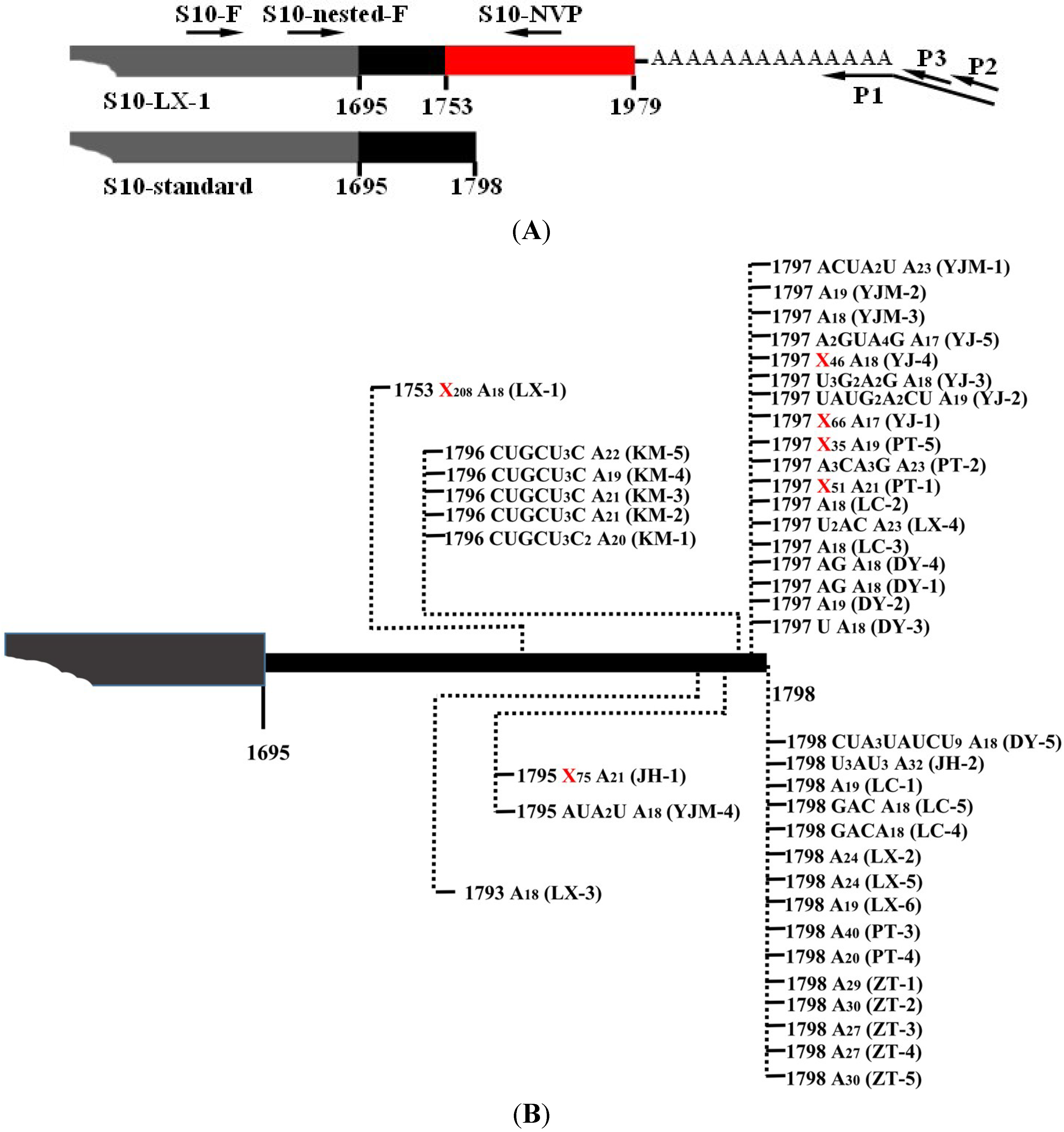{kind=link}
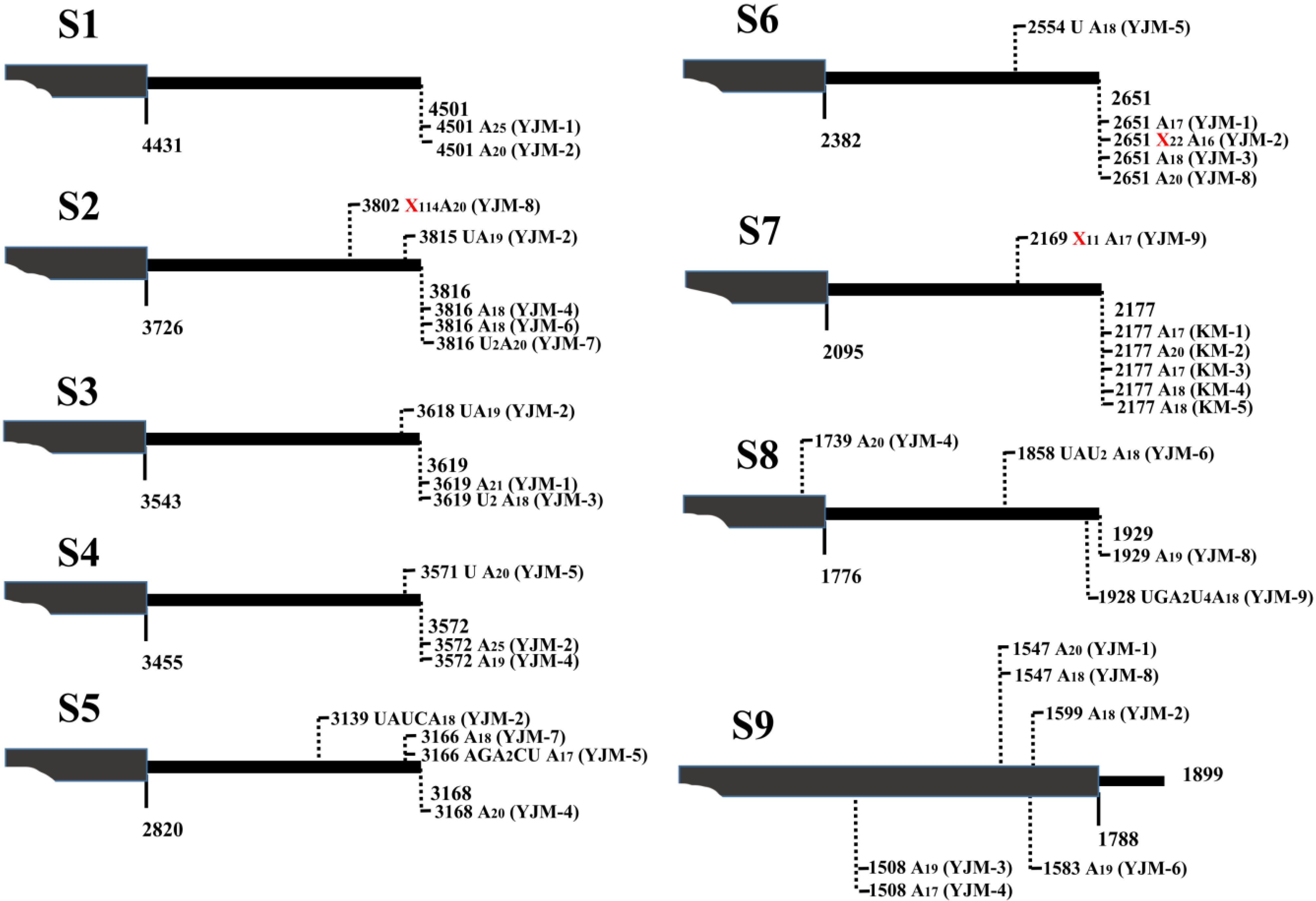{kind=link}
| Primer | Sequence (5'→3') | Target | Reference GenBank No. |
|---|---|---|---|
| S1-F | TCAGTGCTCAAGGCTCACAAGATTGAAG | S1-mRNA | JQ034348 |
| S1-nested-F | ATTCATGAACTTAATGGGCGCAGAGTG | ||
| S2-F | CGGCACATCTTCACCCGCAGACTTC | S2-mRNA | JQ034349 |
| S2-nested-F | CTGATGAATTGCTCGACCGTTACATTAG | ||
| S3-F | GATGGGATTAGCGAAATTGCATTTGGAG | S3-mRNA | JQ034350 |
| S3-nested-F | TGCATGGACATTCATTTTCAGATCAAG | ||
| S4-F | TAGATTTTGTTATTCCCGGTGTTCGAGAAG | S4-mRNA | JQ034351 |
| S4-nested-F | AGTGCGGATGTGGCTGCAGATAAATTC | ||
| S5-F | TGTGATCAGTGCCATGTCCACTAGCATC | S5-mRNA | JQ034352 |
| S5-nested-F | AATCATCCCTGTGCGCTTCGACTTAG | ||
| S6-F | CGATACTCTGATGAAACAGGCGAAGCTC | S6-mRNA | JQ034353 |
| S6-nested-F | TGAGAACCAATGGAGCGCGTATGGA | ||
| S7-F | ACTACTTCAGCTGAAGATGTCGACGCAC | S7-mRNA | JQ034354 |
| S7-nested-F | TTGGCAAGCGATGGAAAGAAGATGG | ||
| S8-F | CGTATTGGACGATGAGCGCAACTTTG | S8-mRNA | JQ034355 |
| S8-nested-F | TGAATTAGCGTTCGTACCTCATTCGCTG | ||
| S9-F | TTGGACTTGGCTAACTACGTTCGACAAC | S9-mRNA | JQ034356 |
| S9-nested-F | GGAATTGGATGATCGAGTTGAAAAATTGG | ||
| S10-F | CTCCCTGCATCGATTACATCAAACTTGG | S10-mRNA | JQ034357 |
| S10-nested-F | GCCAACAATTTATTGAAGGCGGATCG | ||
| S10-NVP | TTCCATCTCTATCATTCAGTCAAG | S10-mRNA | |
| Adapter-oligo(dT) (P1) | GCTGTCAACGATACGCTACGTAACGGCATGACAGTG(T)18VN | Poly(A) tails | |
| Adapter primer P2 | GCTGTCAACGATACGCTACGTAACG | Adapter | |
| Adapter primer P3 | CGCTACGTAACGGCATGACAGTG | Adapter |
3. Results and Discussion


Acknowledgments
Author Contributions
Conflicts of Interest
References
- King, A.M.Q.; Adams, M.J.; Carstens, E.B.; Lefkowitz, E.J. Virus Taxonomy: Classification and Nomenclature of Viruses, Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier: SanDiego, CA, USA, 2012. [Google Scholar]
- Sanfaçon, H.; Brodmann, P.; Hohn, T. A dissection of the cauliflower mosaic virus polyadenylation signal. Genes Dev. 1991, 5, 141–149. [Google Scholar] [CrossRef]
- Weichs an der Glon, C.; Ashe, M.; Eggermont, J.; Proudfoot, N.J. Tat-dependent occlusion of the HIVpoly(A) site. EMBO J. 1993, 12, 2119–2128. [Google Scholar]
- Poon, L.L.; Pritlove, D.C.; Fodor, E.; Brownlee, G.G. Direct evidence that the poly(A) tail of influenza A virus mRNA is synthesized by reiterative copying of a U track in the virion RNA template. J. Virol. 1999, 73, 3473–3476. [Google Scholar] [PubMed]
- Steil, B.P.; Kempf, B.J.; Barton, D.J. Poly(A) at the 3' end of positive-strand RNA and VPg-linked poly(U) at the 5' end of negative-strand RNA are reciprocal templates during replication of poliovirus RNA. J. Virol. 2010, 84, 2843–2858. [Google Scholar] [CrossRef] [PubMed]
- Dreher, T.W. Funcations of the 3'-untranslated regions of positive strand RNA viral genomes. Annu. Rev. Phytopathol. 1999, 37, 151–174. [Google Scholar] [CrossRef] [PubMed]
- Barr, J.N.; Fearns, R. How RNA viruses maintain their genome integrity. J. Gen. Virol. 2010, 91, 1373–1387. [Google Scholar] [CrossRef] [PubMed]
- Slomovic, S.; Fremder, E.; Staals, R.H.; Pruijn, G.J.; Schuster, G. Addition of poly(A) and poly(A)-rich tails during RNA degradation in the cytoplasm of human cells. Proc. Natl. Acad. Sci. USA 2010, 107, 7407–7412. [Google Scholar] [CrossRef] [PubMed]
- Li, W.M.; Zhang, Y.Q.; Zhang, C.; Pei, X.W.; Wang, Z.X.; Jia, S.R. Presence of poly(A) and poly(A)-rich tails in a positive-strand RNA virus known to lack 3' poly(A) tails. Virology 2014, 454–455, 1–10. [Google Scholar]
- Zhou, G.H.; Wen, J.J.; Cai, D.J.; Li, P.; Xu, D.L.; Zhang, S.G. Southern rice black-streaked dwarf virus: A new proposed Fijivirus species in the family Reoviridae. Chin. Sci. Bull. 2008, 53, 3677–3685. [Google Scholar] [CrossRef]
- Cheng, Z.B.; Li, S.; Gao, R.Z.; Sun, F.; Liu, W.C.; Zhou, G.H.; Wu, J.X.; Zhou, X.P.; Zhou, Y.J. Distribution and genetic diversity of Southern rice black-streaked dwarf virus in China. Virol. J. 2013, 10, 307. [Google Scholar]
- Hoang, A.T.; Zhang, H.M.; Yang, J.; Chen, J.P.; Hébrard, E.; Zhou, G.H.; Vinh, V.N.; Cheng, J.A. Identification, characterization, and distribution of Southern rice black-streaked dwarf virus in Vietnam. Plant Dis. 2011, 95, 1063–1069. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, J.; Zhou, G.H.; Zhang, H.M.; Chen, J.P.; Adams, M.J. The complete genome sequence of two isolates of Southern rice black-streaked dwarf virus, a new member of the genus Fijivirus. J. Phytopathol. 2010, 158, 733–737. [Google Scholar] [CrossRef]
- Lu, Y.H.; Zhang, J.F.; Xiong, R.Y.; Xu, Q.F.; Zhou, Y.J. Identification of an RNA silencing suppressor encoded by Southern rice black-streaked dwarf virus S6. Sci. Agri. Sin. 2011, 44, 2909–2917. [Google Scholar]
- Jia, D.S.; Mao, Q.Z.; Chen, H.Y.; Wang, A.M.; Liu, Y.Y.; Wang, H.T.; Xie, L.H.; Wei, T.Y. Virus-induced tubule: A vehicle for rapid spread of virions through basal lamina from midgut epithelium in the insect vector. J. Virol. 2014, 88, 10488–10500. [Google Scholar] [CrossRef] [PubMed]
- Jia, D.S.; Chen, H.Y.; Zheng, A.L.; Chen, Q.; Liu, Q.F.; Xie, L.H.; Wu, Z.J.; Wei, T.Y. Development of an insect vector cell culture and RNA interference system to investigate the functional role of fijivirus replication protein. J. Virol. 2012, 86, 5800–5807. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.M.; Chen, J.P.; Adams, M.J. Molecular characterisation of segments 1 to 6 of Rice black-streaked dwarf virus from China provides the complete genome. Arch. Virol. 2001, 146, 2331–2339. [Google Scholar] [CrossRef] [PubMed]
- Isogai, M.; Uyeda, I.; Lee, B.C. Detection and assignment of proteins encoded by Rice black streaked dwarf fijivirus S7, S8, S9 and S10. J. Gen. Virol. 1998, 79, 1487–1494. [Google Scholar] [PubMed]
- He, P.; Liu, J.J.; He, M.; Wang, Z.C.; Chen, Z.; Guo, R.; Correll, J.C.; Yang, S.; Song, B.A. Quantitative detection of relative expression levels of the whole genome of Southern rice black-streaked dwarf virus and its replication in different hosts. Virol. J. 2013, 10, 136. [Google Scholar] [CrossRef]
- Ji, Y.H.; Gao, R.Z.; Zhang, Y.; Cheng, Z.B.; Zhou, T.; Fan, Y.J.; Zhou, Y.J. A simplified method for quick detection of Rice black-streaked dwarf virus and Southern rice black-streaked dwarf virus. Chin. J. Rice Sci. 2011, 25, 91–94. [Google Scholar]
- Slomovic, S.; Portnoy, V.; Liveanu, V.; Schuster, G. RNA polyadenylation in prokaryotes and organelles; different tails tell different tales. Crit. Rev. Plant Sci. 2006, 25, 65–77. [Google Scholar] [CrossRef]
- Regnier, P.; Hajnsdorf, E. Poly(A)-assisted RNA decay and modulators of RNA stability. Prog. Mol. Biol. Transl. Sci. 2009, 85, 137–185. [Google Scholar] [PubMed]
- Kuai, L.; Das, B.; Sherman, F. A nuclear degradation pathway controls the abundance of normal mRNAs in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2005, 102, 13962–13967. [Google Scholar] [CrossRef]
- Wyers, F.; Rougemaille, M.; Badis, G.; Rousselle, J.C.; Dufour, M.E.; Boulay, J.; Régnault, B.; Devaux, F.; Namane, A.; Séraphin, B.; et al. Cryptic pol II transcripts are degraded by a nuclear quality control pathway involving a new poly(A) polymerase. Cell 2005, 121, 725–737. [Google Scholar]
- Nakamura, R.; Takeuchi, R.; Takata, K.; Shimanouchi, K.; Abe, Y.; Kanai, Y.; Ruike, T.; Ihara, A.; Sakaguchi, K. TRF4 is involved in polyadenylation of snRNAs in Drosophila melanogaster. Mol. Cell Biol. 2008, 28, 6620–6631. [Google Scholar] [CrossRef]
- Lange, H.; Sement, F.M.; Canaday, J.; Gagliardi, D. Polyadenylation-assisted RNA degradation processes in plants. Trends Plant Sci. 2009, 14, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Slomovic, S.; Portnoy, V.; Yehudai-Resheff, S.; Bronshtein, E.; Schuster, G. Polynucleotide phosphorylase and the archaeal exosome as poly(A)-polymerases. Biochim. Biophys. Acta 2008, 1779, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Houseley, J.; Tollervey, D. The many pathways of RNA degradation. Cell 2009, 136, 763–776. [Google Scholar] [CrossRef] [PubMed]
- Lange, H.; Gagliardi, D. The exosome and 3'-5' RNA degradation in plants. Adv. Exp. Med. Biol. 2011, 702, 50–62. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, M.; Jiang, Z.; Li, S.; He, P. Presence of Poly(A) Tails at the 3'-Termini of Some mRNAs of a Double-Stranded RNA Virus, Southern Rice Black-Streaked Dwarf Virus. Viruses 2015, 7, 1642-1650. https://doi.org/10.3390/v7041642
He M, Jiang Z, Li S, He P. Presence of Poly(A) Tails at the 3'-Termini of Some mRNAs of a Double-Stranded RNA Virus, Southern Rice Black-Streaked Dwarf Virus. Viruses. 2015; 7(4):1642-1650. https://doi.org/10.3390/v7041642
Chicago/Turabian StyleHe, Ming, Ziqiong Jiang, Shuo Li, and Peng He. 2015. "Presence of Poly(A) Tails at the 3'-Termini of Some mRNAs of a Double-Stranded RNA Virus, Southern Rice Black-Streaked Dwarf Virus" Viruses 7, no. 4: 1642-1650. https://doi.org/10.3390/v7041642